
Extracted Spectral Signatures from the Water Column as a Tool for the Prediction of the Structure of a Marine Microbial Community


 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Incubations
2.2. Fluorometric Determination of Chlorophyll and Radiometric Determination of Chlorophyll Absorbance Spectra
2.3. Flow Cytometry
2.4. Light Microscopy
2.5. Solar Radiation, Salinity, Temperature, and Depth Measurements
2.6. Preprocessing of Hyperspectral Curves Using Reference Measurements
2.7. Computational Rquirements
3. Results
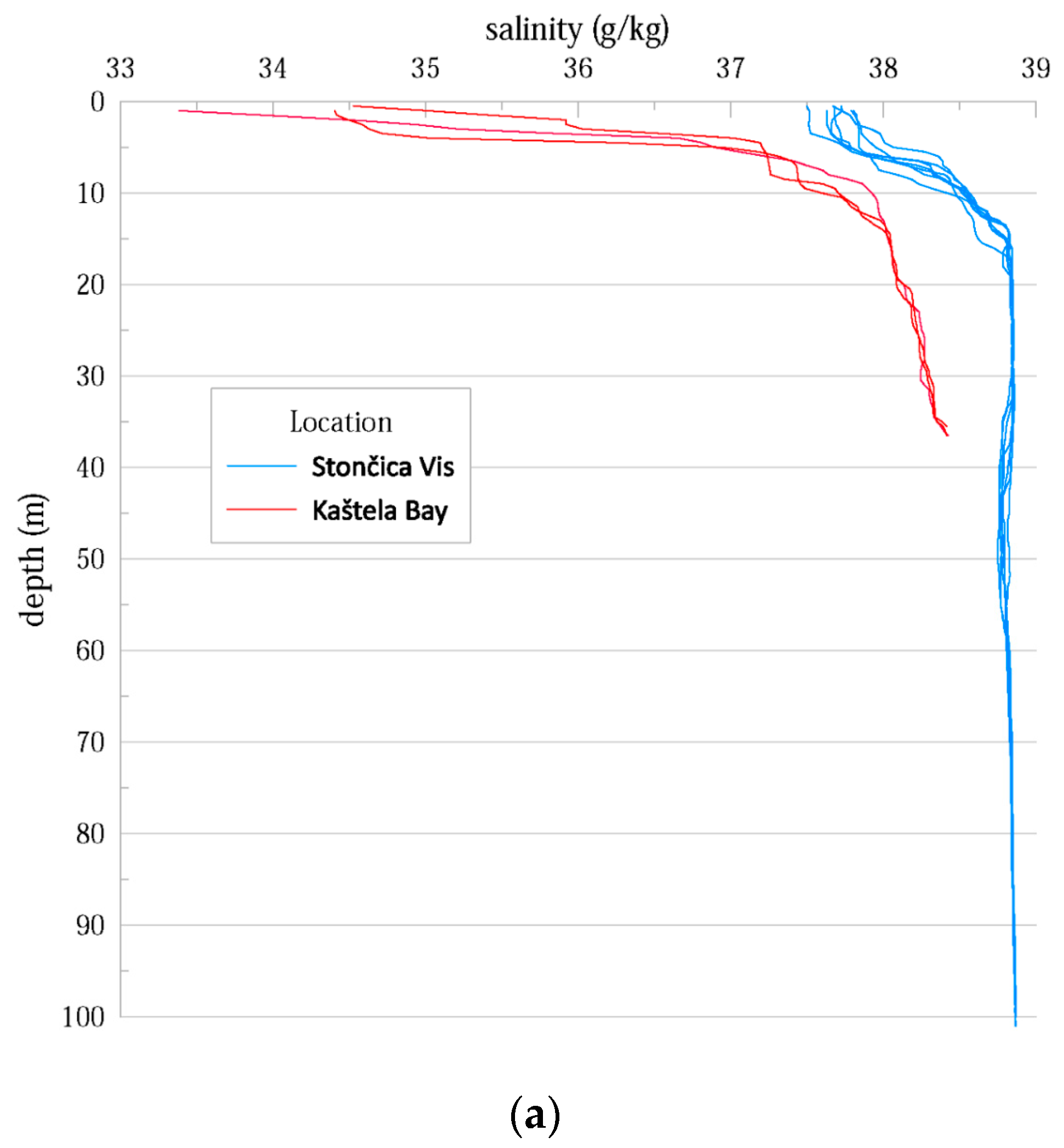
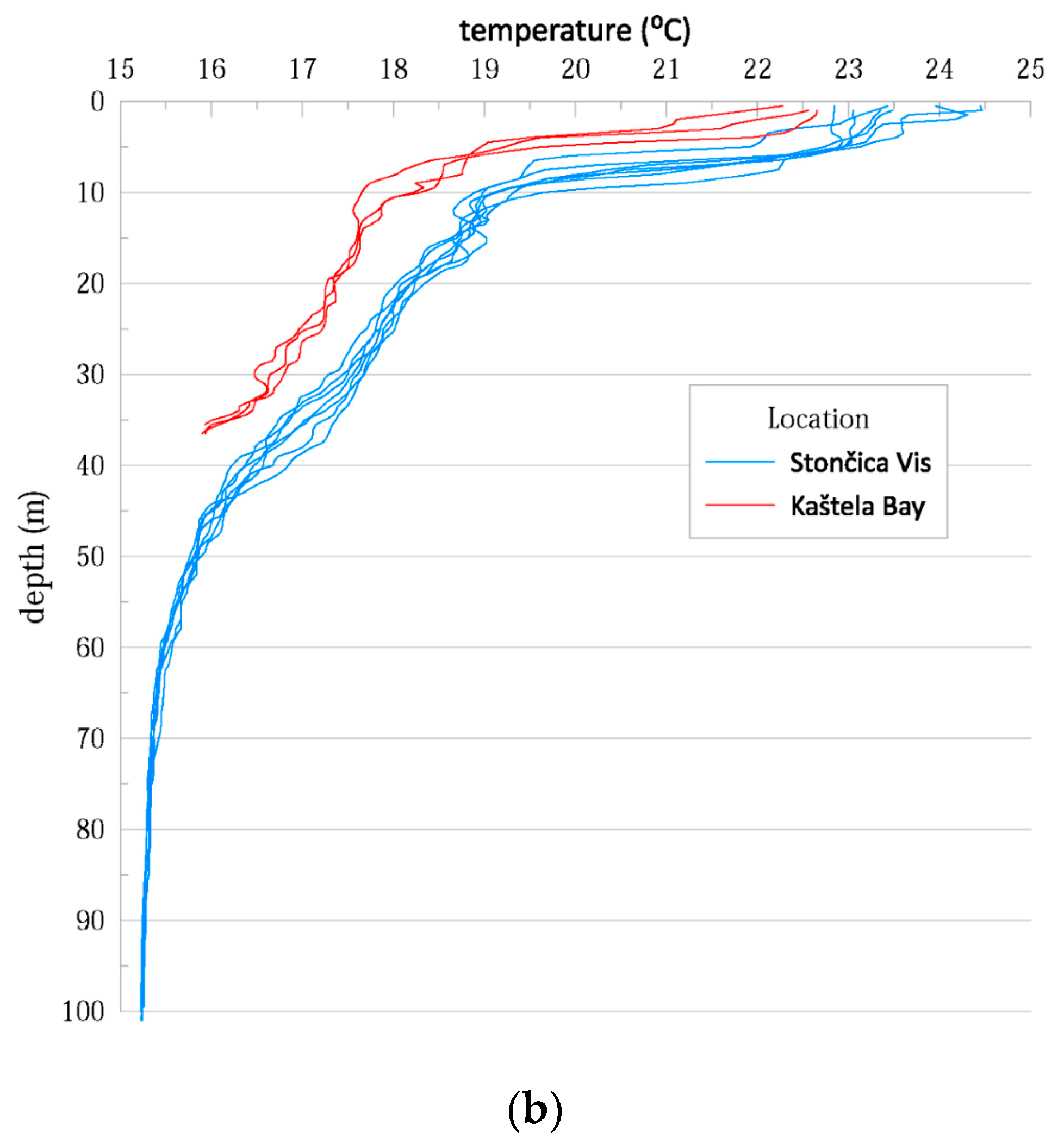
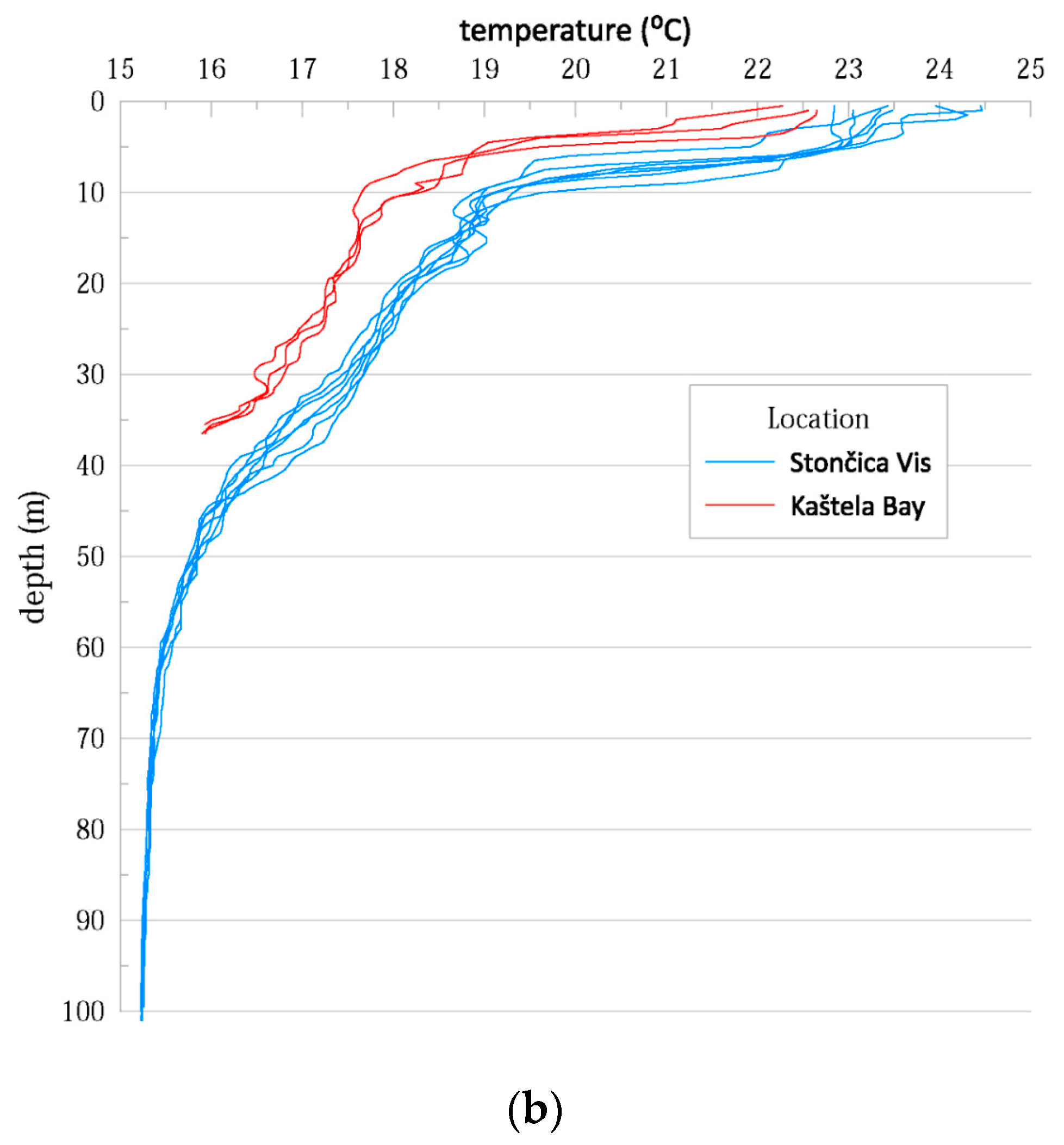
3.1. Hydrographic Properties of the Water Column
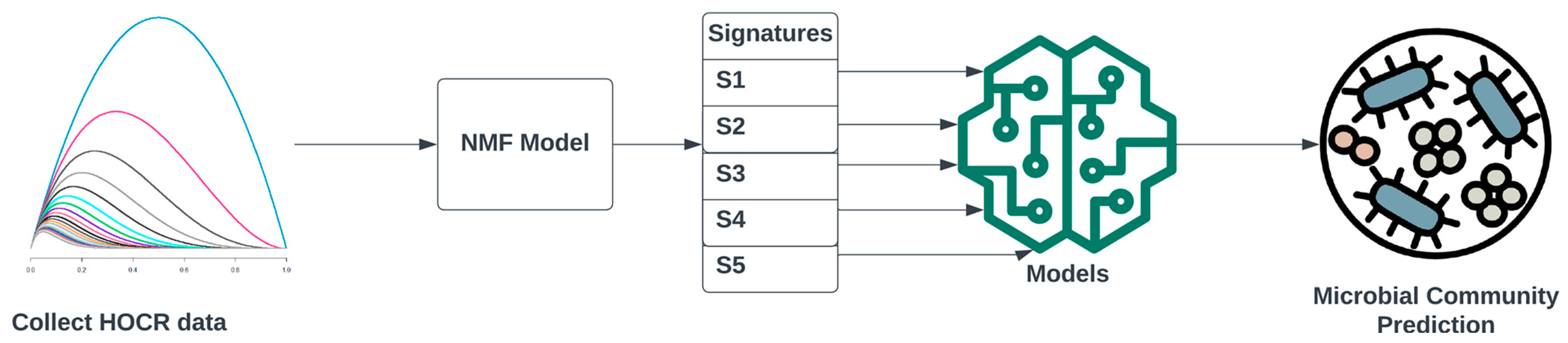
3.2. Developing HOCR Signatures Using a Non-Negative Factorization Model on Training Data
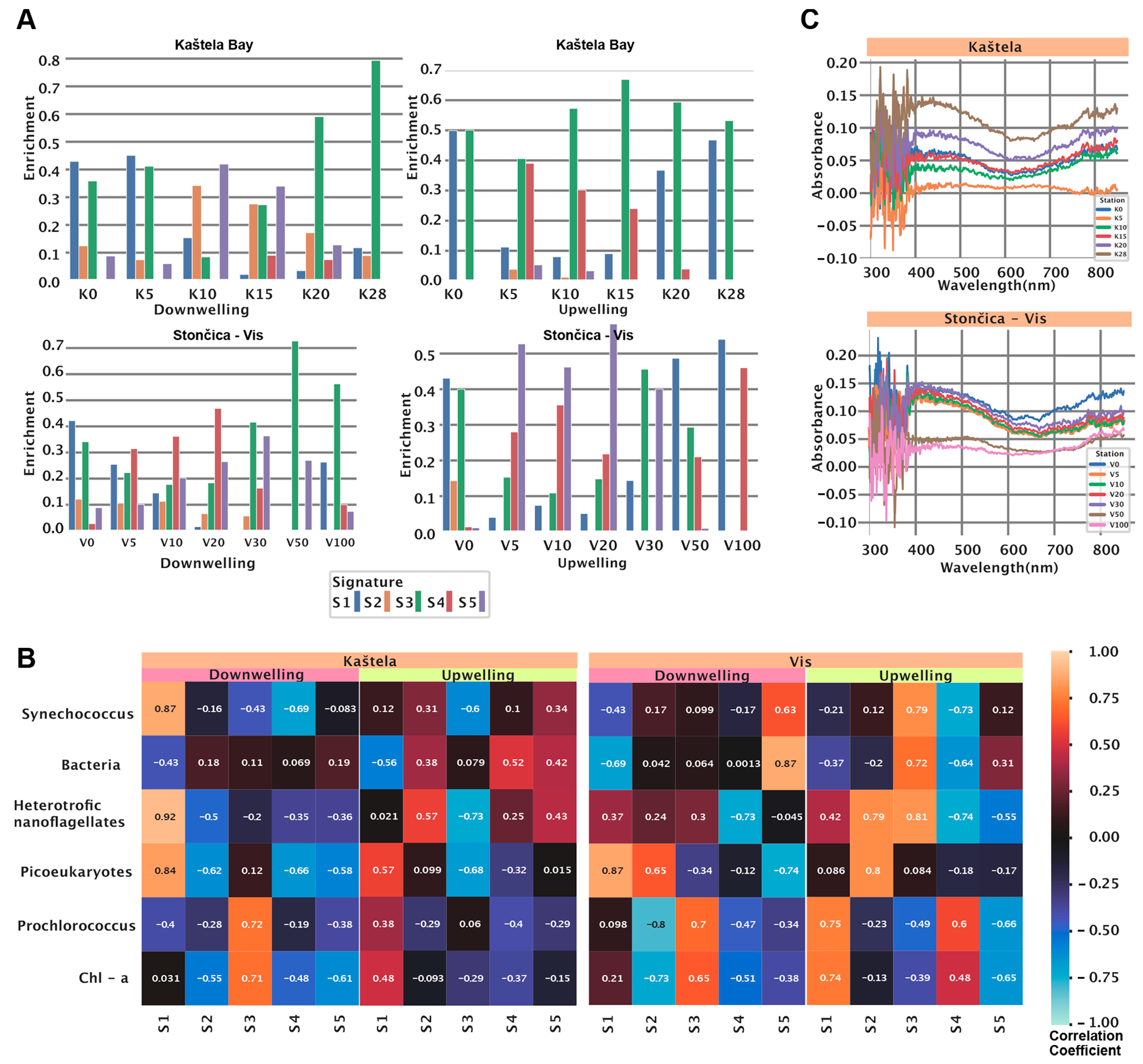
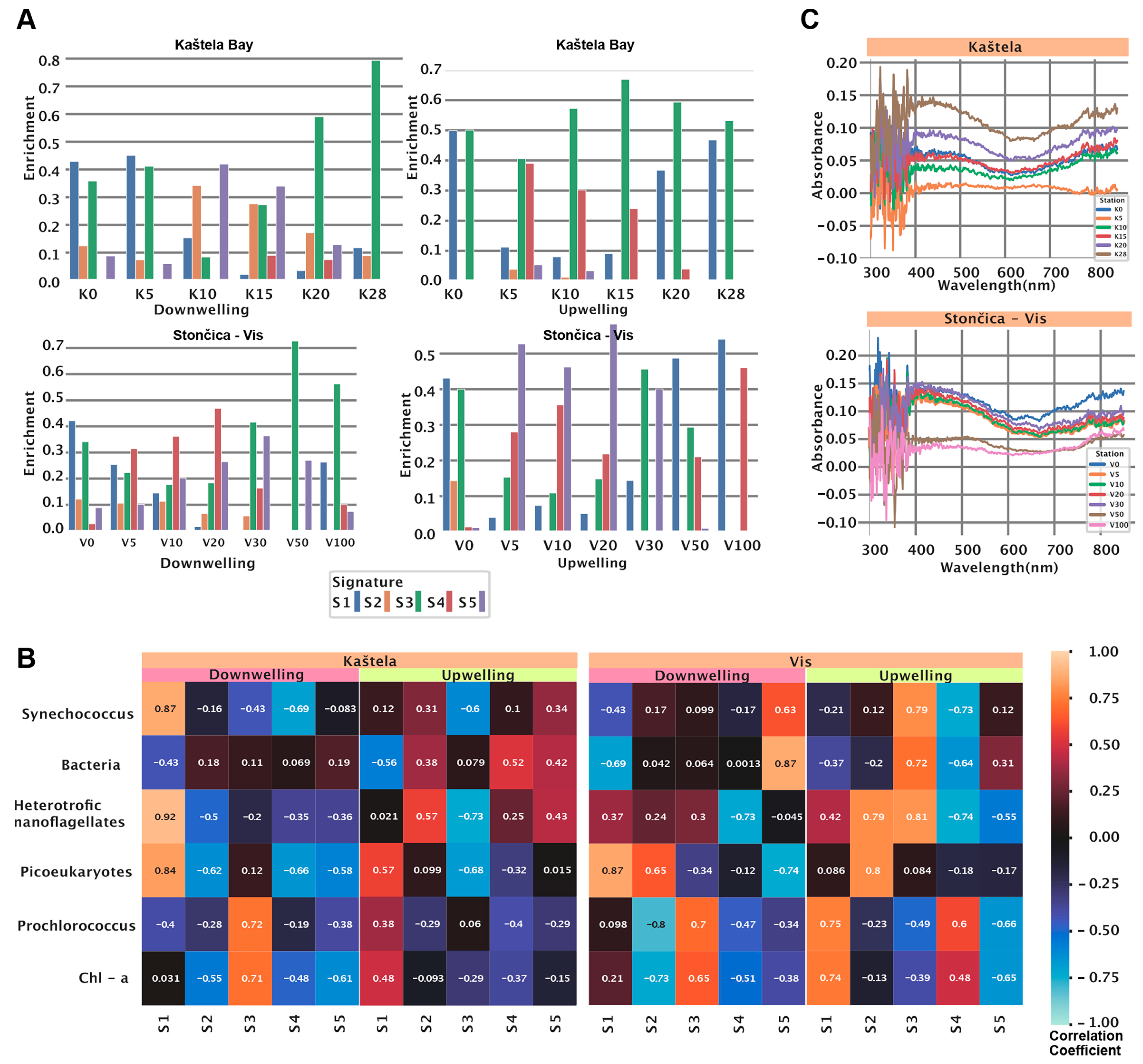
3.3. HOCR Spectral Signatures at the Study Locations
3.4. Microbial Community Structure at the Two Stations
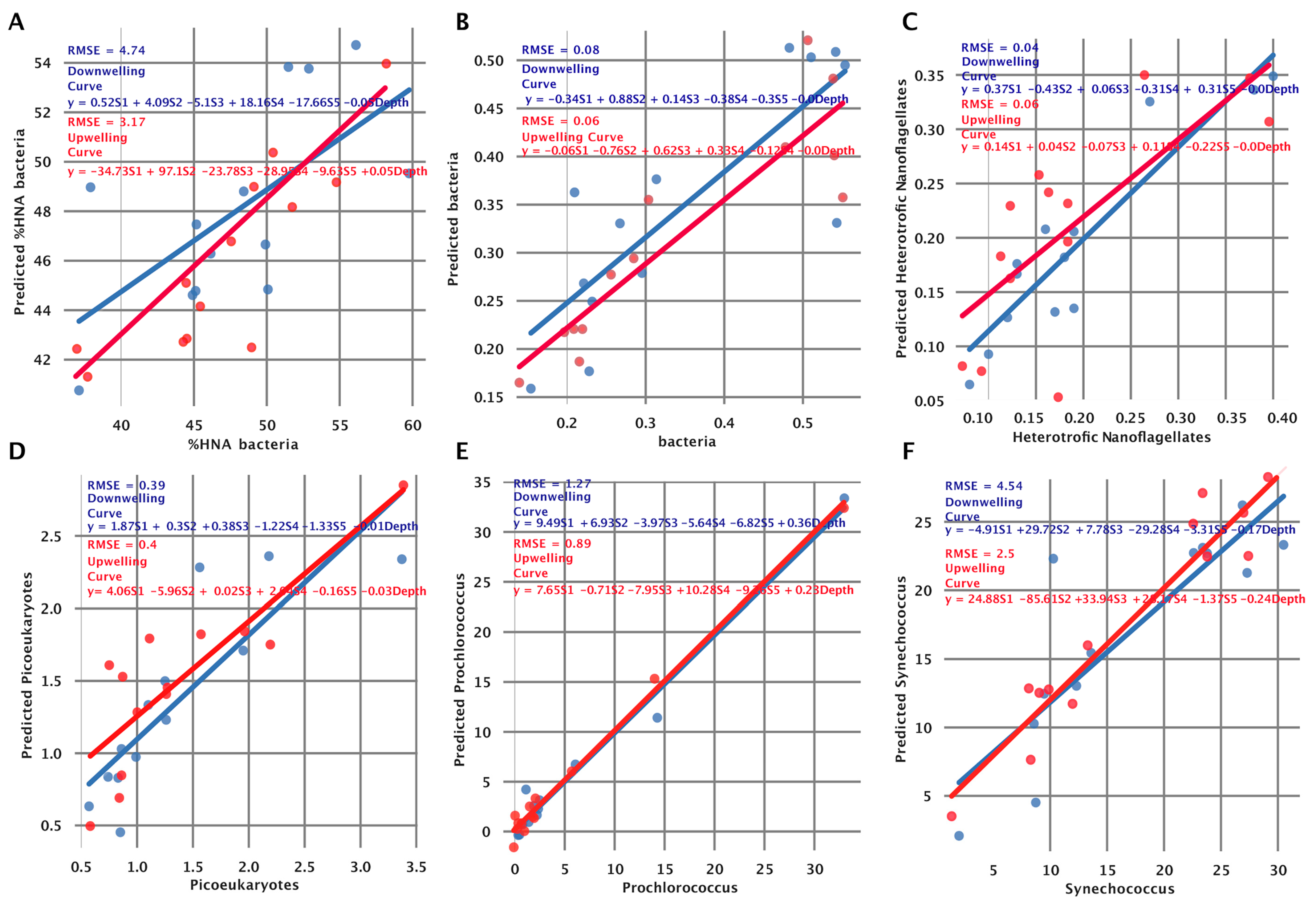
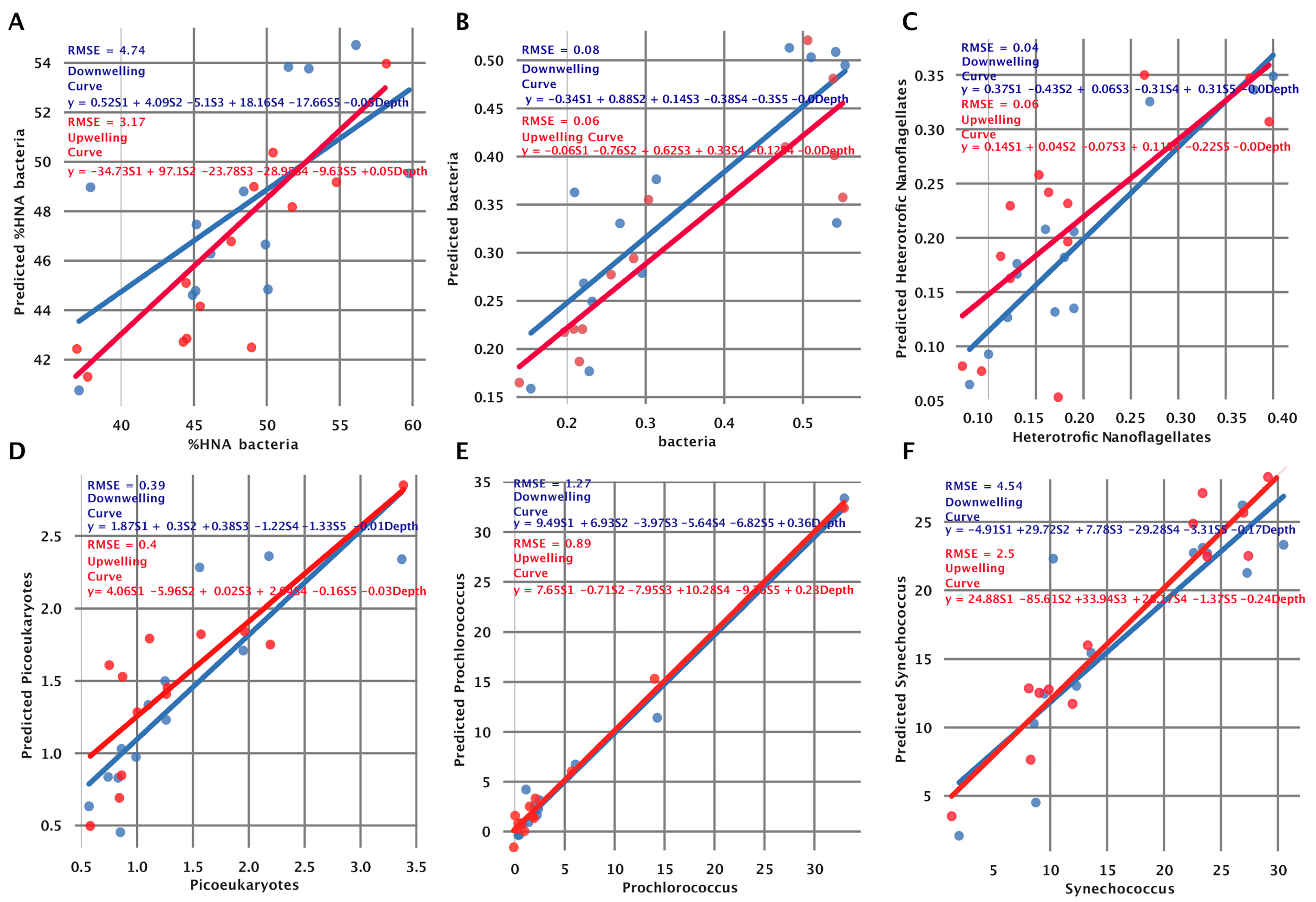
3.5. Fitting the Model to Independent Data Collected at the Two Stations
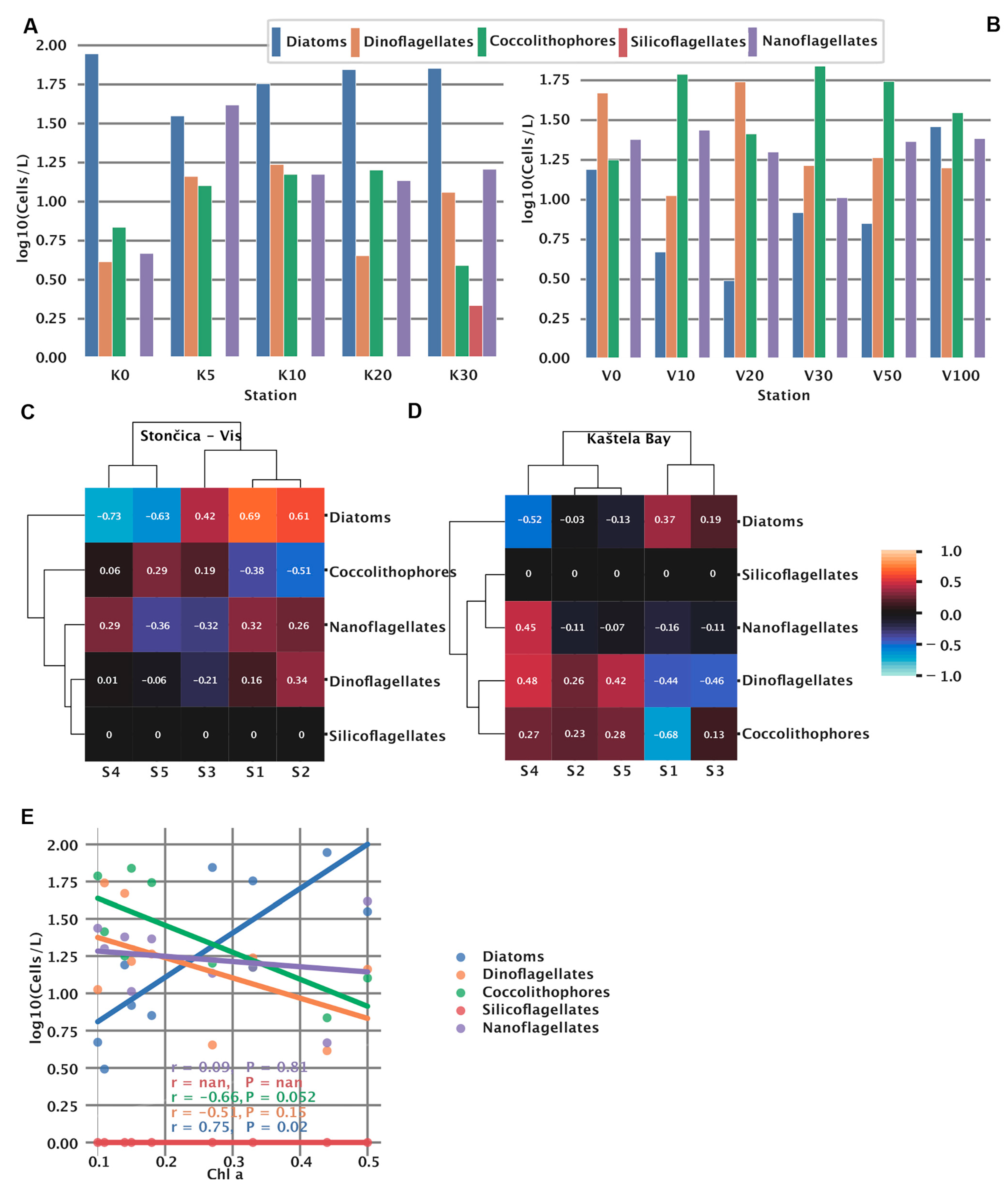
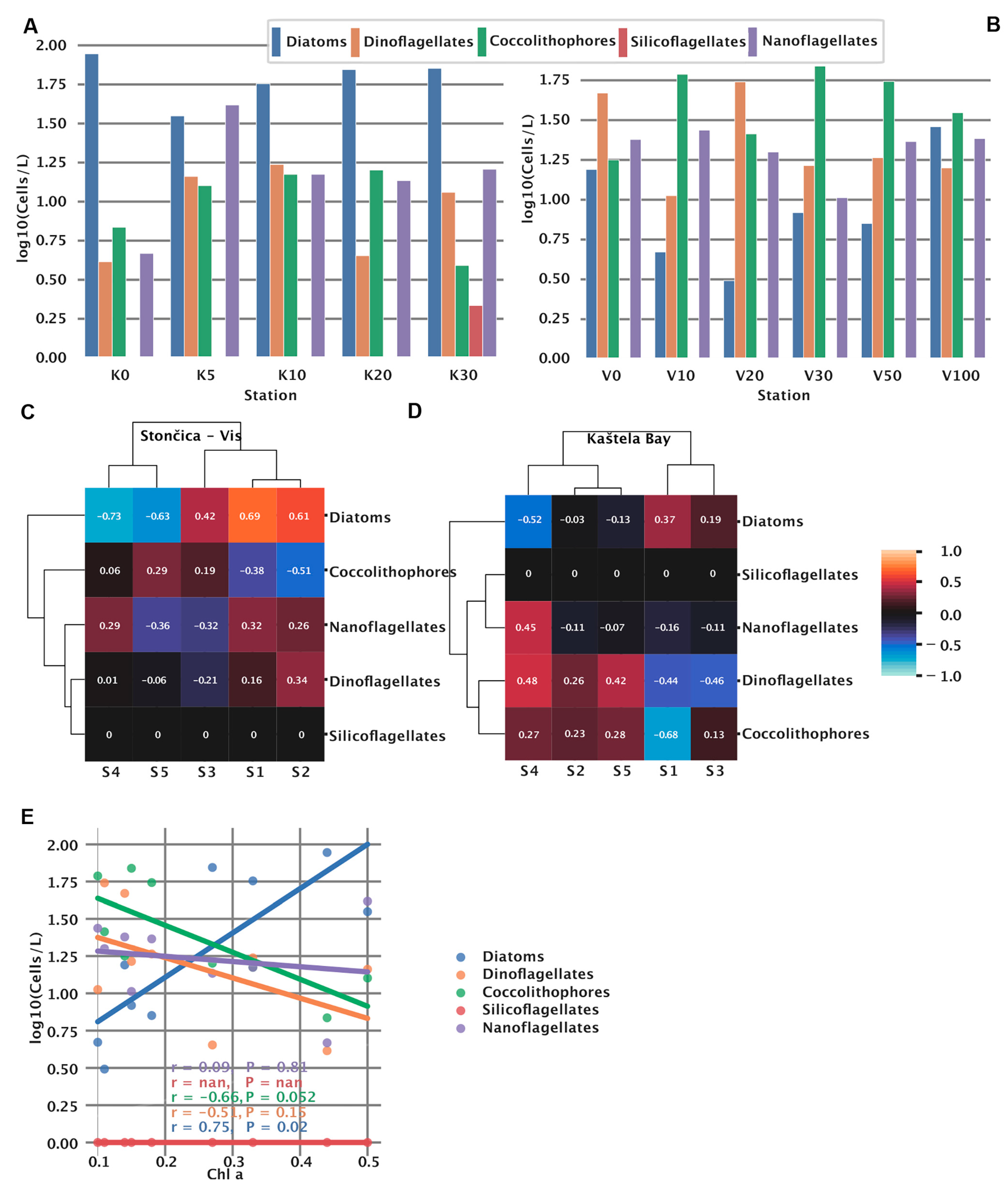
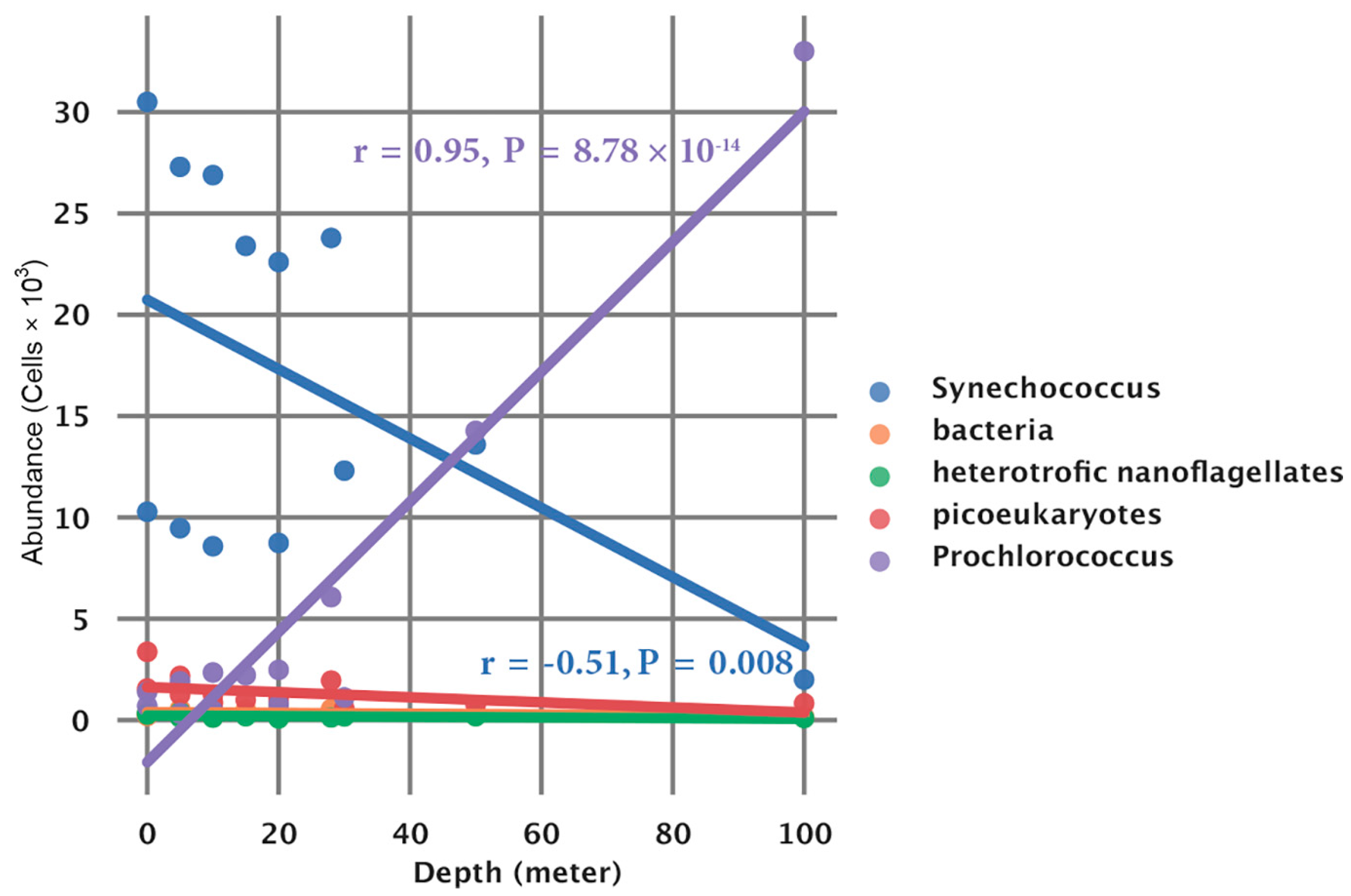
3.6. Phytoplankton Abundance and Community Structure
3.7. Spectral Signatures and Microbial Community Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Bukata, R.P.; Jerome, J.H.; Kondratyev, A.S.; Pozdnyakov, D.V. Optical Properties and Remote Sensing of Inland and Coastal Waters, 1st ed.; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Feng, H.; Campbell, J.W.; Dowell, M.D.; Moore, T.S. Modeling spectral reflectance of optically complex waters using bio-optical measurements from Tokyo Bay. Remote Sens. Environ. 2005, 99, 232–243. [Google Scholar] [CrossRef]
- IOCCG. Remote Sensing of Inherent Optical Properties: Fundamentals, Tests of Algorithms, and Applications; Lee, Z.-P., Ed.; Reports of the International Ocean-Colour Coordinating Group; International Ocean-Colour Coordinating Group (IOCCG): Dartmouth, NS, Canada, 2006; pp. 1–122. [Google Scholar]
- Strömbeck, N.; Pierson, D.C. The effects of variability in the inherent optical properties on estimations of chlorophyll a by remote sensing in Swedish freshwaters. Sci. Total Environ. 2001, 268, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.J.; Ciotti, Á.M.; Davis, R.F.; Lewis, M.R. Optical detection and assessment of algal blooms. Limnol. Oceanogr. 1997, 42, 1223–1239. [Google Scholar] [CrossRef]
- Schofield, O.; Grzymski, J.; Bissett, W.P.; Kirkpatrick, G.J.; Millie, D.F.; Moline, M.; Roesler, C.S. Optical monitoring and forecasting systems for harmful algal blooms: Possibility or pipe dream? J. Phycol. 1999, 35, 1477–1496. [Google Scholar] [CrossRef]
- IOCCG. Phytoplankton Functional Types from Space; Sathyendranath, S., Ed.; Reports of the International Ocean-Colour Coordinating Group; International Ocean-Colour Coordinating Group (IOCCG): Dartmouth, NS, Canada, 2014; pp. 1–154. [Google Scholar]
- Mouw, C.B.; Hardman-Mountford, N.J.; Alvain, S.; Bracher, A.; Brewin, R.J.W.; Bricaud, A.; Ciotti, A.M.; Devred, E.; Fujiwara, A.; Hirata, T.; et al. A Consumer’s Guide to Satellite Remote Sensing of Multiple Phytoplankton Groups in the Global Ocean. Front. Mar. Sci. 2017, 4, 41. [Google Scholar] [CrossRef]
- Ciotti, Á.M.; Lewis, M.R.; Cullen, J.J. Assessment of the relationships between dominant cell size in natural phytoplankton communities and the spectral shape of the absorption coefficient. Limnol. Oceanogr. 2002, 47, 404–417. [Google Scholar] [CrossRef]
- Sathyendranath, S.; Lazzara, L.; Prieur, L. Variations in the spectral values of specific absorption of phytoplankton. Limnol. Oceanogr. 1987, 32, 403–415. [Google Scholar] [CrossRef]
- Bricaud, A.; Claustre, H.; Ras, J.; Oubelkheir, K. Natural variability of phytoplanktonic absorption in oceanic waters: Influence of the size structure of algal populations. J. Geophys. Res. Ocean. 2004, 109, C11010. [Google Scholar] [CrossRef]
- Jemai, A.; Wollschläger, J.; Voβ, D.; Zielinski, O. Radiometry on Argo Floats: From the Multispectral State-of-the-Art on the Step to Hyperspectral Technology. Front. Mar. Sci. 2021, 8, 676537. [Google Scholar] [CrossRef]
- Wang, Y.-X.; Zhang, Y.-J. Nonnegative Matrix Factorization: A Comprehensive Review. IEEE Trans. Knowl. Data Eng. 2013, 25, 1336–1353. [Google Scholar] [CrossRef]
- Pauca, V.P.; Piper, J.; Plemmons, R.J. Nonnegative matrix factorization for spectral data analysis. Linear. Algebra Appl. 2006, 416, 29–47. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Campbell, P.J.; Stratton, M.R. Deciphering signatures of mutational processes operative in human cancer. Cell Rep. 2013, 3, 246–259. [Google Scholar] [CrossRef]
- Marinov, I.; Doney, S.C.; Lima, I.D. Response of ocean phytoplankton community structure to climate change over the 21st century: Partitioning the effects of nutrients, temperature and light. Biogeosciences 2010, 7, 3941–3959. [Google Scholar] [CrossRef]
- Puškarić, S.; Sokač, M.; Matić, K. Application of non-negative matrix factorization for studying short-term physiological changes in grapevine from canopy hyperspectral reflection. RIThink 2021, 10, 1–25. [Google Scholar]
- Holm-Hansen, O.; Lorenzen, C.J.; Holmes, R.W.; Strickland, J.D.H. Fluorometric Determination of Chlorophyll. ICES J. Mar. Sci. 1965, 30, 3–15. [Google Scholar] [CrossRef]
- Gasol, J.M.; Morán, X.A.G. Flow Cytometric Determination of Microbial Abundances and Its Use to Obtain Indices of Community Structure and Relative Activity. In Hydrocarbon and Lipid Microbiology Protocols; Springer: Berlin/Heidelberg, Germany, 2015; pp. 159–187. [Google Scholar]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Marasović, I.; Gačić, M.; Kovačević, V.; Krstulović, N.; Kušpilić, G.; Pucher-Petković, T.; Odzak, N.; Solic, M. Development of the red tide in the Kaštela Bay (Adriatic Sea). Mar. Chem. 1991, 32, 375–387. [Google Scholar] [CrossRef]
- Lee, D.D.; Seung, H.S. Learning the parts of objects by non-negative matrix factorization. Nature 1999, 401, 788–791. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, P.; Tismenetsky, M. The Theory of Matrices: With Applications; Elsevier: Gainesville, FL, USA, 1985. [Google Scholar]
- Paxinos, R. A rapid Utermohl method for estimating algal numbers. J. Plankton. Res. 2000, 22, 2255–2262. [Google Scholar] [CrossRef]
- Vilibić, I.; Šantić, D. Deep water ventilation traced by Synechococcus cyanobacteria. Ocean Dyn. 2008, 58, 119–125. [Google Scholar] [CrossRef]
- Šantić, D.; Kovačević, V.; Bensi, M.; Giani, M.; Vrdoljak Tomaš, A.; Ordulj, M.; Santinelli, C.; Šestanović, S.; Šolić, M.; Grbec, B. Picoplankton Distribution and Activity in the Deep Waters of the Southern Adriatic Sea. Water 2019, 11, 1655. [Google Scholar] [CrossRef]
- Mella-Flores, D.; Mazard, S.; Humily, F.; Partensky, F.; Mahé, F.; Bariat, L.; Courties, C.; Marie, D.; Ras, J.; Mauriac, R.; et al. Is the distribution of Prochlorococcus and Synechococcus ecotypes in the Mediterranean Sea affected by global warming? Biogeosciences 2011, 8, 2785–2804. [Google Scholar] [CrossRef]
- Aktan, Y. Large-scale patterns in summer surface water phytoplankton (except picophytoplankton) in the Eastern Mediterranean. Estuar. Coast Shelf. Sci. 2011, 91, 551–558. [Google Scholar] [CrossRef]
- Navarro, G.; Alvain, S.; Vantrepotte, V.; Huertas, I.E. Identification of dominant phytoplankton functional types in the Mediterranean Sea based on a regionalized remote sensing approach. Remote Sens. Environ. 2014, 152, 557–575. [Google Scholar] [CrossRef]
- Brotas, V.; Tarran, G.A.; Veloso, V.; Brewin, R.J.W.; Woodward, E.M.S.; Airs, R.; Beltran, C.; Ferreira, A.; Groom, S.B. Complementary approaches to assess phytoplankton groups and size classes on a long transect in the Atlantic Ocean. Front. Mar. Sci. 2022, 8, 682621. [Google Scholar] [CrossRef]
- Šolić, M.; Šantić, D.; Šestanović, S.; Bojanić, N.; Grbec, B.; Jozić, S.; Vrdoljak, A.; Ordulj, M.; Matić, F.; Kušpilić, G.; et al. Impact of water column stability dynamics on the succession of plankton food web types in the offshore area of the Adriatic Sea. J. Sea Res. 2020, 158, 101860. [Google Scholar] [CrossRef]
- Šantić, D.; Krstulović, N.; Šolić, M.; Ordulj, M.; Kušpilić, G. Dynamics of prokaryotic picoplankton community in the central and southern Adriatic Sea (Croatia). Helgol. Mar. Res. 2012, 67, 471–481. [Google Scholar] [CrossRef]
- Vilibić, I.; Šepić, J.; Proust, N. Weakening thermohaline circulation in the Adriatic Sea. Clim. Res. 2013, 55, 217–225. [Google Scholar] [CrossRef]
- Organelli, E.; Claustre, H. Small Phytoplankton Shapes Colored Dissolved Organic Matter Dynamics in the North Atlantic Subtropical Gyre. Geophys. Res. Lett. 2019, 46, 12183–12191. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Signature | Spectrum Assoc. | Depth Assoc. (Training Data) | Microbial Assoc. Kaštela | Microbial Assoc. Stončica Vis |
|---|---|---|---|---|
| S1 | Two peaks at 359 nm and 597 nm | UW: Positive | UW: PE (positive), PCO (positive), BAC (negative) | UW: SNCO (negative), HNAN (positive), BAC (negative), PCO (positive) |
| DW: Minimal enrichment, around 40 m | DW: SNCO (positive), HNAN (positive), PE (positive), BAC (negative), PCO (negative) | DW: SNCO (negative),HNAN (positive), PE (positive), BAC (negative) | ||
| S2 | High-intensity broad peak at 506 nm | UW: Minimal enrichment | UW: HNAN (positive), BAC (positive) | UW: HNAN (positive) PE (positive) |
| DW: Negative convex | DW: PCO (negative), PE (negative), HNAN (negative), | DW: PE(positive), PCO (negative) | ||
| S3 | Small peak at 490 nm | UW: Enriched at all depths (0–100 m), mostly around 90 m | UW: HNAN (negative), PE (negative), SNCO (negative) | UW: SNCO (positive), BAC (positive), HNAN (positive), PCO (negative) |
| DW: Positive, mainly between 0–40 m | DW: SNCO (negative), PCO (positive) | DW: PCO (positive), PE (negative) | ||
| S4 | Low-intensity, almost uniform at 350–580 nm | UW: Positive, mostly around 80 m | UW: BAC (positive) | UW: SNCO (negative), HNAN (negative),BAC (negative), PCO (positive) |
| DW: Highly enriched in 0–20 m, minimal in depth > 20 m | DW: SNCO (negative), HNAN (negative), PE (negative) | DW: HNAN (negative), PCO (negative) | ||
| S5 | Low-intensity, Covering a broad spectrum of 350–560 nm | UW: Negative, mostly around 80 m | UW: SNCO (positive), HNAN (positive), BAC (positive), | UW: HNAN (negative), PCO (negative) |
| DW: Enriched at all depths (0–100 m) | DW: HNAN (negative), PE (negative), PCO (negative) | DW: PE (negative), PCO (negative), BAC (positive), SNCO (positive) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puškarić, S.; Sokač, M.; Ninčević, Ž.; Šantić, D.; Skejić, S.; Džoić, T.; Prelesnik, H.; Børsheim, K.Y. Extracted Spectral Signatures from the Water Column as a Tool for the Prediction of the Structure of a Marine Microbial Community. J. Mar. Sci. Eng. 2024, 12, 286. https://doi.org/10.3390/jmse12020286
Puškarić S, Sokač M, Ninčević Ž, Šantić D, Skejić S, Džoić T, Prelesnik H, Børsheim KY. Extracted Spectral Signatures from the Water Column as a Tool for the Prediction of the Structure of a Marine Microbial Community. Journal of Marine Science and Engineering. 2024; 12(2):286. https://doi.org/10.3390/jmse12020286
Chicago/Turabian StylePuškarić, Staša, Mateo Sokač, Živana Ninčević, Danijela Šantić, Sanda Skejić, Tomislav Džoić, Heliodor Prelesnik, and Knut Yngve Børsheim. 2024. "Extracted Spectral Signatures from the Water Column as a Tool for the Prediction of the Structure of a Marine Microbial Community" Journal of Marine Science and Engineering 12, no. 2: 286. https://doi.org/10.3390/jmse12020286
APA StylePuškarić, S., Sokač, M., Ninčević, Ž., Šantić, D., Skejić, S., Džoić, T., Prelesnik, H., & Børsheim, K. Y. (2024). Extracted Spectral Signatures from the Water Column as a Tool for the Prediction of the Structure of a Marine Microbial Community. Journal of Marine Science and Engineering, 12(2), 286. https://doi.org/10.3390/jmse12020286