
A Report on the Artificial Reef Use in Grenada, West Indies
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Coral
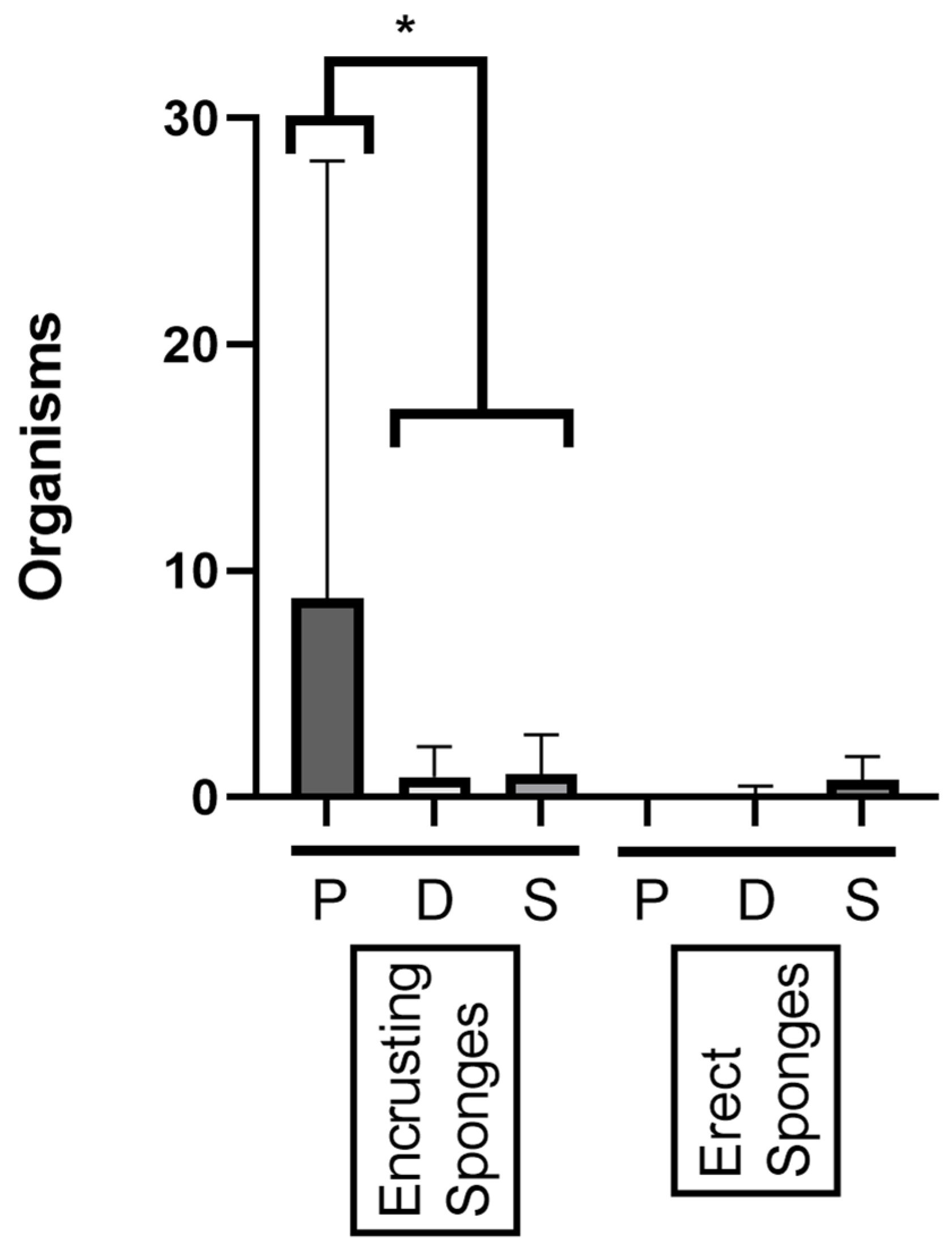
3.2. Sponges
3.3. Algae and Cyanobacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheaves, M.; Baker, R.; Nagelkerken, I.; Connolly, R. True Value of Estuarine and Coastal Nurseries for Fish: Incorporating Complexity and Dynamics. Estuaries Coasts 2015, 38, 401–414. [Google Scholar] [CrossRef]
- Reguero, B.G.; Beck, M.W.; Agostini, V.N.; Kramer, P.; Hancock, B. Coral Reefs for Coastal Protection: A New Methodological Approach and Engineering Case Study in Grenada. J. Environ. Manag. 2018, 210, 146–161. [Google Scholar] [CrossRef]
- Weaver, D.B. Ecotourism in the Small Island Caribbean. GeoJournal 1993, 31, 457–465. [Google Scholar] [CrossRef]
- March, A.; Failler, P.; Bennett, M. Caribbean Fishery and Aquaculture Financing Needs in the Blue Economy: Identifying Opportunities and Constraints in Barbados, Grenada, and St. Vincent and the Grenadines. J. Sustain. Res. 2023, 5, 1–37. [Google Scholar] [CrossRef]
- Burke, L.; Spalding, M. Shoreline Protection by the World’s Coral Reefs: Mapping the Benefits to People, Assets, and Infrastructure. Mar. Policy 2022, 146, 105311. [Google Scholar] [CrossRef]
- Ferrario, F.; Beck, M.W.; Storlazzi, C.D.; Micheli, F.; Shepard, C.C.; Airoldi, L. The Effectiveness of Coral Reefs for Coastal Hazard Risk Reduction and Adaptation. Nat. Commun. 2014, 5, 3794. [Google Scholar] [CrossRef] [PubMed]
- Hockings, M.; Leverington, F.; Cook, C. Protected Area Management Effectiveness. In Protected Area Governance and Management; ANU Press: Canberra, Australia, 2014; pp. 889–928. ISBN 978-1-925021-69-1. [Google Scholar]
- Hayek, M.; Salgues, M.; Souche, J.-C.; De Weerdt, K.; Pioch, S. How To Improve The Bioreceptivity of Concrete Infrastructure Used in Marine Ecosystems? Literature Review for Mechanisms, Key Factors and Colonization Effects. J. Coast. Res. 2023, 39, 553–568. [Google Scholar] [CrossRef]
- Cramer, K.L.; Jackson, J.B.C.; Donovan, M.K.; Greenstein, B.J.; Korpanty, C.A.; Cook, G.M.; Pandolfi, J.M. Widespread Loss of Caribbean Acroporid Corals Was Underway before Coral Bleaching and Disease Outbreaks. Sci. Adv. 2020, 6, eaax9395. [Google Scholar] [CrossRef]
- Fisheries, N. Elkhorn Coral|NOAA Fisheries. Available online: https://www.fisheries.noaa.gov/species/elkhorn-coral (accessed on 12 July 2022).
- Chamberland, V.F.; Vermeij, M.J.A.; Brittsan, M.; Carl, M.; Schick, M.; Snowden, S.; Schrier, A.; Petersen, D. Restoration of Critically Endangered Elkhorn Coral (Acropora palmata) Populations Using Larvae Reared from Wild-Caught Gametes. Glob. Ecol. Conserv. 2015, 4, 526–537. [Google Scholar] [CrossRef]
- Jackson, E.J.; Donovan, M.; Cramer, K.; Lam, V. Status and Trends of Caribbean Coral Reefs: 1970–2012; Global Coral Reef Monitoring Network; International Union for the Conservation of Nature (IUCN): Gland, Switzerland, 2014; p. 891. [Google Scholar]
- Cummings, K.; Zuke, A.; De Stasio, B.; Krumholz, J. Coral Growth Assessment on an Established Artificial Reef in Antigua. Ecol. Rest. 2015, 33, 90–95. [Google Scholar] [CrossRef]
- DeGeorges, A.; Goreau, T.J.; Reilly, B. Land-Sourced Pollution with an Emphasis on Domestic Sewage: Lessons from the Caribbean and Implications for Coastal Development on Indian Ocean and Pacific Coral Reefs. Sustainability 2010, 2, 2919–2949. [Google Scholar] [CrossRef]
- Lapointe, B. Nutrient Thresholds for Bottom-up Control of Macroalgal Blooms on Coral Reefs in Jamaica and Southeast Florida. Limnol. Oceanogr. 1997, 42 Pt 2, 1119–1131. [Google Scholar] [CrossRef]
- McWilliams, J.P.; Côté, I.M.; Gill, J.A.; Sutherland, W.J.; Watkinson, A.R. Accelerating Impacts of Temperature-Induced Coral Bleaching in the Caribbean. Ecology 2005, 86, 2055–2060. [Google Scholar] [CrossRef]
- Eakin, C.M.; Morgan, J.A.; Heron, S.F.; Smith, T.B.; Liu, G.; Alvarez-Filip, L.; Baca, B.; Bartels, E.; Bastidas, C.; Bouchon, C.; et al. Caribbean Corals in Crisis: Record Thermal Stress, Bleaching, and Mortality in 2005. PLoS ONE 2010, 5, e13969. [Google Scholar] [CrossRef]
- Spillman, C.M.; Heron, S.F.; Jury, M.R.; Anthony, K.R.N. Climate Change and Carbon Threats to Coral Reefs: National Meteorological and Ocean Services as Sentinels. Bull. Am. Meteorol. Soc. 2011, 92, 1581–1586. [Google Scholar] [CrossRef][Green Version]
- Lessios, H.A. The Great Diadema Antillarum Die-Off: 30 Years Later. Annu. Rev. Mar. Sci. 2016, 8, 267–283. [Google Scholar] [CrossRef]
- Heron, S.; Morgan, J.; Eakin, M.; Skirving, W. 3. Hurricanes and Their Effects on Coral Reefs. In Status of Caribbean Coral Reefs after Bleaching and Hurricanes; National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 2005; Volume 6. [Google Scholar]
- Wu, L.; Wang, B. What Has Changed the Proportion of Intense Hurricanes in the Last 30 Years? J. Clim. 2008, 21, 1432–1439. [Google Scholar] [CrossRef]
- Anderson, R.; Morrall, C.; Jossart, J.; Nimrod, S.; Bolda, E.; Musser, K.; Berg, C.; Balza, R. Marine Protected Area Monitoring in the Nearshore Waters of Grenada, Eastern Caribbean: Benthic Cover and Fish Populations. Rev. Biol. Trop. 2014, 62, 273–286. [Google Scholar] [CrossRef]
- Paxton, A.B.; Peterson, C.H.; Taylor, J.C.; Adler, A.M.; Pickering, E.A.; Silliman, B.R. Artificial Reefs Facilitate Tropical Fish at Their Range Edge. Commun. Biol. 2019, 2, 168. [Google Scholar] [CrossRef]
- Mofitt, R.B.; Parrish, F.A. Polovina, Jeffrey J Community Structure, Biomass and Productivity of Deepwater Artificial Reefs in Hawaii. Bull. Mar. Sci. 1989, 44, 616–630. [Google Scholar]
- Szedlmayer, S.T.; Shipp, R.L. Movement and Growth of Red Snapper, Lutjanus Campechanus, from an Artificial Reef Area in the Northeastern Gulf of Mexico. Bull. Mar. Sci. 1994, 55, 887–896. [Google Scholar]
- Collins, K.J.; Free, E.K.; Jensen, A.C.; Bannister, R.C.A. Lobster (Homarus Gammarus) Movement On an Artificial Reef: The Potential Use of Artificial Reefs for Stock Enhancement. Crustaceana 1994, 67, 198–211. [Google Scholar] [CrossRef]
- J⊘rgensen, D. An Oasis in a Watery Desert? Discourses on an Industrial Ecosystem in the Gulf of Mexico Rigs-to-Reefs Program. Hist. Technol. 2009, 25, 343–364. [Google Scholar] [CrossRef]
- Abelson, A. Artificial Reefs vs Coral Transplantation as Restoration Tools for Mitigating Coral Reef Deterioration: Benefits, Concerns, and Proposed Guidelines. Bull. Mar. Sci. 2006, 78, 151–159. [Google Scholar]
- Carr, M.; Hixon, M. Artificial Reefs: The Importance of Comparisons with Natural Reefs. Fisheries 1997, 22, 28–33. [Google Scholar] [CrossRef]
- Lymperaki, M.M.; Hill, C.E.L.; Hoeksema, B.W. The Effects of Wave Exposure and Host Cover on Coral-Associated Fauna of a Centuries-Old Artificial Reef in the Caribbean. Ecol. Eng. 2022, 176, 106536. [Google Scholar] [CrossRef]
- Ajemian, M.J.; Wetz, J.J.; Shipley-Lozano, B.; Shively, J.D.; Stunz, G.W. An Analysis of Artificial Reef Fish Community Structure along the Northwestern Gulf of Mexico Shelf: Potential Impacts of “Rigs-to-Reefs” Programs. PLoS ONE 2015, 10, e0126354. [Google Scholar] [CrossRef]
- Nakamura, M. Evolution of Artificial Fishing Reef Concepts in Japan. Bull. Mar. Sci. 1985, 37, 271–278. [Google Scholar]
- Pan, Y.; Tong, H.; Wei, D.; Xiao, W.; Xue, D. Review of Structure Types and New Development Prospects of Artificial Reefs in China. Front. Mar. Sci. 2022, 9, 853452. [Google Scholar] [CrossRef]
- Lukens, R.R. Guidelines For Marine Artificial Reef Materials—Second Edition. Gulf States Mar. Fish. Comm. 1997, 19, 205. [Google Scholar]
- Froehlich, C.Y.M.; Kline, R.J. Using Fish Population Metrics to Compare the Effects of Artificial Reef Density. PLoS ONE 2015, 10, e0139444. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R. Rebuilding, or Supplementing of, the Artificial Fishing Reefs in the Gulf of Mexico—Development Activities in Region V; Texas Parks and Wildlife Department: Austin, TX, USA, 1964. [Google Scholar]
- Pickering, H.; Whitmarsh, D.; Jensen, A. Artificial Reefs as a Tool to Aid Rehabilitation of Coastal Ecosystems: Investigating the Potential. Mar. Pollut. Bull. 1999, 37, 505–514. [Google Scholar] [CrossRef]
- Heery, E.C.; Bishop, M.J.; Critchley, L.P.; Bugnot, A.B.; Airoldi, L.; Mayer-Pinto, M.; Sheehan, E.V.; Coleman, R.A.; Loke, L.H.L.; Johnston, E.L.; et al. Identifying the Consequences of Ocean Sprawl for Sedimentary Habitats. J. Exp. Mar. Biol. Ecol. 2017, 492, 31–48. [Google Scholar] [CrossRef]
- Silva, R.; Mendoza, E.; Mariño-Tapia, I.; Martínez, M.; Escalante, E. An Artificial Reef Improves Coastal Protection and Provides a Base for Coral Recovery. J. Coast. Res. 2016, 75, 467–471. [Google Scholar] [CrossRef]
- Paxton, A.B.; Steward, D.N.; Harrison, Z.H.; Taylor, J.C. Fitting Ecological Principles of Artificial Reefs into the Ocean Planning Puzzle. Ecosphere 2022, 13, e3924. [Google Scholar] [CrossRef]
- Kim, T.; Baek, S.; Kwon, Y.; Lee, J.; Cha, S.M.; Kwon, S. Improved Coastal Erosion Prevention Using a Hybrid Method with an Artificial Coral Reef: Large-Scale 3D Hydraulic Experiment. Water 2020, 12, 2801. [Google Scholar] [CrossRef]
- Hylkema, A.; Hakkaart, Q.C.A.; Reid, C.B.; Osinga, R.; Murk, A.J.; Debrot, A.O. Artificial Reefs in the Caribbean: A Need for Comprehensive Monitoring and Integration into Marine Management Plans. Ocean Coast. Manag. 2021, 209, 105672. [Google Scholar] [CrossRef]
- Andskog, M.A.; Layman, C.; Allgeier, J.E. Seagrass Production around Artificial Reefs Is Resistant to Human Stressors. Proc. R. Soc. B Biol. Sci. 2023, 290, 20230803. [Google Scholar] [CrossRef]
- Risk, M.J. Artificial Reefs in Discovery Bay, Jamaica. Atoll Res. Bull. 1981, 255, 91–100. [Google Scholar]
- Schulze, A.; Erdner, D.L.; Grimes, C.J.; Holstein, D.M.; Miglietta, M.P. Artificial Reefs in the Northern Gulf of Mexico: Community Ecology Amid the “Ocean Sprawl”. Front. Mar. Sci. 2020, 7, 447. [Google Scholar] [CrossRef]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel Extensions (CPCe): A Visual Basic Program for the Determination of Coral and Substrate Coverage Using Random Point Count Methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Crosby, M.P.; Reese, E.S. Manual for Monitoring Coral Reefs with Indicator Species: Butterflyfishes as Indicators of Change on Indo-Pacific Reefs; National Oceanic and Atmospheric Administration: Silver Spring, MD, USA; Office of Ocean and Coastal Resource Management: Washington, DC, USA; Hawaii Institute of Marine Biology: Honolulu, HI, USA; United States Man and the Biosphere Program: Washington, DC, USA; Department of Defense: Washington, DC, USA; Legacy Resource Management: Washington, DC, USA, 1996. [Google Scholar]
- Smith, T.B.; Ennis, R.S.; Kadison, E.; Weinstein, D.W.; Jossart, J.; Gyory, J.; Henderson, L. The United States Virgin Islands Territorial Coral Reef Monitoring Program. Year 15 Annual Report; National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 2015; p. 288. [Google Scholar]
- Folpp, H.; Lowry, M.; Gregson, M.; Suthers, I.M. Colonization and Community Development of Fish Assemblages Associated with Estuarine Artificial Reefs. Braz. J. Ocean. 2011, 59, 55–67. [Google Scholar] [CrossRef]
- Bohnsack, J.A. Are High Densities of Fishes at Artificial Reefs the Result of Habitat Limitation or Behavioral Preference? Bull. Mar. Sci. 1989, 44, 15. [Google Scholar]
- Abelson, A. Comparison of the Development of Coral and Fish Communities on Rock-Aggregated Artificial Reefs in Eilat, Red Sea. ICES J. Mar. Sci. 2002, 59, S122–S126. [Google Scholar] [CrossRef]
- Sherman, R.L.; Gilliam, D.S.; Spieler, R.E. Artificial Reef Design: Void Space, Complexity, and Attractants. ICES J. Mar. Sci. 2002, 59, S196–S200. [Google Scholar] [CrossRef]
- Hylkema, A.; Debrot, A.O.; Osinga, R.; Bron, P.S.; Heesink, D.B.; Izioka, A.K.; Reid, C.B.; Rippen, J.C.; Treibitz, T.; Yuval, M.; et al. Fish Assemblages of Three Common Artificial Reef Designs during Early Colonization. Ecol. Eng. 2020, 157, 105994. [Google Scholar] [CrossRef]
- Saunders, M.I.; Doropoulos, C.; Bayraktarov, E.; Babcock, R.C.; Gorman, D.; Eger, A.M.; Vozzo, M.L.; Gillies, C.L.; Vanderklift, M.A.; Steven, A.D.L.; et al. Bright Spots in Coastal Marine Ecosystem Restoration. Curr. Biol. 2020, 30, R1500–R1510. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.B. Biology and Ecology of the Hydrocoral Millepora on Coral Reefs. Adv. Mar. Biol. 2006, 50, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Dubé, C.E.; Boissin, E.; Planes, S. Overgrowth of Living Scleractinian Corals by the Hydrocoral Millepora Platyphylla in Moorea, French Polynesia. Mar. Biodiv. 2016, 46, 329–330. [Google Scholar] [CrossRef]
- Hill, C.E.L.; Lymperaki, M.M.; Hoeksema, B.W. A Centuries-Old Manmade Reef in the Caribbean Does Not Substitute Natural Reefs in Terms of Species Assemblages and Interspecific Competition. Mar. Pollut. Bull. 2021, 169, 112576. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Diaz, C.P.; Toledo-Hernández, C.; Sánchez-González, J.L.; Betancourt, B. The Effects of Depth-Related Environmental Factors on Traits in Acropora Cervicornis Raised in Nurseries. Water 2022, 14, 212. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Paul, V.J.; Arnold, S.N.; Steneck, R.S. Larval Settlement Preferences and Post-Settlement Survival of the Threatened Caribbean Corals Acropora Palmata and A. Cervicornis. Coral Reefs 2010, 29, 71–81. [Google Scholar] [CrossRef]
- Monchanin, C.; Mehrotra, R.; Haskin, E.; Scott, C.M.; Urgell Plaza, P.; Allchurch, A.; Arnold, S.; Magson, K.; Hoeksema, B.W. Contrasting Coral Community Structures between Natural and Artificial Substrates at Koh Tao, Gulf of Thailand. Mar. Environ. Res. 2021, 172, 105505. [Google Scholar] [CrossRef]
- Blouet, S.; Bramanti, L.; Guizien, K. Artificial Reefs Geographical Location Matters More than Shape, Age and Depth for Sessile Invertebrate Colonization in the Gulf of Lion (NorthWestern Mediterranean Sea). Peer Community J. 2022, 2, e24. [Google Scholar] [CrossRef]
- Blakeway, D.; Byers, M.; Stoddart, J.; Rossendell, J. Coral Colonisation of an Artificial Reef in a Turbid Nearshore Environment, Dampier Harbour, Western Australia. PLoS ONE 2013, 8, e75281. [Google Scholar] [CrossRef]
- La Mesa, G.; Longobardi, A.; Sacco, F.; Marino, G. First Release of Hatchery Juveniles of the Dusky Grouper Epinephelus Marginatus (Lowe, 1834) (Serranidae: Teleostei) at Artificial Reefs in the Mediterranean: Results from a Pilot Study. Sci. Mar. 2008, 72, 743–756. [Google Scholar] [CrossRef]
- Abecasis, D.; Bentes, L.; Lino, P.G.; Santos, M.N.; Erzini, K. Residency, Movements and Habitat Use of Adult White Seabream (Diplodus sargus) between Natural and Artificial Reefs. Estuar. Coast. Shelf Sci. 2013, 118, 80–85. [Google Scholar] [CrossRef]
- Higgins, E.; Metaxas, A.; Scheibling, R.E. A Systematic Review of Artificial Reefs as Platforms for Coral Reef Research and Conservation. PLoS ONE 2022, 17, e0261964. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.; Bell, J. Coastal Sponge Communities of the West Indian Ocean. Afr. J. Ecol. 2002, 40, 350–359. [Google Scholar] [CrossRef]
- Rallis, I.; Chatzigeorgiou, G.; Florido, M.; Sedano, F.; Procopiou, A.; Chertz-Bynichaki, M.; Vernadou, E.; Plaiti, W.; Koulouri, P.; Dounas, C.; et al. Early Succession Patterns of Benthic Assemblages on Artificial Reefs in the Oligotrophic Eastern Mediterranean Basin. J. Mar. Sci. Eng. 2022, 10, 620. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Benayahu, Y. Recruitment of Benthic Organisms onto a Planned Artificial Reef: Shifts in Community Structure One Decade Post-Deployment. Mar. Environ. Res. 2005, 59, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Brandt, M.E.; Olinger, L.K.; Chaves-Fonnegra, A.; Olson, J.B.; Gochfeld, D.J. Coral Recruitment Is Impacted by the Presence of a Sponge Community. Mar. Biol. 2019, 166, 49. [Google Scholar] [CrossRef]
- Rutzler, K.; van Soest, R. Carla Piantoni Sponges (Porifera) of the Gulf of Mexico. In Gulf of Mexico Origins, and Biota; Felder, D.L., Camp, D.K., Eds.; Texas A&M University Press: College Station, TX, USA, 2009; Volume 1, p. 1393. ISBN 1-60344-094-1. [Google Scholar]
- Carter, A.; Prekel, S. Benthic Colonization and Ecological Successional Patterns on a Planned Nearshore Artificial Reef System in Broward County, SE Florida. In Proceedings of the 11th International Coral Reef Symposium, Ft. Lauderdale, FL, USA, 7–11 July 2008. [Google Scholar]
- Carballo, J.L.; Bautista, E.; Nava, H.; Cruz-Barraza, J.A.; Chávez, J.A. Boring Sponges, an Increasing Threat for Coral Reefs Affected by Bleaching Events. Ecol. Evol. 2013, 3, 872–886. [Google Scholar] [CrossRef]
- Rose, C.S.; Risk, M.J. Increase in Cliona Delitrix Infestation of Montastrea Cavernosa Heads on an Organically Polluted Portion of the Grand Cayman Fringing Reef. Mar. Ecol. 1985, 6, 345–363. [Google Scholar] [CrossRef]
- Rutzler, K.; Riegert, G. Sponge Burrowing: Fine Structurc of Cliona Tampa Penetrating Calcareous Substrata. Mar. Biol. 1973, 21, 144–162. [Google Scholar] [CrossRef]
- Nava, H.; Carballo, J.L. Chemical and Mechanical Bioerosion of Boring Sponges from Mexican Pacific Coral Reefs. J. Exp. Biol. 2008, 211, 2827–2831. [Google Scholar] [CrossRef]
- Baird, A.H.; Morse, A.N.C. Induction of Metamorphosis in Larvae of the Brooding Corals Acropora palifera and Stylophora pistillata. Mar. Freshw. Res. 2004, 55, 469–472. [Google Scholar] [CrossRef]
- Kuffner, I.B.; Walters, L.J.; Becerro, M.A.; Paul, V.J.; Ritson-Williams, R.; Beach, K.S. Inhibition of Coral Recruitment by Macroalgae and Cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 11. [Google Scholar] [CrossRef]
- Birrell, C.L.; McCook, L.J.; Willis, B.L.; Harrington, L. Chemical Effects of Macroalgae on Larval Settlement of the Broadcast Spawning Coral Acropora Millepora. Mar. Ecol. Prog. Ser. 2008, 362, 129–137. [Google Scholar] [CrossRef]
- Negri, A.P.; Webster, N.S.; Hill, R.T.; Heyward, A.J. Metamorphosis of Broadcast Spawning Corals in Response to Bacteria Isolated from Crustose Algae. Mar. Ecol. Prog. Ser. 2001, 223, 121–131. [Google Scholar] [CrossRef]
- O’Reilly, L.M.; Willerth, S.M. Evaluating the Biocompatibility of Ceramic Materials for Constructing Artificial Reefs. Front. Mar. Sci. 2024, 10, 1292584. [Google Scholar] [CrossRef]
- Petersen, L.-E.; Moeller, M.; Versluis, D.; Nietzer, S.; Kellermann, M.Y.; Schupp, P.J. Mono- and Multispecies Biofilms from a Crustose Coralline Alga Induce Settlement in the Scleractinian Coral Leptastrea purpurea. Coral Reefs 2021, 40, 381–394. [Google Scholar] [CrossRef]
- Chang Geun, C.; Jung, S.W.; Ahn, J.; Shimasaki, Y.; Kang, I.J. A Study on Marine Algal Succession and Community in Pyramid-Shaped Artificial Reef. J.-Fac. Agric. Kyushu Univ. 2019, 64, 95–99. [Google Scholar] [CrossRef]
- Easton, E.; Gaymer, C.; Friedlander, A.; Herlan, J. Effects of Herbivores, Wave Exposure and Depth on Benthic Coral Communities of the Easter Island Ecoregion. Mar. Freshw. Res. 2018, 69, 997–1006. [Google Scholar] [CrossRef]
- Olds, A.D.; Pitt, K.A.; Maxwell, P.S.; Connolly, R.M. Synergistic Effects of Reserves and Connectivity on Ecological Resilience. J. Appl. Ecol. 2012, 49, 1195–1203. [Google Scholar] [CrossRef]
- Fox, R. Quantifying Herbivory across a Coral Reef Depth Gradient. Mar. Ecol. Prog. Ser. 2007, 339, 49–59. [Google Scholar] [CrossRef]
- Granneman, J.E.; Steele, M.A. Effects of Reef Attributes on Fish Assemblage Similarity between Artificial and Natural Reefs. ICES J. Mar. Sci. 2015, 72, 2385–2397. [Google Scholar] [CrossRef]
- Champion, C.; Suthers, I.; Smith, J.A. Zooplanktivory Is a Key Process for Fish Production on a Coastal Artificial Reef. Mar. Ecol. Prog. Ser. 2015, 541, 1–14. [Google Scholar] [CrossRef]
- Crabbe, M.; Carlin, J.P. Industrial Sedimentation Lowers Coral Growth Rates in a Turbid Lagoon Environment, Discovery Bay, Jamaica. Int. J. Integr. Biol. 2007, 1, 37–40. [Google Scholar]
- Higgins, E.; Scheibling, R.E.; Desilets, K.M.; Metaxas, A. Benthic Community Succession on Artificial and Natural Coral Reefs in the Northern Gulf of Aqaba, Red Sea. PLoS ONE 2019, 14, e0212842. [Google Scholar] [CrossRef]
- Gann, G.D.; McDonald, T.; Walder, B.; Aronson, J.; Nelson, C.R.; Jonson, J.; Hallett, J.G.; Eisenberg, C.; Guariguata, M.R.; Liu, J.; et al. International Principles and Standards for the Practice of Ecological Restoration. Second Edition. Restor. Ecol. 2019, 27, S1–S46. [Google Scholar] [CrossRef]
- Hein, M.Y.; Vardi, T.; Shaver, E.C.; Pioch, S.; Boström-Einarsson, L.; Ahmed, M.; Grimsditch, G.; McLeod, I.M. Perspectives on the Use of Coral Reef Restoration as a Strategy to Support and Improve Reef Ecosystem Services. Front. Mar. Sci. 2021, 8, 299. [Google Scholar] [CrossRef]
- Boström-Einarsson, L.; Babcock, R.C.; Bayraktarov, E.; Ceccarelli, D.; Cook, N.; Ferse, S.C.A.; Hancock, B.; Harrison, P.; Hein, M.; Shaver, E.; et al. Coral Restoration—A Systematic Review of Current Methods, Successes, Failures and Future Directions. PLoS ONE 2020, 15, e0226631. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
LeFebre, B.; Saye, P.; Henkel, J.S. A Report on the Artificial Reef Use in Grenada, West Indies. J. Mar. Sci. Eng. 2024, 12, 253. https://doi.org/10.3390/jmse12020253
LeFebre B, Saye P, Henkel JS. A Report on the Artificial Reef Use in Grenada, West Indies. Journal of Marine Science and Engineering. 2024; 12(2):253. https://doi.org/10.3390/jmse12020253
Chicago/Turabian StyleLeFebre, Brianna, Phil Saye, and James S. Henkel. 2024. "A Report on the Artificial Reef Use in Grenada, West Indies" Journal of Marine Science and Engineering 12, no. 2: 253. https://doi.org/10.3390/jmse12020253
APA StyleLeFebre, B., Saye, P., & Henkel, J. S. (2024). A Report on the Artificial Reef Use in Grenada, West Indies. Journal of Marine Science and Engineering, 12(2), 253. https://doi.org/10.3390/jmse12020253