
Isolation and Cultivation of Carotenoid-Producing Strains from Tidal Flat Sediment and Proposal of Croceibacterium aestuarii sp. nov., a Novel Carotenoid-Producing Species in the Family Erythrobacteraceae



Abstract

1. Introduction
2. Materials and Methods
2.1. Isolation, Cultivation, Preservation, and Identification of Carotenoid-Producing Strains
2.2. Phylogenetic Relationship and Genomic Characteristics of Carotenoid-Producing Strain D39T
2.3. Phenotypic Determinations of Strain D39T
2.4. Carotenoid Determination and Biosynthetic Gene Analysis of Strain D39T
3. Results
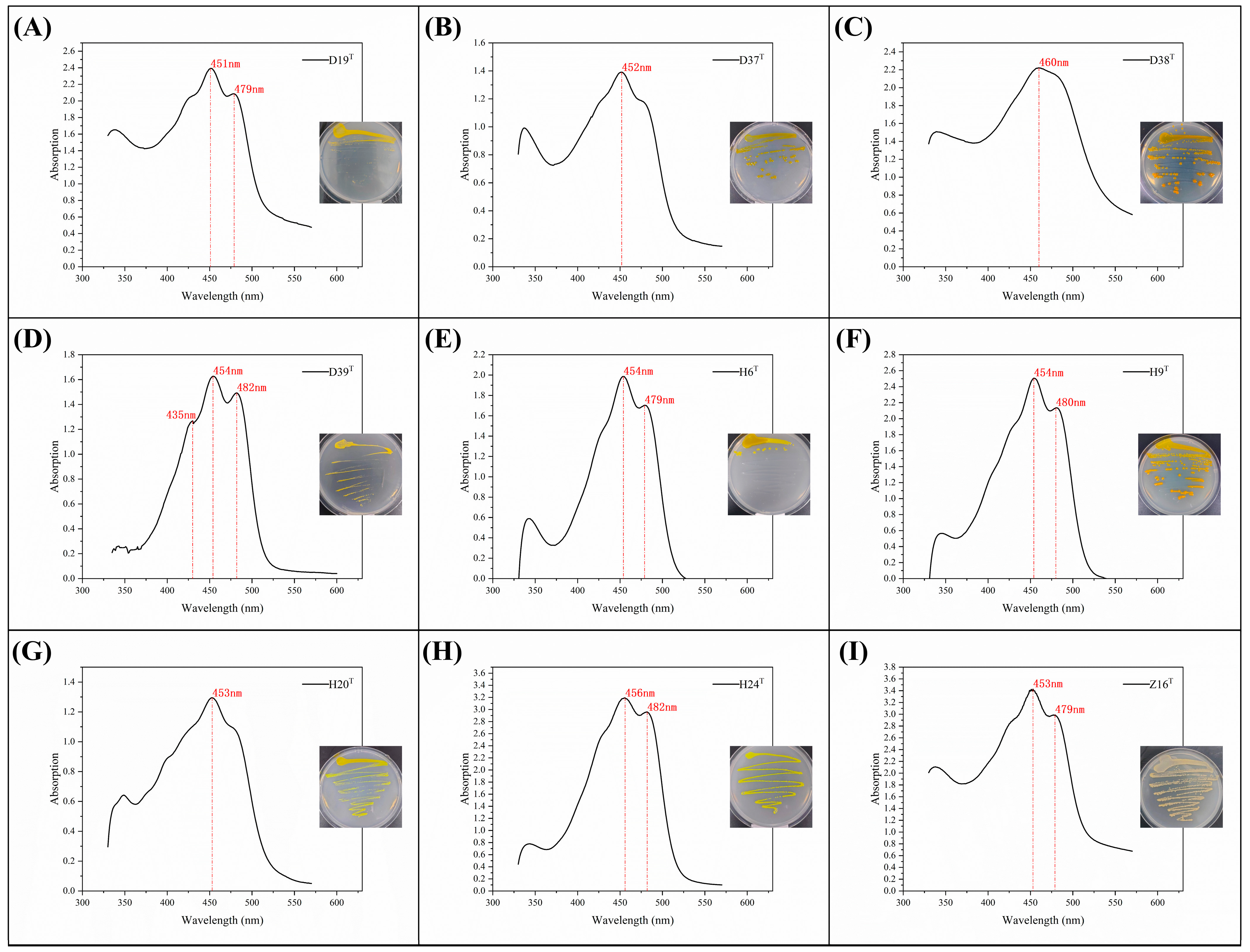
3.1. Bacterial Isolation of Tidal Flat Sediment
3.2. Phylogenetic Relationship and Genomic Comparison of Strain D39T
3.3. Phenotypic Characteristics of Strain D39T
3.4. Carotenoid Composition and Biosynthesis Pathway of Strain D39T
4. Discussion
4.1. Erythrobacteraceae and Flavobacteraceae as Dominant Carotenoid-Producing Taxa
4.2. Carotenoid Synthesis in the Genus Croceibacterium
4.3. Proposal of a Novel Species Croceibacterium aestuarii sp. nov.
- Description of Croceibacterium aestuarii sp. nov.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Soni, N.; Dhandhukia, P.; Thakker, J.N. Carotenoid from marine Bacillus infantis: Production, extraction, partial characterization, and its biological activity. Arch. Microbiol. 2023, 205, 161. [Google Scholar] [CrossRef]
- Maj, A.; Dziewit, L.; Drewniak, L.; Garstka, M.; Krucon, T.; Piatkowska, K.; Gieczewska, K.; Czarnecki, J.; Furmanczyk, E.; Lasek, R.; et al. In vivo creation of plasmid pCRT01 and its use for the construction of carotenoid-producing Paracoccus spp. strains that grow efficiently on industrial wastes. Microb. Cell Fact. 2020, 19, 141. [Google Scholar] [CrossRef]
- Britton, G. Carotenoid research: History and new perspectives for chemistry in biological systems. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158699. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Mumtaz, R.; Tayyab, S.; Rehman, N.U.; Khan, A.L.; Shinwari, Z.K.; Al-Harrasi, A. Therapeutic applications of bacterial pigments: A review of current status and future opportunities. 3 Biotech 2018, 8, 207. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Gómez-Villegas, P.; Gonda, M.L.; León-Vaz, A.; León, R.; Mildenberger, J.; Rebours, C.; Saravia, V.; Vero, S.; Vila, E.; et al. Microalgae, seaweeds and aquatic bacteria, archaea, and yeasts: Sources of carotenoids with potential antioxidant and anti-inflammatory health-promoting actions in the sustainability era. Marine Drugs 2023, 21, 340. [Google Scholar] [CrossRef]
- Béjà, O.; Spudich, E.N.; Spudich, J.L.; Leclerc, M.; DeLong, E.F. Proteorhodopsin phototrophy in the ocean. Nature 2001, 411, 786–789. [Google Scholar] [CrossRef]
- Dieser, M.; Greenwood, M.; Foreman, C.M. Carotenoid pigmentation in antarctic heterotrophic bacteria as a strategy to withstand environmental stresses. Arct. Antarct. Alp. Res. 2010, 42, 396–405. [Google Scholar] [CrossRef]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef]
- Vila, E.; Hornero-Méndez, D.; Lareo, C.; Saravia, V. Biotechnological production of zeaxanthin by an Antarctic Flavobacterium: Evaluation of culture conditions. J. Biotechnol. 2020, 319, 54–60. [Google Scholar] [CrossRef]
- Lorenzo, Y.; Azqueta, A.; Luna, L.; Bonilla, F.; Domínguez, G.; Collins, A.R. The carotenoid beta-cryptoxanthin stimulates the repair of DNA oxidation damage in addition to acting as an antioxidant in human cells. Carcinogenesis 2009, 30, 308–314. [Google Scholar] [CrossRef]
- Setiyono, E.; Heriyanto; Pringgenies, D.; Shioi, Y.; Kanesaki, Y.; Awai, K.; Brotosudarmo, T.H.P. Sulfur-containing carotenoids from a marine coral symbiont Erythrobacter flavus strain KJ5. Mar. Drugs 2019, 17, 349. [Google Scholar] [CrossRef]
- Martín, J.F.; Gudiña, E.; Barredo, J.L. Conversion of β-Carotene into astaxanthin: Two separate enzymes or a bifunctional hydroxylase-ketolase protein? Microb. Cell Fact. 2008, 7, 3. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, Y.; Xu, X.W. Erythrobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; Wiley: Hoboken, NJ, USA, 2023; pp. 1–18. ISBN 978-1-118-96060-8. [Google Scholar]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Park, S.C.; Baik, K.S.; Choe, H.N.; Lim, C.H.; Kim, H.J.; Ka, J.-O.; Seong, C.N. Altererythrobacter namhicola sp. nov. and Altererythrobacter aestuarii sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2011, 61, 709–715. [Google Scholar] [CrossRef][Green Version]
- Gao, Y.; Li, G.; Fang, C.; Shao, Z.; Wu, Y.-H.; Xu, X.-W. Tsuneonella suprasediminis sp. nov., isolated from the Pacific Ocean. Int. J. Syst. Evol. Microbiol. 2019, 71, 004678. [Google Scholar] [CrossRef]
- He, Y.; Liu, Y.; Pei, T.; Duan, J.; Du, J.; Deng, X.; Zhu, H. Tsuneonella litorea sp. nov., a novel carotenoid-producing bacterium isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 2023, 73, 5785. [Google Scholar] [CrossRef]
- Wu, Y.-H.; Xu, L.; Meng, F.-X.; Zhang, D.-S.; Wang, C.-S.; Oren, A.; Xu, X.-W. Altererythrobacter atlanticus sp. nov., isolated from deep-sea sediment. Int. J. Syst. Evol. Microbiol. 2014, 64, 116–121. [Google Scholar] [CrossRef]
- Fidalgo, C.; Rocha, J.; Martins, R.; Proença, D.N.; Morais, P.V.; Henriques, I.; Alves, A. Altererythrobacter halimionae sp. nov. and Altererythrobacter endophyticus sp. nov., two endophytes from the salt marsh plant Halimione portulacoides. Int. J. Syst. Evol. Microbiol. 2017, 67, 3057–3062. [Google Scholar] [CrossRef]
- Nedashkovskaya, O.I.; Cho, S.-H.; Joung, Y.; Joh, K.; Kim, M.N.; Shin, K.-S.; Oh, H.W.; Bae, K.S.; Mikhailov, V.V.; Kim, S.B. Altererythrobacter troitsensis sp. nov., isolated from the sea urchin Strongylocentrotus intermedius. Int. J. Syst. Evol. Microbiol. 2013, 63, 93–97. [Google Scholar] [CrossRef]
- Tareen, S.; Risdian, C.; Müsken, M.; Wink, J. Alteriqipengyuania abyssalis sp. nov., a novel member of the class Alphaproteobacteria isolated from sponge, and emended description of the genus Alteriqipengyuania. Diversity 2021, 13, 670. [Google Scholar] [CrossRef]
- Liu, Y.-H.; Fang, B.-Z.; Dong, Z.-Y.; Li, L.; Mohamad, O.A.A.; Zhang, Y.-G.; Egamberdieva, D.; Xiao, M.; Li, W.-J. Croceibacterium gen. nov., with Description of Croceibacterium ferulae sp. nov., an endophytic bacterium isolated from Ferula sinkiangensis K. M. Shen and reclassification of Porphyrobacter mercurialis as Croceibacterium mercuriale comb. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 2547–2554. [Google Scholar] [CrossRef]
- Xu, L.; Sun, C.; Fang, C.; Oren, A.; Xu, X.-W. Genomic-based taxonomic classification of the family Erythrobacteraceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 4470–4495. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S ribosomal RNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Hildebrand, D.; Palleroni, N.; Hendson, M.; Toth, J.; Johnson, J. Pseudomonas flavescens sp. nov., isolated from walnut blight cankers. Int. J. Syst. Bacteriol. 1994, 44, 410–415. [Google Scholar] [CrossRef]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes de novo assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef]
- Zhang, Y.; Hua, J.; Ying, J.-J.; Dong, H.; Li, H.; Xamxidin, M.; Han, B.-N.; Sun, C.; Xu, L. Erythrobacter aurantius sp. nov., isolated from intertidal seawater in Taizhou. Int. J. Syst. Evol. Microbiol. 2022, 72, 5616. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Evolutionary trees from dna sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Kannan, L.; Wheeler, W.C. Maximum Parsimony on Phylogenetic Networks. Algorithms Mol. Biol. 2012, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Ouk Kim, Y.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Lechner, M.; Findeiss, S.; Steiner, L.; Marz, M.; Stadler, P.F.; Prohaska, S.J. Proteinortho: Detection of (co-)orthologs in large-scale analysis. BMC Bioinform. 2011, 12, 124. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Xu, L.; Wu, Y.-H.; Jian, S.-L.; Wang, C.-S.; Wu, M.; Cheng, L.; Xu, X.-W. Pseudohongiella nitratireducens sp. nov., isolated from seawater, and emended description of the genus Pseudohongiella. Int. J. Syst. Evol. Microbiol. 2016, 66, 5155–5160. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huo, Y.-Y.; Li, Z.-Y.; Wang, C.-S.; Oren, A.; Xu, X.-W. Chryseobacterium profundimaris sp. nov., a new member of the family Flavobacteriaceae isolated from deep-sea sediment. Antonie Van. Leeuwenhoek 2015, 107, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Xu, L.; Yu, X.-Y.; Zhao, Z.; Wu, Y.-H.; Oren, A.; Wang, C.-S.; Xu, X.-W. Minwuia thermotolerans gen. nov., sp. nov., a marine bacterium forming a deep branch in the Alphaproteobacteria, and proposal of Minwuiaceae fam. nov. and Minwuiales ord. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 3856–3862. [Google Scholar] [CrossRef] [PubMed]
- Minnikin, D.E. Chemical principles in the organization of lipid components in the mycobacterial cell envelope. Res. Microbiol. 1991, 142, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant. Cell 1995, 7, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Inbaraj, B.S.; Lu, H.; Hung, C.F.; Wu, W.B.; Lin, C.L.; Chen, B.H. Determination of carotenoids and their esters in fruits of Lycium barbarum Linnaeus by HPLC-DAD-APCI-MS. J. Pharm. Biomed. Anal. 2008, 47, 812–818. [Google Scholar] [CrossRef]
- Siddaramappa, S.; Viswanathan, V.; Thiyagarajan, S.; Narjala, A. Genomewide characterisation of the genetic diversity of carotenogenesis in bacteria of the order Sphingomonadales. Microb. Genom. 2018, 4, e000172. [Google Scholar] [CrossRef]
- Boutet, E.; Lieberherr, D.; Tognolli, M.; Schneider, M.; Bansal, P.; Bridge, A.J.; Poux, S.; Bougueleret, L.; Xenarios, I. UniProtKB/Swiss-Prot, the manually annotated section of the UniProt knowledgebase: How to use the entry view. Methods Mol. Biol. 2016, 1374, 23–54. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16s ribosomal RNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Kim, D.; Yoo, Y.; Khim, J.S.; Yang, D.; Pathiraja, D.; Park, B.; Choi, I.-G.; Kim, J.-J. Altererythrobacter lutimaris sp. nov., a marine bacterium isolated from a tidal flat and reclassification of Altererythrobacter deserti, Altererythrobacter estronivorus and Altererythrobacter muriae as Tsuneonella deserti comb. nov., Croceicoccus estronivorus comb. nov. and Alteripontixanthobacter muriae comb. nov. Int. J. Syst. Evol. Microbiol. 2021, 71, 005134. [Google Scholar] [CrossRef]
- Zhang, K.; Han, C.H.; Ling, J.; Yin, J. Alteriqipengyuania flavescens sp. nov., isolated from pearl river estuary sediment. Int. J. Syst. Evol. Microbiol. 2023, 73, 6161. [Google Scholar] [CrossRef]
- Lee, S.D.; Kim, I.S. Aurantiacibacter rhizosphaerae sp. nov., isolated from a rhizosphere mudflat of a halophyte and proposal to reclassify Erythrobacter suaedae Lee et al. 2019. and Erythrobacter flavus Yoon et al. 2003 as Aurantiacibacter suaedae comb. nov. and Qipengyuania flava comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2020, 70, 6257–6265. [Google Scholar] [CrossRef]
- Kim, C.-H.; Yoo, Y.; Kim, D.; Khim, J.S.; Pathiraja, D.; Kim, B.; Choi, I.-G.; Kim, J.-J. Aurantiacibacter sediminis sp. nov., a marine bacterium isolated from a tidal flat. Int. J. Syst. Evol. Microbiol. 2022, 72, 5406. [Google Scholar] [CrossRef]
- Pei, T.; Liu, Y.; Du, J.; Huang, K.; Deng, M.-R.; Zhu, H. Croceicoccus gelatinilyticus sp. nov., isolated from a tidal flat sediment. Arch. Microbiol. 2021, 204, 93. [Google Scholar] [CrossRef]
- Chen, B.; Ye, Y.; Lin, D.; Zhang, M.; Sun, J.; Tang, K. Croceicoccus hydrothermalis sp. nov., isolated from shallow-sea hydrothermal system off Kueishantao Island. Int. J. Syst. Evol. Microbiol. 2022, 72, 5472. [Google Scholar] [CrossRef]
- Park, S.; Chen, S.; Yoon, J.-H. Erythrobacter insulae sp. nov., isolated from a tidal flat. Int. J. Syst. Evol. Microbiol. 2020, 70, 1470–1477. [Google Scholar] [CrossRef]
- Hu, W.; Li, Z.; Ou, H.; Wang, X.; Wang, Q.; Tao, Z.; Huang, S.; Huang, Y.; Wang, G.; Pan, X. Novosphingobium album sp. nov., Novosphingobium organovorum sp. nov. and Novosphingobium mangrovi sp. nov. with the organophosphorus pesticides degrading ability isolated from mangrove sediments. Int. J. Syst. Evol. Microbiol. 2023, 73, 5843. [Google Scholar] [CrossRef]
- Chaudhary, D.K.; Dahal, R.H.; Kim, D.-U.; Kim, J. Novosphingobium olei sp. nov., with the ability to degrade diesel oil, isolated from oil-contaminated soil and proposal to reclassify Novosphingobium stygium as a later heterotypic synonym of Novosphingobium aromaticivorans. Int. J. Syst. Evol. Microbiol. 2021, 71, 4628. [Google Scholar] [CrossRef]
- He, X.; Lu, H.; Hu, W.; Deng, T.; Gong, X.; Yang, X.; Song, D.; He, M.; Xu, M. Novosphingobium percolationis sp. nov. and Novosphingobium huizhouense sp. nov., isolated from landfill leachate of a domestic waste treatment plant. Int. J. Syst. Evol. Microbiol. 2022, 72, 5394. [Google Scholar] [CrossRef]
- Pira, H.; Risdian, C.; Müsken, M.; Schupp, P.J.; Wink, J. Winogradskyella luteola sp. nov., Erythrobacter ani sp. nov., and Erythrobacter crassostrea sp. nov., isolated from the hemolymph of the pacific oyster Crassostrea gigas. Arch. Microbiol. 2022, 204, 488. [Google Scholar] [CrossRef]
- Bowers, R.M.; Kyrpides, N.C.; Stepanauskas, R.; Harmon-Smith, M.; Doud, D.; Reddy, T.B.K.; Schulz, F.; Jarett, J.; Rivers, A.R.; Eloe-Fadrosh, E.A.; et al. Minimum information about a single amplified genome (MISAG) and a metagenome-assembled genome (MIMAG) of bacteria and archaea. Nat. Biotechnol. 2017, 35, 725–731. [Google Scholar] [CrossRef]
- Liang, X.; Lin, H.; Wang, K.; Liao, Y.; Lai, Q.; Xu, Y.; Wang, C. Altererythrobacter salegens sp. nov., a slightly halophilic bacterium isolated from surface sediment. Int. J. Syst. Evol. Microbiol. 2017, 67, 909–913. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, H.-R.; Han, Q.-Q.; He, A.-L.; Nie, C.-Y.; Wang, S.-M.; Zhang, J.-L. Altererythrobacter soli sp. nov., isolated from desert sand. Int. J. Syst. Evol. Microbiol. 2017, 67, 454–459. [Google Scholar] [CrossRef]
- Ye, Y.-L.; Ma, K.-J.; Fu, Y.-H.; Wu, Z.-C.; Fu, G.-Y.; Sun, C.; Xu, X.-W. The heterogeneity of microbial diversity and its drivers in two types of sediments from tidal flats in Beibu Gulf, China. Front. Mar. Sci. 2023, 10, 1256393. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, B.X.; Ren, H.; Zhang, J. Composition and functional diversity of microbial community across a mangrove-inhabited mudflat as revealed by 16S rDNA gene sequences. Sci. Total Environ. 2018, 633, 518–528. [Google Scholar] [CrossRef]
- Yue, Y.; Rong, H.; Yang, Z.; Pan, X.; Chen, Y.; Yang, M. Microbial diversity and functional profiling in coastal tidal flat sediment with pollution of nutrients and potentially toxic elements. J. Soils Sediments 2023, 23, 2935–2950. [Google Scholar] [CrossRef]
- Zhou, Z.; Meng, H.; Liu, Y.; Gu, J.-D.; Li, M. Stratified bacterial and archaeal community in mangrove and intertidal wetland mudflats revealed by high throughput 16S ribosomal RNA gene sequencing. Front. Microbiol. 2017, 8, 2148. [Google Scholar] [CrossRef]
- Jiang, X.-T.; Peng, X.; Deng, G.-H.; Sheng, H.-F.; Wang, Y.; Zhou, H.-W.; Tam, N.F.-Y. Illumina sequencing of 16S ribosomal RNA tag revealed spatial variations of bacterial communities in a mangrove wetland. Microb. Ecol. 2013, 66, 96–104. [Google Scholar] [CrossRef]
- Wang, C.; Sun, D.; Junaid, M.; Xie, S.; Xu, G.; Li, X.; Tang, H.; Zou, J.; Zhou, A. Effects of tidal action on the stability of microbiota, antibiotic resistance genes, and microplastics in the Pearl River Estuary, Guangzhou, China. Chemosphere 2023, 327, 138485. [Google Scholar] [CrossRef]
- Luo, T.; Huang, Z.; Li, X.; Zhang, Y. Anaerobic microbe mediated arsenic reduction and redistribution in coastal wetland soil. Sci. Total Environ. 2020, 727, 138630. [Google Scholar] [CrossRef]
- Lv, X.; Ma, B.; Yu, J.; Chang, S.X.; Xu, J.; Li, Y.; Wang, G.; Han, G.; Bo, G.; Chu, X. Bacterial community structure and function shift along a successional series of tidal flats in the Yellow River Delta. Sci. Rep. 2016, 6, 36550. [Google Scholar] [CrossRef]
- Dinter, T.; Geihser, S.; Gube, M.; Daniel, R.; Kuzyakov, Y. Impact of sea level change on coastal soil organic matter, priming effects and prokaryotic community assembly. FEMS Microbiol. Ecol. 2019, 95, fiz129. [Google Scholar] [CrossRef]
- Muwawa, E.M.; Obieze, C.C.; Makonde, H.M.; Jefwa, J.M.; Kahindi, J.H.P.; Khasa, D.P. 16S ribosomal RNA gene amplicon-based metagenomic analysis of bacterial communities in the rhizospheres of selected mangrove species from Mida Creek and Gazi Bay, Kenya. PLoS ONE 2021, 16, e0248485. [Google Scholar] [CrossRef]
- Lee, H.; Lee, D.W.; Kwon, S.L.; Heo, Y.M.; Jang, S.; Kwon, B.-O.; Khim, J.S.; Kim, G.-H.; Kim, J.-J. Importance of functional diversity in assessing the recovery of the microbial community after the Hebei Spirit Oil spill in Korea. Environ. Int. 2019, 128, 89–94. [Google Scholar] [CrossRef]
- Choi, H.J.; Jeong, T.-Y.; Yoon, H.; Oh, B.Y.; Han, Y.S.; Hur, M.J.; Kang, S.; Kim, J.-G. Comparative microbial communities in tidal flats sediment on Incheon, South Korea. J. Gen. Appl. Microbiol. 2018, 64, 232–239. [Google Scholar] [CrossRef]
- Lee, D.W.; Lee, H.; Lee, A.H.; Kwon, B.-O.; Khim, J.S.; Yim, U.H.; Kim, B.S.; Kim, J.-J. Microbial community composition and PAHs removal potential of indigenous bacteria in oil contaminated sediment of Taean coast, Korea. Environ. Pollut. 2018, 234, 503–512. [Google Scholar] [CrossRef]
- Boey, J.S.; Mortimer, R.; Couturier, A.; Worrallo, K.; Handley, K.M. Estuarine microbial diversity and nitrogen cycling increase along sand-mud gradients independent of salinity and distance. Environ. Microbiol. 2022, 24, 50–65. [Google Scholar] [CrossRef]
- Vogt, J.C.; Abed, R.M.M.; Albach, D.C.; Palinska, K.A. Bacterial and archaeal diversity in hypersaline cyanobacterial mats along a transect in the intertidal flats of the Sultanate of Oman. Microb. Ecol. 2018, 75, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Leontidou, K.; Abad-Recio, I.L.; Rubel, V.; Filker, S.; Däumer, M.; Thielen, A.; Lanzén, A.; Stoeck, T. Simultaneous analysis of seven 16S ribosomal RNA hypervariable gene regions increases efficiency in marine bacterial diversity detection. Environ. Microbiol. 2023, 25, 3484–3501. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.-W.; Ying, J.-J.; Dong, H.; Liu, W.-J.; He, D.-Y.; Xu, L.; Sun, C. Characterization of Maribacter polysaccharolyticus sp. nov., Maribacter huludaoensis sp. nov., and Maribacter zhoushanensis sp. nov. and illumination of the distinct adaptative strategies of the genus Maribacter. Front. Mar. Sci. 2023, 10, 1248754. [Google Scholar] [CrossRef]
- Takagi, T.; Aoyama, K.; Motone, K.; Aburaya, S.; Yamashiro, H.; Miura, N.; Inoue, K. Mutualistic interactions between dinoflagellates and pigmented bacteria mitigate environmental stress. Microbiol. Spectr. 2023, 11, e0246422. [Google Scholar] [CrossRef] [PubMed]
- Steven, R.; Humaira, Z.; Natanael, Y.; Dwivany, F.M.; Trinugroho, J.P.; Dwijayanti, A.; Kristianti, T.; Tallei, T.E.; Emran, T.B.; Jeon, H.; et al. Marine microbial-derived resource exploration: Uncovering the hidden potential of marine carotenoids. Mar. Drugs 2022, 20, 352. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pei, T.; Du, J.; Yao, Q.; Deng, M.-R.; Zhu, H. Comparative genomics reveals genetic diversity and metabolic potentials of the genus Qipengyuania and suggests fifteen novel species. Microbiol. Spectr. 2022, 10, e0126421. [Google Scholar] [CrossRef] [PubMed]
- Kanso, S.; Greene, A.C.; Patel, B.K.C. Bacillus subterraneus sp. nov., an iron- and manganese-reducing bacterium from a deep subsurface Australian thermal aquifer. Int. J. Syst. Evol. Microbiol. 2002, 52, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Igreja, W.S.; Maia, F.d.A.; Lopes, A.S.; Chisté, R.C. Biotechnological production of carotenoids using low cost-substrates is influenced by cultivation parameters: A review. Int. J. Mol. Sci. 2021, 22, 8819. [Google Scholar] [CrossRef]
- Castresana, J. Topological variation in single-gene phylogenetic trees. Genome Biol. 2007, 8, 216. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Zafar, J.; Aqeel, A.; Shah, F.I.; Ehsan, N.; Gohar, U.F.; Moga, M.A.; Festila, D.; Ciurea, C.; Irimie, M.; Chicea, R. Biochemical and Immunological implications of lutein and zeaxanthin. Int. J. Mol. Sci. 2021, 22, 10910. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Average Nucleotide Identity (%) | In Silico DNA-DNA Hybridization (%) |
|---|---|---|
| C. atlanticum DSM 100738T | 73.4 | 14.8 |
| C. ferulae KCTC 62090T | 73.5 | 14.7 |
| C.mercuriale DSM 29971T | 73.9 | 14.8 |
| C.salegens MCCC 1K01500T | 83.0 | 44.2 |
| C. soli MCCC 1K02066T | 77.2 | 20.5 |
| C. selenioxidans MCCC 1K08007T | 74.4 | 16.6 |
| C. xixiisoli CGMCC 1.12804T | 73.4 | 14.3 |
| Characteristic | 1 | 2 | 3 |
|---|---|---|---|
| Temperature for growth (°C) | |||
| Range | 20–45 | 10–35 | 4–37 |
| Optimum | 37 | 30 | 30–32 |
| NaCl concentration for growth (w/v, %) | |||
| Range | 0.5–5.0 | 2.0–10.0 | 0–2.0 |
| Optimum | 2.0 | 3.0–8.0 | 1.0 |
| pH for growth | |||
| Range | 5.0–9.5 | 6.5–8.0 | 6.5–12.0 |
| Optimum | 7.5–8.5 | 7.0–7.5 | 7.0–9.0 |
| Oxidase | + | − | + |
| Hydrolysis of | |||
| Casein, Tween 80 and tyrosine | − | − | + |
| Tween 20 | − | + | − |
| Tween 60 | + | − | − |
| Utilization of | |||
| L-Alanine and D-glucose | + | − | + |
| D-Galactose and D-maltose | + | − | − |
| Enzymatic activity (API ZYM) | |||
| Leucine arylamidase | − | + | + |
| Valine arylamidase | + | + | − |
| β-Galactosidase and β-glucuronidase | − | + | − |
| α-Glucosidase | − | + | − |
| Reduction of nitrate to nitrite | − | + | + |
| Acid production from | |||
| D-Galactose | + | + | − |
| D-Glucose and glycerol | + | − | + |
| D-Maltose and sucrose | + | − | − |
| Mannitol, salicin and D-trehalose | − | − | + |
| DNA G+C content (%, by genome) | 65.8 | 64.7 | 67.0 |
| Compounds | Class | Molecular Weight | Formula | Content (μg/g) |
|---|---|---|---|---|
| α-Carotene | carotenes | 536.44 | C40H56 | 42.6 ± 0.2 × 10−3 |
| β-Carotene | carotenes | 536.44 | C40H56 | 33.6 ± 0.1 × 10−3 |
| Lutein dimyristate | xanthophylls | 988.80 | C68H108O4 | 58.4 ± 0.9 × 10−3 |
| Zeaxanthin | xanthophylls | 568.43 | C40H56O2 | 412.4 ± 14.3 × 10−3 |
| Zeaxanthin dimyristate | xanthophylls | 989.00 | C68H108O4 | 3.0 ± 0.1 × 10−3 |
| Zeaxanthin dipalmitate | xanthophylls | 1045.10 | C72H116O4 | 57.9 ± 0.1 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.-Y.; Dong, H.; Zhang, Y.; Gao, J.-W.; Zhou, P.; Sun, C.; Xu, L. Isolation and Cultivation of Carotenoid-Producing Strains from Tidal Flat Sediment and Proposal of Croceibacterium aestuarii sp. nov., a Novel Carotenoid-Producing Species in the Family Erythrobacteraceae. J. Mar. Sci. Eng. 2024, 12, 99. https://doi.org/10.3390/jmse12010099
Sun X-Y, Dong H, Zhang Y, Gao J-W, Zhou P, Sun C, Xu L. Isolation and Cultivation of Carotenoid-Producing Strains from Tidal Flat Sediment and Proposal of Croceibacterium aestuarii sp. nov., a Novel Carotenoid-Producing Species in the Family Erythrobacteraceae. Journal of Marine Science and Engineering. 2024; 12(1):99. https://doi.org/10.3390/jmse12010099
Chicago/Turabian StyleSun, Xiao-Yan, Han Dong, Yu Zhang, Jia-Wei Gao, Peng Zhou, Cong Sun, and Lin Xu. 2024. "Isolation and Cultivation of Carotenoid-Producing Strains from Tidal Flat Sediment and Proposal of Croceibacterium aestuarii sp. nov., a Novel Carotenoid-Producing Species in the Family Erythrobacteraceae" Journal of Marine Science and Engineering 12, no. 1: 99. https://doi.org/10.3390/jmse12010099
APA StyleSun, X.-Y., Dong, H., Zhang, Y., Gao, J.-W., Zhou, P., Sun, C., & Xu, L. (2024). Isolation and Cultivation of Carotenoid-Producing Strains from Tidal Flat Sediment and Proposal of Croceibacterium aestuarii sp. nov., a Novel Carotenoid-Producing Species in the Family Erythrobacteraceae. Journal of Marine Science and Engineering, 12(1), 99. https://doi.org/10.3390/jmse12010099