
New Insight into the Genus Cladocroce (Porifera, Demospongiae) Based on Morphological and Molecular Data, with the Description of Two New Species

 ,
,  ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sponge Sample Collection
2.2. Morphological Characterization
2.3. Molecular Investigation
2.3.1. DNA Extraction, Amplification, and Sequencing
2.3.2. Phylogenetic Analysis
2.3.3. Species Delimitation
3. Results
- Class Demospongiae Sollas, 1885;
- Order Haplosclerida Topsent, 1928;
- Family Chalinidae Gray, 1867;
- Genus Cladocroce Topsent, 1892;
- Type species: Cladocroce fibrosa Topsent, 1892: 72;
- Cladocroce aculeata Pulitzer-Finali, 1982
Molecular Species Delimitation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguilar-Camacho, J.M.; Carballo, J.L. First record of Cladocroce (Porifera: Haplosclerida: Chalinidae) from the eastern pacific ocean with the description of Cladocroce reina sp. nov. Zootaxa 2010, 68, 65–68. [Google Scholar] [CrossRef]
- De Weerdt, W.H. Family Chalinidae Gray, 1867. In Systema Porifera; Springer: Boston, MA, USA, 2002; pp. 852–873. [Google Scholar]
- De Voogd, N.J.; Alvarez, B.; Boury-Esnault, N.; Carballo, J.L.; Cárdenas, P.; Díaz, M.-C.; Dohrmann, M.; Downey, R.; Hajdu, E.; Hooper, J.N.A.; et al. World Porifera Database. 2022. Available online: https://www.marinespecies.org/porifera (accessed on 12 October 2022).
- Santos, G.G.; Da Silva, L.P.; Alliz, A.; Pinheiro, U. Cladocroce caelum sp nov from the Brazilian coast; First record of the genus in the South Atlantic. Zootaxa 2014, 3847, 297–300. [Google Scholar] [CrossRef]
- Calcinai, B.; Bastari, A.; Makapedua, D.M.; Cerrano, C. Mangrove sponges from Bangka Island (North Sulawesi, Indonesia) with the description of a new species. J. Mar. Biol. Assoc. 2017, 97, 1417–1422. [Google Scholar] [CrossRef]
- Putchakarn, S.; De Weerdt, W.H.; Sonchaeng, P.; Van Soest, R.W.M. A New Species of Cladocroce Topsent, 1892 (Porifera, Haplosclerida) from the Gulf of Thailand. Beaufortia 2004, 54, 113–116. [Google Scholar]
- Núñez-Pons, L.; Calcinai, B.; Gates, R.D. Who’s there?—First morphological and DNA barcoding catalogue of the shallow Hawai’ian sponge fauna. PLoS ONE 2017, 12, e0189357. [Google Scholar] [CrossRef]
- Cerrano, C.; Azzini, F.; Bavestrello, G.; Calcinai, B.; Pansini, M.; Sarti, M.; Thung, D. Marine lakes of karst islands in Ha Long Bay (Vietnam). Chem. Ecol. 2006, 22, 489–500. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Bertolino, M.; Thung, D.C.; Pansini, M.; Sarti, M.; Núñez Pons, L.; Calcinai, B. The Ha Long Bay marine ecosystem. An unprecedented opportunity for evolutionary studies on marine taxa. In Innovations in Land, Water and Energy for Vietnam’s Sustainable Development; Springer Nature: Cham, Switzerland, 2020; pp. 45–52. [Google Scholar] [CrossRef]
- Di Camillo, C.G.; Gravili, C.; De Vito, D.; Pica, D.; Piraino, S.; Puce, S.; Cerrano, C. The importance of applying Standardised Integrative Taxonomy when describing marine benthic organisms and collecting ecological data. Invertebr. Syst. 2018, 32, 794–802. [Google Scholar] [CrossRef]
- Vicente, J.; Osberg, A.; Marty, M.J.; Rice, K.; Toonen, R.J. Influence of palatability on the feeding preferences of the endemic Hawaiian tiger cowrie for indigenous and introduced sponges. Mar. Ecol. Prog. Ser. 2020, 647, 109–122. [Google Scholar] [CrossRef]
- Vicente, J.; Webb, M.K.; Paulay, G.; Rakchai, W.; Timmers, M.A.; Jury, C.P.; Bahr, K.; Toonen, R.J. Unveiling hidden sponge biodiversity within the Hawaiian reef cryptofauna. Coral Reefs 2022, 41, 727–742. [Google Scholar] [CrossRef]
- Fava, F.; Ponti, M.; Scinto, A.; Calcinai, B.; Cerrano, C. Possible effects of human impacts on epibenthic communities and coral rubble features in the marine Park of Bunaken (Indonesia). Estuar. Coast. Shelf Sci. 2009, 85, 151–156. [Google Scholar] [CrossRef]
- Núñez-Pons, L.; Mazzella, V.; Rispo, F.; Efremova, J.; Calcinai, B. DNA Barcoding Procedures for Taxonomical and Phylogenetic Studies in Marine Animals: Porifera as a Case Study. Methods Mol. Biol. 2022, 2498, 195–223. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.R.; Rachello-Dolmen, P.G.; Parra-Velandia, F.; Schönberg, C.H.L.; Breeuwer, J.A.J.; van Soest, R.W.M. Molecular evidence of cryptic speciation in the “cosmopolitan” excavating sponge Cliona celata (Porifera, Clionaidae). Mol. Phylogenetics Evol. 2010, 56, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Wörheide, G.; Nichols, S.A.; Goldberg, J. Intragenomic variation of the rDNA internal transcribed spacers in sponges (Phylum Porifera): Implications for phylogenetic studies. Mol. Phylogenet. Evol. 2004, 33, 816–830. [Google Scholar] [CrossRef]
- Chombard, C.; Boury-Esnault, N.; Tillier, S. Reassessment of Homology of Morphological Characters in Tetractinellid Sponges Based on Molecular Data. Syst. Biol. 1998, 47, 351–366. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Erpenbeck, D.; Hooper, J.N.A.; Wörheide, G. CO1 phylogenies in diploblasts and the “Barcoding of Life”—Are we sequencing a suboptimal partition? Mol. Ecol. Notes 2006, 6, 550–553. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. 2010 Gateway Computing Environments Workshop, GCE 2010; IEEE Computer Society: Washington, DC, USA, 2010. [Google Scholar]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Smith, T.E.; Hong, W.; Zachariah, M.M.; Harper, M.K.; Matainaho, T.K.; Van Wagoner, R.M.; Ireland, C.M.; Vershinin, M. Single-molecule inhibition of human kinesin by adociasulfate-13 and -14 from the sponge Cladocroce aculeata. Proc. Natl. Acad. Sci. USA 2013, 110, 18880–18885. [Google Scholar] [CrossRef]
- Pulitzer-Finali, G. Some new or little-known sponges from the Great Barrier Reef of Australia. Bollettino dei Musei e degli Istituti Biologici dell’Università di Genova 1982, 48–49, 87–141. [Google Scholar]
- Van Soest, R.W.M. Sponges of the Guyana Shelf. Zootaxa 2017, 4217, 1–225. [Google Scholar] [CrossRef]
- Lundbeck, W. Porifera. (Part I) Homorrhaphidae and Heterorrhaphidae. In The Danish Ingolf-Expedition; Bianco Luno: Copenhagen, Denmark, 1902; Volume 6, pp. 1–108. [Google Scholar]
- Lehnert, H.; Stone, R.P. Two new species of sponges (Porifera, Demospongiae) from the Aleutian Islands, Alaska. J. Mar. Biol. Assoc. 2016, 96, 673–680. [Google Scholar] [CrossRef]
- Huang, D.; Meier, R.; Todd, P.A.; Chou, L.M. Slow mitochondrial COI sequence evolution at the base of the metazoan tree and its implications for DNA barcoding. J. Mol. Evol. 2008, 66, 167–174. [Google Scholar] [CrossRef]
- Tomascik, T.; Mah, A.J.; Anugerah, N.; Moosa, M.K. The Ecology of the Indonesian Sea Part Two; Dalhousie University: Halifax, NS, Canada, 1997; ISBN 978-1-4629-0503-4. [Google Scholar]
- Trang, C.T.T.; Thung, D.C.; Van Nam, L.; Kha, P.T.; Van Bach, N.; Ngoc, D.H. Assessment of Sea Water Quality in some Limestone Island and Archipelagos areas, Viet Nam. VNU J. Sci. Earth Environ. Sci. 2020, 36, 70–78. [Google Scholar] [CrossRef]
- Cárdenas, P.; Pérez, T.; Boury-Esnault, N. Sponge Systematics Facing New Challenges. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 61, pp. 79–209. ISBN 9780123877871. [Google Scholar]
- Calcinai, B.; Cerrano, C.; Núñez-Pons, L.; Pansini, M.; Thung, D.C.; Bertolino, M. A new species of Spongilla (Porifera, demospongiae) from a karst lake in Ha Long Bay (Vietnam). J. Mar. Sci. Eng. 2020, 8, 1008. [Google Scholar] [CrossRef]
- Vrijenhoek, R.C. Cryptic species, phenotypic plasticity, and complex life histories: Assessing deep-sea faunal diversity with molecular markers. Deep. Res. Part II Top. Stud. Oceanogr. 2009, 56, 1713–1723. [Google Scholar] [CrossRef]
- Yang, Q.; Franco, C.M.M.; Sorokin, S.J.; Zhang, W. Development of a multilocus-based approach for sponge (phylum Porifera) identification: Refinement and limitations. Sci. Rep. 2017, 7, 41422. [Google Scholar] [CrossRef] [PubMed]
- Debiasse, M.B.; Hellberg, M.E. Discordance between morphological and molecular species boundaries among Caribbean species of the reef sponge Callyspongia. Ecol. Evol. 2015, 5, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Itskovich, V. Intragenomic variation of rDNA internal transcribed spacers in the endemic Baikal sponge Lubomirskia baikalensis (Pallas, 1776) (Spongillida, Lubomirskiidae): Implications for Porifera barcoding. J. Great Lakes Res. 2020, 46, 62–66. [Google Scholar] [CrossRef]
- Subagio, I.B.; Setiawan, E.; Hariyanto, S.; Irawan, B. Spicule size variation in Xestospongia testudinaria Lamarck, 1815 at Probolinggo-Situbondo coastal. AIP Conf. Proc. 2017, 1854, 020034. [Google Scholar] [CrossRef]
- Bavestrello, G.; Bonito, J.; Sarà, M. Influence of depth on the size of sponge spicules. Sci. Mar. 1993, 57, 415–420. [Google Scholar]







{kind=link}
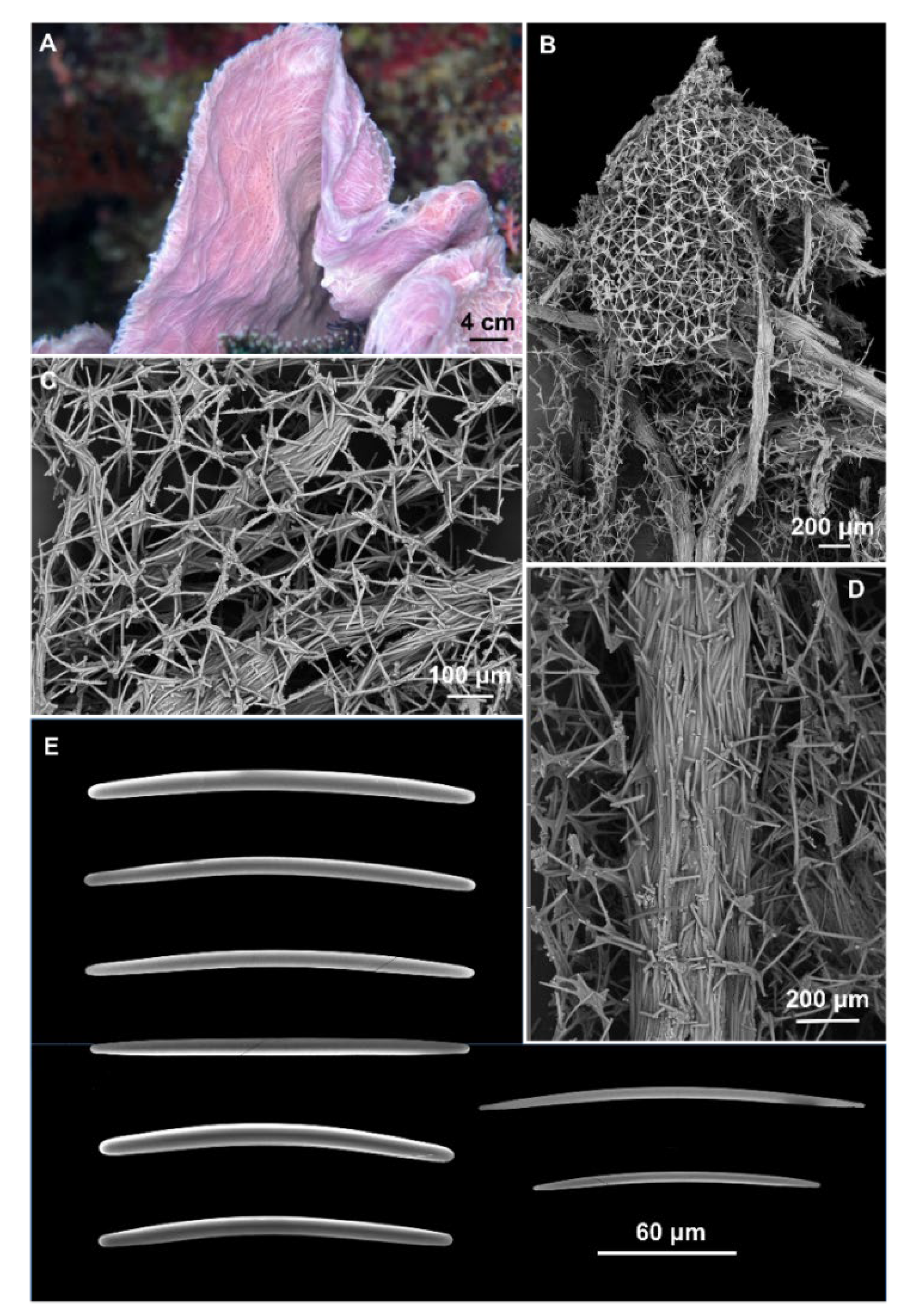{kind=link}
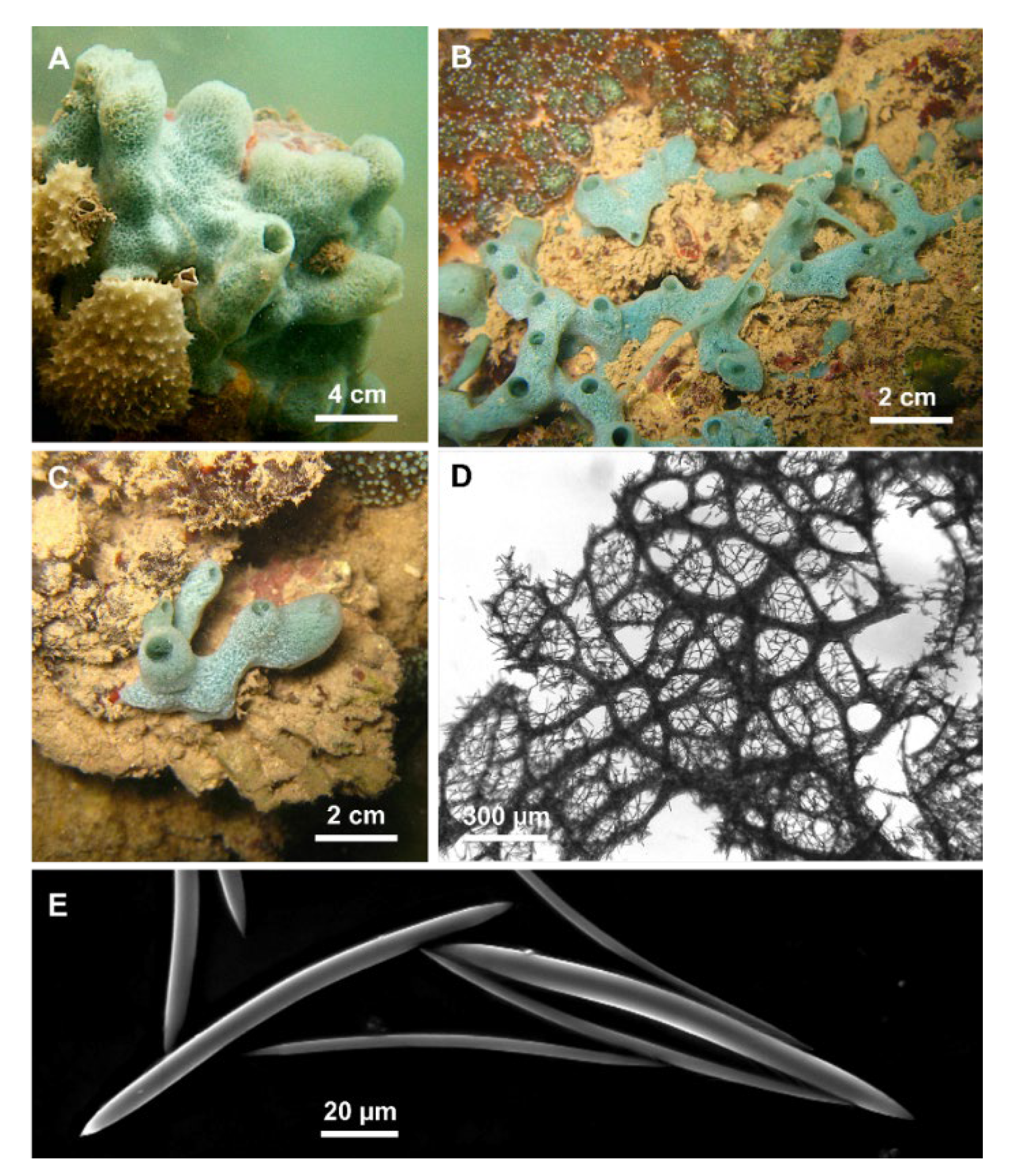{kind=link}
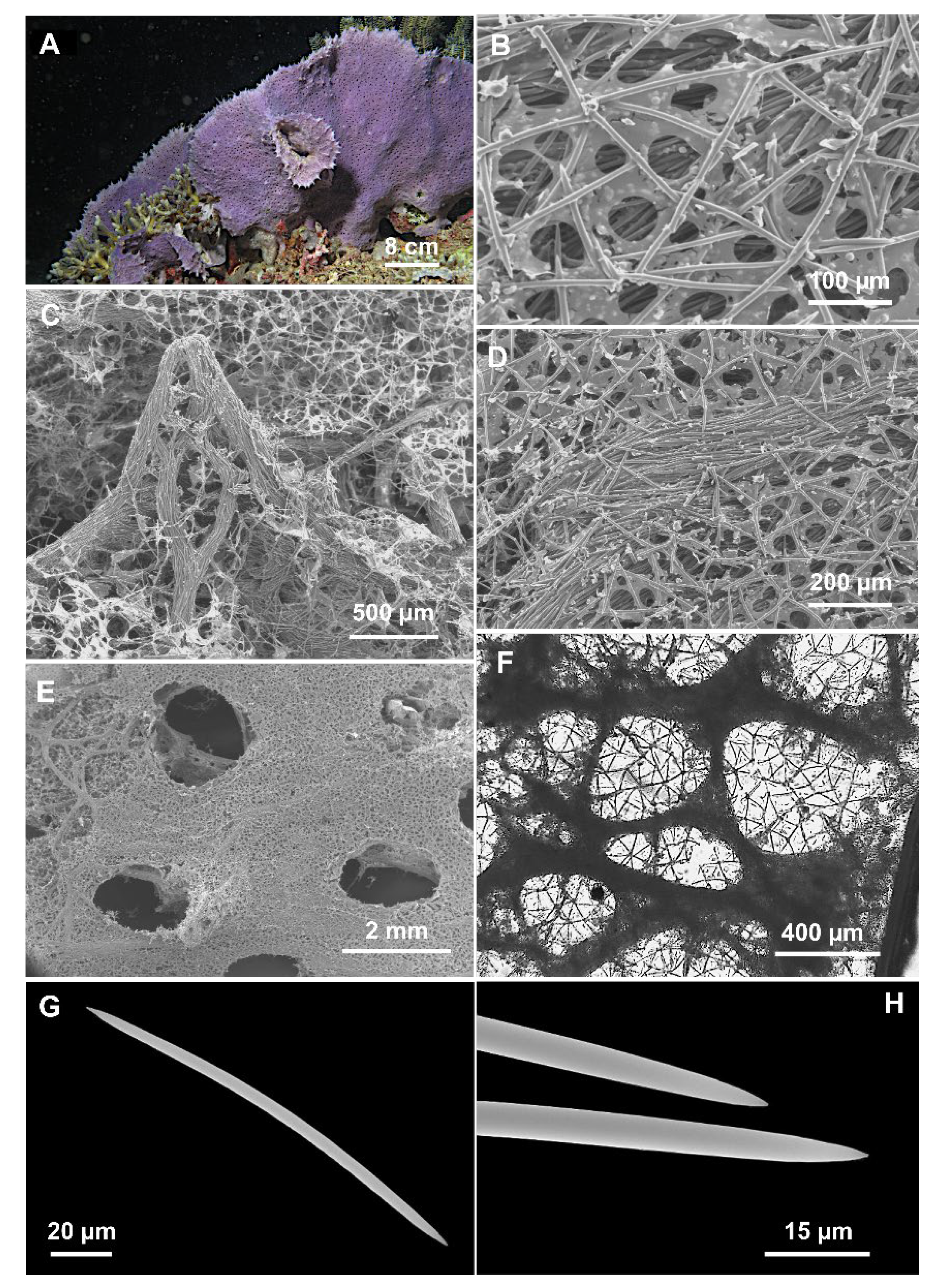{kind=link}
{kind=link}
{kind=link}
| Specimen | Locality | Shape | Color | Oxea Size (µm) | Fiber Thickness (µm) | Mesh Diameter (µm) | Depth (m) | References |
|---|---|---|---|---|---|---|---|---|
| BIMS-I1382 Holotype | Gulf of Thailand | Tubulo-ramose | Off-white to light cream | 105–117 × 6 | 23.5–59 | 131.4–499.6 | 2 | [6] |
| BIMS-I1383 | Gulf of Thailand | Tubulo-ramose | Light blue | 87.5–110 × 6–9.5 | / | / | low tide | [6] |
| MA 6 | Bangka Is. North Sulawesi | Cylindrical, repent | Light green to aquamarine | 90–(101 ± 3.7)–110 × 2.5–(3.9 ± 1.2)–5 | 30–75 | / | 0–1 | Present paper |
| MA 19a | Bangka Is. North Sulawesi | Tubular, repent | Light green to aquamarine | 95–(105.5 ± 5.5)–112 × 2.5–(4.2 ± 1)–5 | 22.7–46.4 | 207.4–562.9 | 0–1 | Present paper |
| HL 116/18 | Out Hang Du I lake, Vietnam | Tubular, repent | Light green to aquamarine | 90–(102 ± 6.7)–115 × 3.7–(4.5 ± 0.6)–5 | / | / | 1 | Present paper |
| HL 119/18 | Out Hang Du I Lake, Vietnam | Tubular, repent | Light green to aquamarine | 80–(97 ± 8.1)–115 × 3.7–(4.3 ± 0.6)–5 | / | / | 1 | Present paper |
| HL 121/18 | Out Hang Du I Lake, Vietnam | Tubular, repent | Light green to aquamarine | 90–(100.8 ± 6.3)–110 × 2.5–(4.5 ± 0.8)–5 | 16.1–69.1 | 104–562.1 | 1 | Present paper |
| HL 139 | Bui Xam Lake Vietnam | Massive, tubular | Light green to aquamarine | 80–(97.3 ± 8.5)–110 × 2.5–(3.9 ± 1)–5 | 20–80 | / | 1 | Present paper |
| HL 139/18 | Bui Xam Lake Vietnam | Tubular, repent | Light green to aquamarine | 95–(108 ± 16.5)–160 × 5 | 26.1–58.6 | 98.7–177 | 1 | Present paper |
| Samples | Oxeas (µm) |
|---|---|
| Holotype (MSNG 61503) TH A | 135–(147.5 ± 9.2)–195 × 3.7–(4.5 ± 0.6)–5 |
| Paratype (MSNG 61504) TH B | 125–(148.2 ± 9.7)–165 × 2.5–(5.3 ± 1.7)–7.5 |
| BA 7 | 145–(150.2 ± 47)–160 × 2.5–(4.1 ± 0.8)–5 |
| PH 9 | 155–(162 ± 8.3)–190 × 3.7–(4.7 ± 0.5)–5 |
| TH C | 130–(150 ± 13.3)–175 × 2.5–(3.8 ± 1)–5 |
| TH D | 145–(154.5 ± 5.5)–165 × 3.7–(5 ± 1)–7.5 |
| Specimen | Locality | Shape | Color | Oxea Size (µm) | Fiber Thickness (µm) | Mesh Diameter (µm) | Depth (m) | References |
|---|---|---|---|---|---|---|---|---|
| C. aculeata Pulitzer-Finali, 1982 | Indonesia | Lamellate | Violet | 120–180 × 2.5–10 | 45–175 | / | Shallow water | Present paper, [32] |
| C. attu Lehnert & Stone, 2013 | Alaska | Funnel | Golden brown | 158–183 × 14–16 | / | / | 358 | [4] |
| C. burapha Putchakarn et al., 2004 | Thailand, Indonesia, Vietnam | Tubulo-ramose, massive, tubular, repent | Off-white to light cream, light blue, light green to aquamarine | 80–160 × 2.5–9.5 | 16.1–80 | 98.7–562.9 | low tide, 0–2 | [6], present paper |
| C. caelum Santo et al., 2014 | Brazil | Massive tubular | Blue | 62–86 × 2–5 | 24–80 | / | 1 | [4] |
| C. fibrosa (Topsent, 1890) | Azores | Vase shaped | Grey | 600 × 18 | / | / | 1300 | [1] |
| C. gaussiana (Hentschel, 1914) | Antarctic | Tubular | Yellow | Oxeas: 230–285 Toxas: 80–180 | / | / | 200 | [1] |
| C. guyanensis Van Soest, 2017 | Guyana Shelf | Flabellate | Pale orange-brown | 336–414 × 9–19 | 80–120 | / | 104–130 | [33] |
| C. hyaline (Lundbeck, 1902) | Faroe Plateau | Erect, lamelliform | Hyaline | 268–315 × 10–14 | / | / | 471 | [34] |
| C. incurvata Lévi & Lévi, 1983 | New Caledonia | Lamellate | Ochre | 180–200 × 8–10 | / | / | 170–190 | [1] |
| C. infundibulum Lehnert & Stone, 2013 | Alaska | Funnel | Golden brown to light brown | 232–281 × 19–23 | 42–128 | / | 180 | [35] |
| C. kiska Lehnert & Stone, 2013 | Alaska | Flabellate | Golden brown | Oxeas: 252–343 × 10–18 Sigmas: 17–25 | / | / | 235 | [4] |
| C. lamellata sp. nov. | Indonesia | Lamellate | Violet, yellowish | 125–195 × 2.5–7.5 | 40–130 up to 500 | 400–1900 | 20 | Present paper |
| C. osculosa Topsent, 1927 | Portugal | Lamellate | Brown | 225 × 9 | / | / | 749–310 | [1] |
| C. pansinii sp. nov. | Vietnam, Thailand, Hawai’i | Massive tubular, tubulo-ramose, repent | Light green, light blue, light grey, violet | 125–200 × 3.7–15 | 25–376 | 166–991 | 1–15 | Present paper, [6,7] |
| C. parenchyma (Lundbeck, 1902) | Greenland | Tubular | Yellow | 239 × 9–12 | / | / | 2260 | [1] |
| C. reina Aguilar-Camacho & Carballo, 2010 | Mexico | Cushion, encrusting | Green, sky blue | 130–175 × 5–7.5 | 30–150 | 150–350 | 3 | [1] |
| C. spathiformis Topsent, 1904 | Azores | Tubular | Transparent | 375 × 17 | / | / | 1165 | [1] |
| C. spatula (Lundbeck, 1902) | Greenland | Lamellate | Yellow | 190–220 × 10–12 | / | / | 100 | [1] |
| C. toxifera Lehnert & Stone, 2016 | Alaska | Lobate | Reddish light brown | Oxeas: 241–297 × 12–23 Toxas: 36–139 × 3–7 | 50–230 | / | 93 | [35] |
| C. tubulosa Pulitzer-Finali, 1993 | Kenya | Tubular | Yellow | 64–74 × 3.5 | / | / | 16 | [1] |
| C. ventilabrum (Fristedt, 1887) | Behring Sea | Lamellate | Brownish | 250 | / | / | 512 | [1] |
| Samples | Locality | Shape | Color | Oxeas (um) | Fiber Thickness (µm) | Mesh Diameter (µm) | Depth (m) | References |
|---|---|---|---|---|---|---|---|---|
| Holotype (MSNG 52833) HL 19 | Coastal Site II, Vietnam | Massive tubular | Light green | 155–(174.4 ± 8.4)–185 × 7.5–(9.7 ± 1.6)–12.5 | 72.5–214 | 259.1–859.9 | 2–4 | Present paper |
| Paratype (MSNG 52834) HL 65 | Hang Toi Dark Cave, Vietnam | Massive tubular | Light green | 155–(176.9 ± 9.4)–190 × 7.5–(10 ± 1)–12.5 | 68–289 | 255.6–987 | 1–1.5 | Present paper |
| ZMA Por. 17921 Paratype | Gulf of Thailand | Tubulo-ramose | Light blue | 141–171 × 6–7.5 | 25–100 | 252–991 | 15 | [6] |
| SPO 29 | Hawai’i | Tubular | Light blue | 130–(147 ± 10)–167 × 5–(6.5 ± 0.9)–8.7 | 60–300 | 200–980 | 3–5 | [7] |
| SPO 32 | Hawai’i | Massive | Light grey | 125–(138.2 ± 10.1)–157.5 × 3.7–(6.2 ± 1.5)–8.7 | 60–300 | 200–980 | 3–5 | [7] |
| UF 3747 | Hawai’i | Massive | Violet | 138–147 | / | / | / | [11] |
| HL 60 | Hang Toi Dark Cave, Vietnam | Massive tubular | Light green | 160–(174 ± 8.2)–200 × 7.5–(9.5 ± 0.8)–10 | 85.3–179.8 | 328.9–629.3 | 1–1.5 | Present paper |
| HL 70 bis | Hang Toi Dark Cave, Vietnam | Massive tubular | Light green | 150–(166.6 ± 8.5)–180 × 7.5–(9.7 ± 1.2)–12.5 | 55.1–94.5 | 208.6–771.1 | 1–1.5 | Present paper |
| HL 95 | Bui Xam Lake, Vietnam | Repent habit | Light grey | 160–(177.8 ± 8.8)–200 × 7.5–(9.7 ± 1.2)–12.5 | 41–376 | 234–759 | 1.5 | Present paper |
| HL 110 | Hang Luong Lake (cove), Vietnam | Massive tubular | Light green | 155–(166.3 ± 7.2)–175 × 5–(9.6 ± 2.6)–12.5 | 70–205 | 182.9–856.7 | 2 | Present paper |
| HL 117/18 | out Hang Dui I Lake, Vietnam | Massive tubular | Light green | 145–(156.8 ± 8.4)–170 × 5–(8.4 ± 2.6)–12 | / | / | 2–3 | Present paper |
| HL 125/18 | Bui Xam Lake, Vietnam | Massive tubular | Light green | 135–(155.8 ± 12.5)–180 × 7.5–(11.7 ± 2.8)–15 | 49–200 | 166–848 | 2 | Present paper |
| HL 162/18 | Me Cung Lake, Vietnam | Massive tubular | Light green | 150–(167.6 ± 8.8)–185 × 7.5–(9.2 ± 1.1)–10 | / | / | 2 | Present paper |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertolino, M.; Cerrano, C.; Bavestrello, G.; Thung, D.C.; Núñez-Pons, L.; Rispo, F.; Efremova, J.; Mazzella, V.; Makapedua, D.M.; Calcinai, B. New Insight into the Genus Cladocroce (Porifera, Demospongiae) Based on Morphological and Molecular Data, with the Description of Two New Species. J. Mar. Sci. Eng. 2023, 11, 1240. https://doi.org/10.3390/jmse11061240
Bertolino M, Cerrano C, Bavestrello G, Thung DC, Núñez-Pons L, Rispo F, Efremova J, Mazzella V, Makapedua DM, Calcinai B. New Insight into the Genus Cladocroce (Porifera, Demospongiae) Based on Morphological and Molecular Data, with the Description of Two New Species. Journal of Marine Science and Engineering. 2023; 11(6):1240. https://doi.org/10.3390/jmse11061240
Chicago/Turabian StyleBertolino, Marco, Carlo Cerrano, Giorgio Bavestrello, Do Cong Thung, Laura Núñez-Pons, Francesca Rispo, Jana Efremova, Valerio Mazzella, Daisy Monica Makapedua, and Barbara Calcinai. 2023. "New Insight into the Genus Cladocroce (Porifera, Demospongiae) Based on Morphological and Molecular Data, with the Description of Two New Species" Journal of Marine Science and Engineering 11, no. 6: 1240. https://doi.org/10.3390/jmse11061240
APA StyleBertolino, M., Cerrano, C., Bavestrello, G., Thung, D. C., Núñez-Pons, L., Rispo, F., Efremova, J., Mazzella, V., Makapedua, D. M., & Calcinai, B. (2023). New Insight into the Genus Cladocroce (Porifera, Demospongiae) Based on Morphological and Molecular Data, with the Description of Two New Species. Journal of Marine Science and Engineering, 11(6), 1240. https://doi.org/10.3390/jmse11061240