
The Distribution of Ferritins in Marine Copepods




Abstract
1. Introduction
2. Materials and Methods
2.1. In Silico Workflow
2.2. Cladogram of Copepod Ferritin Genes
2.3. Expression of Ferritin in Calanus finmarchicus, Calanus helgolandicus and Temora stylifera
3. Results
3.1. Identification of Ferritins in Copepods
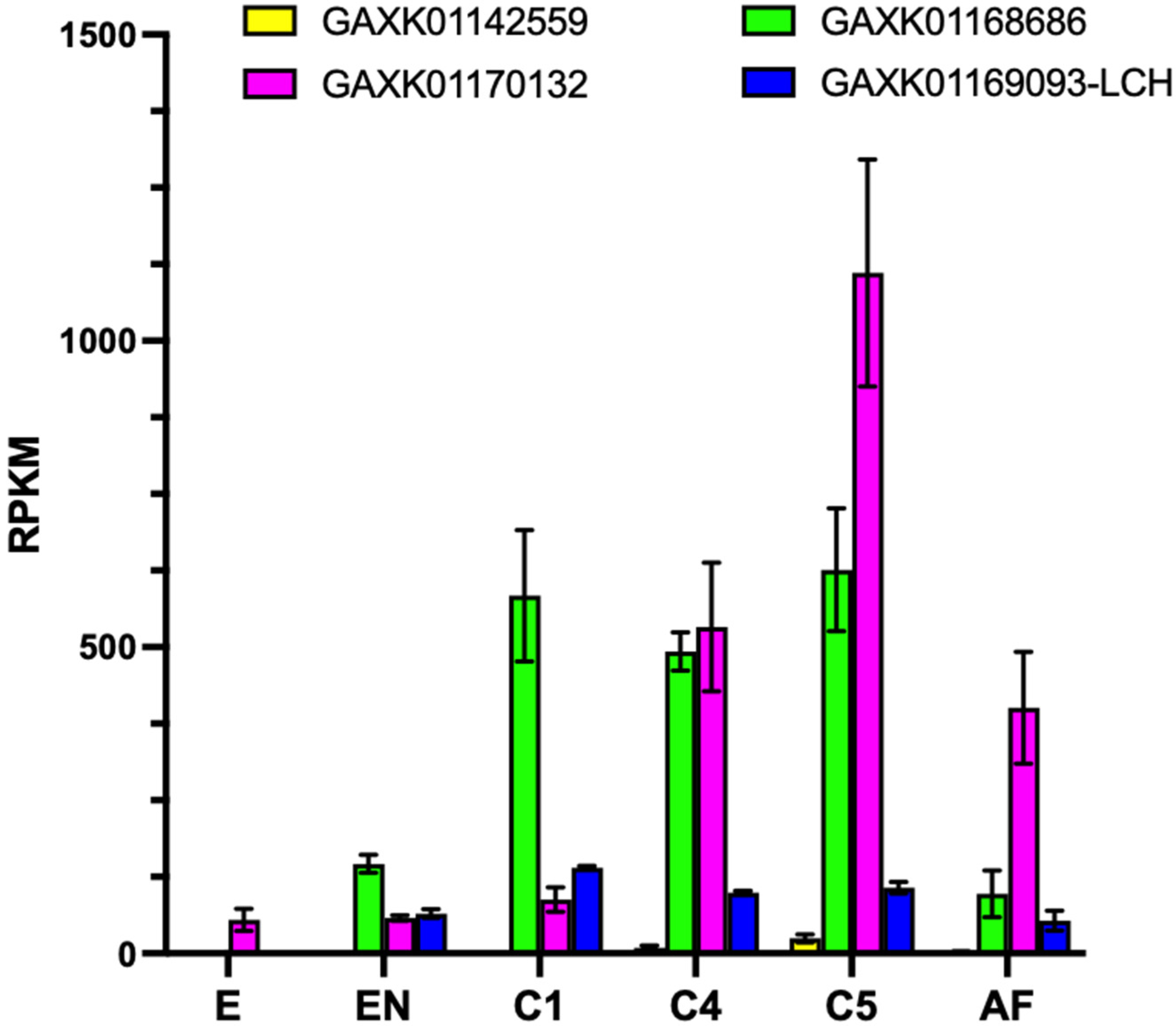
3.2. Expression of ferritin in C. finmarchicus, Calanus helgolandicus and Temora stylifera
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Theil, E.C. Ferritin: Structure, gene regulation, and cellular function in animals, plants, and microorganisms. Annu. Rev. Biochem. 1987, 56, 289–315. [Google Scholar] [CrossRef] [PubMed]
- Nichol, H.; Law, J.H.; Winzerling, J.J. Iron metabolism in insects. Annu. Rev. Entomol. 2002, 47, 535–559. [Google Scholar] [CrossRef] [PubMed]
- Arosio, P.; Ingrassia, R.; Cavadini, P. Ferritins: A family of molecules for iron storage, antioxidation and more. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2009, 1790, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Emerit, J.; Beaumont, C.; Trivin, F. Iron metabolism, free radicals, and oxidative injury. Biomed. Pharmacother. 2001, 55, 333–339. [Google Scholar] [CrossRef]
- Tang, X.; Zhou, B. Ferritin is the key to dietary iron absorption and tissue iron detoxification in Drosophila melanogaster. FASEB J. 2013, 27, 288–298. [Google Scholar] [CrossRef]
- Hamburger, A.E.; West, A.P., Jr.; Hamburger, Z.A.; Hamburger, P.; Bjorkman, P.J. Crystal structure of a secreted insect ferritin reveals a symmetrical arrangement of heavy and light chains. J. Mol. Biol. 2005, 349, 558–569. [Google Scholar] [CrossRef]
- Colbourne, J.K.; Eads, B.D.; Shaw, J.; Bohuski, E.; Bauer, D.J.; Andrews, J. Sampling Daphnia’s expressed genes: Preservation, expansion and invention of crustacean genes with reference to insect genomes. BMC Genom. 2007, 8, 217. [Google Scholar] [CrossRef]
- Zhou, C.; Hou, J.; Lin, D. A ferritin gene in the marine copepod Acartia tonsa as a highly sensitive biomonitor for nano-contamination. Aquat. Toxicol. 2022, 253, 106353. [Google Scholar] [CrossRef]
- Aguilera, V.M.; Vargas, C.A.; Lardies, M.A.; Poupin, M.J. Adaptive variability to low-pH river discharges in Acartia tonsa and stress responses to high PCO2 conditions. Mar. Ecol. 2016, 37, 215–226. [Google Scholar] [CrossRef]
- Petkeviciute, E.; Kania, P.W.; Skovgaard, A. Genetic responses of the marine copepod Acartia tonsa (Dana) to heat shock and epibiont infestation. Aquac. Rep. 2015, 2, 10–16. [Google Scholar] [CrossRef]
- Nilsson, B.; Jepsen, P.M.; Rewitz, K.; Hansen, B.W. Expression of hsp70 and ferritin in embryos of the copepod Acartia tonsa (Dana) during transition between subitaneous and quiescent state. J. Plankton Res. 2014, 36, 513–522. [Google Scholar] [CrossRef]
- Aruda, A.M.; Baumgartner, M.F.; Reitzel, A.M.; Tarrant, A.M. Heat shock protein expression during stress and diapause in the marine copepod Calanus finmarchicus. J. Insect Physiol. 2011, 57, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Tarrant, A.M.; Baumgartner, M.F.; Verslycke, T.; Johnson, C.L. Differential gene expression in diapausing and active Calanus finmarchicus (Copepoda). Mar. Ecol. Prog. Ser. 2008, 355, 193–207. [Google Scholar] [CrossRef]
- Heggland, E.I.; Tröße, C.; Eichner, C.; Nilsen, F. Heavy and light chain homologs of ferritin are essential for blood-feeding and egg production of the ectoparasitic copepod Lepeophtheirus salmonis. Mol. Biochem. Parasitol. 2019, 232, 111197. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Carotenuto, Y.; Roncalli, V. Glutathione S-transferases in marine copepods. J. Mar. Sci. Eng. 2021, 9, 1025. [Google Scholar] [CrossRef]
- Roncalli, V.; Uttieri, M.; Capua, I.D.; Lauritano, C.; Carotenuto, Y. Chemosensory-Related Genes in Marine Copepods. Mar. Drugs 2022, 20, 681. [Google Scholar] [CrossRef]
- Roncalli, V.; Lauritano, C.; Carotenuto, Y. First report of OvoA gene in marine arthropods: A new candidate stress biomarker in copepods. Mar. Drugs 2021, 19, 647. [Google Scholar] [CrossRef]
- Cieslak, M.C.; Castelfranco, A.M.; Roncalli, V.; Lenz, P.H.; Hartline, D.K. t-Distributed Stochastic Neighbor Embedding (t-SNE): A tool for eco-physiological transcriptomic analysis. Mar. Genom. 2020, 51, 100723. [Google Scholar] [CrossRef]
- Roncalli, V.; Cieslak, M.C.; Lenz, P.H. Transcriptomic responses of the calanoid copepod Calanus finmarchicus to the saxitoxin producing dinoflagellate Alexandrium fundyense. Sci. Rep. 2016, 6, 25708. [Google Scholar] [CrossRef]
- Asai, S.; Sanges, R.; Lauritano, C.; Lindeque, P.K.; Esposito, F.; Ianora, A.; Carotenuto, Y. De novo transcriptome assembly and gene expression profiling of the copepod Calanus helgolandicus feeding on the PUA-producing diatom Skeletonema marinoi. Mar. Drugs 2020, 18, 392. [Google Scholar] [CrossRef]
- Russo, E.; Lauritano, C.; d’Ippolito, G.; Fontana, A.; Sarno, D.; von Elert, E.; Ianora, A.; Carotenuto, Y. RNA-Seq and differential gene expression analysis in Temora stylifera copepod females with contrasting non-feeding nauplii survival rates: An environmental transcriptomics study. Bmc Genom. 2020, 21, 693. [Google Scholar] [CrossRef]
- Russo, E.; d’Ippolito, G.; Fontana, A.; Sarno, D.; D’Alelio, D.; Busseni, G.; Ianora, A.; von Elert, E.; Carotenuto, Y. Density-dependent oxylipin production in natural diatom communities: Possible implications for plankton dynamics. ISME J. 2020, 14, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Christie, A.E.; Fontanilla, T.M.; Nesbit, K.T.; Lenz, P.H. Prediction of the protein components of a putative Calanus finmarchicus (Crustacea, Copepoda) circadian signaling systems using a de novo assembled transcriptome. Comp. Biochem. Physiol. D-Genom. Proteom. 2013, 8, 165–193. [Google Scholar] [CrossRef] [PubMed]
- Roncalli, V.; Cieslak, M.C.; Passamaneck, Y.; Christie, A.E.; Lenz, P.H. Glutathione S-transferase (GST) gene diversity in the crustacean Calanus finmarchicus–contributors to cellular detoxification. PLoS ONE 2015, 10, e0123322. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Roncalli, V.; Ambrosino, L.; Cieslak, M.C.; Ianora, A. First de novo transcriptome of the copepod Rhincalanus gigas from Antarctic waters. Biology 2020, 9, 410. [Google Scholar] [CrossRef] [PubMed]
- Hartline, D.K.; Cieslak, M.C.; Castelfranco, A.M.; Lieberman, B.; Roncalli, V.; Lenz, P.H. De novo transcriptomes of six calanoid copepods (Crustacea): A resource for the discovery of novel genes. Sci. Data 2023, 10, 242. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Barreto, F.S.; Watson, E.T.; Lima, T.G.; Willett, C.S.; Edmands, S.; Li, W.; Burton, R.S. Genomic signatures of mitonuclear coevolution across populations of Tigriopus californicus. Nat. Ecol. Evol. 2018, 2, 1250–1257. [Google Scholar] [CrossRef]
- Andrews, S.C. The Ferritin-like superfamily: Evolution of the biological iron storeman from a rubrerythrin-like ancestor. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2010, 1800, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Uttieri, M. Trends in copepod studies. In Trends in Copepod Studies—Distribution, Biology and Ecology; Nova Science Publishers Inc.: New York, NY, USA, 2018; pp. 1–11. [Google Scholar]
- Jiang, J.-L.; Wang, G.-Z.; Mao, M.-G.; Wang, K.-J.; Li, S.-J.; Zeng, C.-S. Differential gene expression profile of the calanoid copepod, Pseudodiaptomus annandalei, in response to nickel exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2013, 157, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Hafez, T.; Bilbao, D.; Etxebarria, N.; Duran, R.; Ortiz-Zarragoitia, M. Application of a biological multilevel response approach in the copepod Acartia tonsa for toxicity testing of three oil Water Accommodated Fractions. Mar. Environ. Res. 2021, 169, 105378. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, B.; Jakobsen, H.H.; Stief, P.; Drillet, G.; Hansen, B.W. Copepod swimming behavior, respiration, and expression of stress-related genes in response to high stocking densities. Aquac. Rep. 2017, 6, 35–42. [Google Scholar] [CrossRef]
- Schminke, H.K. Entomology for the copepodologist. J. Plankton Res. 2007, 29, i149–i162. [Google Scholar] [CrossRef]
- Pham, D.Q.; Winzerling, J.J. Insect ferritins: Typical or atypical? Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2010, 1800, 824–833. [Google Scholar] [CrossRef]
- Whiten, S.R.; Eggleston, H.; Adelman, Z.N. Ironing out the details: Exploring the role of iron and heme in blood-sucking arthropods. Front. Physiol. 2018, 8, 1134. [Google Scholar] [CrossRef]
- Dunkov, B.C.; Georgieva, T.; Yoshiga, T.; Hall, M.; Law, J.H. Aedes aegypti ferritin heavy chain homologue: Feeding of iron or blood influences message levels, lengths and subunit abundance. J. Insect Sci. 2002, 2, 7. [Google Scholar] [CrossRef]
- Geiser, D.L.; Chavez, C.A.; Flores-Munguia, R.; Winzerling, J.J.; Pham, D.Q.D. Aedes aegypti ferritin: A cytotoxic protector against iron and oxidative challenge? Eur. J. Biochem. 2003, 270, 3667–3674. [Google Scholar] [CrossRef]
- Chen, T.; Amons, R.; Clegg, J.S.; Warner, A.H.; MacRae, T.H. Molecular characterization of artemin and ferritin from Artemia franciscana. Eur. J. Biochem. 2003, 270, 137–145. [Google Scholar] [CrossRef]
- Skottene, E.; Tarrant, A.M.; Olsen, A.J.; Altin, D.; Østensen, M.-A.; Hansen, B.H.; Choquet, M.; Jenssen, B.M.; Olsen, R.E. The β-oxidation pathway is downregulated during diapause termination in Calanus copepods. Sci. Rep. 2019, 9, 16686. [Google Scholar] [CrossRef] [PubMed]
- Kučerová, L.; Kubrak, O.I.; Bengtsson, J.M.; Strnad, H.; Nylin, S.; Theopold, U.; Nässel, D.R. Slowed aging during reproductive dormancy is reflected in genome-wide transcriptome changes in Drosophila melanogaster. Bmc Genom. 2016, 17, 50. [Google Scholar] [CrossRef] [PubMed]
- Lenz, P.H.; Roncalli, V. Diapause within the Context of Life-History Strategies in Calanid Copepods (Calanoida: Crustacea). Biol. Bull 2019, 237, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Lenz, P.H.; Roncalli, V.; Cieslak, M.C.; Tarrant, A.M.; Castelfranco, A.M.; Hartline, D.K. Diapause vs. reproductive programs: Transcriptional phenotypes in a keystone copepod. Commun. Biol. 2021, 4, 426. [Google Scholar] [CrossRef]
- Roncalli, V.; Cieslak, M.C.; Castelfranco, A.M.; Hopcroft, R.R.; Hartline, D.K.; Lenz, P.H. Post-diapause transcriptomic restarts: Insight from a high-latitude copepod. BMC Genom. 2021, 22, 409. [Google Scholar]
- Barroeta, Z.; Villate, F.; Uriarte, I.; Iriarte, A. Impact of Colonizer Copepods on Zooplankton Structure and Diversity in Contrasting Estuaries. Estuaries Coasts 2022, 45, 2592–2609. [Google Scholar] [CrossRef]
- Camatti, E.; Pansera, M.; Bergamasco, A. The copepod Acartia tonsa dana in a microtidal Mediterranean lagoon: History of a successful invasion. Water 2019, 11, 1200. [Google Scholar] [CrossRef]
- Carotenuto, Y.; Vitiello, V.; Gallo, A.; Libralato, G.; Trifuoggi, M.; Toscanesi, M.; Lofrano, G.; Esposito, F.; Buttino, I. Assessment of the relative sensitivity of the copepods Acartia tonsa and Acartia clausi exposed to sediment-derived elutriates from the Bagnoli-Coroglio industrial area. Mar. Environ. Res. 2020, 155, 104878. [Google Scholar] [CrossRef]
- Støttrup, J.G.; Richardson, K.; Kirkegaard, E.; Pihl, N.J. The cultivation of Acartia tonsa Dana for use as a live food source for marine fish larvae. Aquaculture 1986, 52, 87–96. [Google Scholar] [CrossRef]
- Gonzàlez-Porta, M.; Frankish, A.; Rung, J.; Harrow, J.; Brazma, A. Transcriptome analysis of human tissues and cell lines reveals one dominant transcript per gene. Genome Biol. 2013, 14, R70. [Google Scholar] [CrossRef]
- Taneri, B.; Snyder, B.; Gaasterland, T. Distribution of alternatively spliced transcript isoforms within human and mouse transcriptomes. J. Omics Res. 2011, 1, 1–5. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Order | Family | Ferritin Transcripts |
|---|---|---|---|
| Acartia clausi | Calanoida | Acartiidae | 1 |
| Acartia tonsa | Calanoida | Acartiidae | 2 |
| Calanoides acutus | Calanoida | Calanidae | 1 |
| Calanus finmarchicus | Calanoida | Calanidae | 4 |
| Calanus glacialis | Calanoida | Calanidae | 1 |
| Calanus helgolandicus | Calanoida | Calanidae | 4 |
| Calanus hyperboreus | Calanoida | Calanidae | 2 |
| Calanus marshallae | Calanoida | Calanidae | 3 |
| Calanus pacificus | Calanoida | Calanidae | 4 |
| Calanus propinquus | Calanoida | Calanidae | 2 |
| Centropages hamatus | Calanoida | Centropagidae | 3 |
| Epischura baikalensis | Calanoida | Temoridae | 1 |
| Eucyclops serrulatus | Cyclopoida | Cyclopidae | 3 |
| Hemidiaptomus amblyodon | Calanoida | Diaptomidae | 1 |
| Labidocera madurae | Calanoida | Pontellidae | 1 |
| Neocalanus cristatus | Calanoida | Calanidae | 1 |
| Neocalanus flemingeri | Calanoida | Calanidae | 6 |
| Neocalanus plumchrus | Calanoida | Calanidae | 1 |
| Paracyclopina nana | Cyclopoida | Cyclopettidae | 1 |
| Platychelipus ittoralis | Harpacticoida | Laophontidae | 4 |
| Pleuromamma xiphias | Calanoida | Metridinidae | 1 |
| Pseudodiaptomus annandalei | Calanoida | Pseudodiaptomidae | 2 |
| Rhincalanus gigas | Calanoida | Rhincalanidae | 3 |
| Temora longicornis | Calanoida | Temoridae | 6 |
| Temora stylifera | Calanoida | Temoridae | 4 |
| Tisbe furcata | Harpacticoida | Tisbidae | 1 |
| Tisbe holothuriae | Harpacticoida | Tisbidae | 1 |
| Species | Stressor | Regulation | Reference Paper |
|---|---|---|---|
| P. annandalei | Ni nanoparticles | + | 33 |
| A. tonsa | Ni nanoparticles | + | 8 |
| CdSe/ZnS quantum dots | + | 8 | |
| Heat shock (salinity dependent) | + | 10 | |
| Infestation C. vesiculosum | + | 10 | |
| Entry/emergence quiescence eggs | + | 11 | |
| Crowding | ns | 11 | |
| High pCO2 (only costal) | + | 9 | |
| WAF | ns | 34 | |
| Calanus spp. | Diapause (CV females) | + | 12 13 42 |
| L. salmonis | Starvation | − | 14 |
| C. finmarchicus | Diapause preparation (CV) vs. reproductive program | ns | 45 |
| Our study | |||
| C. finmarchicus | Copepodites | + | |
| A. fundyense | ns | ||
| C. helgolandicus | Oxylipin (lab) | ns | |
| T. stylifera | Oxylipin (field) | + | |
| N. flemingeri | Diapause emergence | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roncalli, V.; Uttieri, M.; Carotenuto, Y. The Distribution of Ferritins in Marine Copepods. J. Mar. Sci. Eng. 2023, 11, 1187. https://doi.org/10.3390/jmse11061187
Roncalli V, Uttieri M, Carotenuto Y. The Distribution of Ferritins in Marine Copepods. Journal of Marine Science and Engineering. 2023; 11(6):1187. https://doi.org/10.3390/jmse11061187
Chicago/Turabian StyleRoncalli, Vittoria, Marco Uttieri, and Ylenia Carotenuto. 2023. "The Distribution of Ferritins in Marine Copepods" Journal of Marine Science and Engineering 11, no. 6: 1187. https://doi.org/10.3390/jmse11061187
APA StyleRoncalli, V., Uttieri, M., & Carotenuto, Y. (2023). The Distribution of Ferritins in Marine Copepods. Journal of Marine Science and Engineering, 11(6), 1187. https://doi.org/10.3390/jmse11061187