

Echoes of the 2013–2015 Marine Heat Wave in the Eastern Bering Sea and Consequent Biological Responses



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Data and Methods
3. Formation, Evolution, and Propagation of the Blob to the Bering Sea
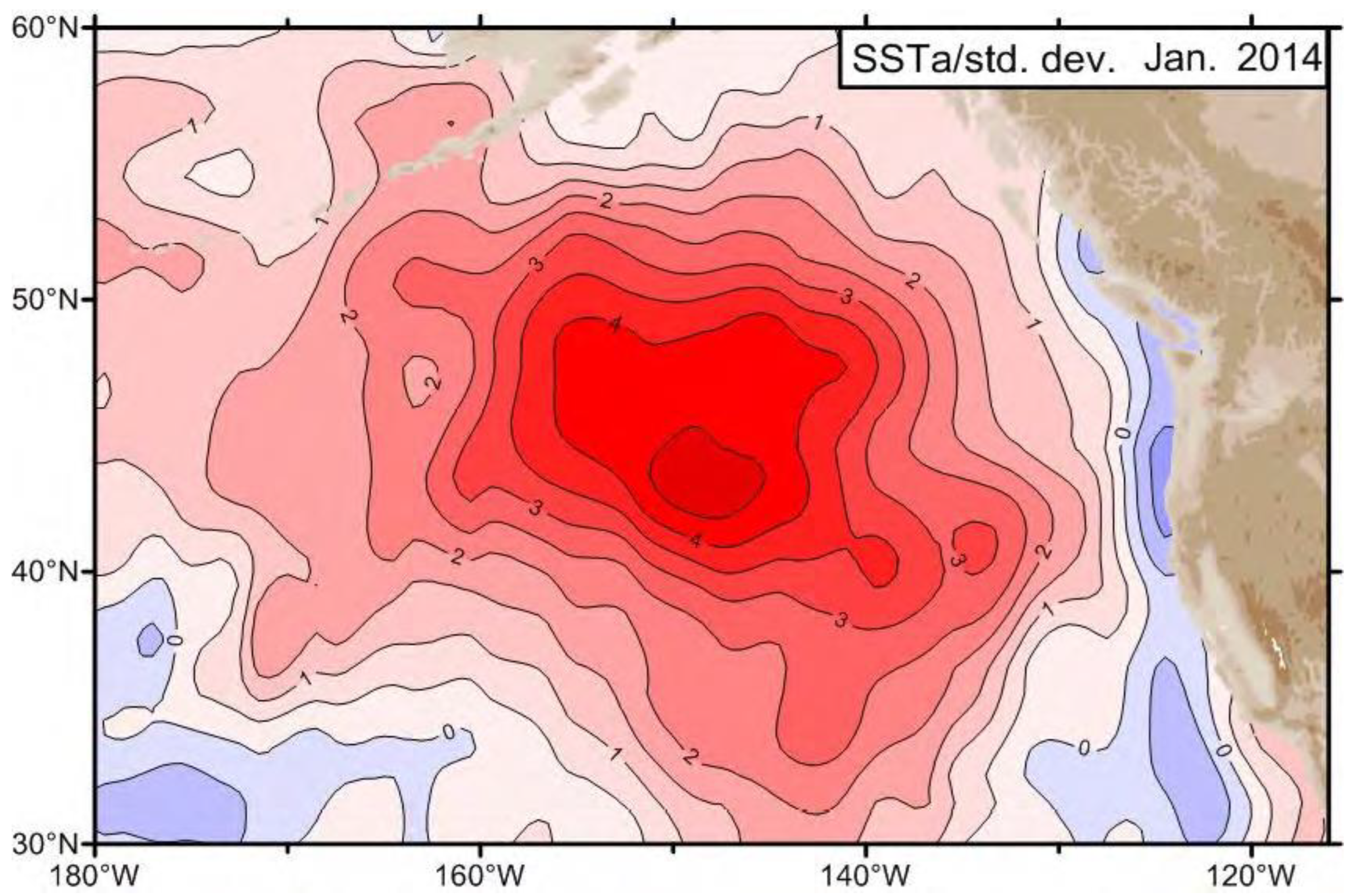
3.1. The Blob’s Emergence and Early Detection
3.2. The Blob’s Preconditioning in the Central North Pacific
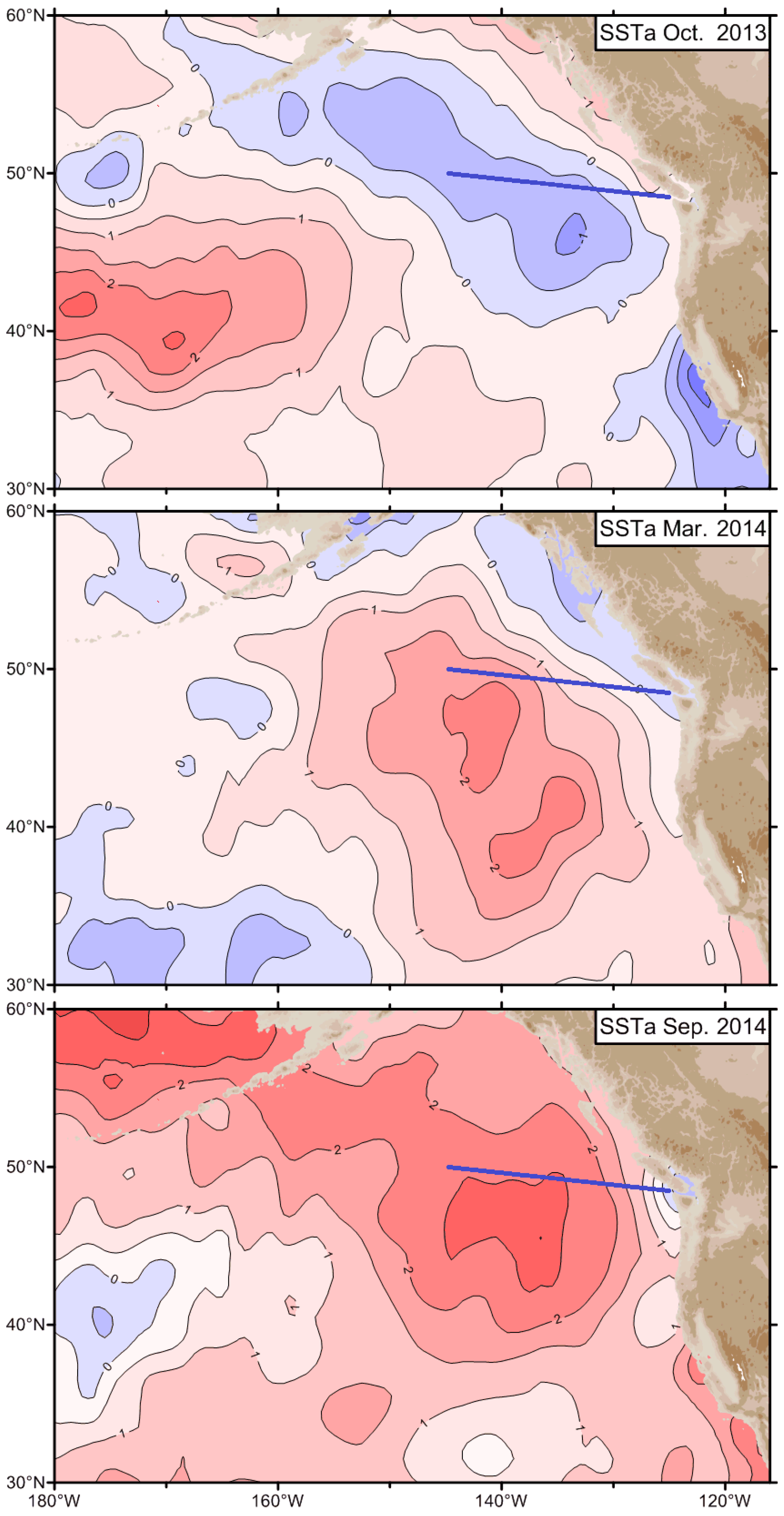
3.3. The Blob’s Advection along the Polar Front
3.4. The Blob’s Expansion into the Gulf of Alaska and Its Advection by Coastal Currents
3.5. The Blob’s Propagation into the Bering Sea and towards the Bering Strait
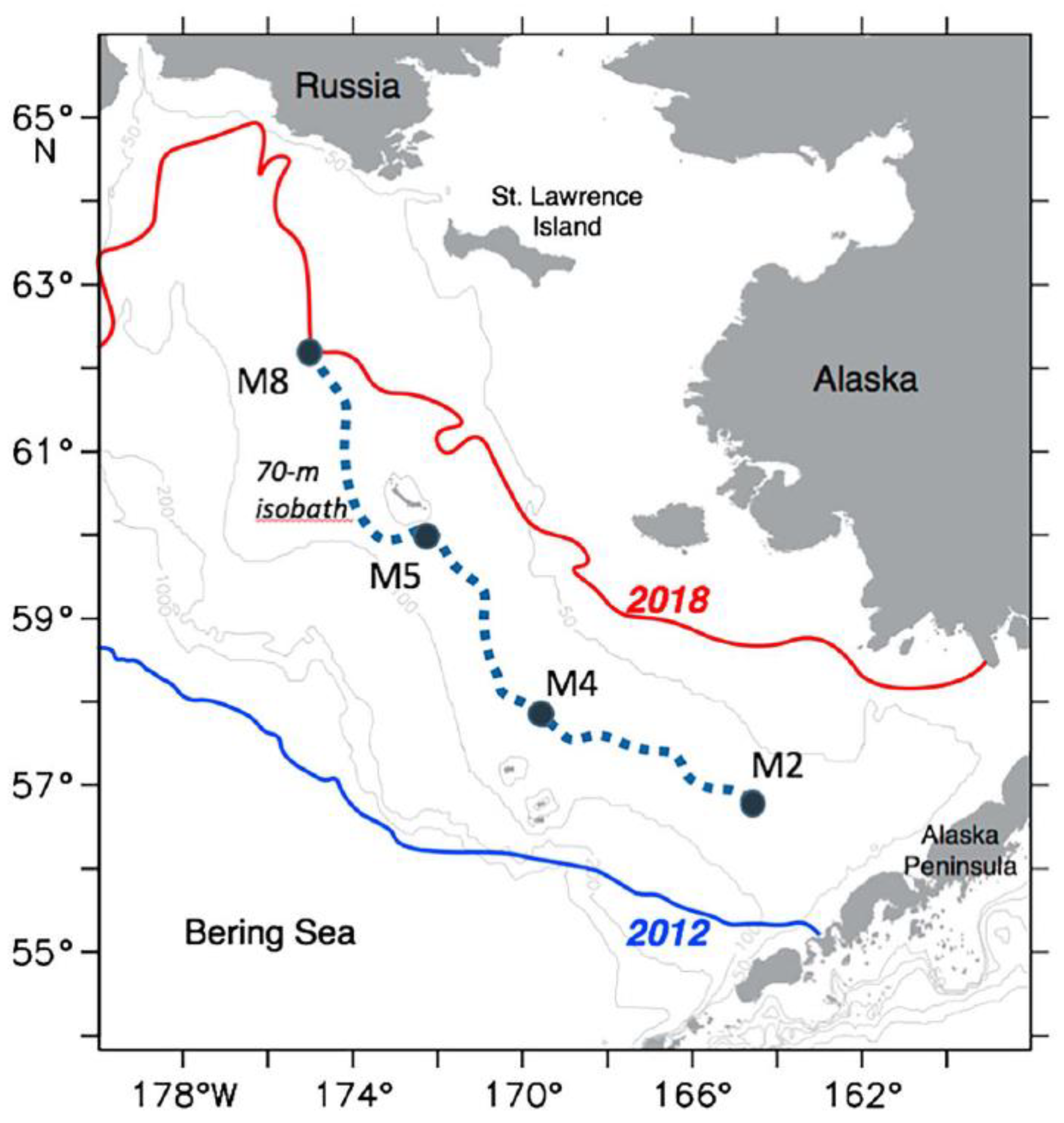
3.6. Unprecedented Loss of Seasonal Sea Ice
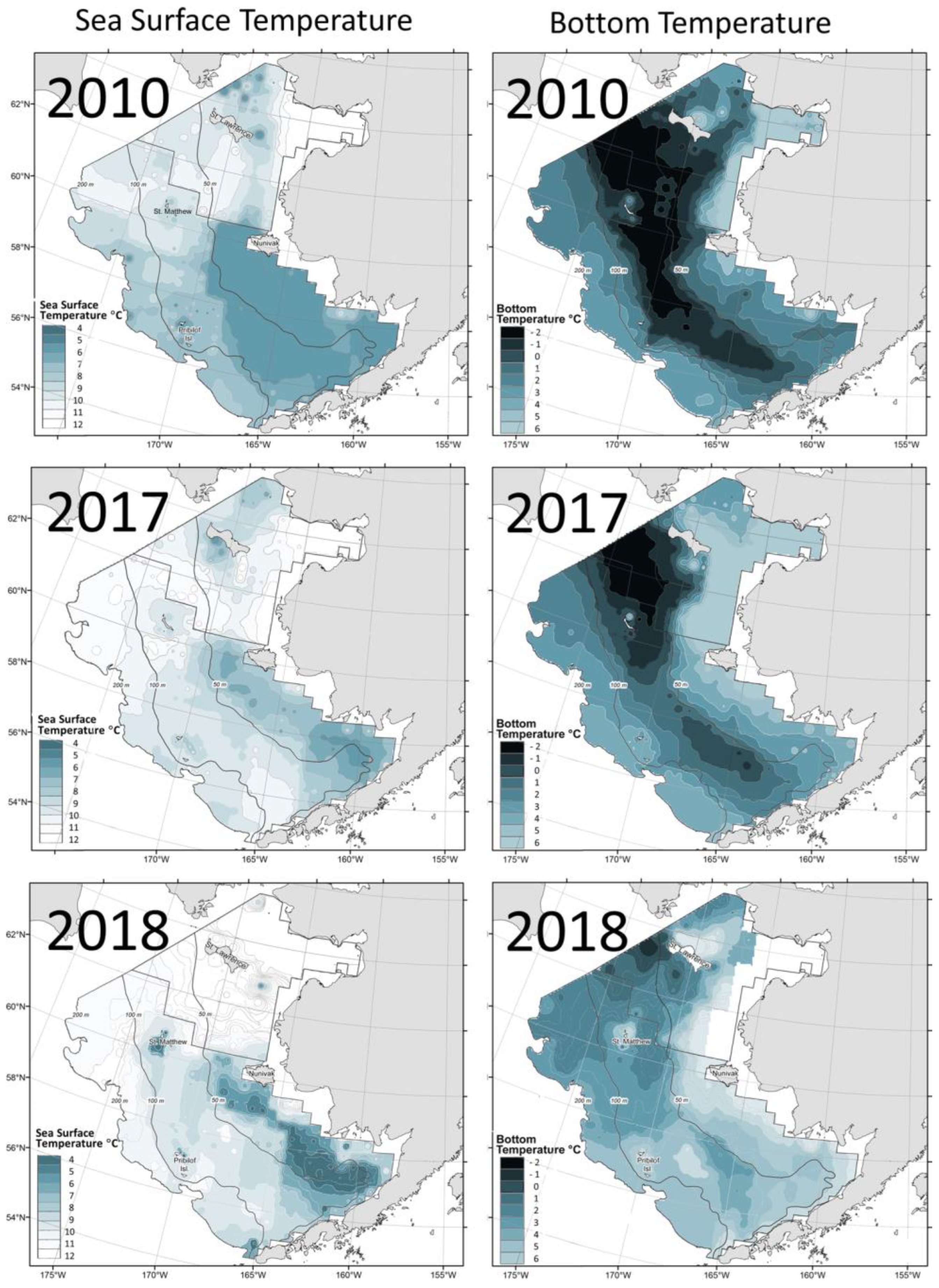
3.7. Disappearance of the Cold Pool on the East Bering Sea Shelf
3.8. Long-Term Variability of the Bering Shelf
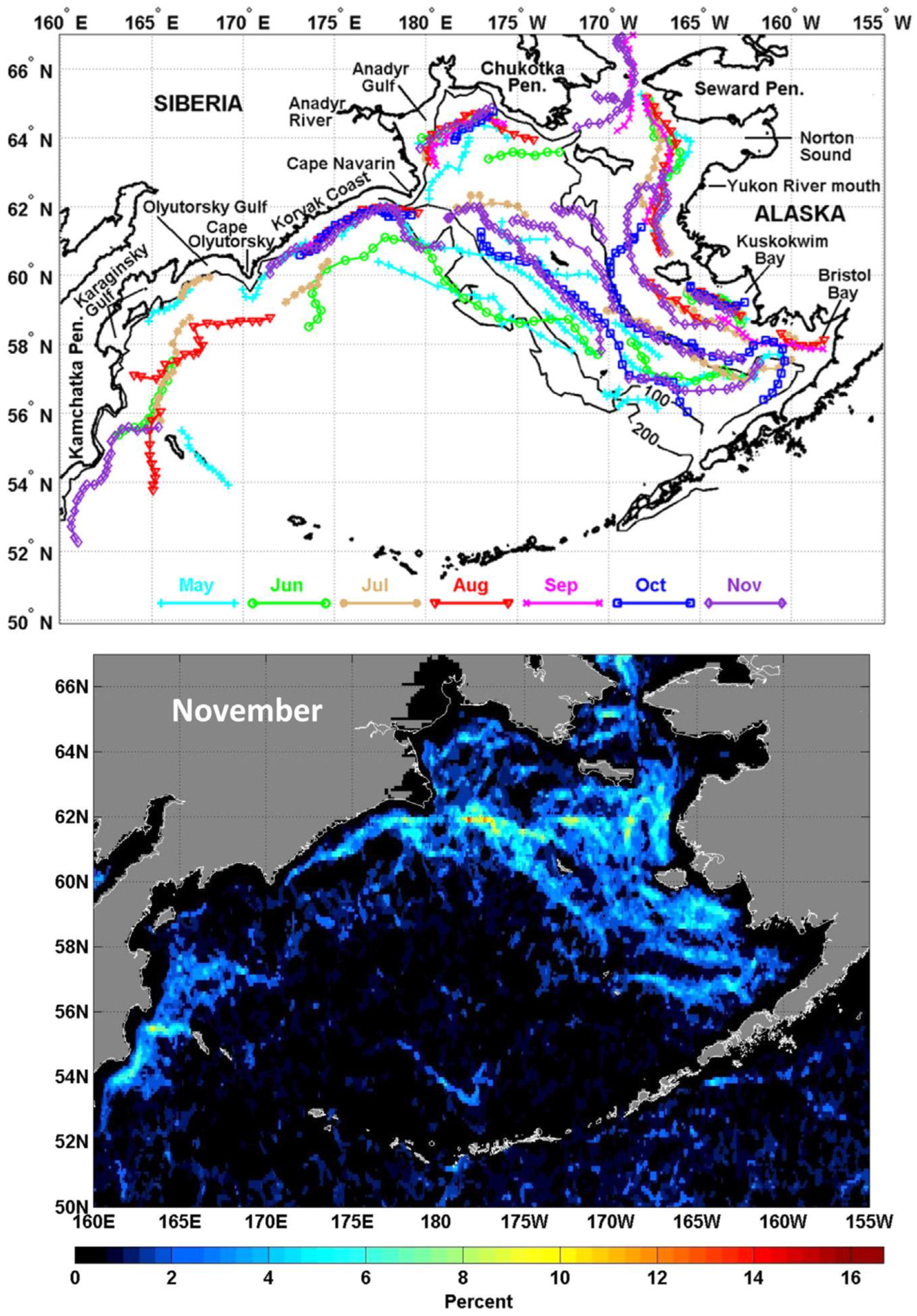
3.9. Physical Fronts
4. Ecosystem Responses to The Blob in the Eastern Bering Sea
4.1. Primary Productivity
4.2. Copepods and Euphausiids
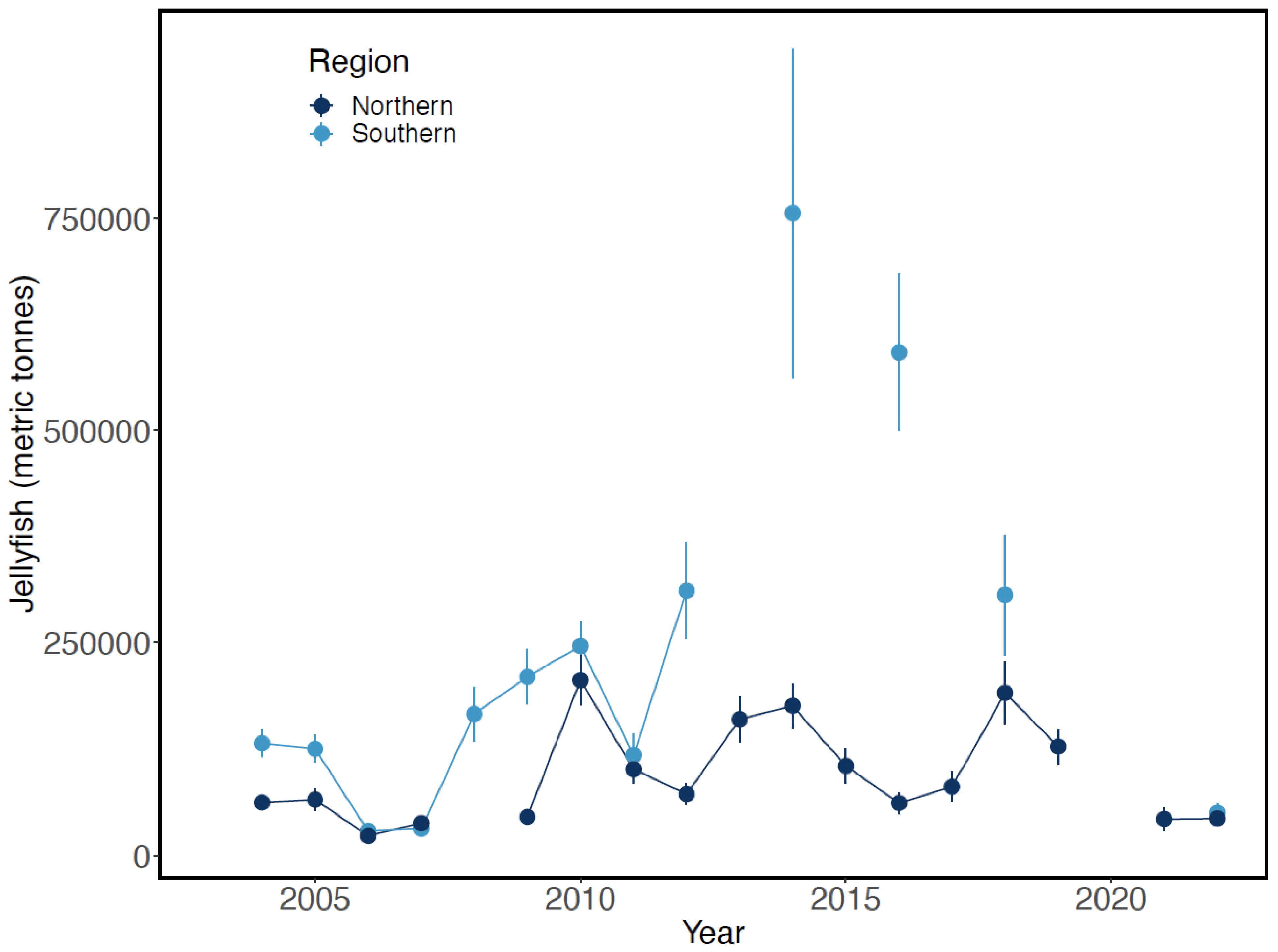
4.3. Jellyfish
4.4. Alaska Pollock and Pacific Cod
4.5. Pacific Halibut
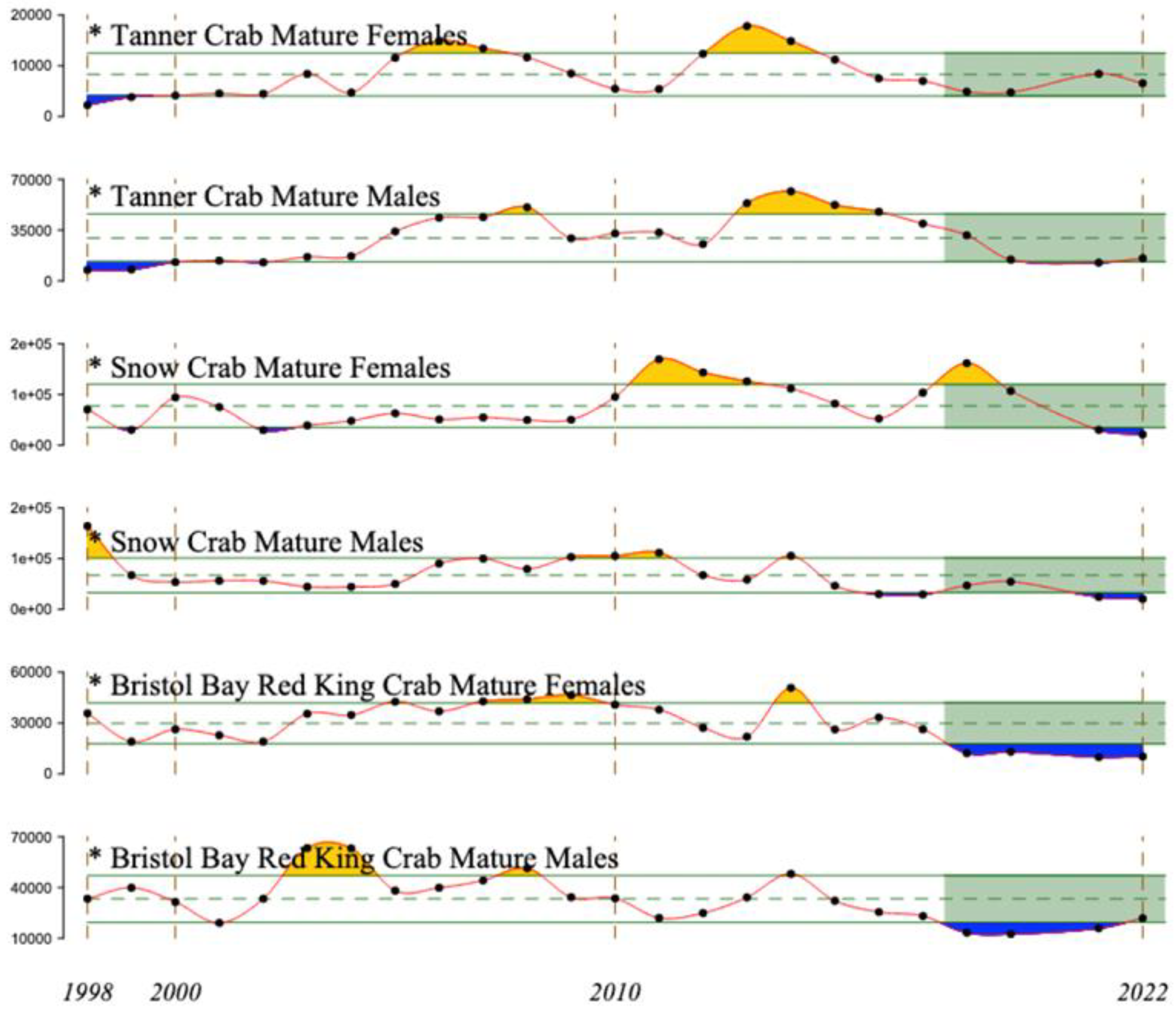
4.6. Red, Snow, and Tanner Crabs
4.7. Salmon
4.8. Seabirds
4.9. Range Extensions
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Freeland, H. Something odd in the Gulf of Alaska. CMOS Bull. SCMO 2014, 42, 57–59. [Google Scholar]
- Freeland, H. The “Blob” or Argo and other views of a large anomaly in the Gulf of Alaska in 2014/15. In State of the Physical, Biological and Selected Fishery Resources of Pacific in 2014; Canadian Technical Report of Fisheries and Aquatic Sciences; Chandler, P.C., King, S.A., Perry, R.I., Eds.; Fisheries & Oceans Canada Institute of Ocean Sciences: Sidney, BC, Canada, 2015; pp. 25–29. [Google Scholar]
- Freeland, H.; Ross, T. ‘The Blob’—Or, how unusual were ocean temperatures in the Northeast Pacific during 2014–2018? Deep. Sea Res. Part I 2019, 150, 103061. [Google Scholar] [CrossRef]
- Bond, N.A.; Cronin, M.F.; Freeland, H.; Mantua, N. Causes and impacts of the 2014 warm anomaly in the NE Pacific. Geophys. Res. Lett. 2015, 42, 3414–3420. [Google Scholar] [CrossRef]
- Scannell, H.A.; Johnson, G.C.; Thompson, L.; Lyman, J.M.; Riser, S.C. Subsurface evolution and persistence of marine heatwaves in the Northeast Pacific. Geophys. Res. Lett. 2020, 47, e2020GL090548. [Google Scholar] [CrossRef]
- Zhi, H.; Lin, P.F.; Zhang, R.H.; Chai, F.; Liu, H.L. Salinity effects on the 2014 warm “Blob” in the Northeast Pacific. Acta Oceanol. Sin. 2019, 38, 24–34. [Google Scholar] [CrossRef]
- Holser, R.R.; Keates, T.R.; Costa, D.P.; Edwards, C.A. Extent and magnitude of subsurface anomalies during the Northeast Pacific Blob as measured by animal-borne sensors. J. Geophys. Res. Ocean. 2022, 127, e2021JC018356. [Google Scholar] [CrossRef]
- Cavole, L.M.; Demko, A.M.; Diner, R.E.; Giddings, A.; Koester, I.; Pagniello, C.M.L.S.; Paulsen, M.L.; Ramirez-Valdez, A.; Schwenck, S.M.; Yen, N.K.; et al. Biological impacts of the 2013–2015 warm-water anomaly in the Northeast Pacific: Winners, losers, and the future. Oceanography 2016, 29, 273–285. [Google Scholar] [CrossRef]
- Suryan, R.M.; Arimitsu, M.L.; Coletti, H.A.; Hopcroft, R.R.; Lindeberg, M.R.; Barbeaux, S.J.; Batten, S.D.; Burt, W.J.; Bishop, M.A.; Bodkin, J.L.; et al. Ecosystem response persists after a prolonged marine heatwave. Sci. Rep. 2021, 11, 6235. [Google Scholar] [CrossRef]
- Schlitzer, R. Ocean Data View. 2022. Available online: https://odv.awi.de (accessed on 25 April 2023).
- Belkin, I.M. Comparative assessment of the West Bering Sea and East Bering Sea Large Marine Ecosystems. Environ. Dev. 2016, 17, 145–156. [Google Scholar] [CrossRef]
- Mueter, F.J.; Planque, B.; Hunt, G.L., Jr.; Alabia, I.D.; Hirawake, T.; Eisner, L.; Dalpadado, P.; Chierici, M.; Drinkwater, K.F.; Harada, N.; et al. Possible future scenarios in the gateways to the Arctic for Subarctic and Arctic marine systems: II. Prey resources, food webs, fish, and fisheries. ICES J. Mar. Sci. 2021, 78, 3017–3045. [Google Scholar] [CrossRef]
- Ladd, C.; Eisner, L.B.; Salo, S.A.; Mordy, C.W.; Iglesias-Rodriguez, M.D. Spatial and temporal variability of coccolithophore blooms in the eastern Bering Sea. J. Geophys. Res. Ocean. 2018, 123, 9119–9136. [Google Scholar] [CrossRef]
- Matson, P.G.; Washburn, L.; Fields, E.A.; Gotschalk, C.; Ladd, T.M.; Siegel, D.A.; Welch, Z.S.; Iglesias-Rodriguez, M.D. Formation, development, and propagation of a rare coastal coccolithophore bloom. J. Geophys. Res. Ocean. 2019, 124, 3298–3316. [Google Scholar] [CrossRef]
- Peterson, W.; Bond, N.; Robert, M. The blob (part three): Going, going, gone? PICES Press 2016, 24, 46–48. [Google Scholar]
- Zimmermann, M.; Prescott, M.M. Passes of the Aleutian Islands: First detailed description. Fish. Oceanogr. 2021, 30, 280–299. [Google Scholar] [CrossRef]
- Belkin, I.M.; Krishfield, R.; Honjo, S. Decadal variability of the North Pacific Polar Front: Subsurface warming versus surface cooling. Geophys. Res. Lett. 2002, 29, 65.1–65.4. [Google Scholar] [CrossRef]
- Belkin, I.M.; Shotwell, S.K. Advection of SST anomalies along the North Pacific Polar Front and their impact on the Gulf of Alaska, Aleutians, and Bering Sea ecosystems. In Proceedings of the Alaska Marine Science Symposium, Anchorage, AK, USA, 16–20 January 2012; p. 78, Alaska Marine Science Symposium Book of Abstracts, 2012. Available online: https://static1.squarespace.com/static/631a53939a3f0f445f25ea48/t/634f32a2155e8b13f868bba9/1666134698078/2012_AMSS_AbstractBook.pdf (accessed on 25 April 2023).
- Shotwell, S.K.; Hanselman, D.H.; Belkin, I.M. Toward biophysical synergy: Investigating advection along the Polar Front to identify factors influencing Alaska sablefish recruitment. Deep. Sea Res. Part II 2014, 107, 40–53. [Google Scholar] [CrossRef]
- Gentemann, C.L.; Fewings, M.R.; García-Reyes, M. Satellite sea surface temperatures along the West Coast of the United States during the 2014–2016 northeast Pacific marine heat wave. Geophys. Res. Lett. 2017, 44, 312–319. [Google Scholar] [CrossRef]
- von Biela, V.R.; Arimitsu, M.L.; Piatt, J.F.; Heflin, B.; Schoen, S.K.; Trowbridge, J.L.; Clawson, C.M. Extreme reduction in nutritional value of a key forage fish during the Pacific marine heatwave of 2014−2016. Mar. Ecol. Prog. Ser. 2019, 613, 171–182. [Google Scholar] [CrossRef]
- Walsh, J.E.; Thoman, R.L.; Bhatt, U.S.; Bieniek, P.A.; Brettschneider, B.; Brubaker, M.; Danielson, S.; Lader, R.; Fetterer, F.; Holderied, K.; et al. The high latitude marine heat wave of 2016 and its impacts on Alaska. Bull. Am. Meteorol. Soc. 2018, 99, S39–S43. [Google Scholar] [CrossRef]
- Stabeno, P.J.; Danielson, S.L.; Kachel, D.G.; Kachel, N.B.; Mordy, C.W. Currents and transport on the Eastern Bering Sea shelf: An integration of over 20 years of data. Deep. Sea Res. Part II 2016, 134, 13–29. [Google Scholar] [CrossRef]
- Thoman, R.L.; Bhatt, U.S.; Bieniek, P.A.; Brettschneider, B.R.; Brubaker, M.; Danielson, S.L.; Labe, Z.; Lader, R.; Meier, W.N.; Sheffield, G.; et al. The record low Bering Sea ice extent in 2018: Context, impacts, and an assessment of the role of anthropogenic climate change. Bull. Am. Meteorol. Soc. 2020, 101, S53–S58. [Google Scholar] [CrossRef]
- Duffy-Anderson, J.T.; Stabeno, P.; Andrews, A.; Cieciel, K.; Deary, A.; Farley, E.; Fugate, C.; Harpold, C.; Heintz, R.; Kimmel, D.; et al. Responses of the northern Bering Sea and southeastern Bering Sea pelagic ecosystems following record-breaking low winter sea ice. Geophys. Res. Lett. 2019, 46, 9833–9842. [Google Scholar] [CrossRef]
- Baker, M.R. Contrast of warm and cold phases in the Bering Sea to understand spatial distributions of Arctic and sub-Arctic gadids. Polar Biol. 2021, 44, 1083–1105. [Google Scholar] [CrossRef]
- Thorson, J.T. Measuring the impact of oceanographic indices on species distribution shifts: The spatially varying effect of cold-pool extent in the eastern Bering Sea. Limnol. Oceanogr. 2019, 64, 2632–2645. [Google Scholar] [CrossRef]
- Johnson, J.J.; Miksis-Olds, J.L.; Lippmann, T.C.; Jech, J.M.; Seger, K.D.; Pringle, J.M.; Linder, E. Decadal community structure shifts with cold pool variability in the eastern Bering Sea shelf. J. Acoust. Soc. Am. 2022, 152, 201–213. [Google Scholar] [CrossRef]
- Danielson, S.L.; Ahkinga, O.; Ashjian, C.; Basyuk, E.; Cooper, L.W.; Eisner, L.; Farley, E.; Iken, K.B.; Grebmeier, J.M.; Juranek, L.; et al. Manifestation and consequences of warming and altered heat fluxes over the Bering and Chukchi Sea continental shelves. Deep. Sea Res. Part II 2020, 177, 104781. [Google Scholar] [CrossRef]
- Stabeno, P.J.; Duffy-Anderson, J.T.; Eisner, L.B.; Farley, E.V.; Heintz, R.A.; Mordy, C.W. Return of warm conditions in the southeastern Bering Sea: Physics to fluorescence. PLoS ONE 2017, 12, e0185464. [Google Scholar] [CrossRef]
- Stabeno, P.J.; Bell, S.W. Extreme conditions in the Bering Sea (2017–2018): Record-breaking low sea-ice extent. Geophys. Res. Lett. 2019, 46, 8952–8959. [Google Scholar] [CrossRef]
- Belkin, I.M.; Cornillon, P.C. Bering Sea thermal fronts from Pathfinder data: Seasonal and interannual variability. Pac. Oceanogr. 2005, 3, 6–20. [Google Scholar]
- Hunt, G.L., Jr.; Stabeno, P.J.; Strom, S.; Napp, J.M. Patterns of spatial and temporal variation in the marine ecosystem of the southeastern Bering Sea, with special reference to the Pribilof Domain. Deep. Sea Res. Part II 2008, 55, 1919–1944. [Google Scholar] [CrossRef]
- Siddon, E. (Ed.) Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022. [Google Scholar]
- Nielsen, J.M.; Eisner, L.; Watson, J.; Gann, J.C.; Callahan, M.W.; Mordy, C.W.; Bell, S.W.; Stabeno, P. Spring satellite chlorophyll-a concentrations in the eastern Bering Sea. In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 68–72. [Google Scholar]
- Springer, A.M.; McRoy, C.P.; Flint, M.V. The Bering Sea Green Belt: Shelf-edge processes and ecosystem production. Fish. Oceanogr. 1996, 5, 205–223. [Google Scholar] [CrossRef]
- Kimmel, D.; Barrett, J.; Cooper, D.; Crouser, D.; Deary, A.; Eisner, L.; Lamb, J.; Murphy, J.; Pinger, C.; Cormack, B.; et al. Current and historical trends for zooplankton in the Bering Sea. In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 79–90. [Google Scholar]
- Yasumiishi, E.; Andrews, A.; Murphy, J.; Dimond, A.; Farley, E.; Siddon, E. Trends in the biomass of jellyfish in the southeastern and northeastern Bering Sea during the late-summer surface trawl survey, 2003–2022. In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 94–95. [Google Scholar]
- Ianelli, J.; Fissel, B.; Stienessen, S.; Honkalehto, T.; Siddon, E.; Allen-Akselrud, C. Chapter 1: Assessment of the Walleye Pollock Stock in the Eastern Bering Sea. In Stock Assessment and Fishery Evaluation Report for the Groundfish Resources of the Bering Sea/Aleutian Islands Regions; Alaska Fisheries Science Center, National Marine Fisheries Service, National Oceanic and Atmospheric Administration: Seattle, WA, USA, 2021; p. 171. [Google Scholar]
- Thompson, G.G.; Barbeaux, S.; Conner, J.; Fissel, B.; Hurst, T.; Laurel, B.; O’Leary, C.A.; Rogers, L.; Shotwell, S.K.; Siddon, E.; et al. Assessment of the Pacific Cod Stock in the Eastern Bering Sea. In NPFMC Bering Sea and Aleutian Islands SAFE; North Pacific Fishery Management Council: Anchorage, AK, USA, 2021; p. 494. [Google Scholar]
- Richar, J. Eastern Bering Sea commercial crab stock biomass. In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 140–141. [Google Scholar]
- Murphy, J.; Garcia, S.; Cooper, D.; Farley, E.; Lee, E.; Dimond, A.; Howard, K. Northern Bering Sea juvenile salmon abundance indices. In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 106–108. [Google Scholar]
- Andrews, A.; Yasumiishi, E.; Farley, E.; Murphy, J.; Dimond, A. Trends in the abundance of juvenile sockeye salmon in the southeastern Bering Sea during the late-summer surface trawl survey, 2003–2020. In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 105–106. [Google Scholar]
- Akiya, A.; Ahkinga, S.; Divine, L.; Jones, T.; Kingeekuk, L.; Lestenkof, A.; Lindsey, J.; Niksik, T.; Padula, V.; Pungowiyi, P.; et al. Integrated seabird information. In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 142–148. [Google Scholar]
- Landeira, J.M.; Matsuno, K.; Tanaka, Y.; Yamaguchi, A. First record of the larvae of tanner crab Chionoecetes bairdi in the Chukchi Sea: A future northward expansion in the Arctic? Polar Sci. 2018, 16, 86–89. [Google Scholar] [CrossRef]
- Thorson, J.T.; Fossheim, M.; Mueter, F.J.; Olsen, E.; Lauth, R.R.; Primicerio, R.; Husson, B.; Marsh, J.; Dolgov, A.; Zador, S.G. Comparison of near-bottom fish densities show rapid community and population shifts in Bering and Barents seas. In Arctic Report Card; Richter-Menge, J., Druckenmiller, M.L., Jeffries, M., Eds.; NOAA: Washington, DC, USA, 2019; pp. 72–80. [Google Scholar]
- Levine, R.M.; De Robertis, A.; Grünbaum, D.; Wildes, S.; Farley, E.V.; Stabeno, P.J.; Wilson, C.D. Climate-driven shifts in pelagic fish distributions in a rapidly changing Pacific Arctic. Deep. Sea Res. Part II 2023, 208, 105244. [Google Scholar] [CrossRef]
- Eddy, T.D.; Bernhardt, J.R.; Blanchard, J.L.; Cheung, W.W.L.; Colléter, M.; du Pontavice, H.; Fulton, E.A.; Gascuel, D.; Kearney, K.A.; Petrik, C.M.; et al. Energy flow through marine ecosystems: Confronting transfer efficiency. Trends Ecol. Evol. 2021, 36, 76–86. [Google Scholar] [CrossRef]
- Cunningham, C.J.; Vega, S.; Head, J. Temporal trend in the annual inshore run size of Bristol Bay sockeye salmon (Oncorhynchus nerka). In Ecosystem Status Report 2022: Eastern Bering Sea, Stock Assessment and Fishery Evaluation Report; Elizabeth, S., Ed.; North Pacific Fishery Management Council: Anchorage, AK, USA, 2022; pp. 109–110. [Google Scholar]
- Yasumiishi, E.; Cunningham, C.; Farley, K.C.; Moss, J.; Strasburger, W.; Eisner, L.; Andrews, A.; Gann, J.; Murphy, J.; Dimond, A.; et al. Mechanisms for shifts in the distribution and abundance of juvenile sockeye salmon in the Eastern Bering Sea during late summer, 2002–2018. In North Pacific Anadromous Fish Commission Technical Report No. 15; North Pacific Fishery Management Council: Anchorage, AK, USA, 2019; pp. 129–131. [Google Scholar]
- Yasumiishi, E.M.; Cieciel, K.; Andrews, A.G.; Murphy, J.; Dimond, J.A. Climate-related changes in the biomass and distribution of small pelagic fishes in the eastern Bering Sea during late summer, 2002–2018. Deep. Sea Res. Part II 2020, 181–182, 104907. [Google Scholar] [CrossRef]
- Natsuike, M.; Saito, R.; Fujiwara, A.; Matsuno, K.; Yamaguchi, A.; Shiga, N.; Hirawake, T.; Kikuchi, T.; Nishino, S.; Imai, I. Evidence of increased toxic Alexandrium tamarense dinoflagellate blooms in the eastern Bering Sea in the summers of 2004 and 2005. PLoS ONE 2017, 12, e0188565. [Google Scholar] [CrossRef]
- Sherman, K.; O’Reilly, J.; Belkin, I.M.; Melrose, C.; Friedland, K.D. The application of satellite remote sensing for assessing productivity in relation to fisheries yields of the world’s large marine ecosystems. ICES J. Mar. Sci. 2011, 68, 667–676. [Google Scholar] [CrossRef]
- Sherman, K.; Belkin, I.M.; Friedland, K.D.; O’Reilly, J. Changing states of North Atlantic large marine ecosystems. Environ. Dev. 2013, 7, 46–58. [Google Scholar] [CrossRef]























Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belkin, I.M.; Short, J.W. Echoes of the 2013–2015 Marine Heat Wave in the Eastern Bering Sea and Consequent Biological Responses. J. Mar. Sci. Eng. 2023, 11, 958. https://doi.org/10.3390/jmse11050958
Belkin IM, Short JW. Echoes of the 2013–2015 Marine Heat Wave in the Eastern Bering Sea and Consequent Biological Responses. Journal of Marine Science and Engineering. 2023; 11(5):958. https://doi.org/10.3390/jmse11050958
Chicago/Turabian StyleBelkin, Igor M., and Jeffrey W. Short. 2023. "Echoes of the 2013–2015 Marine Heat Wave in the Eastern Bering Sea and Consequent Biological Responses" Journal of Marine Science and Engineering 11, no. 5: 958. https://doi.org/10.3390/jmse11050958
APA StyleBelkin, I. M., & Short, J. W. (2023). Echoes of the 2013–2015 Marine Heat Wave in the Eastern Bering Sea and Consequent Biological Responses. Journal of Marine Science and Engineering, 11(5), 958. https://doi.org/10.3390/jmse11050958