
Effects of Global Warming on the Growth and Proliferation of Attached Sargassum horneri in the Aquaculture Area near Gouqi Island, China


 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of S. horneri Samples
2.2. Monitoring of the Growth State of Attached S. horneri in the Aquaculture Area
2.3. Monitoring SST in the Area near Gouqi Island
2.4. Calculation of the Effective Accumulated Temperature
2.5. Data Processing and Analysis
3. Results
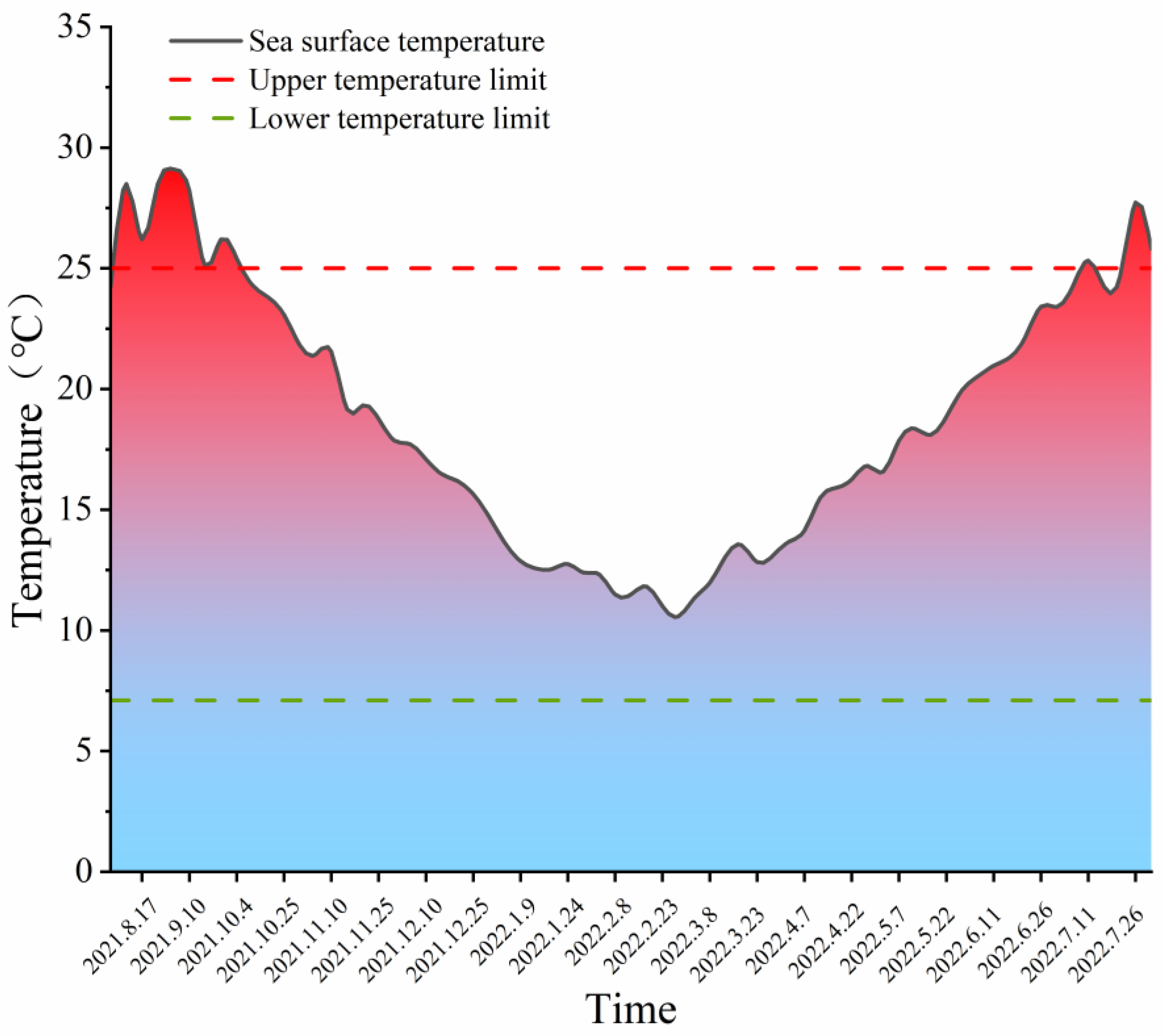
3.1. Variation in SST near Gouqi Island in 2021–2022 and in the Previous 5 Years
3.2. Morphological Characteristics of Attached S. horneri across Four Seasons between 2021 and 2022
3.3. Variation in the Monthly Photosynthetic Activity of Attached S. horneri from 2021 to 2022
3.4. Relationship between Growth Period and Effective Accumulated Temperature in Attached S. horneri
4. Discussion and Conclusions
4.1. Temperature Adaptability of Attached S. horneri in the Mussel Culture Area near Gouqi Island
4.2. Analysis of the Effect of Global Warming on the Growth and Reproduction of Attached S. horneri near Gouqi Island Based on the Effective Accumulated Temperature Theory
4.3. Golden Tides and the Early Warning Problem
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tseng, C.E.U. Common Seaweeds of China; Science Press: Beijing, China, 1983; p. 316. [Google Scholar]
- Zeng, C.K.; Lu, B.R. Records of China Seaweed (Vol. 3 Phaeophyta Vol. 2 Chaetoceros); Science Press: Beijing, China, 2000; pp. 43–44. [Google Scholar]
- Sun, J.Z.; Chen, W.D.; Zhuang, D.G.; Zheng, H.Y.; Lin, L.; Pang, S.J. In situ ecological studies of the subtidal brown alga Sargasssum horneri at Nanji Island of China. South China Fish. Sci. 2008, 3, 58–63. [Google Scholar] [CrossRef]
- Sun, J.Z.; Zhuang, D.G.; Yang, J.B.; Chen, W.D.; Wang, T.G.; Pang, S.J. Primary study on enhancement technique of Sargassum horneri around Nanji Island. Mod. Fish. Inf. 2010, 25, 23–27. [Google Scholar] [CrossRef]
- Sun, J.Z.; Zhuang, D.G.; Chen, W.D.; Zheng, H.Y.; Lin, L.; Pang, S.J. Studies on sexual reproduction and seedling production of the brown alga Sargassum horneri. South China Fish. Sci. 2008, 2, 6–14. [Google Scholar] [CrossRef]
- Li, D.P.; Li, H.; Zhang, G.J.; Ma, Z.L.; Xin, M.L.; Ding, G.; Wu, H.Y.; Guo, W. Studies on the reproduction and development of Sargassum horneri. Mar. Sci. 2018, 42, 40–45. [Google Scholar] [CrossRef]
- Qi, L.; Hu, C.; Wang, M.; Shang, S.; Wilson, C. Floating Algae Blooms in the East China Sea. Geophys. Res. Lett. 2017, 44, 11501–11509. [Google Scholar] [CrossRef]
- Zhang, P.; Cai, Y.F.; Wang, T.G.; Zhong, C.H.; Xie, Q.L.; Chen, S.B.; Wang, N. AFLP analysis of different geographic populations of Sargassum horneri along the coast of Zhejiang Province. Acta Agric. Zhejiangensis 2015, 27, 1586–1592. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Q.; Chen, J.J.; Zhu, Q.; Wu, X.K.; Luo, Q.J.; Yang, R.; Chen, H.M. Analysis of pigments in floating and fixed Sargassum horneri by high performance liquid chromatography coupled with triple quadrupole tandem mass spectrometry. J. Nucl. Agric. Sci. 2019, 33, 1173–1180. [Google Scholar] [CrossRef]
- Ding, X.W.; Zhang, J.H.; Zhuang, M.M.; Kang, X.Y.; Zhao, X.H.; He, P.M.; Liu, S.R.; Liu, J.F.; Wen, Y.; Shen, H.; et al. Growth of Sargassum horneri distribution properties of golden tides in the Yangtze Estuary and adjacent waters. Mar. Fish. 2019, 41, 188–196. [Google Scholar] [CrossRef]
- Wu, Z.L.; Cheng, L.R.; Wang, K.; Zhang, S.Y.; Bi, Y.X. Morphological characteristics of vesicle of Sargassum horneri and its relationship to environmental factors in Gouqi Island. J. Fish. China 2020, 44, 793–804. [Google Scholar] [CrossRef]
- Zhang, J.H.; Ding, X.W.; Zhuang, M.M.; Wang, S.Y.; Chen, L.; Shen, H.; He, P.M. An increase in new Sargassum (Phaeophyceae) blooms along the coast of the East China Sea and Yellow Sea. Phycologia 2019, 58, 374–381. [Google Scholar] [CrossRef]
- Xing, Q.; Guo, R.; Wu, L.; An, D.; Cong, M.; Qin, S.; Li, X. High-Resolution Satellite Observations of a New Hazard of Golden Tides Caused by Floating Sargassum in Winter in the Yellow Sea. IEEE Geosci. Remote Sens. Lett. 2017, 14, 1815–1819. [Google Scholar] [CrossRef]
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef]
- State Oceanic Administration. Bulletin of China Marine Ecological Environment Status. 2017; pp. 55–56. Available online: http://gc.mnr.gov.cn/201806/t20180619_1797652.html. (accessed on 1 July 2022).
- Hiraoka, M.; Masao, O.; Shigeo, K.; Goro, Y. Crossing test among floating Ulva thalli forming ′green tide′ in Japan. Hydrobiologia 2004, 512, 239–245. [Google Scholar] [CrossRef]
- Abdelrhman, A.M.; Mohamed, A.; Mohamed, A.A.; Zaka, Z.S.; Hany, N.; Mohamed, A.A.Z.; Norhan, H.A.; Sherine, R.A.; Ehab, E.; Hien, V.D.; et al. Effect of polysaccharides derived from brown macroalgae Sargassum dentifolium on growth performance, serum biochemical, digestive histology and enzyme activity of hybrid red tilapia. Aquac. Rep. 2022, 25, 101212. [Google Scholar] [CrossRef]
- Abdallah, T.M.; Ahmed, E.A.; Khamael, M.A.; Hossam, S.E.; Khaled, M.A.R.; Mohamed, A. Dried Brown Seaweed’s Phytoremediation Potential for Methylene Blue Dye Removal from Aquatic Environments. Polymers 2022, 14, 1375. [Google Scholar] [CrossRef]
- Goro, Y.; Koji, Y.; Toshinobu, T. Growth and maturation of two populations of Sargassum horneri (Fucales, Phaeophyta) in Hiroshima Bay, the Seto Inland Sea. Fish. Sci. 2001, 67, 1023–1029. [Google Scholar] [CrossRef]
- Ogawa, H. The maturation and early development of Sargassaceous plants and the effects of environment to them. Gekkan Kaiyo-Kagaku 1985, 17, 26–31. [Google Scholar]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef]
- Meehl, G.A.; Stocker, T.F.; Collins, W.D.; Friedlingstein, P.; Gaye, A.T.; Gregory, J.M.; Kitoh, A.; Knutti, R.; Murphy, J.M.; Noda, A.; et al. Global climate projections. In Climate Change 2007: The Physical Science Basis: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 749–846. [Google Scholar]
- Ji, Y.; Xu, Z.; Zou, D.; Gao, K. Ecophysiological responses of marine macroalgae to climate change factors. J. Appl. Phycol. 2016, 28, 2953–2967. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; Gouezo, M.; Tilbrook, B.; Dove, S.; Anthony, K.R.N. High CO2 enhances the competitive strength of seaweeds over corals. Ecol. Lett. 2011, 14, 156–162. [Google Scholar] [CrossRef]
- Wernberg, T.; Bennett, S.; Babcock, R.; De Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.; Hovey, R.; et al. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Levac, E.; Guelphen, L.; Pohle, G.; Chmura, G. The effect of global climate change on the future distribution of economically important macroalgae (seaweeds) in the Northwest Atlantic. Facets 2018, 3, 275–286. [Google Scholar] [CrossRef]
- Zhang, J. Indoor Cultivation of the Brown Alga Sargassum Vachellianum and Sargassum horneri: Morphological Observation and Techniques of Artificial Seeding. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2012; pp. 50–51. [Google Scholar]
- Wang, Y.; Zhong, Z.H.; Qin, S.; Li, J.L.; Li, J.J.; Liu, Z.Y. Effects of temperature and light on growth rate and photosynthetic characteristics of Sargassum horneri. J. Ocean Univ. China 2021, 20, 101–110. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Liu, H.S.; Zhang, J.W. Research on the benthic algae distribution around Gouqi Island using digital echo sounding system and interpolation. J. Shanghai Ocean. Univ. 2012, 21, 445–451. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Zhao, S.J. Marine Algae Ecology; Ocean Press: Beijing, China, 2014; p. 164. [Google Scholar]
- Wu, Z.L.; Zhang, S.Y. Effect of typhoon on the distribution of macroalgae in the seaweed beds of Gouqi Island, Zhejiang Province. J. Agric. Sci. Technol. 2019, 21, 159–168. [Google Scholar] [CrossRef]
- Li, X.M.; Zhang, S.Y.; Wang, K.; Chen, L.R.; Chen, T.H. Effect of annual temperature variation on growth of Sargassum horneri in Gouqi Island. Oceanol. Limnol. Sin. 2020, 51, 1136–1143. [Google Scholar]
- Choi, H.G.; Lee, K.H.; Yoo, H.I.; Kang, P.J.; Kim, Y.S.; Nam, K.W. Physiological differences in the growth of Sargassum horneri between the germling and adult stages. J. Appl. Phycol. 2008, 5, 29–735. [Google Scholar] [CrossRef]
- Okuda, T. Monoecism and autumn-fruiting of S. horneri. Jpn. J. Phycol. 1987, 37, 279–283. [Google Scholar]
- Okuda, T.; Satoh, Y. Conceptacular development in Sargassum filicinum and autumnal S. horneri (Phaeophyceae). Jpn. J. Phycol. 1989, 37, 279–283. [Google Scholar]
- Uchida, T.; Arima, S. Crossing experiments between autumn and spring fruiting types of Sargassum horneri (Phaeophyta). Nippon. Suisan Gakkaishi 1993, 59, 1685–1688. [Google Scholar] [CrossRef][Green Version]
- Chen, L.R.; Zhang, S.Y.; Chen, Y.; Zhao, X.; Zhou, X.J.; Chen., Y.Z. Life history and morphology of Sargassum horneri from the Sargassum seaweed bed of Gouqi Island. J. Fish. China 2015, 39, 1218–1229. [Google Scholar]
- McCourt, R.M. Seasonal patterns of abundance, distributions, and phenology in relation to growth strategies of three Sargassum species. J. Exp. Mar. Biol. Ecol. 1984, 74, 141–156. [Google Scholar] [CrossRef]
- Hurtado, A.Q.; Ragaza, A.R. Sargassum studies in Currimao, Ilocos Norte, Northern Philippines I. Seasonal variations in the biomass of Sargassum carpophyllum J. Agardh, Sargassum ilicifolium (Turner) C. Agardh and Sargassum siliquosum J. Agardh(Phaeophyta, Sargassaceae). Bot. Marina 1999, 42, 321–325. [Google Scholar] [CrossRef]
- Díaz-Villa, T.; Sansón, M.; Afonso-Carrillo, J. Seasonal variations in growth and reproduction of Sargassum orotavicum (Fucales, Phaeophyceae) from the Canary Islands. Bot. Marina 2005, 48, 18–29. [Google Scholar] [CrossRef]
- Rivera, M.; Scrosati, R. Population dynamics of Sargassum lapazeanum (Fucales, Phaeophyta) from the Gulf of California, Mexico. Phycologia 2006, 45, 178–189. [Google Scholar] [CrossRef]
- Taylor, W.R. Marine Algae of the Eastern Tropical and Subtropical Coasts of the Americas; University of Michigan Press: Ann Arbor, MI, USA, 1960; p. 167. [Google Scholar]
- Deysher, L.E. Reproductive phenology of newly introduced populations of the brown alga, Sargassum muticum (Yendo) Fensholt. Hydrobiologia 1984, 116–117, 403–407. [Google Scholar] [CrossRef]
- Ang, P.O. Phenology of Sargassum spp. in Tung Ping Chau Marine Park, Hong Kong SAR, China. J. Appl. Phycol. 2006, 18, 629–636. [Google Scholar] [CrossRef]
- Komatsu, T.; Fukuda, M.; Mikami, A.; Mizuno, S.; Kantachumpoo, A.; Tanoue, H.; Kawamiya, M. Possible change in distribution of seaweed, Sargassum horneri, in northeast Asia under A2 scenario of global warming and consequent effect on some fish. Mar. Pollut. Bull. 2014, 85, 317–324. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.; Xia, L.; Zhuang, M.; Pan, J.; Liu, J.; Dai, W.; Zhao, Z.; Zhang, M.; Shen, X.; He, P.; et al. Effects of Global Warming on the Growth and Proliferation of Attached Sargassum horneri in the Aquaculture Area near Gouqi Island, China. J. Mar. Sci. Eng. 2023, 11, 9. https://doi.org/10.3390/jmse11010009
Wu T, Xia L, Zhuang M, Pan J, Liu J, Dai W, Zhao Z, Zhang M, Shen X, He P, et al. Effects of Global Warming on the Growth and Proliferation of Attached Sargassum horneri in the Aquaculture Area near Gouqi Island, China. Journal of Marine Science and Engineering. 2023; 11(1):9. https://doi.org/10.3390/jmse11010009
Chicago/Turabian StyleWu, Tingjian, Lihua Xia, Minmin Zhuang, Jiayu Pan, Jinlin Liu, Wei Dai, Zitao Zhao, Meijing Zhang, Xiyu Shen, Peimin He, and et al. 2023. "Effects of Global Warming on the Growth and Proliferation of Attached Sargassum horneri in the Aquaculture Area near Gouqi Island, China" Journal of Marine Science and Engineering 11, no. 1: 9. https://doi.org/10.3390/jmse11010009
APA StyleWu, T., Xia, L., Zhuang, M., Pan, J., Liu, J., Dai, W., Zhao, Z., Zhang, M., Shen, X., He, P., Zhang, J., & Qin, Y. (2023). Effects of Global Warming on the Growth and Proliferation of Attached Sargassum horneri in the Aquaculture Area near Gouqi Island, China. Journal of Marine Science and Engineering, 11(1), 9. https://doi.org/10.3390/jmse11010009