
From Plates to Baits: Using a Remote Video Foraging System to Study the Impact of Foraging on Fouling Non-Indigenous Species

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
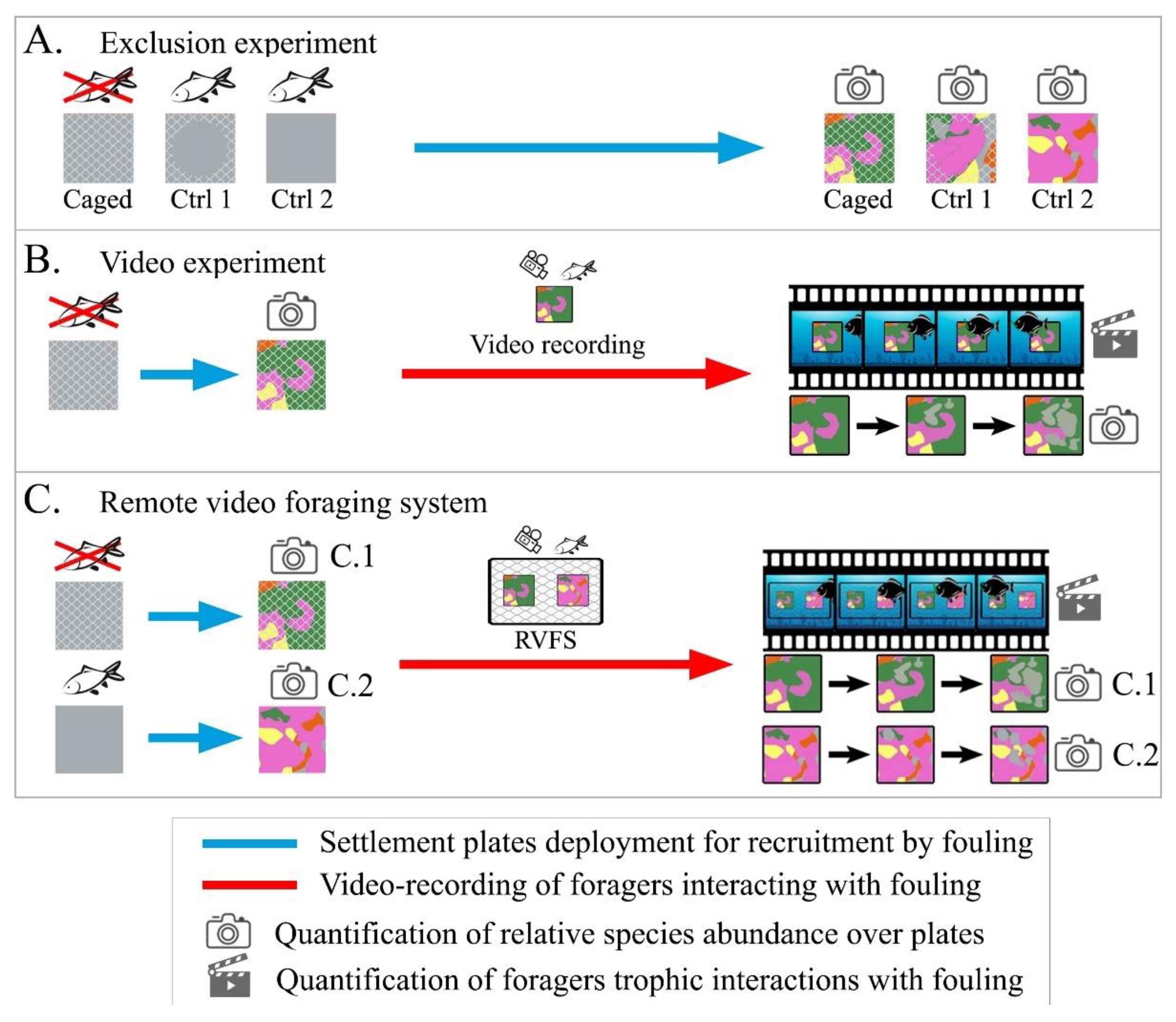
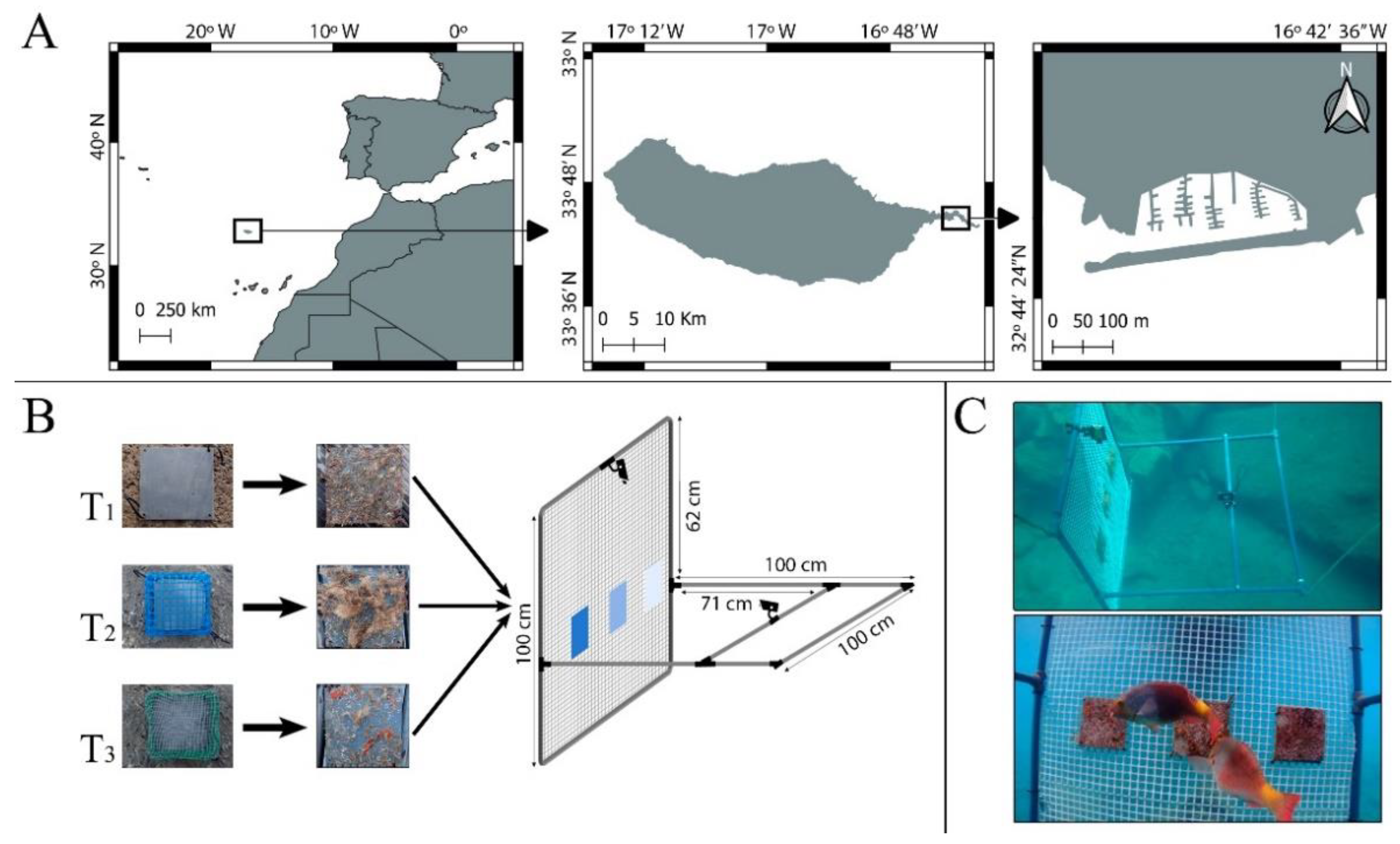
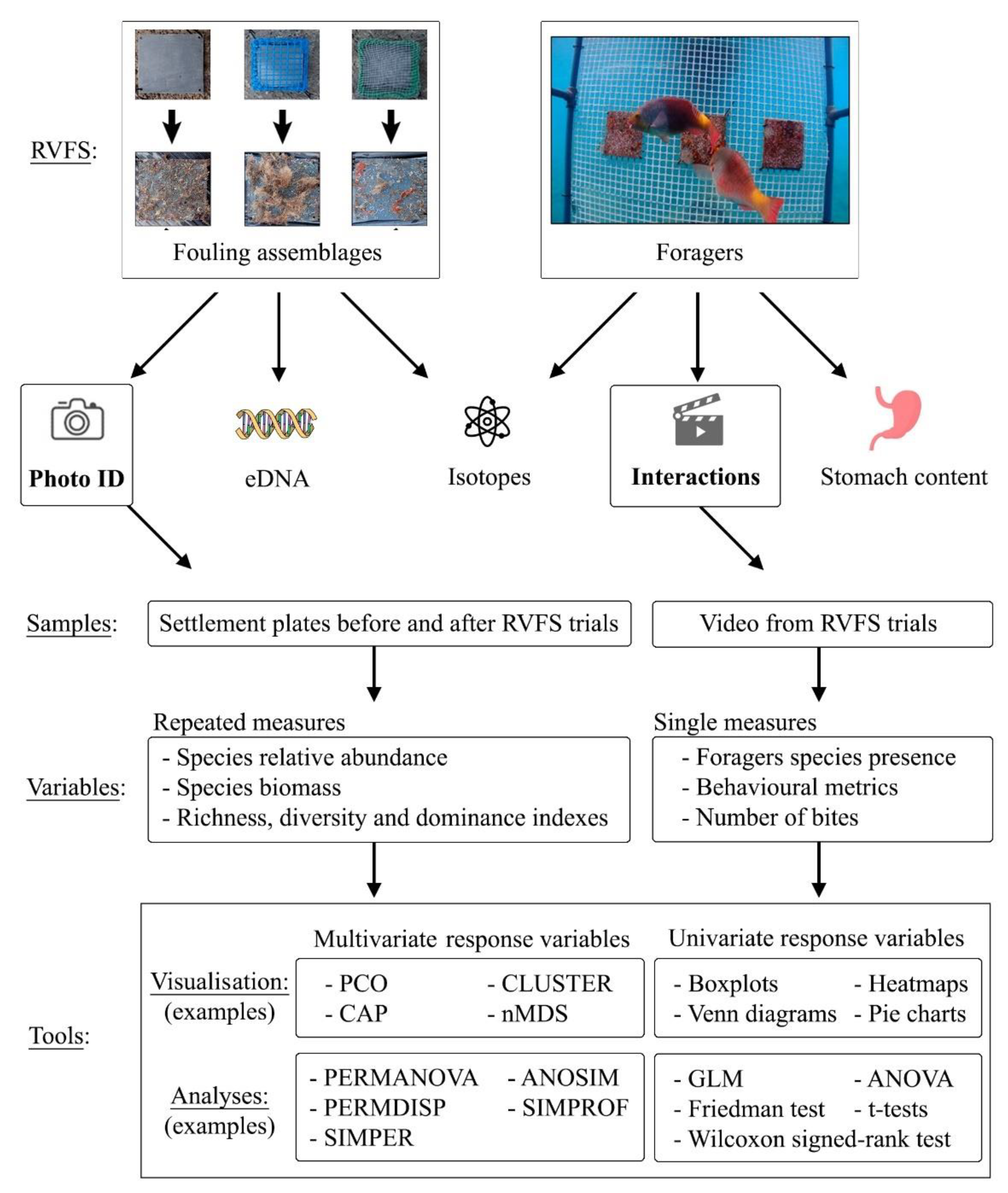
2.1. Remote Video Foraging System
2.2. Pilot Study
2.2.1. Experimental Setup
2.2.2. Data Acquisition and Treatment
2.2.3. Statistical Analyses
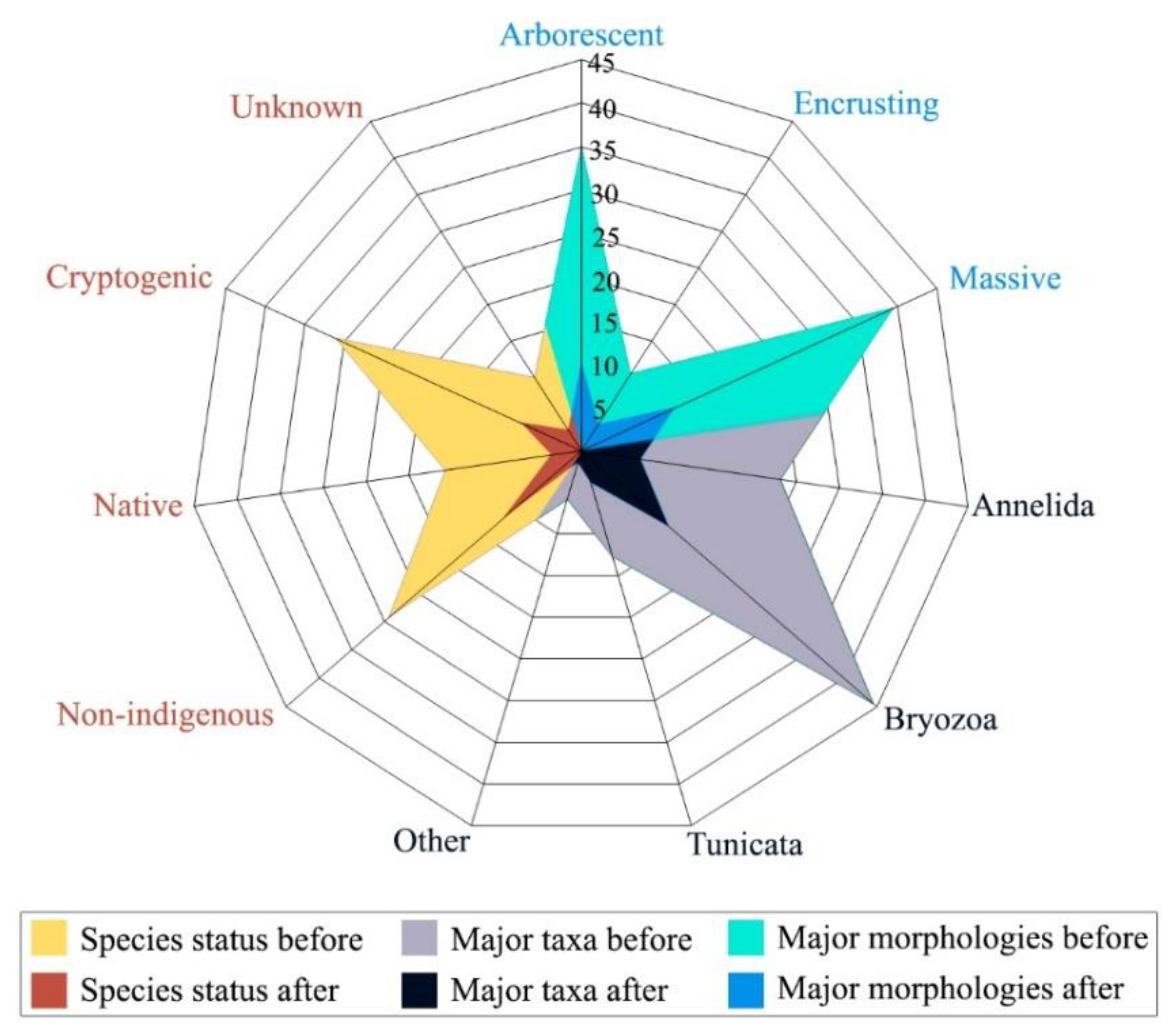
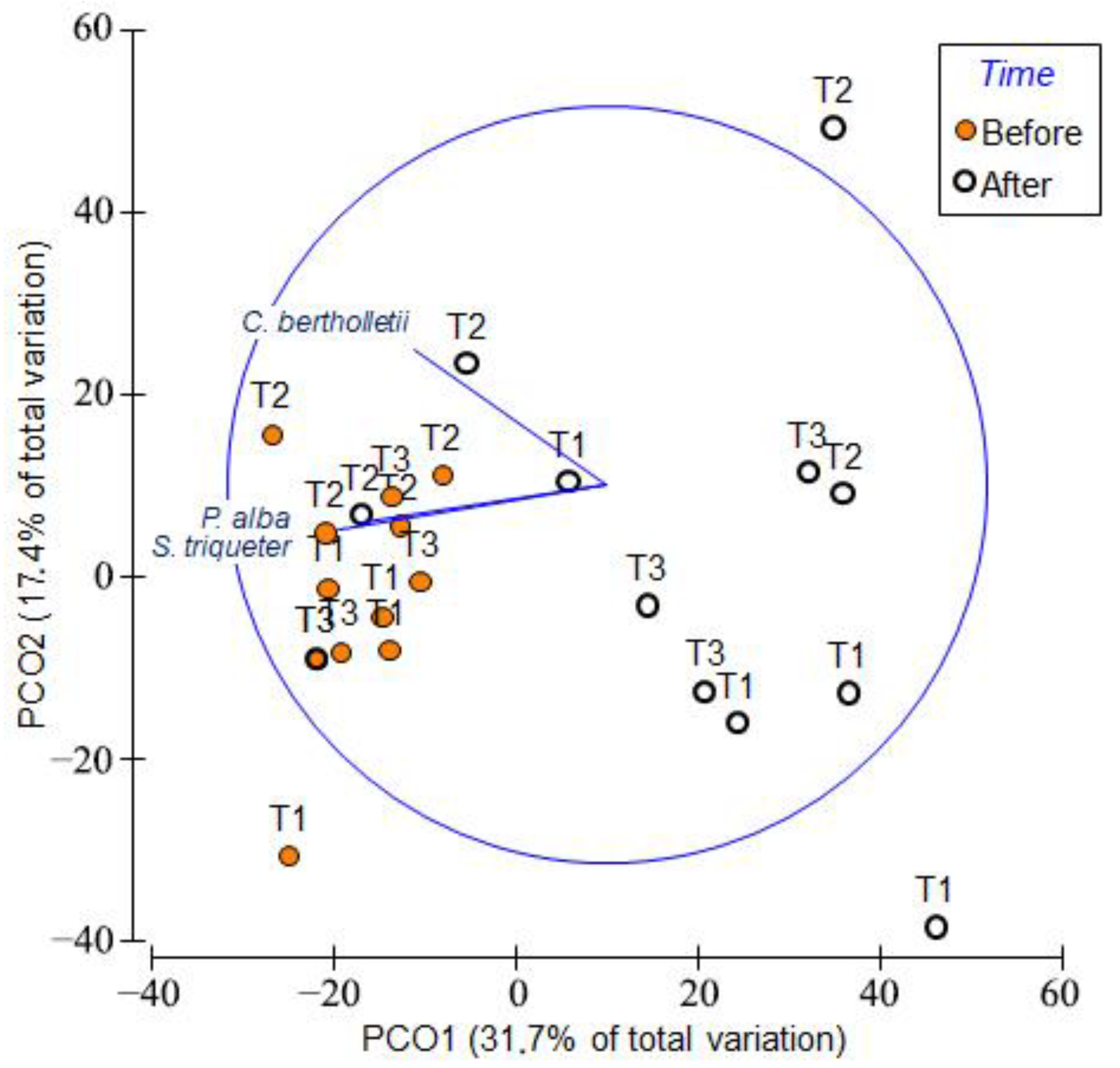
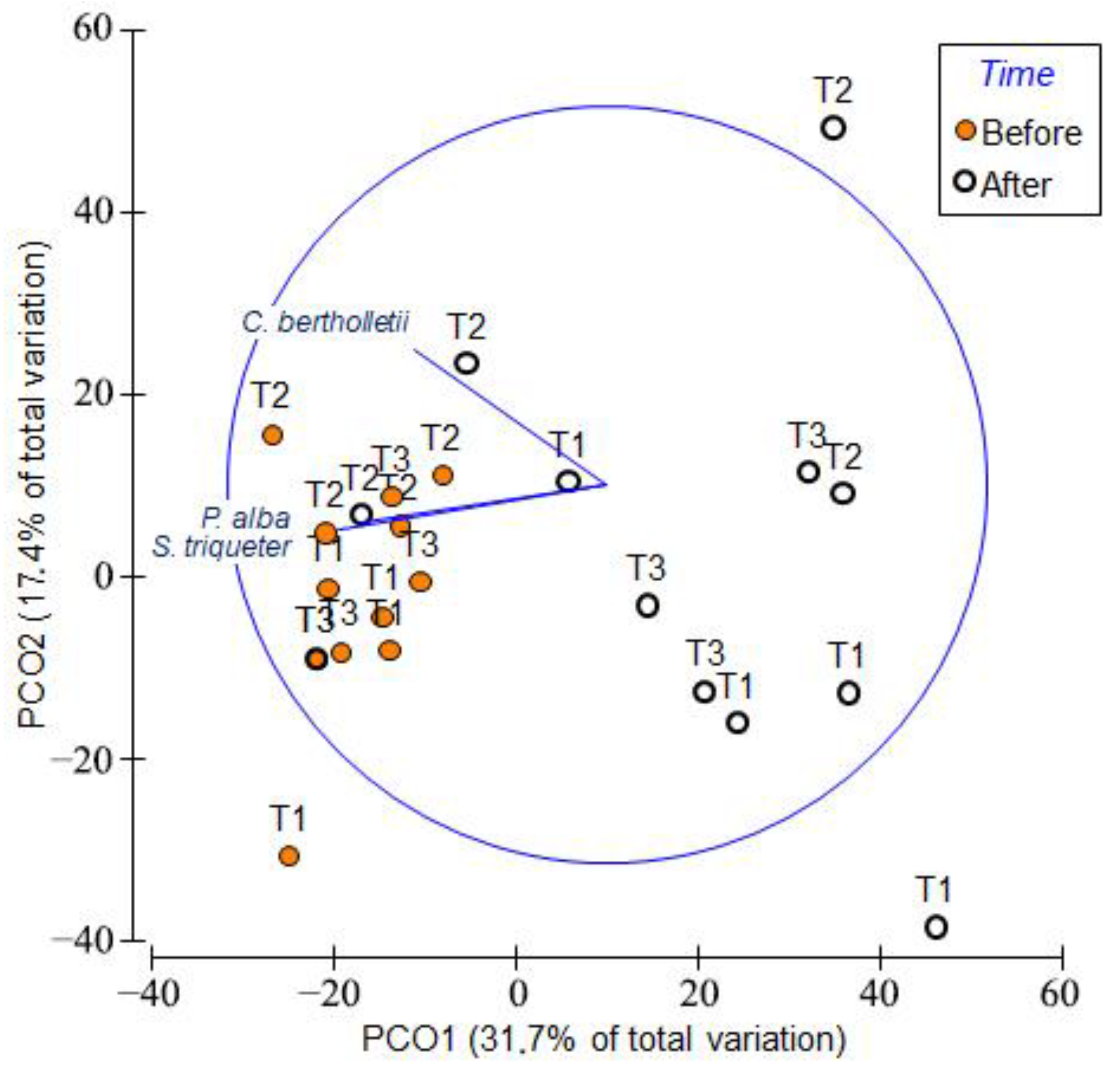
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carlton, J.T. Biological invasions and cryptogenic species. Ecol. Soc. Am. 1996, 77, 1653–1655. [Google Scholar] [CrossRef]
- Bax, N.; Williamson, A.; Aguero, M.; Gonzalez, E.; Geeves, W. Marine Invasive Alien Species: A Threat to Global Biodiversity. Mar. Policy 2003, 27, 313–323. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global Ecological Impacts of Invasive Species in Aquatic Ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.A.; Brown, L.; Campbell, M.L.; Canning-Clode, J.; Carlton, J.T.; Castro, N.; Chainho, P.; Chan, F.T.; Creed, J.C.; Curd, A.; et al. Trends in the Detection of Aquatic Non-Indigenous Species across Global Marine, Estuarine and Freshwater Ecosystems: A 50-Year Perspective. Divers. Distrib. 2020, 26, 1780–1797. [Google Scholar] [CrossRef]
- Lezzi, M.; Del Pasqua, M.; Pierri, C.; Giangrande, A. Seasonal Non-Indigenous Species Succession in a Marine Macrofouling Invertebrate Community. Biol. Invasions 2018, 20, 937–961. [Google Scholar] [CrossRef]
- Carlton, J.T. Global Change and Biological Invasions in the Oceans. In Invasive Species in a Changing World; Mooney, H.A., Hobbs, .J., Eds.; Island Press: Washington, DC, USA, 2000; pp. 31–53. [Google Scholar]
- Ricciardi, A. Are Modern Biological Invasions an Unprecedented Form of Global Change? Conserv. Biol. 2007, 21, 329–336. [Google Scholar] [CrossRef]
- Russell, J.C.; Blackburn, T.M. The Rise of Invasive Species Denialism. Trends Ecol. Evol. 2017, 32, 3–6. [Google Scholar] [CrossRef]
- García-Gómez, J.C.; Sempere-Valverde, J.; González, A.R.; Martínez-Chacón, M.; Olaya-Ponzone, L.; Sánchez-Moyano, E.; Ostalé-Valriberas, E.; Megina, C. From Exotic to Invasive in Record Time: The Extreme Impact of Rugulopteryx Okamurae (Dictyotales, Ochrophyta) in the Strait of Gibraltar. Sci. Total Environ. 2020, 704, 135408. [Google Scholar] [CrossRef]
- Rai, P.K.; Singh, J.S. Invasive Alien Plant Species: Their Impact on Environment, Ecosystem Services and Human Health. Ecol. Indic. 2020, 111, 106020. [Google Scholar]
- Castro, N.; Ramalhosa, P.; Cacabelos, E.; Costa, J.L.; Canning-Clode, J.; Gestoso, I. Winners and Losers: Prevalence of Non-Indigenous Species under Different Simulated Marine Heatwaves and High Propagule Pressure in an Oceanic Island. Mar. Ecol. Prog. Ser. 2021, 668, 21–38. [Google Scholar] [CrossRef]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing Redundancy in Invasion Ecology by Integrating Hypotheses into a Single Theoretical Framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef] [Green Version]
- Keller, R.P.; Geist, J.; Jeschke, J.M.; Kühn, I. Invasive Species in Europe: Ecology, Status, and Policy. Environ. Sci. Eur. 2011, 23, 23. [Google Scholar] [CrossRef] [Green Version]
- Geburzi, J.C.; McCarthy, M.L. How Do They Do It?–Understanding the Success of Marine Invasive Species. In YOUMARES 8–Oceans Across Boundaries: Learning from Each Other; Springer: Cham, Switzerland, 2018; pp. 109–124. [Google Scholar]
- Sareyka, J.; Kraufvelin, P.; Lenz, M.; Lindström, M.; Tollrian, R.; Wahl, M. Differences in Stress Tolerance and Brood Size between a Non-Indigenous and an Indigenous Gammarid in the Northern Baltic Sea. Mar. Biol. 2011, 158, 2001–2008. [Google Scholar] [CrossRef]
- Papacostas, K.J.; Rielly-carroll, E.W.; Georgian, S.E.; Long, D.J.; Princiotta, S.D.; Quattrini, A.M.; Reuter, K.E. Biological Mechanisms of Marine Invasions. Mar. Ecol. Prog. Ser. 2017, 565, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Terlizzi, A.; Felline, S.; Lionetto, M.G.; Caricato, R.; Perfetti, V.; Cutignano, A.; Mollo, E. Detrimental Physiological Effects of the Invasive Alga Caulerpa Racemosa on the Mediterranean White Seabream Diplodus Sargus. Aquat. Biol. 2011, 12, 109–117. [Google Scholar] [CrossRef]
- Felline, S.; Caricato, R.; Cutignano, A.; Gorbi, S.; Lionetto, M.G.; Mollo, E.; Regoli, F.; Terlizzi, A. Subtle Effects of Biological Invasions: Cellular and Physiological Responses of Fish Eating the Exotic Pest Caulerpa racemosa. PLoS ONE 2012, 7, e38763. [Google Scholar] [CrossRef] [Green Version]
- López, D.P.; Freestone, A.L. History of Co-occurrence Shapes Predation Effects on Functional Diversity and Structure at Low Latitudes. Funct. Ecol. 2021, 35, 535–545. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.J. Exotic Plant Invasions and the Enemy Release Hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Shea, K.; Chesson, P. Community Ecology Theory as a Framework for Biological Invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is Invasion Success Explained by the Enemy Release Hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Riera, L.; Ramalhosa, P.; Canning-Clode, J.; Gestoso, I. Variability in the Settlement of Non-Indigenous Species in Benthic Communities from an Oceanic Island. Helgol. Mar. Res. 2018, 72, 15. [Google Scholar] [CrossRef] [Green Version]
- Byers, J.E. Impact of Non-indigenous Species on Natives Enhanced by Anthropogenic Alteration of Selection Regimes. Oikos 2002, 97, 449–458. [Google Scholar] [CrossRef]
- DeRivera, C.E.; Ruiz, G.M.; Hines, A.H.; Jivoff, P. Biotic Resistance to Invasion: Native Predator Limits Abundance and Distribution of an Introduced Crab. Ecology 2005, 86, 3364–3376. [Google Scholar] [CrossRef]
- Freestone, A.L.; Ruiz, G.M.; Torchin, M.E. Stronger Biotic Resistance in Tropics Relative to Temperate Zone: Effects of Predation on Marine Invasion Dynamics. Ecology 2013, 94, 1370–1377. [Google Scholar] [CrossRef]
- Santamaría, J.; Tomas, F.; Ballesteros, E.; Ruiz, J.M.; Terrados, J.; Cebrian, E. A Little Can Be Enough. Native Fish from the Western Mediterranean Sea Can Act as a Control Agent for the Invasive Alga Caulerpa cylindracea. Peer J. Prepr. 2018, 6, e26772v1. [Google Scholar]
- Schoener, T.W.; Spiller, D.A. Effect of Predators and Area on Invasion: An Experiment with Island Spiders. Science 1995, 267, 1811–1813. [Google Scholar] [CrossRef]
- Forrest, B.M.; Fletcher, L.M.; Atalah, J.; Piola, R.F.; Hopkins, G.A. Predation Limits Spread of Didemnum Vexillum into Natural Habitats from Refuges on Anthropogenic Structures. PLoS ONE 2013, 8, e82229. [Google Scholar] [CrossRef] [Green Version]
- Simkanin, C.; Dower, J.F.; Filip, N.; Jamieson, G.; Therriault, T.W. Biotic Resistance to the Infiltration of Natural Benthic Habitats: Examining the Role of Predation in the Distribution of the Invasive Ascidian Botrylloides violaceus. J. Exp. Mar. Bio. Ecol. 2013, 439, 76–83. [Google Scholar] [CrossRef]
- Järv, L.; Kotta, J.; Kotta, I.; Raid, T. Linking the Structure of Benthic Invertebrate Communities and the Diet of Native and Invasive Fish Species in a Brackish Water Ecosystem. In Annales Zoologici Fennici; BioOne: Helsinki, Finland, 2011; Volume 48, pp. 129–141. [Google Scholar]
- Vonesh, J.; McCoy, M.; Altwegg, R.; Landi, P.; Measey, J. Functional Responses Can’t Unify Invasion Ecology. Biol. Invasions 2017, 19, 1673–1676. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Ibáñez, A.; Gestoso, I.; Wirtz, P.; Kaufmann, M.; Serrão, E.A.; Canning-Clode, J.; Cacabelos, E. The Collapse of Marine Forests: Drastic Reduction in Populations of the Family Sargassaceae in Madeira Island (NE Atlantic). Reg. Environ. Chang. 2021, 21, 71. [Google Scholar] [CrossRef]
- Calizza, E.; Rossi, L.; Careddu, G.; Caputi, S.S.; Costantini, M.L. A Novel Approach to Quantifying Trophic Interaction Strengths and Impact of Invasive Species in Food Webs. Biol. Invasions 2021, 23, 2093–2107. [Google Scholar] [CrossRef]
- Parretti, P.; Ros, M.; Gestoso, I.; Ramalhosa, P.; Costa, A.C.; Canning-Clode, J. Assessing Biotic Interactions between a Non-Indigenous Amphipod and Its Congener in a Future Climate Change Scenario. Aquat. Invasions 2021, 16, 186–207. [Google Scholar] [CrossRef]
- Hall, S.J.; Raffaelli, D.; Turrell, W.R. Predator-Caging Experiments in Marine Systems: A Reexamination of Their Value. Am. Nat. 1990, 136, 657–672. [Google Scholar] [CrossRef]
- Miranda, R.J.; José de Anchieta, C.C.; Mariano-Neto, E.; Sippo, J.Z.; Barros, F. Do Invasive Corals Alter Coral Reef Processes? An Empirical Approach Evaluating Reef Fish Trophic Interactions. Mar. Environ. Res. 2018, 138, 19–27. [Google Scholar] [CrossRef]
- Whitmarsh, S.K.; Fairweather, P.G.; Huveneers, C. What Is Big BRUVver up to? Methods and Uses of Baited Underwater Video. Rev. Fish Biol. Fish. 2017, 27, 53–73. [Google Scholar] [CrossRef]
- Zarco-Perello, S.; Enríquez, S. Remote Underwater Video Reveals Higher Fish Diversity and Abundance in Seagrass Meadows, and Habitat Differences in Trophic Interactions. Sci. Rep. 2019, 9, 6596. [Google Scholar] [CrossRef]
- Oricchio, F.T.; Pastro, G.; Vieira, E.A.; Flores, A.A.V.; Gibran, F.Z.; Dias, G.M. Distinct Community Dynamics at Two Artificial Habitats in a Recreational Marina. Mar. Environ. Res. 2016, 122, 85–92. [Google Scholar] [CrossRef]
- Chebaane, S.; Sempere-Valverde, J.; Dorai, S.; Kacem, A.; Sghaier, Y.R. A Preliminary Inventory of Alien and Cryptogenic Species in Monastir Bay, Tunisia: Spatial Distribution, Introduction Trends and Pathways. Mediterr. Mar. Sci. 2019, 20, 616–626. [Google Scholar] [CrossRef]
- Martínez-Laiz, G.; Ulman, A.; Ros, M.; Marchini, A. Is Recreational Boating a Potential Vector for Non-Indigenous Peracarid Crustaceans in the Mediterranean Sea? A Combined Biological and Social Approach. Mar. Pollut. Bull. 2019, 140, 403–415. [Google Scholar] [CrossRef]
- Castro, N.; Ramalhosa, P.; Jiménez, J.; Costa, J.L.; Gestoso, I.; Canning-Clode, J. Exploring Marine Invasions Connectivity in a NE Atlantic Island through the Lens of Historical Maritime Traffic Patterns. Reg. Stud. Mar. Sci. 2020, 37, 101333. [Google Scholar] [CrossRef]
- Canning-Clode, J.; Fofonoff, P.W.; McCann, L.; Carlton, J.T.; Ruiz, G.M. Marine Invasions on a Subtropical Island: Fouling Studies and New Records in a Recent Marina on Madeira Island (Eastern Atlantic Ocean). Aquat. Invasions 2013, 8, 261–270. [Google Scholar] [CrossRef]
- Afonso, I.; Berecibar, E.; Castro, N.; Costa, J.L.; Frias, P.; Henriques, F.; Moreira, P.; Oliveira, P.M.; Silva, G.; Chainho, P. Assessment of the Colonization and Dispersal Success of Non-Indigenous Species Introduced in Recreational Marinas along the Estuarine Gradient. Ecol. Indic. 2020, 113, 106147. [Google Scholar] [CrossRef]
- Olyarnik, S.V.; Bracken, M.E.S.; Byrnes, J.E.; Hughes, A.R.; Hultgren, K.M.; Stachowicz, J.J. Ecological Factors Affecting Community Invasibility BT—Biological Invasions in Marine Ecosystems: Ecological, Management, and Geographic Perspectives; Rilov, G., Crooks, J.A., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2009; pp. 215–238. [Google Scholar] [CrossRef]
- Como, S.; Rossi, F.; Lardicci, C. Caging Experiment: Relationship between Mesh Size and Artifacts. J. Exp. Mar. Bio. Ecol. 2006, 335, 157–166. [Google Scholar] [CrossRef]
- Gestoso, I.; Ramalhosa, P.; Canning-Clode, J. Biotic Effects during the Settlement Process of Non-Indigenous Species in Marine Benthic Communities. Aquat. Invasions 2018, 13, 247–259. [Google Scholar] [CrossRef]
- Marraffini, M.L.; Ashton, G.V.; Brown, C.W.; Chang, A.L.; Ruiz, G.M. Settlement Plates as Monitoring Devices for Non-Indigenous Species in Marine Fouling Communities. Manag. Biol. Invasions 2017, 8, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel Extensions (CPCe): A Visual Basic Program for the Determination of Coral and Substrate Coverage Using Random Point Count Methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Canning-Clode, J. Global Diversity Patterns in Marine Fouling Communities-Exploring Latitudinal Effects and the Local-Regional Richness Relationship. Ph.D. Thesis, Christian-Albrechts-Universität, Kiel, Germany, 2008. [Google Scholar]
- Castro, N.; Carlton, J.T.; Costa, A.C.; Marques, C.S.; Hewitt, C.L.; Cacabelos, E.; Lopes, E.; Gizzi, F.; Gestoso, I.; Monteiro, J.G.; et al. Diversity and patterns of marine non-native species in the archipelagos of Macaronesia. Divers. Distrib. 2022, 28, 667–684. [Google Scholar] [CrossRef]
- Chainho, P.; Fernandes, A.; Amorim, A.; Ávila, S.P.; Canning-Clode, J.; Castro, J.J.; Costa, A.C.; Costa, J.L.; Cruz, T.; Gollasch, S.; et al. Non-indigenous species in Portuguese coastal areas, coastal lagoons, estuaries and islands. Estuar. Coast. Shelf Sci. 2015, 167, 199–211. [Google Scholar] [CrossRef]
- Cruz, T. Esponjas Marinas de Canarias. Cons. Política Territ. Y Medio Ambient. Del Gob. Canar. S/C Tenerife 2002, 1–260. Available online: https://mdc.ulpgc.es/utils/getfile/collection/MDC/id/181302/filename/262673.pdf (accessed on 2 April 2022).
- Gestoso, I.; Ramalhosa, P.; Oliveira, P.; Canning-Clode, J. Marine Protected Communities against Biological Invasions: A Case Study from an Offshore Island. Mar. Pollut. Bull. 2017, 119, 72–80. [Google Scholar] [CrossRef]
- Knight-Jones, P.; Knight-Jones, E.W.; Buzhinskaya, G. Distribution and Interrelationships of Northern Spirorbid Genera. Bull. Mar. Sci. 1991, 48, 189–197. [Google Scholar]
- Levring, T. The Marine Algae of the Archipelago of Madeira. Bol. Mus. Munic. Funchal 1974, 28, 5–111. Available online: https://publications.cm-funchal.pt/jspui/handle/100/1231 (accessed on 5 April 2022).
- Neto, A.I.; Cravo, D.C.; Haroun, R.T. Checklist of the Benthic Marine Plants of the Madeira Archipelago. Bot. Mar. 2001, 44, 391–414. [Google Scholar] [CrossRef] [Green Version]
- Norman, A.M. The Polyzoa of Madeira and Neighbouring Islands. Zool. J. Linn. Soc. 1909, 30, 275–314. [Google Scholar] [CrossRef]
- Núnez, J.; Talavera, J.A. Fauna of the Polychaetous Annelids from Madeira. Bol. Mus. Munic. Funchal 1995, 4, 511–530. Available online: https://publications.cm-funchal.pt/jspui/bitstream/100/958/1/Bolmmf_s04B_1995_pp511-530.pdf (accessed on 5 April 2022).
- Png-Gonzalez, L.; Ramalhosa, P.; Gestoso, I.; Álvarez, S.; Nogueira, N. Non-Indigenous Species on Artificial Coastal Environments: Experimental Comparison between Aquaculture Farms and Recreational Marinas. J. Mar. Sci. Eng. 2021, 9, 1121. [Google Scholar] [CrossRef]
- Ramalhosa, P.; Gestoso, I.; Duarte, B.; Caçador, I.; Canning-Clode, J. Metal Pollution Affects Both Native and Non-Indigenous Biofouling Recruitment in a Subtropical Island System. Mar. Pollut. Bull. 2019, 141, 373–386. [Google Scholar] [CrossRef]
- Ramalhosa, P.; Gestoso, I.; Rocha, R.M.; Lambert, G.; Canning-Clode, J. Ascidian Biodiversity in the Shallow Waters of the Madeira Archipelago: Fouling Studies on Artificial Substrates and New Records. Reg. Stud. Mar. Sci. 2021, 43, 101672. [Google Scholar] [CrossRef]
- Ramalhosa, P.; Souto, J.; Canning-Clode, J. Diversity of Bugulidae (Bryozoa, Cheilostomata) Colonizing Artificial Substrates in the Madeira Archipelago (NE Atlantic Ocean). Helgol. Mar. Res. 2017, 71, 1. [Google Scholar] [CrossRef] [Green Version]
- Souto, J.; Ramalhosa, P.; Canning-Clode, J. Three Non-Indigenous Species from Madeira Harbors, Including a New Species of Parasmittina (Bryozoa). Mar. Biodivers. 2018, 48, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, P. Twelve Invertebrate and Eight Fish Species New to the Marine Fauna of Madeira, and a Discussion of the Zoogeography of the Area. Helgoländer Meeresunters. 1998, 52, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, P. On a Collection of Hydroids (Cnidaria, Hydrozoa) from the Madeira Archipelago. Arquipélago-Life Mar. Sci. 2007, 24, 11–16. [Google Scholar]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods. Prim. Plymouth UK 2008, 214pp. Available online: http://updates.primer-e.com/primer7/manuals/PERMANOVA+_manual.pdf (accessed on 5 March 2022).
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER V7. In PRIMER-E: Plymouth; Plymouth Marine Laboratory: Devon, UK, 2015. [Google Scholar]
- Monteiro, J.G.; Jiménez, J.L.; Gizzi, F.; Přikryl, P.; Lefcheck, J.S.; Santos, R.S.; Canning-Clode, J. Novel Approach to Enhance Coastal Habitat and Biotope Mapping with Drone Aerial Imagery Analysis. Sci. Rep. 2021, 11, 574. [Google Scholar] [CrossRef]
- Madenjian, C.P.; Stapanian, M.A.; Witzel, L.D.; Einhouse, D.W.; Pothoven, S.A.; Whitford, H.L. Evidence for Predatory Control of the Invasive Round Goby. Biol. Invasions 2011, 13, 987–1002. [Google Scholar] [CrossRef]
- Warburton, K.; Hughes, R. Learning of Foraging Skills by Fish. In Fish Cognition and Behavior; Wiley Online Books: Oxford, UK, 2011; pp. 10–35. [Google Scholar] [CrossRef]
- Felline, S.; Mollo, E.; Cutignano, A.; Grauso, L.; Andaloro, F.; Castriota, L.; Consoli, P.; Falautano, M.; Sinopoli, M.; Terlizzi, A. Preliminary Observations of Caulerpin Accumulation from the Invasive Caulerpa Cylindracea in Native Mediterranean Fish Species. Aquat. Biol. 2017, 26, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Magliozzi, L.; Almada, F.; Robalo, J.; Mollo, E.; Polese, G.; Gonçalves, E.J.; Felline, S.; Terlizzi, A.; D’Aniello, B. Cryptic Effects of Biological Invasions: Reduction of the Aggressive Behaviour of a Native Fish under the Influence of an “Invasive” Biomolecule. PLoS ONE 2017, 12, e0185620. [Google Scholar] [CrossRef]
- Costa, P.R.; Estevez, P.; Castro, D.; Soliño, L.; Gouveia, N.; Santos, C.; Rodrigues, S.M.; Leao, J.M.; Gago-Martínez, A. New Insights into the Occurrence and Toxin Profile of Ciguatoxins in Selvagens Islands (Madeira, Portugal). Toxins 2018, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Tortonese, E. Fauna d’Italia: Osteichthyes (Pesci Ossei); Calderini: Bologna, Italy, 1970. [Google Scholar]
- González, J.A. Sparisoma (Euscarus) Cretense (L.)(Pisces, Scaridae): Zoogeografía y Ciclo Reproductor En Las Islas Canarias. Courier Forsch. Inst. Senckenb. 1993, 159, 429–435. [Google Scholar]
- Jiménez-Alvarado, D.; Sarmiento-Lezcano, A.; Guerra-Marrero, A.; Tuya, F.; Santana Del Pino, Á.; Sealey, M.J.; Castro, J.J. Historical Photographs of Captures of Recreational Fishers Indicate Overexploitation of Nearshore Resources at an Oceanic Island. J. Fish Biol. 2019, 94, 857–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrakis, G.; Papaconstantinou, C. Biology of Sparisoma Cretense in the Dodecanese (Greece). J. Appl. Ichthyol. 1990, 6, 14–23. [Google Scholar] [CrossRef]
- Bullock, A.E.; Monod, T. Myologie Céphalique de Deux Poissons Perroquets (Teleostei: Scaridae). Cybium 1997, 21, 173–199. [Google Scholar]
- Tuya, F.; Boyra, A.; Sanchez-Jerez, P.; Barbera, C.; Haroun, R.J. Relationships between Rocky-Reef Fish Assemblages, the Sea Urchin Diadema Antillarum and Macroalgae throughout the Canarian Archipelago. Mar. Ecol. Prog. Ser. 2004, 278, 157–169. [Google Scholar] [CrossRef]
- Azzurro, E.; Fanelli, E.; Mostarda, E.; Catra, M.; Andaloro, F. Resource Partitioning among Early Colonizing Siganus Luridus and Native Herbivorous Fish in the Mediterranean: An Integrated Study Based on Gut-Content Analysis and Stable Isotope Signatures. J. Mar. Biol. Assoc. UK 2007, 87, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Afonso, P.; Fontes, J.; Holland, K.N.; Santos, R.S. Social Status Determines Behaviour and Habitat Usage in a Temperate Parrotfish: Implications for Marine Reserve Design. Mar. Ecol. Prog. Ser. 2008, 359, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Clemente, S.; Hernández, J.C.; Rodríguez, A.; Brito, A. Identifying Keystone Predators and the Importance of Preserving Functional Diversity in Sublittoral Rocky-Bottom Areas. Mar. Ecol. Prog. Ser. 2010, 413, 55–67. [Google Scholar] [CrossRef]
- Alomar, C.; Deudero, S.; Andaloro, F.; Castriota, L.; Consoli, P.; Falautano, M.; Sinopoli, M. Caulerpa Cylindracea Sonder Invasion Modifies Trophic Niche in Infralittoral Rocky Benthic Community. Mar. Environ. Res. 2016, 120, 86–92. [Google Scholar] [CrossRef]
- Marić, M.; De Troch, M.; Occhipinti-Ambrogi, A.; Olenin, S. Trophic Interactions between Indigenous and Non-Indigenous Species in Lampedusa Island, Mediterranean Sea. Mar. Environ. Res. 2016, 120, 182–190. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Ballesteros, E.; Clemente, S.; Gonçalves, E.J.; Estep, A.; Rose, P.; Sala, E. Contrasts in the Marine Ecosystem of Two Macaronesian Islands: A Comparison between the Remote Selvagens Reserve and Madeira Island. PLoS ONE 2017, 12, e0187935. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.S.; Ruiz, G.M.; Altieri, A.H.; Torchin, M.E. The Biogeography of Invasion in Tropical and Temperate Seagrass Beds: Testing Interactive Effects of Predation and Propagule Pressure. Divers. Distrib. 2019, 25, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Freestone, A.L.; Torchin, M.E.; Jurgens, L.J.; Bonfim, M.; López, D.P.; Repetto, M.F.; Schlöder, C.; Sewall, B.J.; Ruiz, G.M. Stronger Predation Intensity and Impact on Prey Communities in the Tropics. Ecology 2021, 102, e03428. [Google Scholar] [CrossRef] [PubMed]
- Flecker, A.S.; Taylor, B.W. Tropical Fishes as Biological Bulldozers: Density Effects on Resource Heterogeneity and Species Diversity. Ecology 2004, 85, 2267–2278. [Google Scholar] [CrossRef] [Green Version]
- Janiak, D.S.; Branson, D. Impacts of Habitat and Predation on Epifaunal Communities from Seagrass Beds and Artificial Structures. Mar. Environ. Res. 2021, 163, 105225. [Google Scholar] [CrossRef]
- Whitney, K.D.; Gabler, C.A. Rapid Evolution in Introduced Species,‘Invasive Traits’ and Recipient Communities: Challenges for Predicting Invasive Potential. Divers. Distrib. 2008, 14, 569–580. [Google Scholar] [CrossRef]
- Skein, L.; Alexander, M.E.; Robinson, T.B. Characteristics of Native Predators Are More Important than Those of Alien Prey in Determining the Success of Biotic Resistance in Marine Systems. Aquat. Ecol. 2021, 55, 97–106. [Google Scholar] [CrossRef]
- Micael, J.; Tempera, F.; Berning, B.; López-Fé, C.M.; Occhipinti-Ambrogi, A.; Costa, A.C. Shallow-Water Bryozoans from the Azores (Central North Atlantic): Native vs. Non-Indigenous Species, and a Method to Evaluate Taxonomic Uncertainty. Mar. Biodivers. 2019, 49, 469–480. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Whitlatch, R.B.; Osman, R.W. Species Diversity and Invasion Resistance in a Marine Ecosystem. Science 1999, 286, 1577–1579. [Google Scholar] [CrossRef] [Green Version]
- Glasby, T.M.; Connell, S.D.; Holloway, M.G.; Hewitt, C.L. Nonindigenous Biota on Artificial Structures: Could Habitat Creation Facilitate Biological Invasions? Mar. Biol. 2007, 151, 887–895. [Google Scholar] [CrossRef]
- Ruiz, G.M.; Freestone, A.L.; Fofonoff, P.W.; Simkanin, C. Habitat Distribution and Heterogeneity in Marine Invasion Dynamics: The Importance of Hard Substrate and Artificial Structure. In Marine Hard Bottom Communities; Springer: Berlin/Heidelberg, Germany, 2009; pp. 321–332. [Google Scholar]
- Steele, L.; Ray, C.; Guidone, M. High Phenolic Content Fails to Deter Mesograzer Consumption of Myriophyllum Spicatum (Eurasian Watermilfoil) in New England. Aquat. Ecol. 2018, 52, 255–267. [Google Scholar] [CrossRef]
- Price, N.W.; Chen, K.-S.; Chen, C.A.; Wen, C.K.-C. Scraping and Grazing Herbivorous/Detritivorous fish display opposite feeding behaviours under different protection regulations. Res. Sq. 2021, 17, 876–891. [Google Scholar] [CrossRef]
- Tyrrell, M.C.; Byers, J.E. Do Artificial Substrates Favor Nonindigenous Fouling Species over Native Species? J. Exp. Mar. Bio. Ecol. 2007, 342, 54–60. [Google Scholar] [CrossRef]
- Tepolt, C.K.; Somero, G.N. Master of All Trades: Thermal Acclimation and Adaptation of Cardiac Function in a Broadly Distributed Marine Invasive Species, the European Green Crab, Carcinus Maenas. J. Exp. Biol. 2014, 217, 1129–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.R.; Alcover, J.A.; Blackburn, T.M.; Bover, P.; Duncan, R.P.; Hume, J.P.; Louys, J.; Meijer, H.J.M.; Rando, J.C.; Wilmshurst, J.M. Island Extinctions: Processes, Patterns, and Potential for Ecosystem Restoration. Environ. Conserv. 2017, 44, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.C.; Kueffer, C. Island Biodiversity in the Anthropocene. Annu. Rev. Environ. Resour. 2019, 44, 31–60. [Google Scholar] [CrossRef]
- Kremer, L.P.; da Rocha, R.M. The Biotic Resistance Role of Fish Predation in Fouling Communities. Biol. Invasions 2016, 18, 3223–3237. [Google Scholar] [CrossRef]
- Humphries, A.T.; McClanahan, T.R.; McQuaid, C.D. Algal Turf Consumption by Sea Urchins and Fishes Is Mediated by Fisheries Management on Coral Reefs in Kenya. Coral Reefs 2020, 39, 1137–1146. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foragers Interest | Foragers Feeding | ||||||
|---|---|---|---|---|---|---|---|
| Source | df | MS | Pseudo-F | P (Perm) | MS | Pseudo-F | P (Perm) |
| Treatment | 2 | 826.48 | 70.8584 | 0.0172 * | 541.58 | 2.8368 | 0.1381 |
| Trials | 3 | 155.17 | 10.4754 | 0.3144 | 408.75 | 2.141 | 0.1962 |
| Residuals | 6 | 105.17 | 190.92 | ||||
| Pair-wise | (T1 = T2) > T3 | ||||||
| Relative Abundance | Wet Biomass | ||||||
|---|---|---|---|---|---|---|---|
| Source | df | MS | Pseudo-F | P (Perm) | MS | Pseudo-F | P (Perm) |
| Treatment | 2 | 180.54 | 0.4924 | 0.6376 | 1.7689 | 7.4793 | 0.0059 * |
| Time | 1 | 22083 | 60.23 | 0.0001 * | 23.764 | 100.48 | 0.0001 * |
| Tr × Ti | 2 | 351.04 | 0.9575 | 0.419 | 1.9138 | 8.092 | 0.0032 * |
| Residuals | 18 | 366.64 | 0.2365 | ||||
| Pair-wise | Before > After | Before: T2 > (T1 = T3) After: T1 = T2 = T3 | |||||
| PERMANOVA | PERMDISP | |||||||
|---|---|---|---|---|---|---|---|---|
| Source | df | MS | Pseudo-F | P (Perm) | df | res. df | F | P (Perm) |
| Treatment | 2 | 4224.8 | 3.3431 | 0.0007 * | 2 | 21 | 1.555 | 0.4229 |
| Time | 1 | 7617.8 | 6.028 | 0.0001 * | 1 | 22 | 19.836 | 0.0002 * |
| Tr × Ti | 2 | 1219.8 | 0.9652 | 0.4983 | 2 | 18 | 24.108 | 0.0001 * |
| Residuals | 18 | 1263.7 | 18 | |||||
| Pair-wise | Treatment: T1 ≠ T2 ≠ T3 |
Before: (T1 = T3) > T2 After: T1 = T2 = T3 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chebaane, S.; Canning-Clode, J.; Ramalhosa, P.; Belz, J.; Castro, N.; Órfão, I.; Sempere-Valverde, J.; Engelen, A.H.; Pais, M.P.; Monteiro, J.G. From Plates to Baits: Using a Remote Video Foraging System to Study the Impact of Foraging on Fouling Non-Indigenous Species. J. Mar. Sci. Eng. 2022, 10, 611. https://doi.org/10.3390/jmse10050611
Chebaane S, Canning-Clode J, Ramalhosa P, Belz J, Castro N, Órfão I, Sempere-Valverde J, Engelen AH, Pais MP, Monteiro JG. From Plates to Baits: Using a Remote Video Foraging System to Study the Impact of Foraging on Fouling Non-Indigenous Species. Journal of Marine Science and Engineering. 2022; 10(5):611. https://doi.org/10.3390/jmse10050611
Chicago/Turabian StyleChebaane, Sahar, João Canning-Clode, Patrício Ramalhosa, Janina Belz, Nuno Castro, Inês Órfão, Juan Sempere-Valverde, Aschwin Hillebrand Engelen, Miguel Pessanha Pais, and João Gama Monteiro. 2022. "From Plates to Baits: Using a Remote Video Foraging System to Study the Impact of Foraging on Fouling Non-Indigenous Species" Journal of Marine Science and Engineering 10, no. 5: 611. https://doi.org/10.3390/jmse10050611
APA StyleChebaane, S., Canning-Clode, J., Ramalhosa, P., Belz, J., Castro, N., Órfão, I., Sempere-Valverde, J., Engelen, A. H., Pais, M. P., & Monteiro, J. G. (2022). From Plates to Baits: Using a Remote Video Foraging System to Study the Impact of Foraging on Fouling Non-Indigenous Species. Journal of Marine Science and Engineering, 10(5), 611. https://doi.org/10.3390/jmse10050611