
Estimation of the Benthic Habitat Zonation by Photo-Quadrat Image Analysis along the Fringing Reef of Weno Island, Chuuk, Micronesia

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
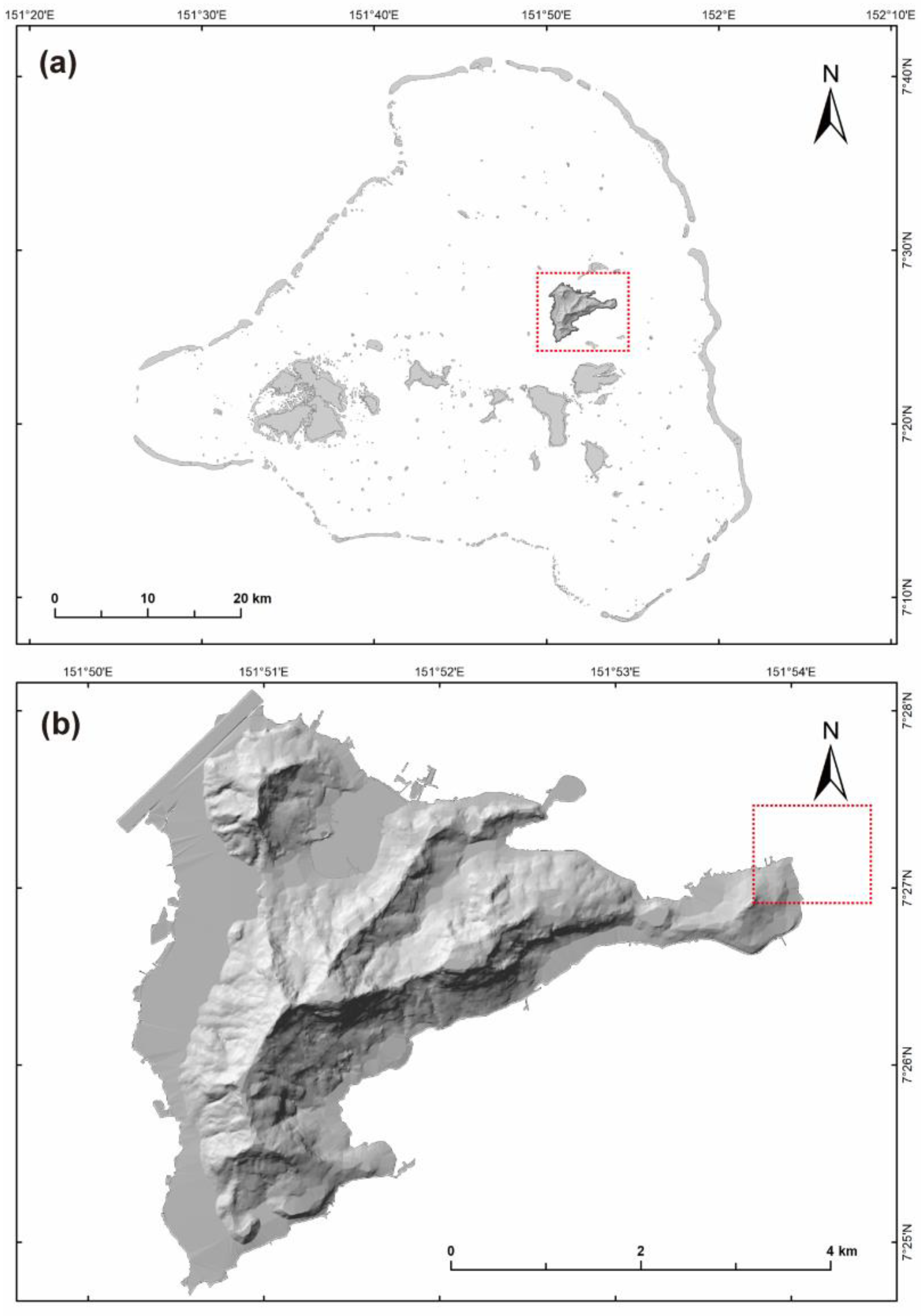
2.1. Study Site
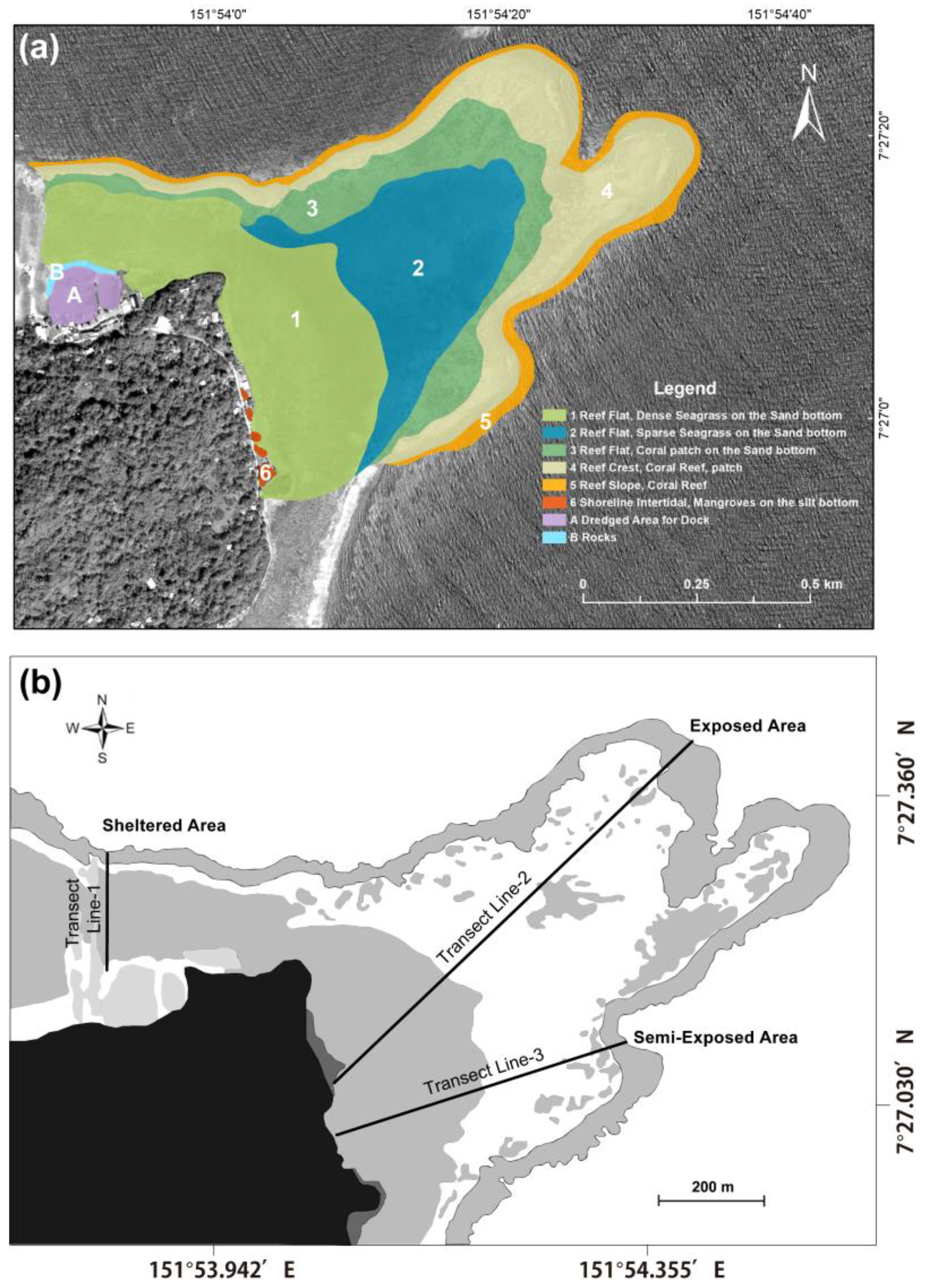
2.2. Mapping and Transect Lines Installation
2.3. Photo Analysis
2.4. Habitat Classification
2.5. Statistical Analysis
3. Results
3.1. Overall Distribution of Benthic Habitat
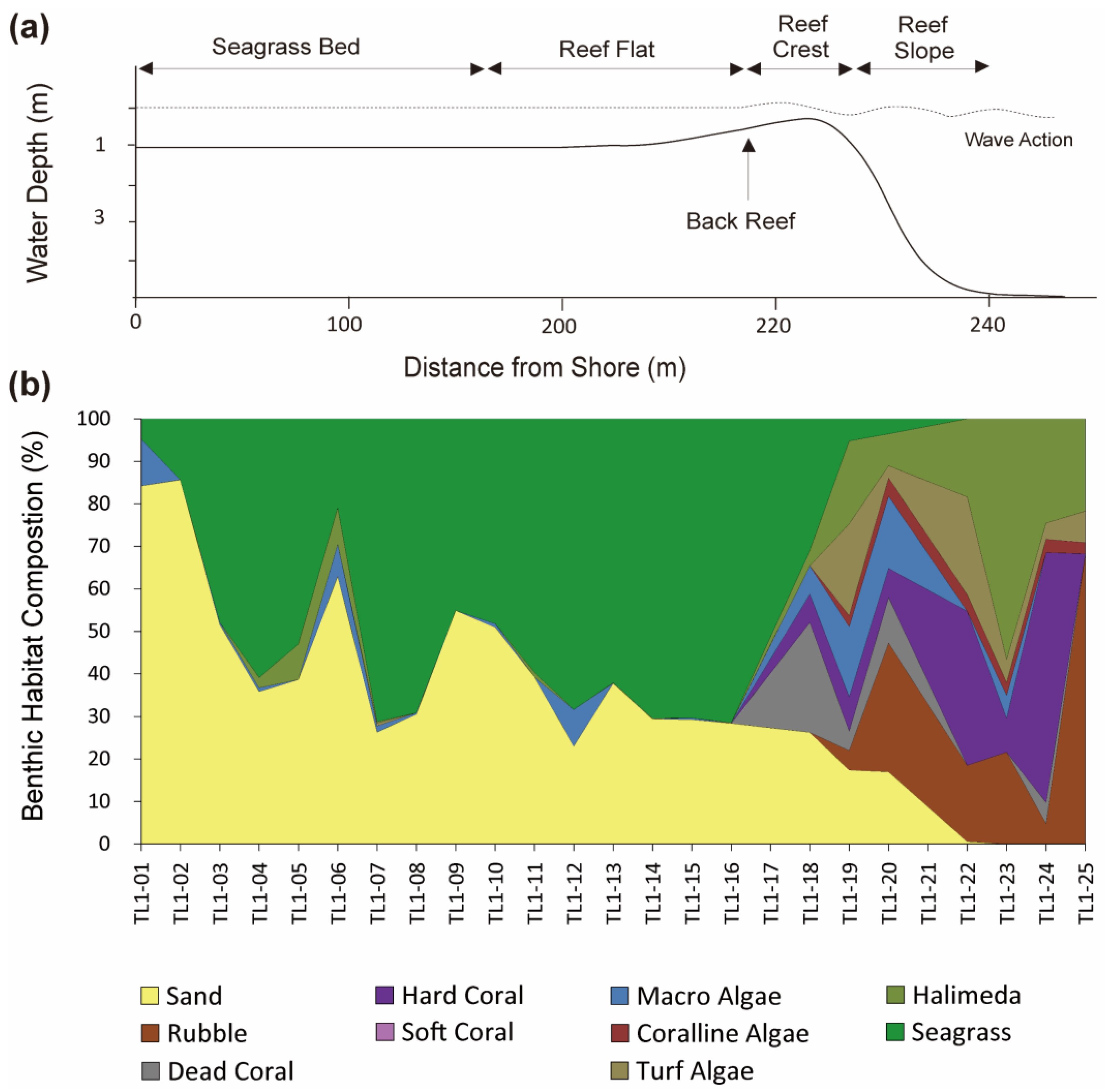
3.2. Transect Line-1 (TL-1)


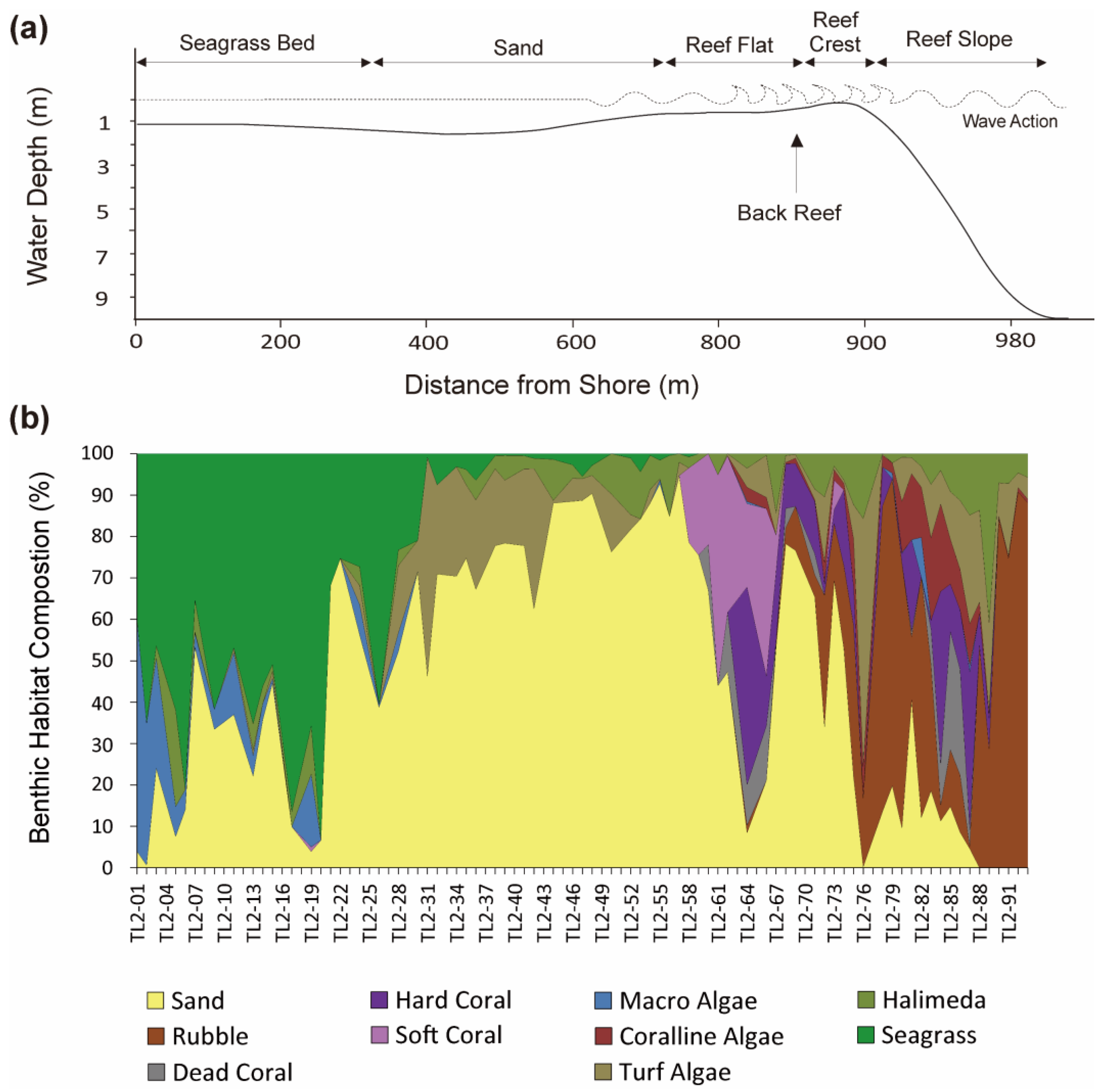
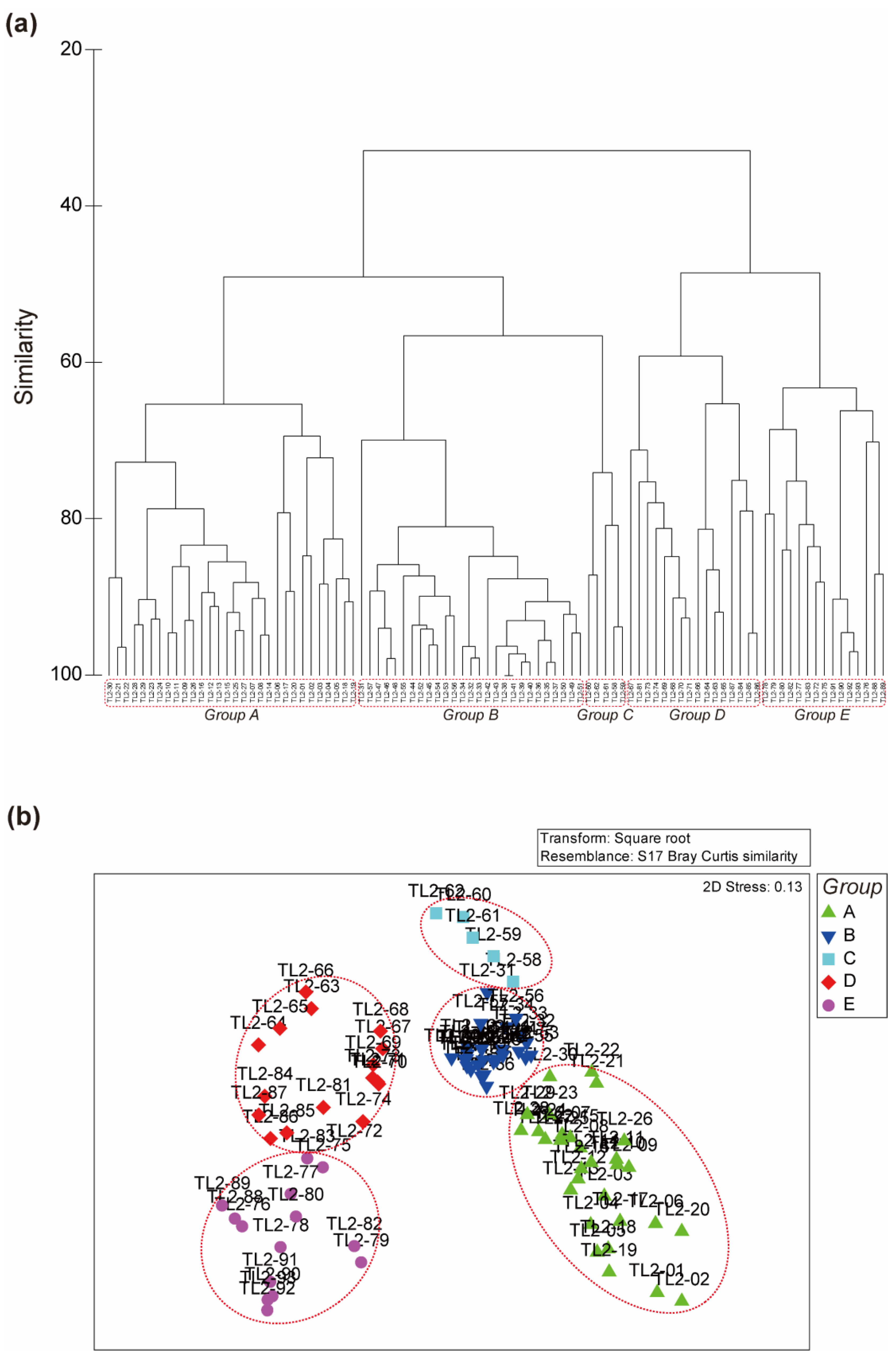
3.3. Transect Line-2 (TL-2)


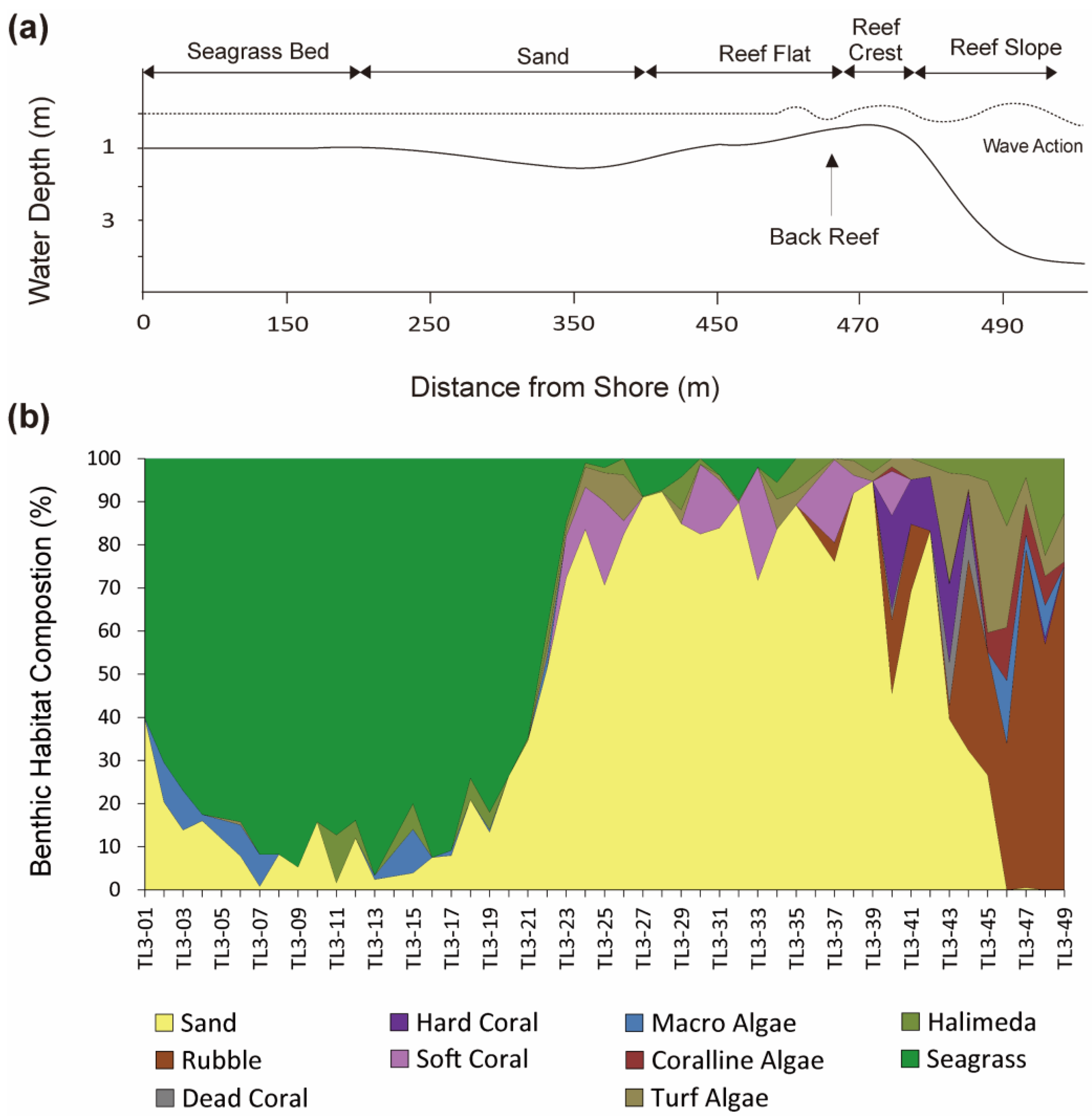
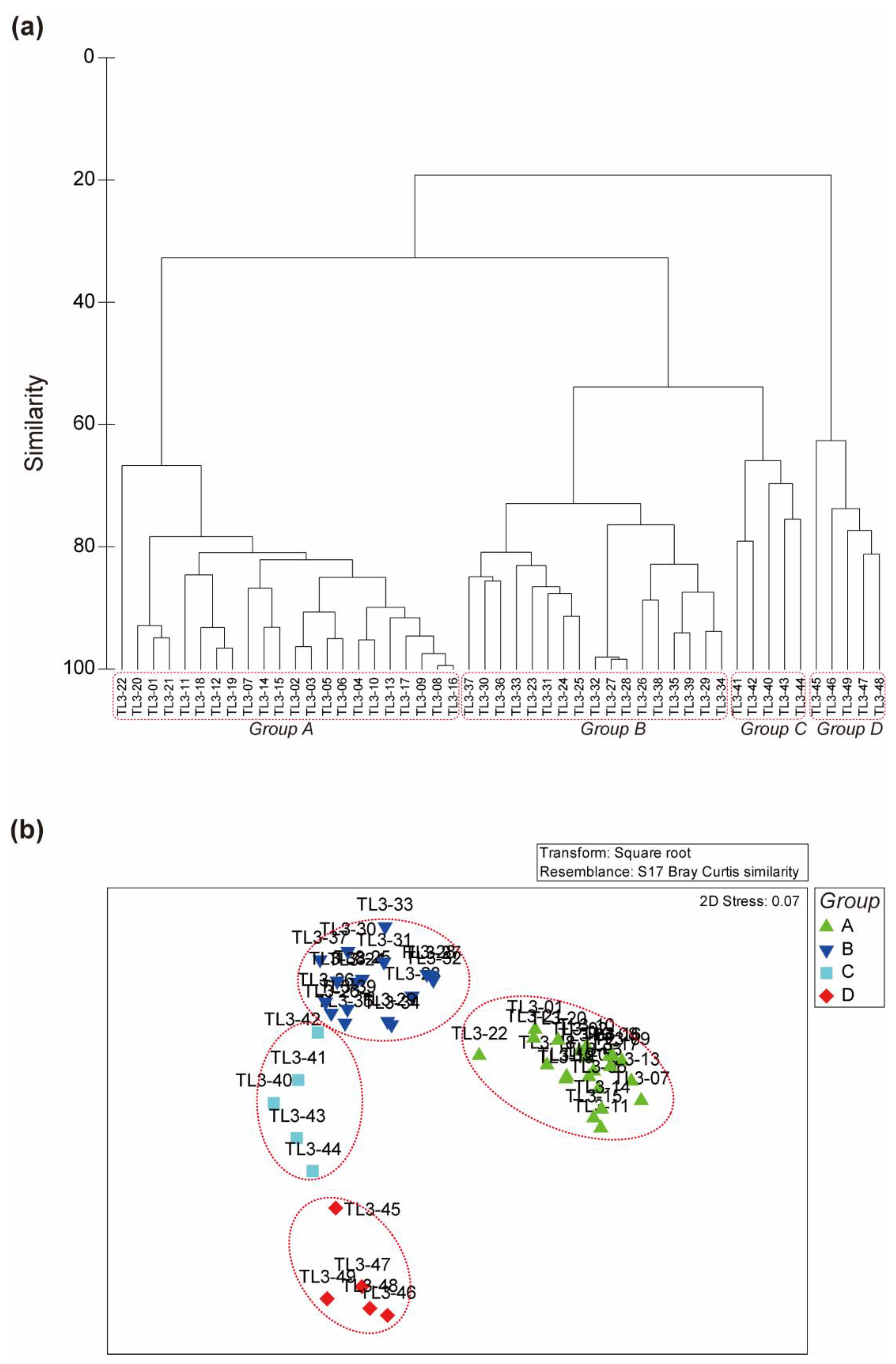
3.4. Transect Line-3 (TL-3)
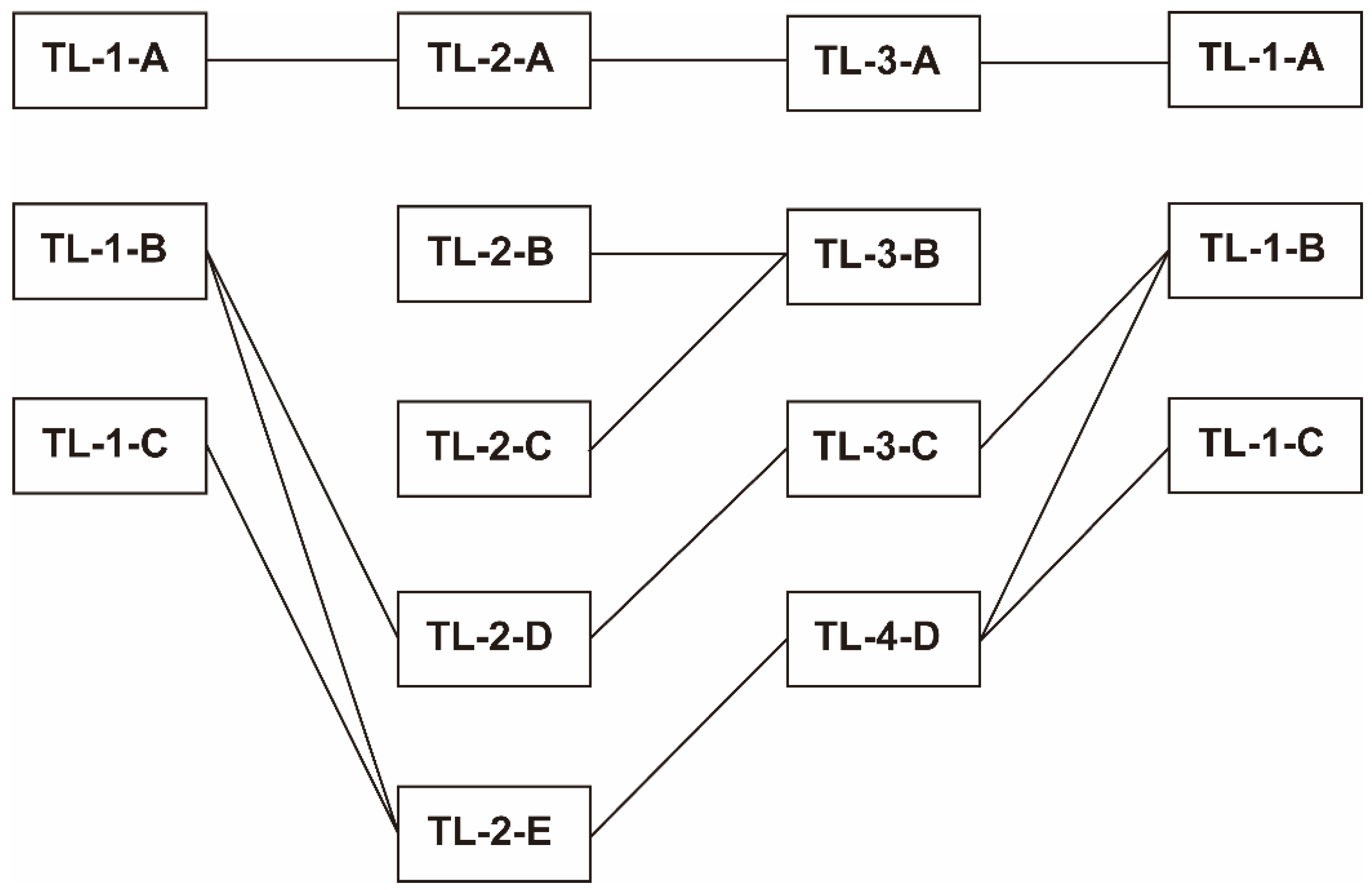
3.5. Comparison of Inter-Group Distance between the Three Transects



4. Discussion
4.1. Assessment of Methodology
4.2. Benthic Habitat Zonation Patterns by Habitat Zone
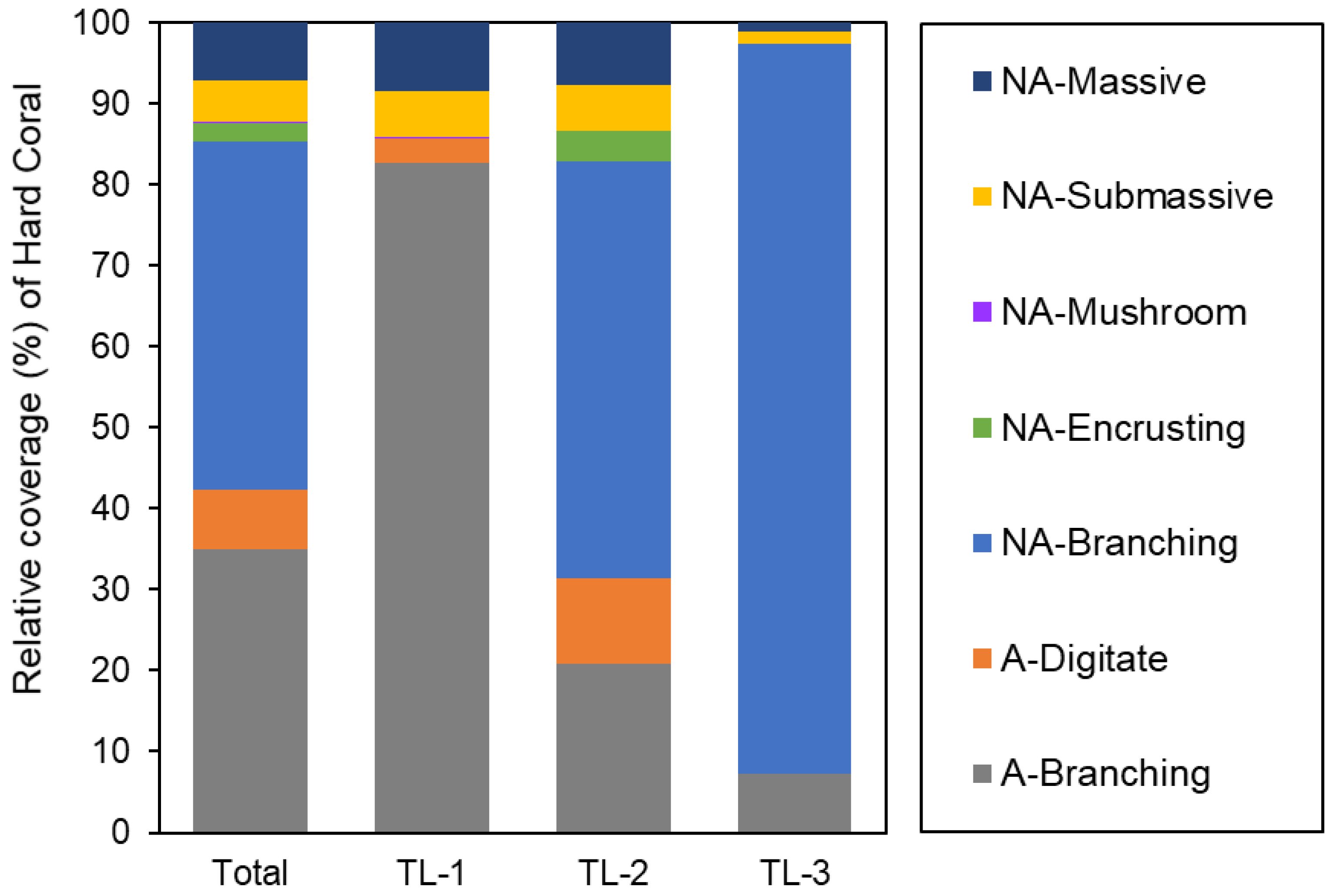
4.3. Benthic Habitat Zonation Patterns by Habitat Type Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lohrer, A.M.; Fukui, Y.; Wada, K.; Whitlatch, R.B. Structural complexity and vertical zonation of intertidal crabs, with focus on habitat requirements of the invasive Asian shore crab, Hemigrapsus sanguineus (de Haan). J. Exp. Mar. Biol. Ecol. 2000, 244, 203–217. [Google Scholar] [CrossRef]
- Oakley-Cogan, A.; Tebbett, S.B.; Bellwood, D.R. Habitat zonation on coral reefs: Structural complexity, nutritional resources and herbivorous fish distributions. PLoS ONE 2020, 15, e0233498. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Mi, C.-R.; Zhao, Y.-Z.; Yang, N.; Sun, Y.-H.; Xu, J.-L. Zonation of nature reserve according to the habitat requirement of conservation target: A case study on the endangered Brown Eared-Pheasant at Baihuashan Nature Reserve. Glob. Ecol. Conserv. 2021, 32, e01941. [Google Scholar] [CrossRef]
- Ninio, R.; Meekan, M.G. Spatial patterns in benthic communities and the dynamics of a mosaic ecosystem on the Great Barrier Reef, Australia. Coral Reefs 2002, 21, 95–104. [Google Scholar] [CrossRef]
- Harborne, A.R.; Mumby, P.J.; Micheli, F.; Perry, C.T.; Dahlgren, C.P.; Holmes, K.E.; Brumbaugh, D.R. The functional value of Caribbean coral reef, seagrass and mangrove habitats to ecosystem processes. Adv. Mar. Biol. 2006, 50, 57–189. [Google Scholar] [CrossRef]
- De Mazières, J. Spatial Distribution of Reefr Fish Communities an Investigation of the Coral Coast, Fiji Islands. Master’s Thesis, The University of the South Pacific, Suva, Fiji, 2008. [Google Scholar]
- Jackson, E.; Rowden, A.A.; Attrill, M.J.; Bossey, S.J.; Jones, M.B. The importance of seagrass beds as a habitat for fishery species. Oceanogr. Mar. Biol. Annu. Rev. 2001, 39, 269–303. [Google Scholar]
- Gillanders, B.M. Seagrasses, fish, and fisheries. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Borum, J.; Sand-Jensen, K.; Binzer, T.; Pedersen, O. Oxygen movement in seagrasses. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Mateo, M.A.; Cebrián, J.; Dunton, K.; Mutchler, T. Carbon flux in seagrass ecosystems. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Marbà, N.; Holmer, M.; Gacia, E.; Barrón, C. Seagrass beds and coastal biogeochemistry. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Romero, J.; Lee, K.-S.; P´erez, M.; Mateo, M.A.; Alcoverro, T. Nutrient dynamics in seagrass ecosystems. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Sheppard, C.R.C.; Davy, S.K.; Pilling, G.M.; Graham, N.A.J. The Biology of Coral Reefs; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Wilkinson, C. Status of Coral Reefs of the World; Australian Institute of Marine Science: Townsville, Australia, 2004.
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.; Paredes, G.; et al. Global trajectories of the long-term decline of coral reef ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef]
- George, A.; Luckymis, M.; Palik, S.; Joseph, E.; Mathias, D.; Malakai, S.; Nakayama, M.R.; Graham, C.; Rikim, K.; Marcus, A.; et al. The State of Coral Reef Ecosystems of the Federated States of Micronesia; Secretariat of the Pacific Regional Environment Programme (SPREP): Apia, Samoa, 2007. [Google Scholar]
- Hill, J.; Wilkinson, C. Methods for Ecological Monitoring of Coral Reefs; Australian Institute of Marine Science: Townsville, Australia, 2004.
- Jonker, M.; Johns, K.; Osborne, K. Surveys of Benthic Reef Communities Using Underwater Digital Photography and Counts of Juvenile Corals; Australian Institute of Marine Science: Townsville, Australia, 2008.
- Littler, M.M.; Littler, D.S.; Brooks, B.L. Harmful algae on tropical coral reefs: Bottom-up eutrophication and top-down herbivory. Harmful Algae 2006, 5, 565–585. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Morrissey, J. Community structure and zonation of macroalgae and hermatypic corals on a fringing reef flat of Magnetic Island (Queensland, Australia). Aquat. Bot. 1980, 8, 91–139. [Google Scholar] [CrossRef]
- Reichelt, R.E.; Loya, Y.; Bradbury, R.H. Patterns in the use of space by benthic communities on two coral reefs of the Great Barrier Reef. Coral Reefs 1986, 5, 73–79. [Google Scholar] [CrossRef]
- Iryu, Y.; Nakamori, T.; Matsuda, S.; Abe, O. Distribution of marine organisms and its geological significance in the modern reef complex of the Ryukyu Islands. Sediment. Geol. 1995, 99, 243–258. [Google Scholar] [CrossRef]
- Nakamura, T.; Nakamori, T. Population dynamics of hermatypic coral communities on reef slope vs. reef flat at Shiraho, Ishigaki Island, southwest Japan. In Proceedings of the 10th International Coral Reef Symposium, Okinawa, Japan, 28 June–2 July 2004; pp. 460–477. [Google Scholar]
- Riegl, B.; Piller, W.E. Mapping of benthic habitats in northern Safaga Bay (Red Sea, Egypt): A tool for proactiUe management. Aquat. Conserv. Mar. Freshw. Ecosyst. 2000, 10, 127–140. [Google Scholar] [CrossRef]
- Mehrabi, Z.; Slade, E.M.; Solis, A.; Mann, D.J. The importance of microhabitat for biodiversity sampling. PLoS ONE 2014, 9, e114015. [Google Scholar] [CrossRef] [PubMed]
- Done, T.J. Coral zonation: Its nature and significance. In Perspectives on Coral Reefs; Barnes, D.J., Ed.; Australian Institute of Marine Science: Townsville, Australia, 1983; p. 277. [Google Scholar]
- Done, T.J. Coral community adaptability to environmental change at the scales of regions, reefs and reef zones. Am. Zool. 1999, 39, 66–79. [Google Scholar] [CrossRef]
- Riegl, B.; Piller, W.E. Distribution and Environmental Control of Coral Assemblages in Northern Safaga Bay (Red Sea, Egypt). Facies 1997, 36, 141–162. [Google Scholar] [CrossRef]
- Loya, Y.; Sakai, K.; Yamazato, K.; Nakano, Y.; Sambali, H.; van Woesik, R. Coral bleaching: The winners and the losers. Ecol. Lett. 2001, 4, 122–131. [Google Scholar] [CrossRef]
- Marshall, P.A.; Baird, A.H. Bleaching of corals on the Great Barrier Reef: Differential susceptibilities among taxa. Coral Reefs 2000, 19, 155–163. [Google Scholar] [CrossRef]
- Pichon, M. Porites. In Encyclopedia of Earth Sciences Series; Springer: Berlin/Heidelberg, Germany, 2011; pp. 653–655. [Google Scholar]
- Roelfsema, C.; Kovacs, E.; Ortiz, J.C.; Wolff, N.H.; Callaghan, D.; Wettle, M.; Ronan, M.; Hamylton, S.M.; Mumby, P.J.; Phinn, S. Coral reef habitat mapping: A combination of object-based image analysis and ecological modelling. Remote Sens. Environ. 2018, 208, 27–41. [Google Scholar] [CrossRef]
- Babcock, R.C.; Bull, G.D.; Harrison, P.L.; Heyward, A.J.; Oliver, J.K.; Wallace, C.C.; Willis, B.L. Synchronous spawnings of 105 scleractinian coral species on the Great Barrier Reef. Mar. Biol. 1986, 90, 379–394. [Google Scholar] [CrossRef]
- Connell, J.H.; Hughes, T.P.; Wallace, C.C.; Kerr, A.M. A long-term study of competition and diversity of corals. Ecol. Monogr. 2004, 74, 179–210. [Google Scholar] [CrossRef]
- Freeman, L.A.; Miller, A.J.; Norris, R.D.; Smith, J.E. Classification of remote Pacific coral reefs by physical oceanographic environment. J. Geophys. Res. Oceans 2012, 117, C02007. [Google Scholar] [CrossRef]
- Onuf, C.P. Seagrasses, dredging and light in Laguna Madre, Texas, U.S.A. Estuar. Coast. Shelf. Sci. 1994, 39, 75–91. [Google Scholar] [CrossRef]
- Duke, N.C. Mangrove islands. In Encyclopedia of Earth Sciences Series; Springer: Berlin/Heidelberg, Germany, 2011; pp. 653–655. [Google Scholar]
- Benayahu, Y.; Loya, Y. Competition for space among coral-reef sessile organisms at Eilat, Red Sea. Bull. Mar. Sci. 1981, 31, 514–522. [Google Scholar]
- Done, T.J. Phase shifts in coral reef communities and their ecological significance. Hydrobiologia 1992, 247, 121–132. [Google Scholar] [CrossRef]
- Chadwick, N.E.; Morrow, K.M. Competition among sessile organisms on coral reefs. In Coral Reefs: An Ecosystem in Transition; Springer: Berlin/Heidelberg, Germany, 2011; pp. 347–371. [Google Scholar]
- Diaz, R.J.; Solan, M.; Valente, R.M. A review of approaches for classifying benthic habitats and evaluating habitat quality. J. Environ. Manag. 2004, 73, 165–181. [Google Scholar] [CrossRef]
- Roelfsema, C.M.; Kovacs, E.M.; Ortiz, J.C.; Callaghan, D.P.; Hock, K.; Mongin, M.; Johansen, K.; Mumby, P.J.; Wettle, M.; Ronan, M.; et al. Habitat maps to enhance monitoring and management of the Great Barrier Reef. Coral Reefs 2020, 39, 1039–1054. [Google Scholar] [CrossRef]
- Roelfsema, C.M.; Lyons, M.B.; Castro-Sanguino, C.; Kovacs, E.M.; Callaghan, D.; Wettle, M.; Markey, K.; Borrego-Acevedo, R.; Tudman, P.; Roe, M.; et al. How Much Shallow Coral Habitat Is There on the Great Barrier Reef? Remote Sens. 2021, 13, 4343. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Type | Classification | Taxon | Morphology |
|---|---|---|---|---|
| Abiotic | Sand | - | - | - |
| Rubble | - | - | - | |
| Dead coral | - | - | - | |
| Rock | - | - | - | |
| Coral | Hard coral | Acropora (Cnidaria: Anthozoa: Hexacorallia: Scleractinia: Acroporidae) | Acropora millepora (Ehrenberg, 1834) Acropora monticulosa (Brüggemann, 1879) Acropora nobilis (Dana, 1846) Acropora sp. | Bottlebrush Branching Digitate Tabulate Encrusting Submassive |
| Non-Acropora | Fungiidae spp. Montipora spp. Pavona spp. Pocillopora spp. Porites spp. Porites cylindrica Dana, 1846 | Branching Encrusting Foliaceous Massive Submassive Mushroom Solitary | ||
| Soft coral | Cnidaria: Anthozoa: Octocorallia: Alcyonacea | Alcyonium sp. Sinularia sp. | - | |
| Algae | Macroalgae | Brown macroalgae (Phaeophyta) | Dictyota spp. Padina spp. Sargassum spp. Turbinaria spp. | - |
| Red macroalgae (Rhodophyta) | Actinotrichia spp. | - | ||
| Green macroalgae (Chlorophyta) | Bornetella spp. Caulerpa spp. Codium spp. | - | ||
| Coralline algae | Rhodophyta: Corallinales | - | - | |
| Turf algae | Microalgae, Juvenile macroalgae, Cyanobacteria | - | - | |
| Halimeda | Chlorophyta: Ulvophyceae | Halimeda spp. | - | |
| Seagrass | Angiospermae: Alismatales | Thalassia hemprichii Ascherson, 1871 Cymodocea rotundata Ascherson & Schweinfurt, 1870 Syringodium isoetifolium Dandy, 1939 Enhalus acoroides Royle, 1839 | - | |
| Other | Macrofauna | Porifera | - | - |
| Tunicata | - | |||
| Echinodermata | - | |||
| Arthropoda | - | |||
| Mollusca | - | |||
| Cnidaria: Anthozoa: Hexacorallia: Zoantharia | - | |||
| Cnidaria: Hydrozoa: Milleporidae | Millepora spp. | |||
| Cnidaria: Anthozoa: Octocorallia: Helioporacea | Heliopora coerulea (Pallas, 1766) | |||
| Indeterminate | Indeterminate | - | - | - |
| Type | Classification | Taxon | Total (%) | TL1 (%) | TL2 (%) | TL3 (%) |
|---|---|---|---|---|---|---|
| Sand (S) | - | - | 42.4 | 32.2 | 45.5 | 41.8 |
| Rubble (R) | - | - | 8.9 | 6.8 | 10.4 | 7.3 |
| Dead coral (DC) | - | - | 1.5 | 2.6 | 1.7 | 0.5 |
| Rock (RK) | - | - | - | - | - | - |
| Hard coral (HC) | Cnidaria: Anthozoa: Hexacorallia: Scleractinia | Scleractinia total | 3.5 | 6.0 | 4.0 | 1.4 |
| Acropora | Acropora spp. total | 1.5 | 5.1 | 1.2 | 0.1 | |
| Acropora millepora | 0.2 | - | 0.3 | - | ||
| Acropora monticulosa | - | - | 0.1 | - | ||
| Acropora nobilis | 0.7 | 2.7 | 0.5 | 0.1 | ||
| Acropora sp. | 0.6 | 2.4 | 0.4 | - | ||
| Non-Acropora | Non-Acropora spp. total | 2.0 | 0.9 | 2.7 | 1.3 | |
| Fungiidae spp. | - | - | - | - | ||
| Montipora spp. | 0.1 | - | 0.1 | - | ||
| Pavona spp. | 0.2 | - | 0.4 | - | ||
| Pocillopora spp. | 0.2 | 0.3 | 0.2 | - | ||
| Porites cylindrica | 1.2 | - | 1.7 | 0.8 | ||
| Porites spp. | 0.4 | 0.5 | 0.3 | 0.5 | ||
| Soft coral (SC) | Cnidaria: Anthozoa: Octocorallia: Alcyonacea | Alcyonacea total | 2.6 | - | 3.1 | 2.8 |
| Alcyonium spp. | - | - | 0.1 | - | ||
| Sinularia spp. | 2.5 | - | 3.0 | 2.8 | ||
| Macroalgae (MA) | Macroalgae total | - | 2.7 | 3.6 | 2.9 | 1.7 |
| Brown macroalgae (Phaeophyta) | Phaeophyta total | 2.5 | 3.4 | 2.7 | 1.7 | |
| Dictyota spp. | 0.8 | 0.8 | 0.9 | 0.4 | ||
| Padina spp. | 1.3 | 0.5 | 1.8 | 0.8 | ||
| Sargassum spp. | 0.3 | 1.9 | - | - | ||
| Turbinaria spp. | 0.2 | 0.2 | - | 0.5 | ||
| Green macroalgae (Chlorophyta) | Chlorophyta total | 0.1 | 0.1 | 0.1 | - | |
| Bornetella spp. | - | 0.1 | - | - | ||
| Caulerpa spp. | 0.1 | - | 0.1 | - | ||
| Codium spp. | - | - | - | - | ||
| Red macroalgae (Rhodophyta) | Rhodophyta total | - | - | - | 0.1 | |
| Actinotrichia spp. | - | - | - | 0.1 | ||
| Coralline algae (CA) | Rhodophyta: Corallinales | Corallinales total | 1.4 | 0.9 | 1.9 | 0.7 |
| Turf algae (TA) | Microalgae, juvenile macroalgae, Cyanobacteria | Turf algae total | 6.0 | 3.1 | 8.1 | 3.4 |
| Halimeda (HA) | Chlorophyta: Ulvophyceae | Halimeda spp. total | 4.9 | 7.5 | 5.2 | 2.9 |
| Seagrass (SG) | Angiospermae: Alismatales | Alismatales total | 26.1 | 37.2 | 17.1 | 37.4 |
| Thalassia hemprichii | NA | NA | NA | NA | ||
| Cymodocea rotundata | NA | NA | NA | NA | ||
| Syringodium isoetifolium | NA | NA | NA | NA | ||
| Enhalus acoroides | NA | NA | NA | NA | ||
| Macrofauna (MF) | Macrofauna total | - | 0.1 | 0.1 | 0.1 | - |
| Cnidaria: Hydrozoa: Milleporidae | Millepora spp. | - | - | - | - | |
| Cnidaria: Anthozoa: Octocorallia: Helioporacea | Heliopora coerulea | - | - | - | - | |
| Porifera | - | 0.1 | 0.1 | 0.1 | - | |
| Tunicata | - | - | - | - | - | |
| Cnidaria: Anthozoa: Hexacorallia: Zoantharia | - | - | - | - | - | |
| Echinodermata | - | - | - | - | - | |
| Mollusca | - | - | - | - | - | |
| Arthropoda | - | - | - | - | - | |
| Indeterminate (IN) | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.; Lee, D.-W.; Kim, H.-J.; Jung, Y.-H.; Choi, Y.-U.; Oh, J.-H.; Kim, T.-H.; Kang, D.-H.; Park, H.-S. Estimation of the Benthic Habitat Zonation by Photo-Quadrat Image Analysis along the Fringing Reef of Weno Island, Chuuk, Micronesia. J. Mar. Sci. Eng. 2022, 10, 1643. https://doi.org/10.3390/jmse10111643
Kim T, Lee D-W, Kim H-J, Jung Y-H, Choi Y-U, Oh J-H, Kim T-H, Kang D-H, Park H-S. Estimation of the Benthic Habitat Zonation by Photo-Quadrat Image Analysis along the Fringing Reef of Weno Island, Chuuk, Micronesia. Journal of Marine Science and Engineering. 2022; 10(11):1643. https://doi.org/10.3390/jmse10111643
Chicago/Turabian StyleKim, Taihun, Dae-Won Lee, Han-Jun Kim, Yun-Hwan Jung, Young-Ung Choi, Jung-Hee Oh, Tae-Hoon Kim, Do-Hyung Kang, and Heung-Sik Park. 2022. "Estimation of the Benthic Habitat Zonation by Photo-Quadrat Image Analysis along the Fringing Reef of Weno Island, Chuuk, Micronesia" Journal of Marine Science and Engineering 10, no. 11: 1643. https://doi.org/10.3390/jmse10111643
APA StyleKim, T., Lee, D.-W., Kim, H.-J., Jung, Y.-H., Choi, Y.-U., Oh, J.-H., Kim, T.-H., Kang, D.-H., & Park, H.-S. (2022). Estimation of the Benthic Habitat Zonation by Photo-Quadrat Image Analysis along the Fringing Reef of Weno Island, Chuuk, Micronesia. Journal of Marine Science and Engineering, 10(11), 1643. https://doi.org/10.3390/jmse10111643