Color for Life: Biosynthesis and Distribution of Phenolic Compounds in Pepper (Capsicum annuum)



Abstract
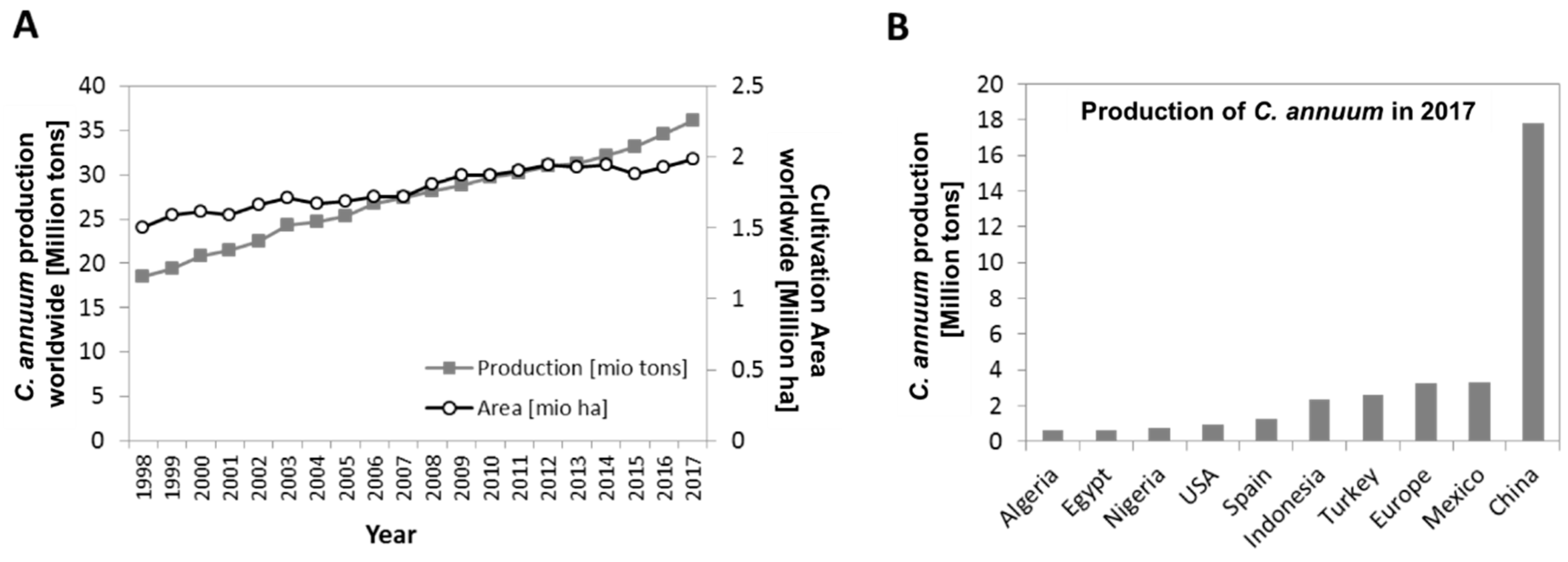
:1. Secondary Metabolites from Fruits and Vegetables are of High Value for Human Nutrition and Industrial Applications
2. Phenolic Compounds—Structure and Biosynthesis
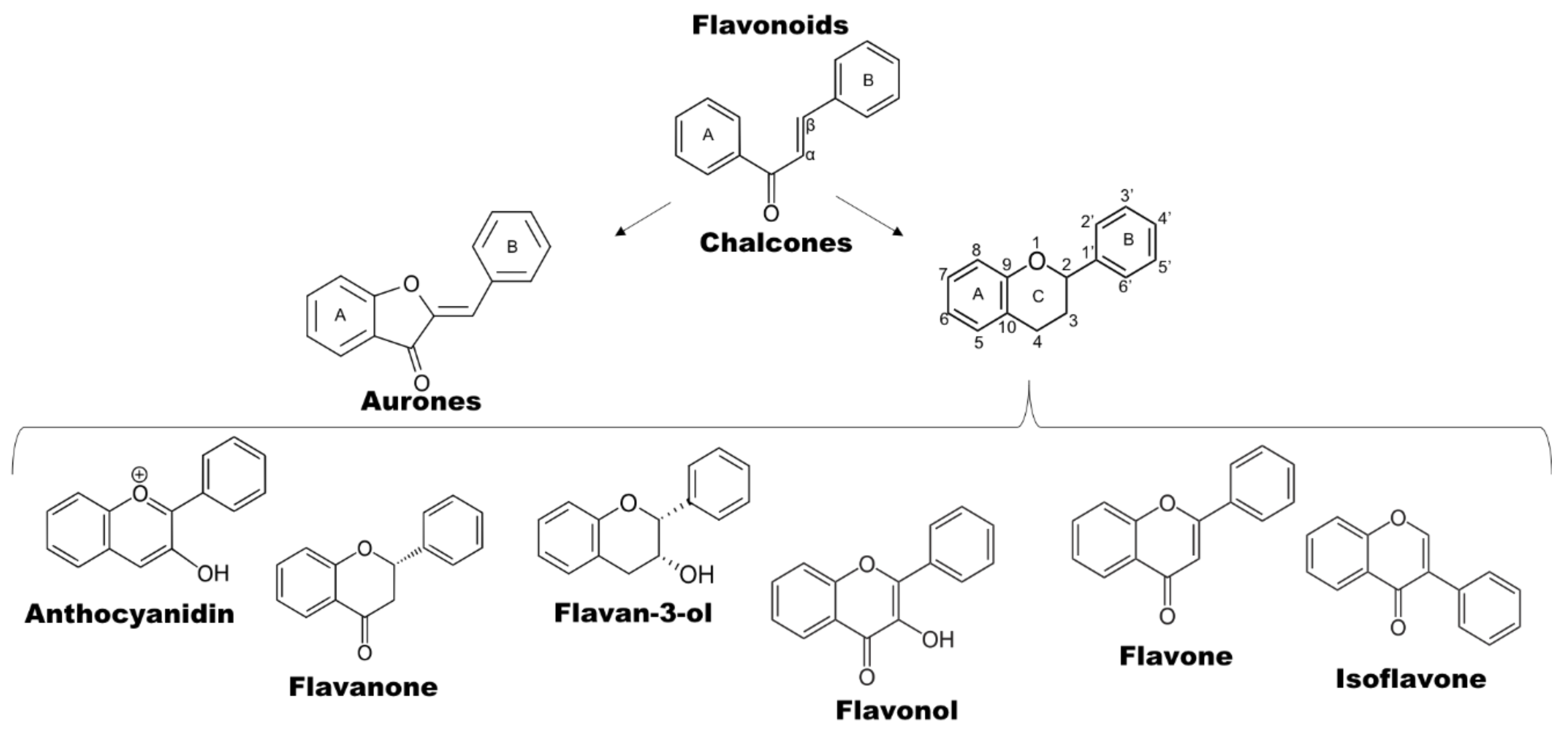
2.1. Definition and Structure of Phenolic Compounds
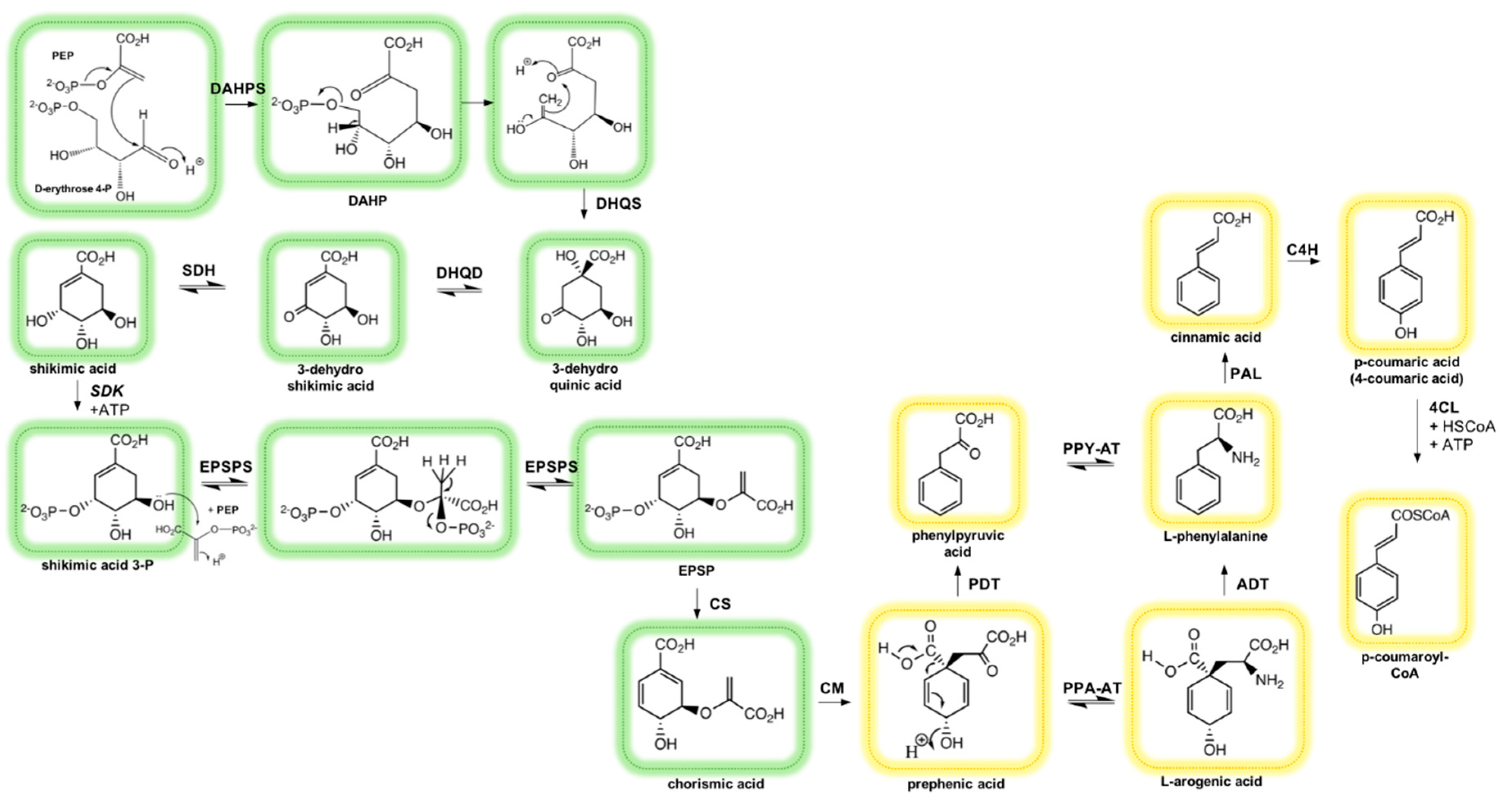
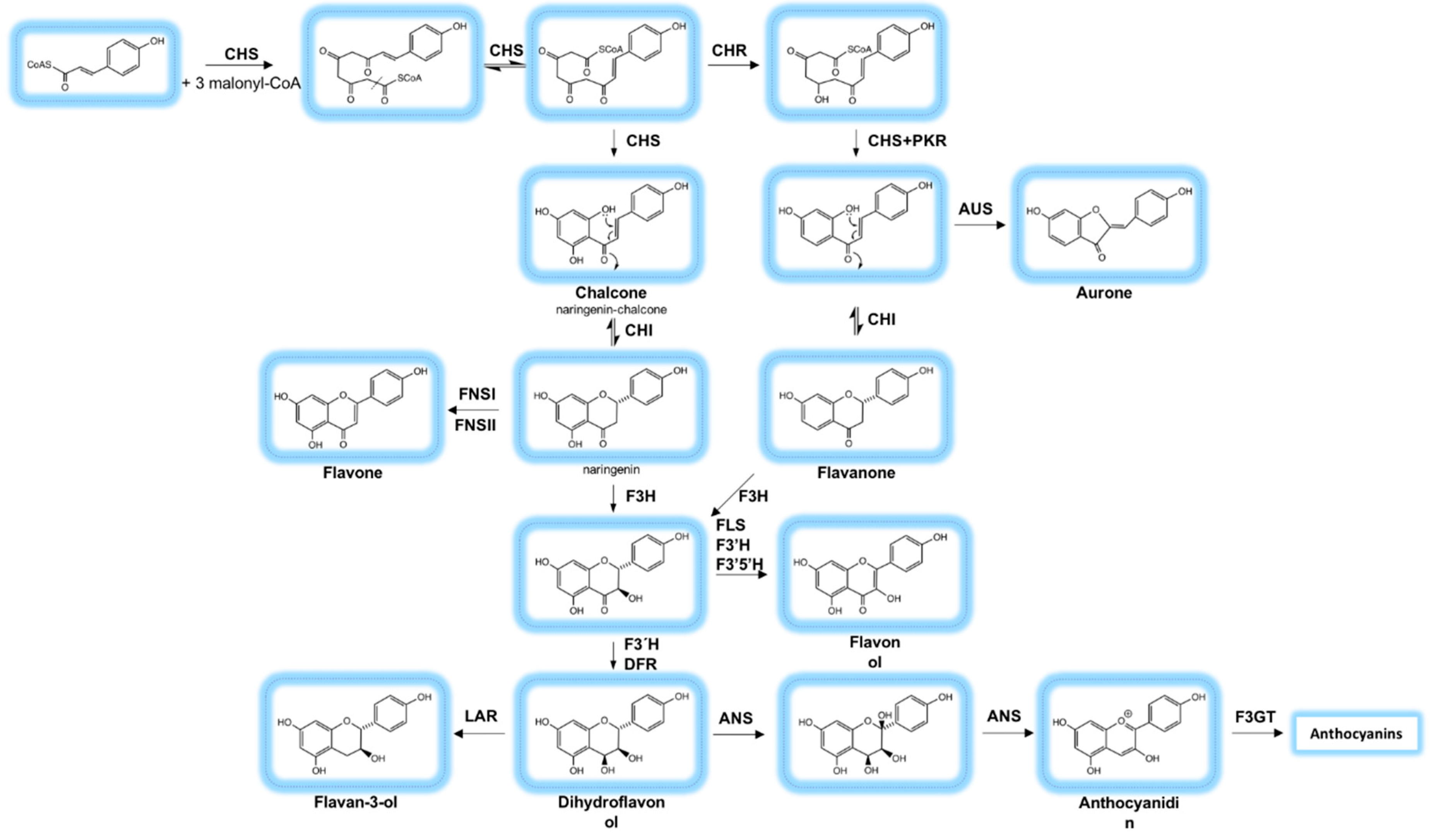
2.2. Biosynthesis of Phenolic Compounds
2.3. Genes and Regulation of the Biosynthesis of Phenolic Compounds in General
3. Phenolic Composition of Capsicum Annuum
3.1. Preparation and Measurement of Phenolic Compounds from Plants
3.2. Total Phenolic and Flavonoid Content in Capsicum
3.3. Quantification of Flavonoids in Capsicum
4. Possible Strategies to Increase Phenolic Compounds in Capsicum annuum
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organisation (WHO). Available online: https://www.who.int/news-room/fact-sheets/detail/healthy-diet (accessed on 29 March 2019).
- 2015–2020 Dietary Guidelines. Available online: https://health.gov/dietaryguidelines/2015/guidelines/ (accessed on 29 March 2019).
- Food and Agriculture Organization of the United Nations | Food-Based Dietary Guidelines. Available online: http://www.fao.org/nutrition/education/food-dietary-guidelines/regions/europe/en/ (accessed on 29 March 2019).
- Wu, S.; Chappell, J. Metabolic engineering of natural products in plants; tools of the trade and challenges for the future. Curr. Opin. Biotechnol. 2008, 19, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Li, J.; Ou-Lee, T.M.; Raba, R.; Amundson, R.G.; Last, R.L. Arabidopsis flavonoid mutants are hypersensitive to UV-B irradiation. Plant Cell 1993, 5, 171–179. [Google Scholar] [CrossRef]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defense mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Wink, M. Importance of plant secondary metabolites for protection against insects and microbial infections. Adv. Phytomed. 2006, 3, 251–268. [Google Scholar] [CrossRef]
- Wittstock, U.; Gershenzon, J. Constitutive plant toxins and their role in defense against herbivores and pathogens. Curr. Opin. Plant Biol. 2002, 5, 300–307. [Google Scholar] [CrossRef]
- López-Goldar, X.; Villari, C.; Bonello, P.; Borg-Karlson, A.K.; Grivet, D.; Zas, R.; Sampedro, L. Inducibility of plant secondary metabolites in the stem predicts genetic variation in resistance against a key insect herbivore in maritime pine. Front. Plant Sci. 2018, 9, 1651. [Google Scholar] [CrossRef]
- Schreiner, M.; Mewis, I.; Huyskens-Keil, S.; Jansen, M.A.K.; Zrenner, R.; Winkler, J.B.; O’Brien, N.; Krumbein, A. UV-B-induced secondary plant metabolites - Potential benefits for plant and human health. CRC. Crit. Rev. Plant Sci. 2012, 31, 229–240. [Google Scholar] [CrossRef]
- Tohge, T.; Wendenburg, R.; Ishihara, H.; Nakabayashi, R.; Watanabe, M.; Sulpice, R.; Hoefgen, R.; Takayama, H.; Saito, K.; Stitt, M.; et al. Characterization of a recently evolved flavonol-phenylacyltransferase gene provides signatures of natural light selection in Brassicaceae. Nat. Commun. 2016, 7, 12399. [Google Scholar] [CrossRef]
- Stevenson, P.C.; Nicolson, S.W.; Wright, G.A. Plant secondary metabolites in nectar: Impacts on pollinators and ecological functions. Funct. Ecol. 2017, 31, 65–75. [Google Scholar] [CrossRef]
- Harborne, J.B. Secondary Metabolites: Attracting Pollinators. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2011. [Google Scholar] [CrossRef]
- Wink, M. Plant secondary metabolites modulate insect behavior-steps toward addiction? Front. Physiol. 2018, 9, 364. [Google Scholar] [CrossRef]
- Crozier, A.; Clifford, M.N.; Ashihara, H. Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; Blackwell: Oxford, UK, 2006; ISBN 9780470988558. [Google Scholar]
- Fernandes, I.; Pérez-Gregorio, R.; Soares, S.; Mateus, N.; de Freitas, V.; Fernandes, I.; Pérez-Gregorio, R.; Soares, S.; Mateus, N.; De Freitas, V. Wine flavonoids in health and disease prevention. Molecules 2017, 22, 292. [Google Scholar] [CrossRef] [PubMed]
- Silva-Beltrán, N.P.; Ruiz-Cruz, S.; Chaidez, C.; Ornelas-Paz, J.d.J.; López-Mata, M.A.; Márquez-Ríos, E.; Estrada, M.I. Chemical constitution and effect of extracts of tomato plants byproducts on the enteric viral surrogates. Int. J. Environ. Health Res. 2015, 25, 299–311. [Google Scholar] [CrossRef]
- Taveira, M.; Silva, L.R.; Vale-Silva, L.A.; Pinto, E.; Valentão, P.; Ferreres, F.; Guedes de Pinho, P.; Andrade, P.B. Lycopersicon esculentum seeds: An industrial byproduct as an antimicrobial agent. J. Agric. Food Chem. 2010, 58, 9529–9536. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, P. Phenolic compounds of cereals and their antioxidant capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Ahmad, B.; Bawazeer, S.; Atif, M.; Peters, D.G.; et al. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phyther. Res. 2019, 33, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.; Novellino, E.; et al. The therapeutic potential of apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J.; Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; et al. The therapeutic potential of naringenin: A review of clinical trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.I.; Jain, P.; Khan, Z.K. Potential role of flavonoids in treating chronic inflammatory diseases with a special focus on the anti-inflammatory activity of apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Serino, A.; Salazar, G.; Serino, A.; Salazar, G. Protective role of polyphenols against vascular inflammation, aging and cardiovascular disease. Nutrients 2018, 11, 53. [Google Scholar] [CrossRef]
- Krga, I.; Milenkovic, D. Anthocyanins: From sources and bioavailability to cardiovascular-health benefits and molecular mechanisms of action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, Y.; Rasool, S.; Geetha, T.; Babu, J.R. Effects and underlying mechanisms of bioactive compounds on type 2 diabetes mellitus and Alzheimer’s disease. Oxid. Med. Cell. Longev. 2019, 2019, 1–25. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Caltagirone, S.; Rossi, C.; Poggi, A.; Ranelletti, F.O.; Natali, P.G.; Brunetti, M.; Aiello, F.B.; Piantelli, M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int. J. Cancer 2000, 87, 595–600. [Google Scholar] [CrossRef]
- Oksman-Caldentey, K.M.; Inzé, D. Plant cell factories in the post-genomic era: New ways to produce designer secondary metabolites. Trends Plant Sci. 2004, 9, 433–440. [Google Scholar] [CrossRef] [PubMed]
- McChesney, J.D.; Venkataraman, S.K.; Henri, J.T. Plant natural products: Back to the future or into extinction? Phytochemistry 2007, 68, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Farhoudi, R.; Mehrnia, M.A.; Lee, D.-J. Antioxidant activities and bioactive compounds of five Jalopeno peppers (Capsicum annuum) cultivars. Nat. Prod. Res. 2017. [Google Scholar] [CrossRef]
- Bosland, P.W.; Votava, E.J. Peppers: Vegetable and Spice Capsicums; CABI: Oxfordshire, UK, 2012; ISBN 178064020X. [Google Scholar]
- Wahyuni, Y.; Ballester, A.-R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Secondary metabolites of Capsicum species and their importance in the human diet. J. Nat. Prod. 2013, 76, 783–793. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: http://faostat3.fao.org/home/E (accessed on 13 March 2015).
- Salvatore, S.; Pellegrini, N.; Serafini, M.; Del Rio, D.; Brighenti, F.; Bianchi, M.; Colombi, B. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr. 2018, 133, 2812–2819. [Google Scholar] [CrossRef]
- Zimmer, A.R.; Leonardi, B.; Miron, D.; Schapoval, E.; Oliveira, J.R. De Gosmann, G. Antioxidant and anti-inflammatory properties of Capsicum baccatum: From traditional use to scientific approach. J. Ethnopharmacol. 2012, 139, 228–233. [Google Scholar] [CrossRef]
- Morales-Soto, A.; García-Salas, P.; Rodríguez-Pérez, C.; Jiménez-Sánchez, C.; de la Luz Cádiz-Gurrea, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Antioxidant capacity of 44 cultivars of fruits and vegetables grown in Andalusia (Spain). Food Res. Int. 2014, 58, 35–46. [Google Scholar] [CrossRef]
- Olatunji, T.L.; Afolayan, A.J. The suitability of chili pepper (Capsicum annuum L.) for alleviating human micronutrient dietary deficiencies: A review. Food Sci. Nutr. 2018, 6, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Kantar, M.B.; Anderson, J.E.; Lucht, S.A.; Mercer, K.; Bernau, V.; Case, K.A.; Le, N.C.; Frederiksen, M.K.; DeKeyser, H.C.; Wong, Z.-Z.; et al. Vitamin variation in Capsicum spp. provides opportunities to improve nutritional value of human diets. PLoS ONE 2016, 11, e0161464. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Serrani-Yarce, J.C.; Chen, F.; Baxter, D.; Venables, B.J.; Dixon, R.A. Role of bifunctional ammonia-lyase in grass cell wall biosynthesis. Nat. Plants 2016, 2, 16050. [Google Scholar] [CrossRef] [PubMed]
- Vermerris, W.; Nicholson, R. Phenolic Compound Biochemistry; Springer: Berlin, Germany, 2008; ISBN 978-1-4020-5163-0. [Google Scholar]
- Vukics, V.; Guttman, A. Structural characterization of flavonoid glycosides by multi-stage mass spectrometry. Mass Spectrom. Rev. 2010, 29, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Belitz, H.D.; Grosch, W.; Schieberle, P. Food Chemistry; Springer-Verlag: Berlin/Heidelberg, Germany, 2009; ISBN 9783540699330. [Google Scholar]
- Dewick, P.M. The shikimic pathway: Aromatic aminoacids and phenylpropanoids. In Medicinal Natural Products: A Biosynthetic Approach; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; ISBN 9780470741689. [Google Scholar]
- Nishiumi, S.; Miyamoto, S.; Kawabata, K.; Ohnishi, K.; Mukai, R.; Murakami, A.; Ashida, H.; Terao, J. Dietary flavonoids as cancer-preventive and therapeutic biofactors. Front. Biosci. 2011, 3, 1332–1362. [Google Scholar] [CrossRef]
- Santos, E.L.; Maia, B.H.L.N.S.; Ferriani, A.P.; Teixeira, S.D. Flavonoids: Classification, Biosynthesis and Chemical Ecology. In Flavonoids—From Biosynthesis to Human Health; IntechOpen Limited: London, UK, 2017; ISBN 978-953-51-3424-4. [Google Scholar]
- Hui, Y.H.; Nip, W.K.; Nollet, L.M.L.; Paliyath, G.; Simpson, B.K. Food Biochemistry and Food Processing; Blackwell: Oxford, UK, 2007; ISBN 0813803780. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products. A Biosynthetic Approach, 2nd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; ISBN 9780470846278. [Google Scholar]
- Andersen, O.M.; Markham, K.R. Flavonoids. Chemistry, Biochemistry and Applications; CRC Press: Boca Raton, FL, USA, 2006; ISBN 0-8493-2021-6. [Google Scholar]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Bomati, E.K.; Austin, M.B.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structural elucidation of chalcone reductase and implications for deoxychalcone biosynthesis. J. Biol. Chem. 2005, 280, 30496–30503. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The flavonoid biosynthetic pathway in Arabidopsis: Structural and genetic diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Xie, D.-Y.; Sharma, S.B. Proanthocyanidins—A final frontier in flavonoid research? New Phytol. 2004, 165, 9–28. [Google Scholar] [CrossRef]
- Abrahams, S.; Lee, E.; Walker, A.R.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. The Arabidopsis TDS4 gene encodes leucoanthocyanidin dioxygenase (LDOX) and is essential for proanthocyanidin synthesis and vacuole development. Plant J. 2003, 35, 624–636. [Google Scholar] [CrossRef]
- Austin, M.B.; Noel, J.P. The chalcone synthase superfamily of type III polyketide synthases. Nat. Prod. Rep. 2003, 20, 79–110. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2002, 126, 485–493. [Google Scholar] [CrossRef]
- Turnbull, J.J.; Nakajima, J.I.; Welford, R.W.D.; Yamazaki, M.; Saito, K.; Schofield, C.J. Mechanistic studies on three 2-oxoglutarate-dependent oxygenases of flavonoid biosynthesis: Anthocyanidin synthase, flavonol synthase, and flavanone 3β-hydroxylase. J. Biol. Chem. 2004, 279, 1206–1216. [Google Scholar] [CrossRef] [PubMed]
- Schoenbohm, C.; Martens, S.; Eder, C.; Forkmann, G.; Weisshaar, B. Identification of the Arabidopsis thaliana flavonoid 3′-hydroxylase gene and functional expression of the encoded P450 enzyme. Biol. Chem. 2000, 381, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Cone, K.C.; Burr, F.A.; Burr, B. Molecular analysis of the maize anthocyanin regulatory locus C1. Proc. Natl. Acad. Sci. USA 1986, 83, 9631–9635. [Google Scholar] [CrossRef] [PubMed]
- Larson, R.; Bussard, J.B.; Coe, E.H. Gene-dependent flavonoid 3′-hydroxylation in maize. Biochem. Genet. 1986, 24, 615–624. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, S.D.; Tong, Y.; Spörlein, B.; Forkmann, G.; Yoder, J.I. Molecular genetic analysis of chalcone synthase in Lycopersicon esculentum and an anthocyanin-deficient mutant. Mol. Gen. Genet. 1990, 224, 279–288. [Google Scholar] [CrossRef]
- Kang, J.-H.; Mcroberts, J.; Shi, F.; Moreno, J.E.; Jones, A.D.; Howe, G.A. The flavonoid biosynthetic enzyme chalcone isomerase modulates terpenoid production in glandular trichomes of tomato. Plant Physiol. 2014, 164, 1161–1174. [Google Scholar] [CrossRef]
- Tohge, T.; Zhang, Y.; Peterek, S.; Matros, A.; Rallapalli, G.; Tandrõn, Y.A.; Butelli, E.; Kallam, K.; Hertkorn, N.; Mock, H.P.; et al. Ectopic expression of snapdragon transcription factors facilitates the identification of genes encoding enzymes of anthocyanin decoration in tomato. Plant J. 2015, 83, 686–704. [Google Scholar] [CrossRef]
- Zhang, Y.; Butelli, E.; Alseekh, S.; Tohge, T.; Rallapalli, G.; Luo, J.; Kawar, P.G.; Hill, L.; Santino, A.; Fernie, A.R.; et al. Multi-level engineering facilitates the production of phenylpropanoid compounds in tomato. Nat. Commun. 2015, 26, 8635. [Google Scholar] [CrossRef] [PubMed]
- Burbulis, I.E.; Winkel-Shirley, B. Interactions among enzymes of the Arabidopsis flavonoid biosynthetic pathway. Proc. Natl. Acad. Sci. USA 2002, 96, 12929–12934. [Google Scholar] [CrossRef]
- Eloy, N.B.; Voorend, W.; Lan, W.; Saleme, M.D.L.S.; Cesarino, I.; Vanholme, R.; Smith, R.A.; Goeminne, G.; Pallidis, A.; Morreel, K.; et al. Silencing CHALCONE SYNTHASE in maize impedes the incorporation of tricin into lignin and increases lignin content. Plant Physiol. 2017, 173, 998–1016. [Google Scholar] [CrossRef]
- Bongue-Bartelsman, M.; O’Neill, S.D.; Yusen, T.; Yoder, J.I. Characterization of the gene encoding dihydroflavonol 4-reductase in tomato. Gene 1994, 138, 153–157. [Google Scholar] [CrossRef]
- Ralston, E.J.; English, J.J.; Dooner, H.K. Sequence of three bronze alleles of maize and correlation with the genetic fine structure. Genetics 1988, 119, 185–197. [Google Scholar]
- Devic, M.; Guilleminot, J.; Debeaujon, I.; Bechtold, N.; Bensaude, E.; Koornneef, M.; Pelletier, G.; Delseny, M. The BANYULS gene encodes a DFR-like protein and is a marker of early seed coat development. Plant J. 1999, 19, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.-M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Nesi, N.; Jond, C.; Debeaujon, I.; Caboche, M.; Lepiniec, L. The Arabidopsis TT2 gene encodes an R2R3 MYB domain protein that acts as a key determinant for proanthocyanidin accumulation in developing seed. Plant Cell 2001, 13, 2099–2114. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 Repeat protein. Plant Cell Online 1999, 11, 1337–1350. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.; Robatzek, S.; Somssich, I. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Gonzalez, A.; Brown, M.; Hatlestad, G.; Akhavan, N.; Smith, T.; Hembd, A.; Moore, J.; Montes, D.; Mosley, T.; Resendez, J.; et al. TTG2 controls the developmental regulation of seed coat tannins in Arabidopsis by regulating vacuolar transport steps in the proanthocyanidin pathway. Dev. Biol. 2016, 419, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Barragán, A.; Ochoa-Alejo, N. Virus-induced silencing of MYB and WD40 transcription factor genes affects the accumulation of anthocyanins in chili pepper fruit. Biol. Plant. 2014, 58, 567–574. [Google Scholar] [CrossRef]
- Aza-González, C.; Herrera-Isidrón, L.; Núñez-Palenius, H.G.; Martínez De La Vega, O.; Ochoa-Alejo, N. Anthocyanin accumulation and expression analysis of biosynthesis-related genes during chili pepper fruit development. Biol. Plant. 2013, 57, 49–55. [Google Scholar] [CrossRef]
- Stommel, J.R.; Lightbourn, G.J.; Winkel, B.S.; Griesbach, R.J. Transcription factor families regulate the anthocyanin biosynthetic pathway in Capsicum annuum. J. Am. Soc. Hortic. Sci. 2009, 134, 244–251. [Google Scholar] [CrossRef]
- Li, J.-G.; Li, H.-L.; Peng, S.-Q. Three R2R3 MYB transcription factor genes from Capsicum annuum showing differential expression during fruit ripening. African J. Biotechnol. 2011, 10. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.-W.; Jin, J.-H.; Yin, Y.-X.; Zhang, H.-X.; Chai, W.-G.; Gong, Z.-H. VIGS approach reveals the modulation of anthocyanin biosynthetic genes by CaMYB in chili pepper leaves. Front. Plant Sci. 2015, 6, 500. [Google Scholar] [CrossRef]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, Y.; Ballester, A.-R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef]
- Noreen, H.; Semmar, N.; Farman, M.; McCullagh, J.S.O. Measurement of total phenolic content and antioxidant activity of aerial parts of medicinal plant Coronopus didymus. Asian Pac. J. Trop. Med. 2017, 10, 792–801. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteau, V. Tyrosine and tryptophane determination in proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin-Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Contreras Guzman, E.S.; Strong F., C. Determination of tocopherols (vitamin e) by reduction of cupric ion. J. Assoc. Off. Anal. Chem. 1982, 65, 1215–1221. [Google Scholar]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76. [Google Scholar] [CrossRef]
- Stratil, P.; Klejdus, B.; Kubáň, V. Determination of total content of phenolic compounds and their antioxidant activity in vegetables—Evaluation of spectrophotometric methods. J. Agric. Food Chem. 2006, 54, 607–616. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and mechanisms of antioxidant activity using the DPPH• free radical method. LWT Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Butkovic, V.; Klasinc, L.; Bors, W. Kinetic study of flavonoid reactions with stable radicals. J. Agric. Food Chem. 2004, 52, 2816–2820. [Google Scholar] [CrossRef]
- Kim, D.O.; Lee, K.W.; Lee, H.J.; Lee, C.Y. Vitamin C equivalent antioxidant capacity (VCEAC) of phenolic phytochemicals. J. Agric. Food Chem. 2002, 50, 3713–3717. [Google Scholar] [CrossRef]
- Awika, J.M.; Rooney, L.W.; Wu, X.; Prior, R.L.; Cisneros-Zevallos, L. Screening methods to measure antioxidant activity of orghum (Sorghum bicolor) and Sorghum products. J. Agric. Food Chem. 2003, 51, 6657–6662. [Google Scholar] [CrossRef]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of antiradical properties of antioxidants using DPPH- assay: A critical review and results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Papariello, G.J.; Janish, M.A.M. Diphenylpicrylhydrazyl as an Organic Analytical Reagent in the Spectrophotometric Analysis of Phenols. Anal. Chem. 1966, 38, 211–214. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Wianowska, D.; Olszowy, M. On practical problems in estimation of antioxidant activity of compounds by DPPH method (Problems in estimation of antioxidant activity). Food Chem. 2012, 131, 1037–1043. [Google Scholar] [CrossRef]
- McGowan, J.C.; Powell, T.; Raw, R. The rates of reaction of αα-diphenyl-β-picrylhydrazyl with certain amines and phenols. J. Chem. Soc. 1959, 630, 3103–3110. [Google Scholar] [CrossRef]
- Xie, J.; Schaich, K.M. Re-evaluation of the 2,2-diphenyl-1-picrylhydrazyl free radical (DPPH) assay for antioxidant activity. J. Agric. Food Chem. 2014, 62, 4251–4260. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Huang, D.; Boxin, O.U.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Walker, R.B.; Everette, J.D. Comparative Reaction Rates of Various Antioxidants with ABTS Radical Cation. J. Agric. Food Chem. 2009, 57, 1156–1161. [Google Scholar] [CrossRef]
- Chandra, S.; Khan, S.; Avula, B.; Lata, H.; Yang, M.H.; Elsohly, M.A.; Khan, I.A. Assessment of total phenolic and flavonoid content, antioxidant properties, and yield of aeroponically and conventionally grown leafy vegetables and fruit crops: A comparative study. Evid. Based. Complement. Altern. Med. 2014, 2014, 253875. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Ahn, J.; Lee, S.J.; Moon, B.K.; Ha, T.Y.; Kim, S. Phytochemicals and Antioxidant Activity of Fruits and Leaves of Paprika (Capsicum annuum L., var. Special) Cultivated in Korea. J. Food Sci. 2011, 76, C193–C198. [Google Scholar] [CrossRef]
- Marín, A.; Ferreres, F.; Tomás-Barberán, F.A.; Gil, M.I. Characterization and quantitation of antioxidant constituents of sweet pepper (Capsicum annuum L.). J. Agric. Food Chem. 2004, 52, 3861–3869. [Google Scholar] [CrossRef]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Morales-Soto, A.; Gómez-Caravaca, A.M.; García-Salas, P.; Segura-Carretero, A.; Fernández-Gutiérrez, A. High-performance liquid chromatography coupled to diode array and electrospray time-of-flight mass spectrometry detectors for a comprehensive characterization of phenolic and other polar compounds in three pepper (Capsicum annuum L.) samples. Food Res. Int. 2013, 51, 977–984. [Google Scholar] [CrossRef]
- Medina-Juárez, L.Á.; Molina-Quijada, D.M.A.; Del Toro-Sánchez, C.L.; González-Aguilar, G.A.; Gámez-Meza, N. Antioxidant activity of peppers (Capsicum annum L.) extracts and characterization of their phenolic constituents. Interciencia 2012, 37, 588–593. [Google Scholar]
- Chen, L.; Kang, Y.-H. Anti-inflammatory and antioxidant activities of red pepper (Capsicum annuum L.) stalk extracts: Comparison of pericarp and placenta extracts. J. Funct. Foods 2013, 5, 1724–1731. [Google Scholar] [CrossRef]
- Mudrić, S.; Gašić, U.M.; Dramićanin, A.M.; Ćirić, I.Ž.; Milojković-Opsenica, D.M.; Popović-Đorđević, J.B.; Momirović, N.M.; Tešić, Ž.L. The polyphenolics and carbohydrates as indicators of botanical and geographical origin of Serbian autochthonous clones of red spice paprika. Food Chem. 2017, 217, 705–715. [Google Scholar] [CrossRef]
- Kim, W.-R.; Kim, E.O.; Kang, K.; Oidovsambuu, S.; Jung, S.H.; Kim, B.S.; Nho, C.W.; Um, B.-H. Antioxidant activity of phenolics in leaves of three red pepper (Capsicum annuum) cultivars. J. Agric. Food Chem. 2014, 62, 850–859. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Horibata, Y.; Mekuria, D.B.; Tebayashi, S.-I.; Kim, C.-S. Ovipositional deterrent in the sweet pepper, Capsicum annuum, at the mature stage against Liriomyza trifolii (Burgess). Biosci. Biotechnol. Biochem. 2005, 69, 1831–1835. [Google Scholar] [CrossRef]
- Del Rocio Moreno-Ramírez, Y.; Martínez-Ávila, G.C.G.; González-Hernández, V.A.; Castro-López, C.; Torres-Castillo, J.A. Free radical-scavenging capacities, phenolics and capsaicinoids in wild piquin chili (Capsicum annuum var. glabriusculum). Molecules 2018, 23, 2655. [Google Scholar] [CrossRef] [PubMed]
- Ogunruku, O.O.; Oboh, G.; Passamonti, S.; Tramer, F.; Boligon, A.A. Capsicum annuum var. grossum (Bell Pepper) Inhibits β-Secretase Activity and β-Amyloid 1–40 Aggregation. J. Med. Food 2017, 20, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, M.; Soukup, J.; Donato, P.; Cacciola, F.; Dugo, P.; Riazi, A.; Jandera, P.; Mondello, L. Determination of the polyphenolic content of a Capsicum annuum L. extract by liquid chromatography coupled to photodiode array and mass spectrometry detection and evaluation of its biological activity. J. Sep. Sci. 2015, 38, 171–178. [Google Scholar] [CrossRef]
- Park, S.; Jeong, W.Y.; Lee, J.H.; Kim, Y.H.; Jeong, S.W.; Kim, G.S.; Bae, D.W.; Lim, C.S.; Jin, J.S.; Lee, S.J.; et al. Determination of polyphenol levels variation in Capsicum annuum L. cv. Chelsea (yellow bell pepper) infected by anthracnose (Colletotrichum gloeosporioides) using liquid chromatography-tandem mass spectrometry. Food Chem. 2012, 130, 981–985. [Google Scholar] [CrossRef]
- Jeong, W.Y.; Jin, J.S.; Cho, Y.A.; Lee, J.H.; Park, S.; Jeong, S.W.; Kim, Y.H.; Lim, C.S.; Abd El-Aty, A.M.; Kim, G.S.; et al. Determination of polyphenols in three Capsicum annuum L. (bell pepper) varieties using high-performance liquid chromatography tandem mass spectrometry: Their contribution to overall antioxidant and anticancer activity. J. Sep. Sci. 2011, 34, 2967–2974. [Google Scholar] [CrossRef]
- Materska, M.; Piacente, S.; Stochmal, A.; Pizza, C.; Oleszekc, W.; Perucka, I. Isolation and structure elucidation of flavonoid and phenolic acid glycosides from pericarp of hot pepper fruit Capsicum annuum L. Phytochemistry 2003, 63, 893–898. [Google Scholar] [CrossRef]
- Materska, M.; Perucka, I. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef]
- Lee, Y.; Howard, L.R.; Villarón, B. Flavonoids and antioxidant activity of fresh pepper (Capsicum annuum) cultivars. J. Food Sci. 1995, 60, 473–476. [Google Scholar] [CrossRef]
- Jang, Y.K.; Jung, E.S.; Lee, H.A.; Choi, D.; Lee, C.H. Metabolomic characterization of hot pepper (Capsicum annuum “cM334”) during fruit development. J. Agric. Food Chem. 2015, 63, 9452–9460. [Google Scholar] [CrossRef]
- Sadilova, E.; Stintzing, F.C.; Carle, R. Anthocyanins, color and antioxidant properties of eggplant (Solanum melongena L.) and violet pepper (Capsicum annuum L.) peel extracts. Z. Naturforsch. C J. Biosci. 2006, 61, 527–535. [Google Scholar] [CrossRef]
- Lightbourn, G.J.; Griesbach, R.J.; Novotny, J.A.; Clevidence, B.A.; Rao, D.D.; Stommel, J.R. Effects of anthocyanin and carotenoid combinations on foliage and immature fruit color of Capsicum annuum L. J. Hered. 2008, 99, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Bonesi, M.; Conforti, F.; Statti, G.; De Luca, D.; de Cindio, B.; Menichini, F. Comparative study on the chemical composition, antioxidant properties and hypoglycaemic activities of two Capsicum annuum L. cultivars (Acuminatum small and Cerasiferum). Plant Foods Hum. Nutr. 2011, 66, 261–269. [Google Scholar] [CrossRef]
- Sun, T.; Xu, Z.; Wu, C.T.; Janes, M.; Prinyawiwatkul, W.; No, H.K. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L.). J. Food Sci. 2007, 72, S98–S102. [Google Scholar] [CrossRef]
- Ichiyanagi, T.; Kashiwada, Y.; Shida, Y.; Ikeshiro, Y.; Kaneyuki, T.; Konishi, T. Nasunin from eggplant consists of cis-trans isomers of delphinidin 3-[4-(p-coumaroyl)-l-rhamnosyl (1→6)glucopyranoside]-5-glucopyranoside. J. Agric. Food Chem. 2005, 53, 9472–9477. [Google Scholar] [CrossRef] [PubMed]
- Poiroux-Gonord, F.; Bidel, L.P.R.; Fanciullino, A.L.; Gautier, H.; Lauri-Lopez, F.; Urban, L. Health benefits of vitamins and secondary metabolites of fruits and vegetables and prospects to increase their concentrations by agronomic approaches. J. Agric. Food Chem. 2010, 58, 12065–12082. [Google Scholar] [CrossRef] [PubMed]
- Perry, L.; Dickau, R.; Zarrillo, S.; Holst, I.; Pearsall, D.M.; Piperno, D.R.; Berman, M.J.; Cooke, R.G.; Rademaker, K.; Ranere, A.J.; et al. Starch fossils and the domestication and dispersal of chili peppers (Capsicum spp. L.) in the Americas. Science 2007, 315, 986–988. [Google Scholar] [CrossRef]
- Paran, I.; Van Der Knaap, E. Genetic and molecular regulation of fruit and plant domestication traits in tomato and pepper. J. Exp. Bot. 2007, 58, 3841–3852. [Google Scholar] [CrossRef]
- Rousseaux, M.C.; Jones, C.M.; Adams, D.; Chetelat, R.; Bennett, A.; Powell, A. QTL analysis of fruit antioxidants in tomato using Lycopersicon pennellii introgression lines. Theor. Appl. Genet. 2005, 111, 1396–1408. [Google Scholar] [CrossRef]
- Di Matteo, A.; Ruggieri, V.; Sacco, A.; Rigano, M.M.; Carriero, F.; Bolger, A.; Fernie, A.R.; Frusciante, L.; Barone, A. Identification of candidate genes for phenolics accumulation in tomato fruit. Plant Sci. 2013, 205–206, 87–96. [Google Scholar] [CrossRef]
- McCown, B.; Zeldin, E. “HyRed”, an early, high fruit color cranberry hybrid. HortScience 2003, 23, 304–305. [Google Scholar] [CrossRef]
- Khanizadeh, S.; Ehsani-Moghaddam, B.; Levasseur, A. Antioxidant capacity in June-bearing and day-neutral strawberry. Can. J. Plant Sci. 2006, 86, 1387–1390. [Google Scholar] [CrossRef]
- Cevallos-Casals, B.A.; Byrne, D.; Okie, W.R.; Cisneros-Zevallos, L. Selecting new peach and plum genotypes rich in phenolic compounds and enhanced functional properties. Food Chem. 2006, 96, 273–280. [Google Scholar] [CrossRef]
- Verhoeyen, M.E. Increasing antioxidant levels in tomatoes through modification of the flavonoid biosynthetic pathway. J. Exp. Bot. 2002, 53, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Giovinazzo, G.; D’Amico, L.; Paradiso, A.; Bollini, R.; Sparvoli, F.; DeGara, L. Antioxidant metabolite profiles in tomato fruit constitutively expressing the grapevine stilbene synthase gene. Plant Biotechnol. J. 2005, 3, 57–69. [Google Scholar] [CrossRef]
- Schijlen, E.; Ric De Vos, C.H.; Jonker, H.; Van Den Broeck, H.; Molthoff, J.; Van Tunen, A.; Martens, S.; Bovy, A. Pathway engineering for healthy phytochemicals leading to the production of novel flavonoids in tomato fruit. Plant Biotechnol. J. 2006, 4, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Van Der Rest, B.; Danoun, S.; Boudet, A.M.; Rochange, S.F. Down-regulation of cinnamoyl-CoA reductase in tomato (Solanum lycopersicum L.) induces dramatic changes in soluble phenolic pools. J. Exp. Bot. 2006, 57, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Bovy, A. High-flavonol tomatoes resulting from the heterologous expression of the maize transcription factor genes LC and C1. Plant Cell Online 2002, 14, 2509–2526. [Google Scholar] [CrossRef]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.P.; Matros, A.; Peterek, S.; Schijlen, E.G.W.M.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Løvdal, T.; Olsen, K.M.; Slimestad, R.; Verheul, M.; Lillo, C. Synergetic effects of nitrogen depletion, temperature, and light on the content of phenolic compounds and gene expression in leaves of tomato. Phytochemistry 2010, 71, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D.; Kleinwächter, M. Stress enhances the synthesis of secondary plant products: The impact of stress-related over-reduction on the accumulation of natural products. Plant Cell Physiol. 2013, 54, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crops Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Bénard, C.; Gautier, H.; Bourgaud, F.; Grasselly, D.; Navez, B.; Caris-Veyrat, C.; Weiss, M.; Génard, M. Effects of low nitrogen supply on tomato (Solanum lycopersicum) fruit yield and quality with special emphasis on sugars, acids, ascorbate, carotenoids, and phenolic compounds. J. Agric. Food Chem. 2009, 57, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Schonhof, I.; Kläring, H.P.; Krumbein, A.; Claußen, W.; Schreiner, M. Effect of temperature increase under low radiation conditions on phytochemicals and ascorbic acid in greenhouse grown broccoli. Agric. Ecosyst. Environ. 2007, 119, 103–111. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zheng, W. Effect of plant growth temperature on antioxidant capacity in strawberry. J. Agric. Food Chem. 2001, 49, 4977–4982. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Duan, Y.; Ju, Z. Effects of covering the orchard floor with reflecting films on pigment accumulation and fruit coloration in “Fuji” apples. Sci. Hortic. 1999, 82, 47–56. [Google Scholar] [CrossRef]
- Wilkens, R.T.; Spoerke, J.M.; Stamp, N.E. Differential responses of growth and two soluble phenolics of tomato to resource availability. Ecology 1996, 77, 247–258. [Google Scholar] [CrossRef]
- Gautier, H.; Diakou-Verdin, V.; Bénard, C.; Reich, M.; Buret, M.; Bourgaud, F.; Poëssel, J.L.; Caris-Veyrat, C.; Génard, M. How does tomato quality (sugar, acid, and nutritional quality) vary with ripening stage, temperature, and irradiance? J. Agric. Food Chem. 2008, 56, 1241–1250. [Google Scholar] [CrossRef]
- Torres, C.A.; Andrews, P.K.; Davies, N.M. Physiological and biochemical responses of fruit exocarp of tomato (Lycopersicon esculentum Mill.) mutants to natural photo-oxidative conditions. J. Exp. Bot. 2006, 57, 1933–1947. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.M.; Flores, P.; Garrido, C.; Martinez, V. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006, 96, 66–73. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Modifications of strawberry fruit antioxidant pools and fruit quality under NaCl stress. J. Agric. Food Chem. 2007, 55, 4066–4072. [Google Scholar] [CrossRef]
- De Pascale, S.; Martino, A.; Raimondi, G.; Maggio, A. Comparative analysis of water and salt stress-induced modifications of quality parameters in cherry tomatoes. J. Hortic. Sci. Biotechnol. 2007, 82, 283–289. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Hoppula, K.I.; Nestby, R.; Verheul, M.J.; Karjalainen, R.O. Influence of fertilization, mulch color, early forcing, fruit order, planting date, shading, growing environment, and genotype on the contents of selected phenolics in strawberry (Fragaria x ananassa Duch.) fruits. J. Agric. Food Chem. 2006, 54, 2614–2620. [Google Scholar] [CrossRef]
- Lekala, C.S.; Madani, K.S.H.; Phan, A.D.T.; Maboko, M.M.; Fotouo, H.; Soundy, P.; Sultanbawa, Y.; Sivakumar, D. Cultivar-specific responses in red sweet peppers grown under shade nets and controlled-temperature plastic tunnel environment on antioxidant constituents at harvest. Food Chem. 2019, 175, 85–94. [Google Scholar] [CrossRef]
- Ribes-Moya, A.M.; Raigón, M.D.; Moreno-Peris, E.; Fita, A.; Rodríguez-Burruezo, A. Response to organic cultivation of heirloom Capsicum peppers: Variation in the level of bioactive compounds and effect of ripening. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Wasternack, C.; Strnad, M. Jasmonates are signals in the biosynthesis of secondary metabolites—Pathways, transcription factors and applied aspects—A brief review. New Biotechnol. 2019, 48, 1–11. [Google Scholar] [CrossRef]
- Heredia, J.B.; Cisneros-Zevallos, L. The effect of exogenous ethylene and methyl jasmonate on pal activity, phenolic profiles and antioxidant capacity of carrots (Daucus carota) under different wounding intensities. Postharvest Biol. Technol. 2009, 51, 242–249. [Google Scholar] [CrossRef]
- Hendrawati, O.; Yao, Q.; Kim, H.K.; Linthorst, H.J.M.; Erkelens, C.; Lefeber, A.W.M.; Choi, Y.H.; Verpoorte, R. Metabolic differentiation of Arabidopsis treated with methyl jasmonate using nuclear magnetic resonance spectroscopy. Plant Sci. 2006, 170, 1118–1124. [Google Scholar] [CrossRef]
- Mejía-Teniente, L.; de Dalia Duran-Flores, F.; Chapa-Oliver, A.M.; Torres-Pacheco, I.; Cruz-Hernández, A.; González-Chavira, M.M.; Ocampo-Velázquez, R.V.; Guevara-González, R.G. Oxidative and molecular responses in Capsicum annuum L. after hydrogen peroxide, salicylic acid and chitosan foliar applications. Int. J. Mol. Sci. 2013, 14, 10178–10196. [Google Scholar] [CrossRef]
- Groher, T.; Schmittgen, S.; Noga, G.; Hunsche, M. Limitation of mineral supply as tool for the induction of secondary metabolites accumulation in tomato leaves. Plant Physiol. Biochem. 2018, 130, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Padilla, F.M.; Peña-Fleitas, M.T.; Fernández, M.D.; del Moral, F.; Thompson, R.B.; Gallardo, M. Responses of soil properties, crop yield and root growth to improved irrigation and N fertilization, soil tillage and compost addition in a pepper crop. Sci. Hortic. 2017, 225, 422–430. [Google Scholar] [CrossRef]
- Usadel, B.; Wormit, A. InducTomE. Available online: http://www.biosc.de/inductome (accessed on 29 March 2019).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Content | Content Expressed In | Reference | |||
|---|---|---|---|---|---|---|
| Fruit (Pericarp) | Leaves | Stems | Other | |||
| Phenolic Content | ||||||
| Total Phenolic Content | 6.97–12.19 | - | - | - | mg CE/g f.w | [108] |
| 6.92–7.32 | 17.14 | - | - | mg CE/g dw | [107] | |
| 1.03 * | - | - | - | mg GAE/g fw | [111] | |
| 52.27 | - | 71.34 | 64.07 (placenta) | mg GAE/g dw | [112] | |
| Total Flavonoid Content | 0.017–0.2024 | - | - | - | mg/g fw | [108,109] |
| 0.00366–0.0054 | 0.0206 | - | - | mg/g dw | [107] | |
| 46.71 | - | 69.84 | 58.96 (placenta) | mg NAE/g dw | [112] | |
| 0.3864 | - | - | - | mg QE/g fw | [111] | |
| Total O-glycosyl-flavones | 1.00–9.43 | - | - | - | mg/100g fw | [108] |
| Total C-glycosyl-flavones | 0.84–2.45 | - | - | - | mg/100g fw | [108] |
| Total hydroxyl-cinnamics | 0.33–1.99 | - | - | - | mg/100g fw | [108] |
| Antioxidant Activity | ||||||
| Radical scaveng-ing activity (RSA) | 17.32–48.95 | - | - | - | TEAC/kg dw | [113] |
| DPPH | 15.40–286.23 | 463.23 | - | - | IC50 μg 80% EtOH extract (v/v)/mL | [107] |
| - | 49.2–62.2 | - | - | IC50 μg/mL | [114] | |
| 60.41 * | - | - | - | % of inhibition | [111] | |
| ABTS | 55.23–118.00 | 68.67 | - | - | IC50 μg 80% EtOH extract(v/v)/mL | [107] |
| - | 26.2–33.7 | - | - | IC50 μg/mL | [114] | |
| 34.44 * | - | - | - | μM TEAC/g fw | [111] | |
| β-carotene bleaching | 42.75–66.98 | - | - | - | % of inhibition | [109] |
| Identified In | Reference | ||||
|---|---|---|---|---|---|
| Fruit | Leaves | Stems | Other | ||
| Class of Phenolic Compounds | a | a | a | a | |
| Phenolic acids | |||||
| Quinic Acid | ID | - | - | - | [116] |
| Hydroxybenzoic acids | |||||
| Gallic acid | 2.53–1013.0 | - | - | - | [111,113,116,117] |
| 1-O-galloyl-β-D-glucose | ID | - | - | - | [116] |
| Protocatechuic acid | 0.29–0.83 | - | - | - | [113] |
| p-Hydroxybenxoic acid | 420 b | - | - | 260 b (placenta) | [112] |
| 2.88–6.27 | - | - | - | [113] | |
| p-Hydroxyphenylacetic acid | 0.41–1.77 | - | - | - | [113] |
| p-Hydroxybenzoic acid β-glucoside | ID | - | - | - | [110] |
| Vanillic acid | - | - | 130 b | 1020 b (placenta) | [112] |
| 3.07–13.29 | - | - | - | [113] | |
| Vanillic acid glucoside | 4.02 | - | - | - | [118] |
| Vanillic-acid-1-O-β-D-gluco-pyranosylester | ID | - | - | - | [110,116] |
| Hydroxycinnamic acids | |||||
| Caffeic acid | 1320 b | - | - | 120 b (placenta) | [112] |
| 0.55–13.7 | - | - | - | [108,111,113] | |
| Hydrocaffeic acid | 3.03 | - | - | - | [118] |
| Caffeoyl glucoside | 2.59 | - | - | - | [118] |
| 5-O-Caffeoylquinic acid | 0.36–23.33 | 260–360 | - | - | [113,114] |
| 5-O-Caffeoylquinic acid methyl ester | - | 10–340 | - | - | [114] |
| 5-O-Caffeoylquinic acid butyl ester | - | 20 | - | - | [114] |
| Caffeic acid-4-O-β-D-glucopyranoside | ID | - | - | - | [110] |
| 1-O-Caffeoyl-β-D-glucopyranoside | ID | - | - | - | [110] |
| Caffeoyl-O-hexoside | ID | - | - | - | [119] |
| 2-Caffeoyl-L-tartaric acid | ID | - | - | - | [116] |
| 4-O-caffeoylquinic acid | ID | - | - | - | [116] |
| Chlorogenic acid | 1560 b | - | 3820 b | 230 b (placenta) | [112] |
| 2–17.9 | - | - | - | [111,117] | |
| p-Coumaric acid | 4560 b | - | 2980 b | 820 b (placenta) | [112] |
| 0.09–6.97 | - | - | - | [113,118] | |
| Coumaroyl quinic acid | ID | - | - | - | [116] |
| p-Coumaryl tyrosine | 6.81 | - | - | - | [118] |
| p-Coumaryl glycolic acid | 6.47 | - | - | - | [118] |
| trans-p-coumaroyl-α-D-gluco-pyranoside | 0.5–1.0 | [108] | |||
| Ferulic acid | 120 b | - | - | - | [112] |
| 2.18–12.45 | - | - | - | [113] | |
| Feruloyl-O-glucoside | 2.5 | - | - | - | [119] |
| Feruloyl hexoside | 32–53.4 | - | - | - | [120] |
| Feruloyl-β-D-glucose | ID | - | - | - | [116] |
| trans-feruloyl-α-D-gluco-pyranoside | 0.9–1.0 | [108] | |||
| trans-p-feruloyl-β-D-gluco-pyranoside | 84–359 b | - | - | - | [121,122] |
| Syringic acid | 0.66–1.21 | - | - | - | [113] |
| Sinapic acid | 0.45–1.89 | - | - | - | [113] |
| Sinapoyl hexoside | 65.0–72.4 | - | - | - | [120] |
| trans-sinapoyl-α-D-gluco-pyranoside | 1.4–3.0 | - | - | - | [108] |
| trans-p-sinapoyl-β-D-gluco-pyranoside | 74–419 b | - | - | - | [121,122] |
| Cinnamic acid | 0.03–0.24 | - | - | - | [113] |
| Coumarins | |||||
| Hydroxycoumarin | 2.42 | - | - | - | [118] |
| Daphnetine (7,8-Dihydroxycoumarin) | 16.29 | - | - | - | [118] |
| Aesculin (Esculetin-6-β-D-glucoside) | 0.16–0.20 | - | - | - | [113] |
| Umbelliferone (7-hydroxycoumarin) | 10.66–14.61 | - | - | - | [113] |
| Chalcones | |||||
| Phloretin dihexoside | ID | - | - | - | [110] |
| Flavonoids | |||||
| Flavonols | |||||
| Quercetin | 8860 b | 5.72 b | 6510 b | 10270 b (placenta) | [112,116] |
| 3.29–783.83 | - | - | - | [107,109,113,117,118,123] | |
| Quercetin-3-O-hexoside | 13.5 | - | - | - | [120] |
| Quercetin rhamnoside | 82.60 | - | - | - | [118] |
| Quercetin dihexoside | ID | - | - | - | [110] |
| Quercitrin (Quercetin-3-O-rhamnoside) | 0.5–1501.8 | - | - | - | [108,117,119,120] |
| Quercetin-O-rhamnosyl-O-hexoside | 17.6–23.1 | - | - | - | [120] |
| Quercetin glucoside | 19.86 | - | - | - | [118] |
| Quercetin-3-vicianoside | ID | - | - | - | [110] |
| Quercetin rhamnoside-glucoside | ID | - | - | - | [84] |
| Quercetin-3-O-glucoside-7-O-rhamnoside | ID | - | - | - | [84] |
| Quercetin-3,7-di-O-α-L-rhamno-pyranoside | ID | - | - | - | [110] |
| Quercetin-3,7-diglucuronide | ID | - | - | - | [116] |
| Quercetin-3-rhamnopyranoside | ID | - | - | - | [110] |
| Quercetin-3′-O-β-D-gluco-pyranoside | ID | - | - | - | [110] |
| Quercetin-4′-β-D-glucopyranoside | ID | - | - | - | [110] |
| Quercetin-3-O-α-L-rhamno-pyranoside | 113–993 b | - | - | - | [121,122] |
| Quercetin-3-O-α-L-rhamnopyranoside-7-O-β-D-glucopyranoside | ID | - | - | - | [121] |
| Quercetin-3-O-(6′-O-acetyl)-β-D-glucopyranoside | ID | - | - | - | [116] |
| Hyperoside (Quercetin-3-O-galactoside) | 1.81–27.28 | - | - | - | [113] |
| Isoquercitrin | 31.9–52.5 | - | - | - | [117] |
| Rutin | 1.05–52.0 | - | - | - | [111,113,117] |
| Rutin pentoside | ID | - | - | - | [110] |
| Kaempferol | 50 b | - | 160 b | - | [112] |
| 128.0–143.0 | - | - | - | [117] | |
| Kaempferol-3-O-hexoside | ID | - | - | - | [124] |
| Kaempferol-di-glucoside | 17.17 | - | - | - | [118] |
| Kaempferol-feruloyl dihexoside | ID | - | - | - | [124] |
| Kaempferol-pentosyldihexoside | 42.1 | - | - | - | [120] |
| Kaempferol-O-pentosyldihexoside | 1.3 | - | - | - | [119] |
| Kaempferol-7,4′-dimethoxy-8-butyryl ester | ID | - | - | - | [116] |
| Myricetin | 2280 b | - | - | 1380 b (placenta) | [112] |
| Flavan-3-ols | |||||
| Catechin | - | - | 30 b | - | [112] |
| 1.1–36.8 | - | - | - | [111] | |
| Epicatechin | 1–79.3 | - | - | - | [111] |
| Flavones | |||||
| Dihydroxyflavone-O-hexoside | 3.1 | - | - | - | [119] |
| Chrysoeriol-6,8-di-C-hexoside | ID | - | - | - | [124] |
| Apigenin | 1.38 | - | - | - | [113] |
| Apigenin-7-O-glucopyranoside | ID | - | - | - | [124] |
| Apigenin-C-pentosyl-C-hexoside | 5.5–7.4 | - | - | - | [120] |
| Apigenin-6,8-di-C-hexoside | ID | - | - | - | [84] |
| Apigenin-6-C-pentoside-8-C-hexoside | ID | - | - | - | [84] |
| Vicenin 2 (Apigenin-6,8-di-C-glucoside) | 12.7 | - | - | - | [120] |
| Apiin (Apigenin-7-O-apioglucoside) | 0.27–0.43 | - | - | - | [113] |
| Apigenin-7-O-β-D-apiofuranosyl (1→2)-β-D-glucopyranoside | ID | 6–3530 | - | - | [110,114] |
| Apigenin-8-C-glucoside | ID | - | - | - | [116] |
| Apigenin-8-C-α-L-arabinoside-6-C-β-D-glucoside | ID | - | - | - | [110] |
| Schaftoside (Apigenin-6-C-β-D-gluco-pyranoside-8-C-α-L-arabinopyranosid) | 18–109 b | - | - | - | [121,122] |
| Diosmetin-7-O-β-D-glucoside | ID | - | - | - | [110] |
| Vitexin | 0.15–3.50 | - | - | - | [113] |
| Luteolin | 260 b | 14.34 b | 280 b | 310b (placenta) | [112] |
| 0.16–103.50 | - | - | - | [107,109,111,113,118,123] | |
| Luteolin-glucoside | 5.09 | - | - | - | [118] |
| Luteolin-di-glucoside | 5.66 | - | - | - | [118] |
| Luteolin-C-pentosyl-C-hexoside | 8.0–17.4 | - | - | - | [120] |
| Luteolin-6-C-hexoside | 7.7 | - | - | - | [120] |
| Luteolin-8-C-hexoside | 19–26.5 | - | - | - | [120,124] |
| Orientin (Luteolin-8-C-glucoside) | 12.7 | - | - | - | [120] |
| Isoscoparin (Chrysoeriol-6-C-glucoside) | 23.4 | - | - | - | [120] |
| Luteolin-6-C-pentoside-8-C-hexoside | ID | - | - | - | [84] |
| Luteolin-6,8-di-C-hexoside | 1.7–12.9 | - | - | - | [120] |
| Luteolin-C-6-malonyl-C-pentoside | ID | - | - | - | [124] |
| Luteolin-6-C-β-D-glucopyranoside-8-C-α-L-arabinopyranoside | 59–299 b | - | - | - | [121,122] |
| Luteolin-O-(apiosylacetyl)-glucoside | 6.1-19.2 | - | - | - | [120] |
| Luteolin-O-maloylpentosyldi-hexoside | 105.3 | - | - | - | [120] |
| Luteolin-O-(apiosylmalonyl)-glucoside | 60.1–206.6 | - | - | - | [120] |
| Graveobioside A (Luteolin-7-O-[2-(α-D-apiofuranosyl)-α-D-glucopyranoside]) | 160–1367 b | - | - | - | [121,122] |
| Graveobioside A (Luteolin-7-O-β-D-apiofuranosyl-(1->2)-β-D-glucopyranoside) | 4.179 | 4.1 –121.36 | - | - | [114,115,121] |
| Graveobioside A (Luteolin-7-O-apiosyl-glucoside) | ID | - | - | - | [124] |
| Graveobioside A (Luteolin-7-O-(2-apiosyl)glucoside) | 0.1 | - | - | - | [119] |
| Luteolin-7-O-(2-apiosyl-6-malonyl)glucoside | 3.9–41.4 | - | - | - | [108] |
| Luteolin-7-O-[2-(β-D-apiofurano-syl)-4-(β-D-glucopyranosyl)-6-malonyl]-β-D-glucopyranoside | 47-412 b | - | - | - | [121,122] |
| Cynaroside (Luteolin-7-O-glucopyranoside) | 1.00–7.21 | 30-370 | - | - | [113,114] |
| Isoflavones | |||||
| Genistein-4,7′-dimethyl ether | ID | - | - | - | [116] |
| 6′-O-acetyl-Daidzin | ID | - | - | - | [116] |
| Flavanones | |||||
| Naringenin | 1.12–4.46 | - | - | - | [113] |
| Naringenin-chalcone-hexose | ID | - | - | - | [110] |
| Naringenin-7-O-β-D-glucoside | ID | - | - | - | [110] |
| Narigenin-7-O-β-D-(3″-p-couma-royl)-glucopyranoside | ID | - | - | - | [110] |
| Theaflavanoside I | ID | - | - | - | [110] |
| Flavanonols | |||||
| 7-O-β-D-Glucopyranosyl-dihydroquercetin-3-O-α-D-glucopyranoside | ID | - | - | - | [110] |
| Anthocyanidins | |||||
| Nasunin (Delphinidin-3-(p-coumaroylrutino-side)-5-glucoside) | 14.7–286.2 | 1525 | - | - | [125,126] |
| Delphinidin-3-[4-(trans-coumaroyl)-L-rhamnosyl(1→6)glucopyrano-side]-5-O-glucopyranoside | - | 860 | - | - | [114] |
| Myrtillin (Delphinidin-3-O-gluco-side) | 2.2 | - | - | - | [125] |
| Tulipanin (Delphinidin-3-O-ru-tinoside) | 2.9 | - | - | - | [125] |
| Delphinidin-3-rutinoside-5-glucoside | 1.5 | - | - | - | [125] |
| Delphinidin-3-rhamnoside | 2.4 | - | - | - | [125] |
| Delphinidin-3-caffeoylrutinoside-5-glucoside | 7.8 | - | - | - | [125] |
| Stilbenes | |||||
| Resveratrol | 3.8–12.2 | - | - | - | [111] |
| Other phenolics | |||||
| Vanillin | 0.19–0.85 | - | - | - | [113] |
| Conyferyl aldehyde | 0.26–0.43 | - | - | - | [113] |
| 2′-O-β-Apiofuranosyl-arbutin | ID | - | - | - | [110] |
| Lariciresinol-glucopyranoside | ID | - | - | - | [110] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho Lemos, V.; Reimer, J.J.; Wormit, A. Color for Life: Biosynthesis and Distribution of Phenolic Compounds in Pepper (Capsicum annuum). Agriculture 2019, 9, 81. https://doi.org/10.3390/agriculture9040081
Carvalho Lemos V, Reimer JJ, Wormit A. Color for Life: Biosynthesis and Distribution of Phenolic Compounds in Pepper (Capsicum annuum). Agriculture. 2019; 9(4):81. https://doi.org/10.3390/agriculture9040081
Chicago/Turabian StyleCarvalho Lemos, Virgínia, Julia Jessica Reimer, and Alexandra Wormit. 2019. "Color for Life: Biosynthesis and Distribution of Phenolic Compounds in Pepper (Capsicum annuum)" Agriculture 9, no. 4: 81. https://doi.org/10.3390/agriculture9040081
APA StyleCarvalho Lemos, V., Reimer, J. J., & Wormit, A. (2019). Color for Life: Biosynthesis and Distribution of Phenolic Compounds in Pepper (Capsicum annuum). Agriculture, 9(4), 81. https://doi.org/10.3390/agriculture9040081