1. Introduction
Situated between 20° and 45° north latitude, Japan shares a similar geographical position with other major grape-growing regions in the Northern Hemisphere. However, its unique climatic conditions—characterized by high rainfall and humidity from late May through August, coinciding with critical grapevine phenological stages (flowering to berry set)—make the environment particularly susceptible to fungal and oomycetes diseases. These include gray mold, grape rot, powdery mildew, powdery black grape mildew, grape rust, and downy mildew. Among them, downy mildew poses a significant threat, necessitating stringent control measures for the cultivation of Vitis vinifera grapes.
Downy mildew, caused by the oomycete pathogen
Plasmopara viticola (Berk. & Curtis) Berl. & De Toni, primarily affects grapevine leaves in Japan but can also infect shoots, tendrils, and young fruits (
Figure 1). The coalescence of foliar lesions into large chlorotic areas leads to defoliation, reducing photosynthetic capacity and consequently impairing shoot elongation and berry development [
1]. Additionally, the pathogen preferentially infects young grape clusters and berries. Infected grapes develop a characteristic leaden coloration, followed by skin rupture or shriveling and eventual berry abscission, resulting in substantial yield losses [
2]. In Japan, where
V. vinifera was introduced from France in 1877, downy mildew infection was first confirmed in 1890 [
3]. Initially, the disease was recognized for its role in accelerating the defoliation of wine grapevines during the late August and September rains. However, it was largely overlooked due to the absence of fruit damage, as seen in other European countries. The introduction of the low downy mildew-resistant Neo-Muscat cultivar (
V. vinifera cv. Muscat of Alexandria ×
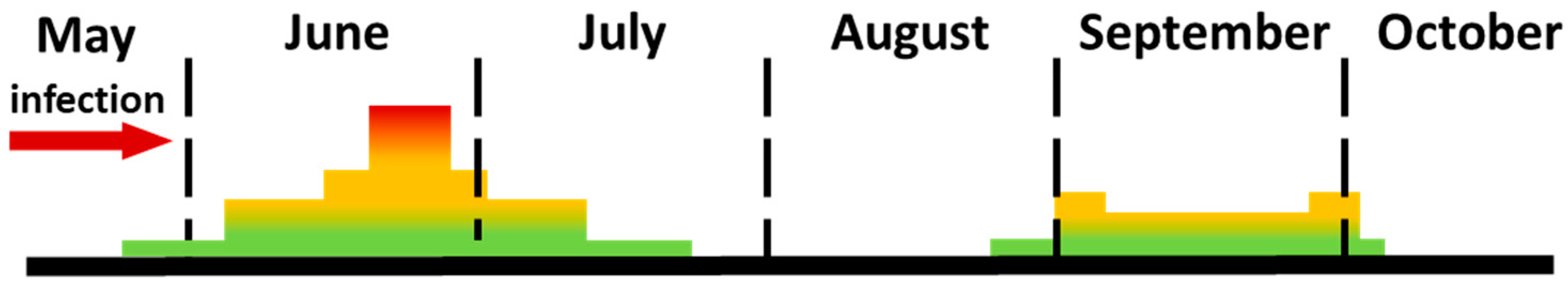
V. vinifera cv. Koshu Sanjyaku) around 1950 coincided with the emergence of infections during the rainy season from late May to June. By the 1970s, downy mildew had become widespread among both table and wine grape cultivars across Japan. Since then, disease outbreaks in Japan have exhibited a bimodal distribution, peaking in June and September under conditions of high humidity (≥70%) and temperatures ranging from 10 °C to 32 °C (
Figure 2) [
3].
Probability of infection increases as it approaches a red color.
Traditionally, downy mildew has been managed primarily with Bordeaux mixture and chemical agents designed for fungal pathogens. Oomycetes, though historically classified as fungi, are phylogenetically distinct and reproduce sexually via occurred oospores in leaf. A key difference between oomycetes and fungi lies in their cell wall and membrane composition. Fungal cell walls primarily contain chitin, chitosan, and glucan, whereas oomycete cell walls consist of cellulose and glucan. Similarly, ergosterol is the dominant sterol in fungal cell membranes, while oomycetes primarily utilize cholesterol.
2. Chronology and Effectiveness of Fungicides in Japan Viticulture
A diverse array of chemical agents has been employed for grapevine disease management in Japan, ranging from traditional compounds like copper sulfate and dithiocarbamate fungicides to more recently developed fungicides from various chemical classes. These include phthalimides, phosphonates, phenylamides, 2,6-dinitroanilines, cyano-acetamide oximes, benzamides, quinones, quinone outside inhibitors (QoI), quinone inside inhibitors (QiI), carboxylic acid amides (CAA), piperidinyl thiazole isoxazolines, thiazole carboxamides, and oxysterol-binding protein inhibitors (OSBPI) (Fungicide Resistance Action Committee, FRAC,
Table 1).
In 1899, the Bordeaux mixture, a combination of lime and copper sulfate, was introduced and subsequently adopted in Yamanashi Prefecture, Japan’s primary grape-growing region. This fungicide remained the sole control measure for grape downy mildew until the 1960s. The registration of organosulfur dithiocarbamate fungicides, such as mancozeb and maneb, in 1969 marked a significant advancement in disease management. In 1972, quinone fungicides (e.g., dithianon) and phthalimide fungicides (e.g., captan) were registered for use alongside copper sulfate and dithiocarbamates.
Following captan registration, efforts focused on introducing highly selective chemical agents, leading to the approval of phosphonate fungicides like fosetyl-aluminium in 1983, either as a standalone treatment or in combination with phthalimides. These chemical agents exhibit diverse modes of action, reducing the likelihood of resistance development (see FRAC,
http://www.frac.info, accessed on 2 September 2024). Dithiocarbamate fungicides, in particular, remain a cornerstone of disease management due to their broad-spectrum efficacy against a range of grapevine pathogens (
Table 2).
Phenylamide fungicides, exemplified by metalaxyl, were introduced in 1986. Metalaxyl inhibits mycelial growth and sporulation by disrupting RNA, DNA, and lipid synthesis through the inhibition of uridine incorporation. This marked a shift towards the development of single-mode-of-action agents, leading to the registration of 2,6-dinitroaniline fungicides, such as fluazinam, in 1990. Between 1997 and 2001, the QoI fungicides famoxadone and azoxystrobin, along with the QiI fungicide cyazofamid, were registered. These compounds exert fungicidal action by inhibiting mitochondrial ubiquinol oxidase (Qo site) and ubiquinone reductase (Qi site), respectively. In 1997, the CAA fungicide dimethomorph was introduced, targeting cellulose synthase, a key enzyme in fungal cell wall biosynthesis. Additional CAA fungicides, benthiavalicarb-isopropyl and mandipropamid, were registered in 2007 and 2009, respectively.
Subsequently, the piperidinyl thiazole isoxazoline fungicide fluopicolide and the thiazole carboxamide fungicide ethaboxam were introduced in 2012 and 2014, respectively, both primarily targeting cell division. Fluopicolide disrupts cell membrane integrity by delocalizing spectrin-like proteins, whereas ethaboxam inhibits β-tubulin polymerization, a critical process in chromosome division [
4]. The combined application of CAA fungicides and fluopicolide is expected to enhance cell division inhibition through synergistic effects. In 2019, oxathiapiprolin, an OSBPI fungicide, was registered. This compound disrupts lipid homeostasis by inhibiting oxysterol-binding proteins involved in lipid transport across membranes.
To date, approximately 60 chemical agents have been registered for grape downy mildew control in Japan, with oxathiapiprolin being the most recent addition. However, many of these agents have been withdrawn from disease management programs due to the emergence of resistant downy mildew populations. For example, metaraxyl was found to cause the emergence of drug-resistant grapevine mildew in France in 1981 [
5]. In Japan, a decrease in efficacy was observed in 2009 and the emergence of resistant strains in 2010. For this reason, some metalaxyl mixtures have been removed from the pest management calendar since 2010 [
6]. In the following sections, we will examine the development of resistance to QoI, CAA, and OSBPI fungicides in Japan.
3. QoI Fungicide Resistance
The widespread adoption of azoxystrobin, a QoI fungicide, in Japan around 2000 coincided with growing global and domestic concerns over climate change. During this period, rising temperatures and increased precipitation levels were documented in Japan [
7]. Moreover, elevated nighttime temperatures associated with accelerated global warming have been linked to the increased susceptibility of Japanese grapevines to downy mildew infections [
8]. The heightened incidence of grape downy mildew has necessitated multiple applications of azoxystrobin-based fungicides starting in mid-May, a critical period for grape development. Already by this year, however, QoI resistance in grape downy mildew had first been detected in populations in northern Italy and France [
9], and has since been confirmed in almost all major viticultural areas of these two countries, dramatically reducing the use of QoI fungicides for downy mildew control [
7]. A severe grape downy mildew outbreak in Yamanashi Prefecture in 2010 highlighted the challenges posed by this disease, resulting in significant economic losses for grape growers [
6]. Consequently, wine production in Yamanashi Prefecture plummeted by 26% compared to the previous year [
10]. Subsequent investigations revealed that the devastation was attributable to QoI fungicide-resistant grape downy mildew [
11].
Azoxystrobin binds to ubiquinol within the cytochrome bc1 complex, inhibiting ATP synthesis at the Qo site [
12]. This high selectivity disrupts mitochondrial respiration by blocking the electron transfer system [
13]. QoI fungicides were registered in France and Italy in 1997 and 1998, respectively, and subsequently marketed in Japan in 2000. Initially, there was no evidence of resistance development in Japanese grapevine populations. The first report of QoI fungicide resistance in Japanese grapevine downy mildew emerged in 2009 [
14]. Subsequent studies confirmed the widespread occurrence of point mutations, as shown in
Figure 3, in downy mildew populations across major Japanese grape-growing regions [
11]. In Yamanashi Prefecture, the detection rate of QoI-resistant downy mildew surged from 25.3% in 2008 to 56.5% in 2009, indicating a rapid increase in resistance prevalence by 2010. These data suggest that the decline in Yamanashi Prefecture’s wine production in 2010 was exacerbated by insufficient monitoring of QoI resistance development in Japanese grapevine downy mildew populations.
Fungicide resistance testing in grapevine downy mildew, an obligate parasite, is complex and resource-intensive due to the requirement for host plants [
15]. If, as reported by Furuya et al. [
14], fungicide resistance is attributed to a single gene substitution within the pathogen’s genome, PCR-RFLP (polymerase chain reaction–restriction fragment length polymorphism) analysis could serve as a rapid and efficient diagnostic tool. Building upon this knowledge, Aoki et al. [
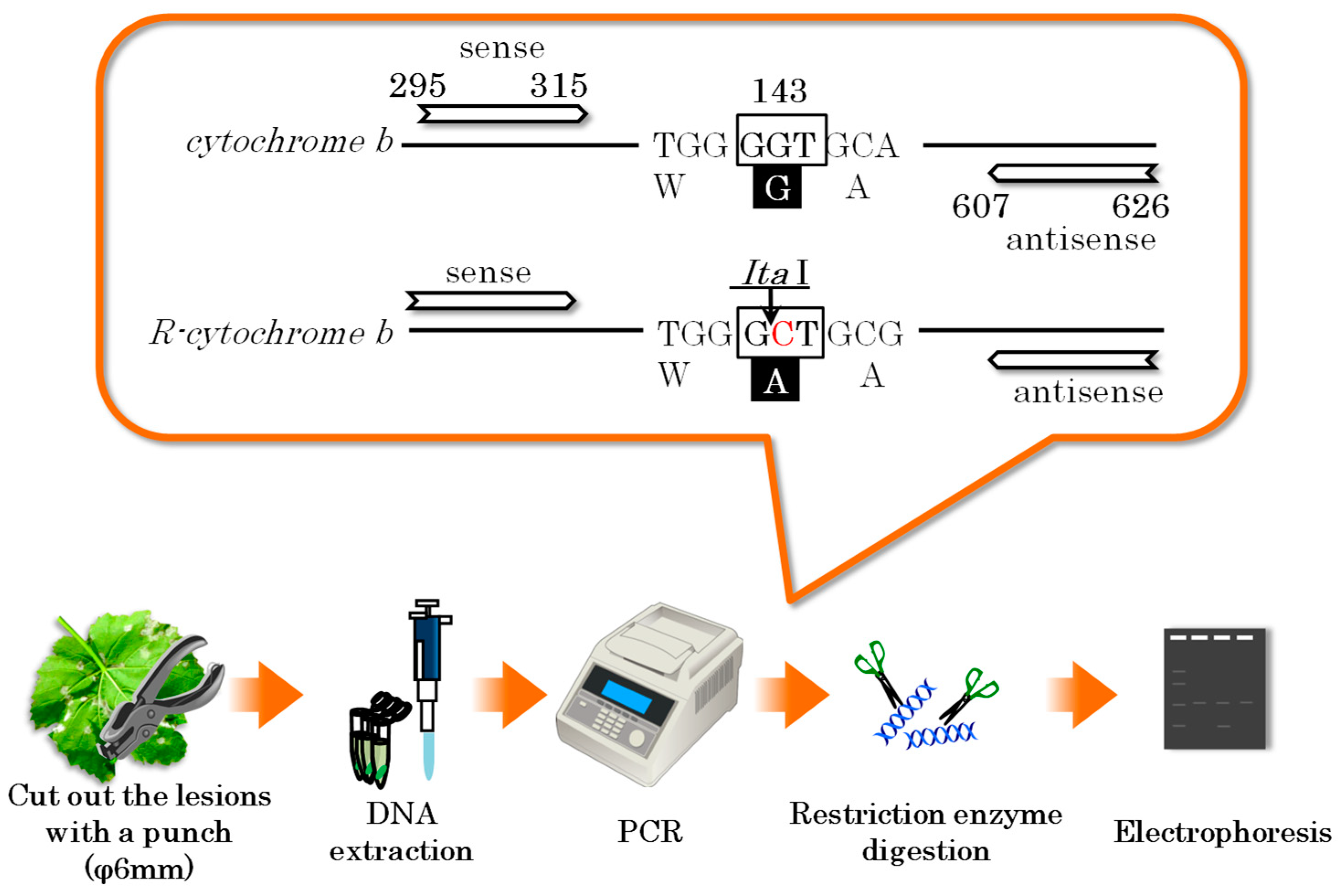
16] intensified surveillance efforts for QoI-resistant grapevine downy mildew. The PCR-RFLP method is a method for detecting nucleotide sequence variation by determining the difference in length of DNA fragments amplified by PCR and digested with restriction enzymes, or by determining whether the DNA fragments have been digested with restriction enzymes (
Figure 3). The PCR-RFLP method was originally developed for rapid identification of pathogens and diagnosis of genetic diseases in infectious diseases based on PCR and has recently become a mainstream method for testing fungicide resistance of plant pathogens due to its reliability and simplicity. Specifically, an annual monitoring program was initiated in Hokkaido, Japan, where no QoI resistance was detected in 2008 or 2009 [
16]. However, in 2014, 30 of 131 isolates from a field in Urausu, Hokkaido, Japan exhibited the G143A mutation. Subsequent monitoring in 2015 and 2016 revealed the persistent presence of QoI-resistant downy mildew in a field in Urausu, Hokkaido, Japan. A field in Urausu, Hokkaido, Japan received a single azoxystrobin application in both 2004 and 2009. Subsequently, the field was subjected to two QoI fungicide applications annually since 2014, specifically azoxystrobin (alone) in June and famoxadone (mixed with cymoxanil) in July. In contrast, a field in Furano, Hokkaido, Japan remained untreated with QoI fungicides until 2016. It is widely accepted that resistant fungi are present in low-frequency populations before exposure to pesticides [
17]. Consequently, the persistent application of QoI fungicides, specifically azoxystrobin and famoxadone, in a field in Urausu, Hokkaido, Japan is likely responsible for the emergence of resistant grapevine downy mildew populations. In particular, the susceptibility of downy mildew to famoxadone is known to decrease by more than 90% in four asexual generations [
18]; it is likely that multiple sprays have resulted in the development of resistant strains. Moreover, famoxadone exerts stronger selective pressure than azoxystrobin [
18], potentially reducing the effectiveness of tank mixtures with cymoxanil in mitigating resistance development. Considering the dominant nature of QoI resistance [
19], the use of QoI fungicides for grapevine downy mildew management in Japan is currently discouraged.
This figure shows the procedure of the PCR-RFLP method for the detection of QoI fungicide-resistant downy mildew. The lower part of the figure shows the main procedure, and the orange speech bubble indicates the susceptible and resistant cytochrome b genes. These genes are amplified by PCR and digested by Ita I, the restriction enzyme corresponding to the resistant gene, to identify resistant downy mildew.
4. CAA Fungicide Resistance
Dimethomorph, the first CAA fungicide, has been used for grapevine downy mildew control since the late 1980s. The emergence of CAA-resistant strains was first documented in France in 1994 [
20], with subsequent detections reported across numerous European countries [
21]. CAA-resistant isolates have also been identified in Asian countries, including India and China [
22,
23]. The FRAC CAA Working Group acknowledges the frequent detection of resistant strains in France, Germany, Switzerland, Italy, and other countries. However, they suggest that CAA fungicides can still be effective when thorough preventive or prophylactic spraying is implemented. The rapid decline in CAA fungicide efficacy can be masked by their frequent combination with other fungicides that have distinct modes of action, such as fosetyl-aluminium. Upon its initial market introduction in 1997, dimethomorph was prohibited for application to fruits, particularly peaches, in Japan; it was only approved for use on grapes. Consequently, the use of dimethomorph in grape vineyards adjacent to peach orchards was restricted or completely avoided due to concerns over spray drift. However, the approval of mandipropamid, a novel CAA fungicide for peach powdery mildew control, in 2009 marked a significant shift. This approval coincided with the removal of QoI fungicides from the recommended grapevine disease management guidelines due to widespread resistance. As a result, the use of CAA fungicides in grapevine fields substantially increased.
By 2024, Japanese viticulture relied on three CAA fungicides for grapevine downy mildew control: mandipropamid, benthiavalicarb-isopropyl, and dimethomorph. Notably, the 2009 introduction of mandipropamid in Japan coincided with the detection of mandipropamid-resistant downy mildew strains in Europe [
24]. This prompted discussions within the FRAC CAA Working Group regarding the potential risk of fungicide resistance in Japanese grapevine mildew populations, given the recent introduction of CAA fungicides [
25]. In response to these concerns, genetic-based monitoring of fungicide-resistant downy mildew was initiated in Yamanashi Prefecture, a region where CAA fungicide application was recommended [
26,
27]. Mandipropamid resistance in
P. viticola has been linked to a single amino acid substitution at codon 1105 of the
PvCesA3 cellulose synthase gene [
28]. However, the recessive nature of this genetic mutation necessitates homozygosity for resistance expression [
24].
Building on this knowledge, an ASP-PCR (allele specific primer–polymerase chain reaction) assay was developed to rapidly detect the CAA fungicide resistance-associated gene mutation in
P. viticola [
27]. The ASP-PCR method is a variant of the PCR method that takes advantage of the property of PCR primers to deviate from the template DNA depending on the number of mismatches between the PCR primers and the template DNA [
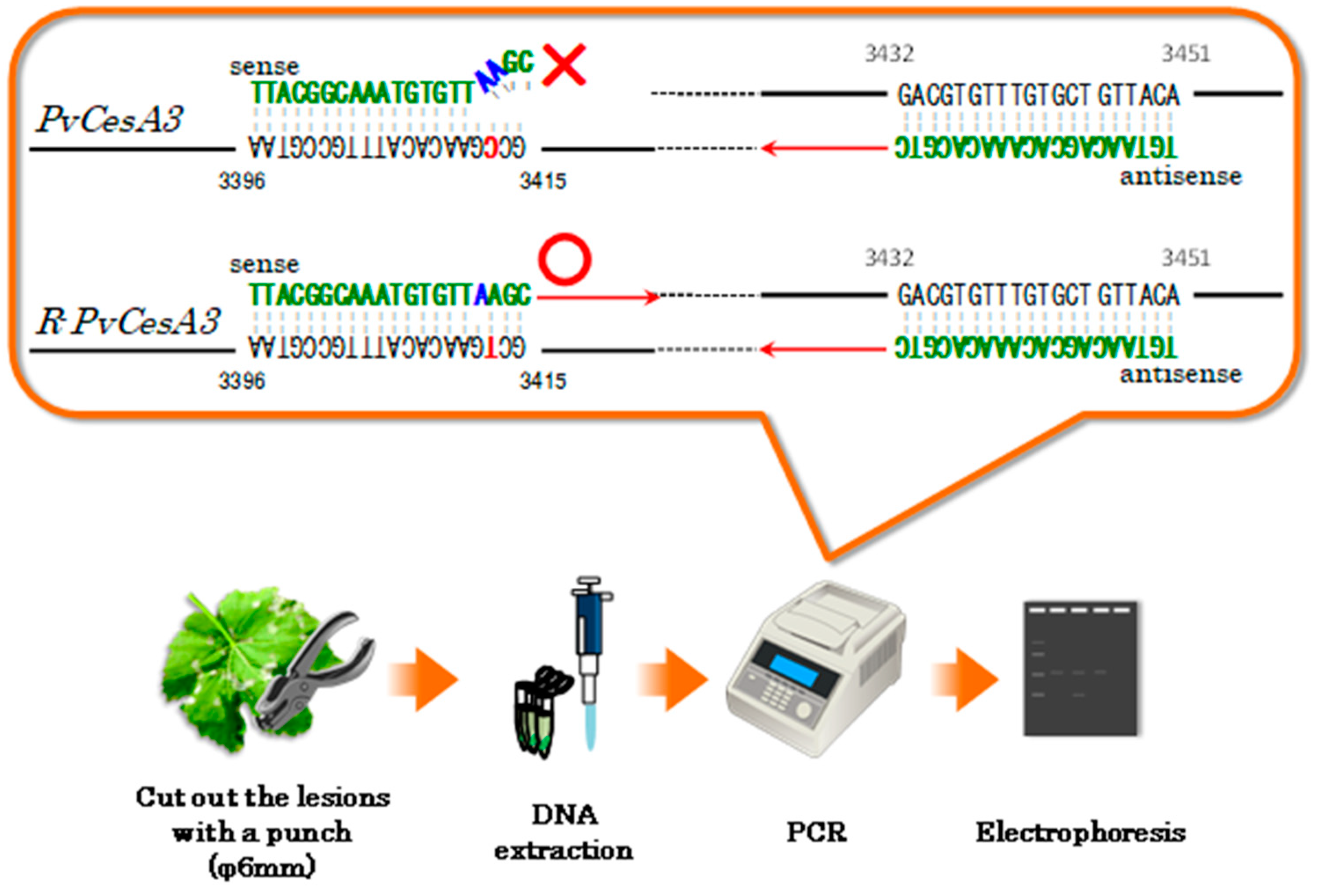
28]. Generally, the divergence of PCR primers from the template DNA occurs when there are two or more mismatch sequences in a row, and DNA polymerase stops DNA synthesis at this divergence. Using this property of DNA polymerase, we developed an ASP-PCR method that can detect the G1105S mutation, which imparts CAA fungicide resistance acquisition (
Figure 4). To differentiate between CAA fungicide-resistant homozygotes and susceptible heterozygous carrying the mutation, a diagnostic scheme incorporating both PCR-RFLP and ASP-PCR analyses was established [
26]. Aoki et al. reported that all 134 samples collected in Yamanashi Prefecture during the 2010 and 2011 monitoring campaigns were homozygotes susceptible to CAA fungicides based on ASP-PCR analysis, with no evidence of resistance-conferring gene mutations [
29]. However, a subsequent analysis of 80 samples in 2012 detected the target gene mutation in a single isolate [
29]. Further ASP-PCR analysis of 157 samples in 2013 identified the mutant gene in five isolates. PCR-RFLP and leaf-disc bioassays confirmed that these isolates were heterozygous for CAA fungicide susceptibility [
29]. The phenotype of this CAA fungicide-resistant heterozygous is the same as that of susceptible homozygotes. Grapevine plots harboring these mutant isolates had a history of intensive CAA fungicide applications. The presence of multiple CAA fungicide-susceptible heterozygotes within a single plot underscores the heightened risk of homozygotes development through natural hybridization [
24].
Although unpublished data from Aoki et al. [
30] have confirmed the presence of CAA fungicide-resistant homozygotes in Yamanashi Prefecture, the phenomenon of resistance gene silencing has also been observed. As reported by the FRAC CAA Working Group [
19], subsequent unpublished data [
30] confirmed resistance gene silencing as a result of a two-year CAA fungicide failure in Yamanashi, Japan, because CAA fungicide-resistant strains are recessive homozygotes.
Given this knowledge, continuous monitoring of the distribution and frequency of fungicide resistance genes is crucial for managing the emergence of recessive resistance traits. High-throughput genetic diagnostic methods are essential for effective surveillance.
This figure shows the procedure of the ASP-PCR method for the detection of CAA fungicide-resistant downy mildew. The lower part of the figure shows the main procedure, and the orange speech bubble indicates the susceptible and resistant PvCesA3 genes. These genes are amplified by PCR to identify resistant PvCesA3 genes of downy mildew.
5. Oxathiapiprolin Fungicide Resistance
Oxathiapiprolin is the most recently introduced fungicide in Japanese viticulture for managing grapevine mildew. Classified as an OSBPI fungicide, oxathiapiprolin targets oxysterol-binding proteins, which are essential for the intracellular transport of sterols, key components of cell membranes and organelles. By inhibiting these proteins, oxathiapiprolin disrupts lipid formation, a critical process for cellular maintenance, including intracellular membrane signaling and cell membrane biogenesis. This ultimately inhibits sporocyst formation and the germination of spores. Oxathiapiprolin was registered and marketed as a fungicide in Japan in 2016, following its commercialization in France and other European countries in 2012. According to a 2018 FRAC survey, reduced susceptibility to oxathiapiprolin has been documented in several oomycetes, including the pathogens responsible for potato blight and cucumber downy mildew. Furthermore, the ease with which resistant mutants have been generated in laboratory studies involving pepper blight and strawberry blight oomycetes underscores the susceptibility of oomycetes to developing oxathiapiprolin resistance [
31,
32]. Consequently, FRAC has categorized the risk of oxathiapiprolin resistance as medium to high.
At present, no reports of oxathiapiprolin-resistant grapevine downy mildew strains have emerged in Japan. To mitigate the risk of resistance development, the use of oxathiapiprolin as a standalone fungicide has been discontinued in Japan. Instead, combination products featuring oxathiapiprolin with either famoxadone hydrate or mancozeb hydrate have been developed.
6. IPM and Monitoring Fungicide Resistance in Obligate Plant Pathogens
Monitoring fungicide resistance in obligate plant pathogens, such as grapevine powdery mildew (
Erysiphe necator) and grapevine downy mildew, presents significant logistical challenges due to the requirement for host plants [
15]. In Japanese viticulture, genetic diagnostics for grapevine downy mildew fungicide resistance have been established [
8,
26]. Genetic diagnosis is crucial for tracking the distribution and frequency of recessive fungicide resistance traits, such as those associated with CAA fungicides. High-throughput genetic diagnostic methods are well-suited for this task. Similar diagnostic tools have been developed for detecting sterol demethylation inhibitor- and QoI fungicide-resistant strains of grapevine powdery mildew [
33,
34].
Effective genetic diagnosis requires a clear understanding of the underlying genetic mechanisms conferring fungicide resistance, including the specific genes and mutation sites involved. For instance, oxathiapiprolin resistance in
Phytophthora capsici has been attributed to a point mutation in the oxysterol-binding protein gene, resulting in a glycine-to-tryptophan substitution at position 769 [
31]. However, this specific gene is absent in grapevine downy mildew, necessitating an alternative approach for resistance diagnosis. In 2022, two grapevine downy mildew isolates exhibiting reduced susceptibility to oxathiapiprolin were identified in an Italian vineyard with a history of intensive oxathiapiprolin applications. Genetic analysis revealed identical mutations in the
PvORP1 gene, a putative oxysterol-binding protein [
35]. Notably, oxathiapiprolin-insensitive downy mildew strains had already been detected across Europe even before the development of genetic diagnostic tools [
36].
On the other hand, the concept of Integrated Pest Management (IPM) has begun to attract attention from producers who are concerned about reducing the risk of fungicide-resistant downy mildew caused by chemical agents and the impression it may have on consumers. IPM is the concept of “taking appropriate measures to reduce or minimize risks to humans and the environment in a comprehensive manner, while taking into account economic efficiency, by using all available pest control technologies” [
37]. In IPM, chemical agents that have been used as core pest control are used only as a supplement to the existing chemical agents, which enables a smooth transition to fungicide-reduced cultivation by reducing the number of chemical agents used without difficulty. This is expected to reduce the emergence of drug-resistant fungi and oomycetes. In Japanese viticulture, the development of microbial fungicides against filamentous fungal diseases is also being pursued [
38,
39,
40]. Monitoring for downy mildew and other diseases in vineyards is essential for the effective use of these microbial fungicides.
Given this knowledge, the effective management of fungicide resistance requires advance monitoring for the emergence of resistant strains using the genetic diagnostic method as soon as the efficacy of the method is perceived to be declining and detecting the emergence of resistant strains before they become widespread. To facilitate such monitoring, agrochemical manufacturers should consider sharing data to the best of their knowledge, such as the target sites of novel fungicides, with researchers.
7. Conclusions and Prospects
The 2019 FRAC assessment classified grapevine downy mildew as a high-risk pathogen for fungicide resistance development. The rapid emergence and spread of resistant strains have severely limited effective chemical control options in Japanese viticulture. Even oxathiapiprolin, the newest fungicide introduced in Japan, has shown reduced efficacy against grapevine downy mildew in Europe [
34,
35]. While no resistance cases have been reported in Japan, the historical pattern of resistance development suggests that oxathiapiprolin resistance is highly probable.
To combat fungicide resistance, chemical control strategies should meet two key criteria: avoiding cross-resistance between combined fungicides and ensuring resistance reversibility after fungicide discontinuation through monitoring. However, these theoretical principles face practical challenges: many commercial fungicides share similar modes of action, and the dominant inheritance of resistance genes in grapevine downy mildew often prevents the restoration of fungicide susceptibility even after use cessation. A more practical approach involves diversifying fungicide applications by using multiple agents with distinct mechanisms of action, as has been performed for the past 40 years for many crops worldwide [
41]. For instance, fungicides with low resistance risk, such as copper sulfate or dithiocarbamates, serve as primary agents. CAA fungicides and oxathiapiprolin can then be alternated biennially during high-risk periods (e.g., June’s high humidity and temperature conditions). While this strategy cannot completely prevent disease development, it significantly reduces incidence compared to annual applications of the same fungicides [
19,
35,
42].
Japanese guidelines require strict adherence to a rotation of fungicides from different strains every two years. However, maintaining this rotation system without introducing novel fungicides periodically will become increasingly challenging as resistant strains continue to emerge. Therefore, continuous monitoring of fungicide resistance remains essential.






{kind=link}
{kind=link}
{kind=link}
{kind=link}