
Characterization and Whole-Genome Sequencing of Phytobacter palmae WL65, a Plant Growth-Promoting Rhizobacterium First Isolated from Rice Rhizosphere Soil in Thailand


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Physiological and Biochemical Characteristics
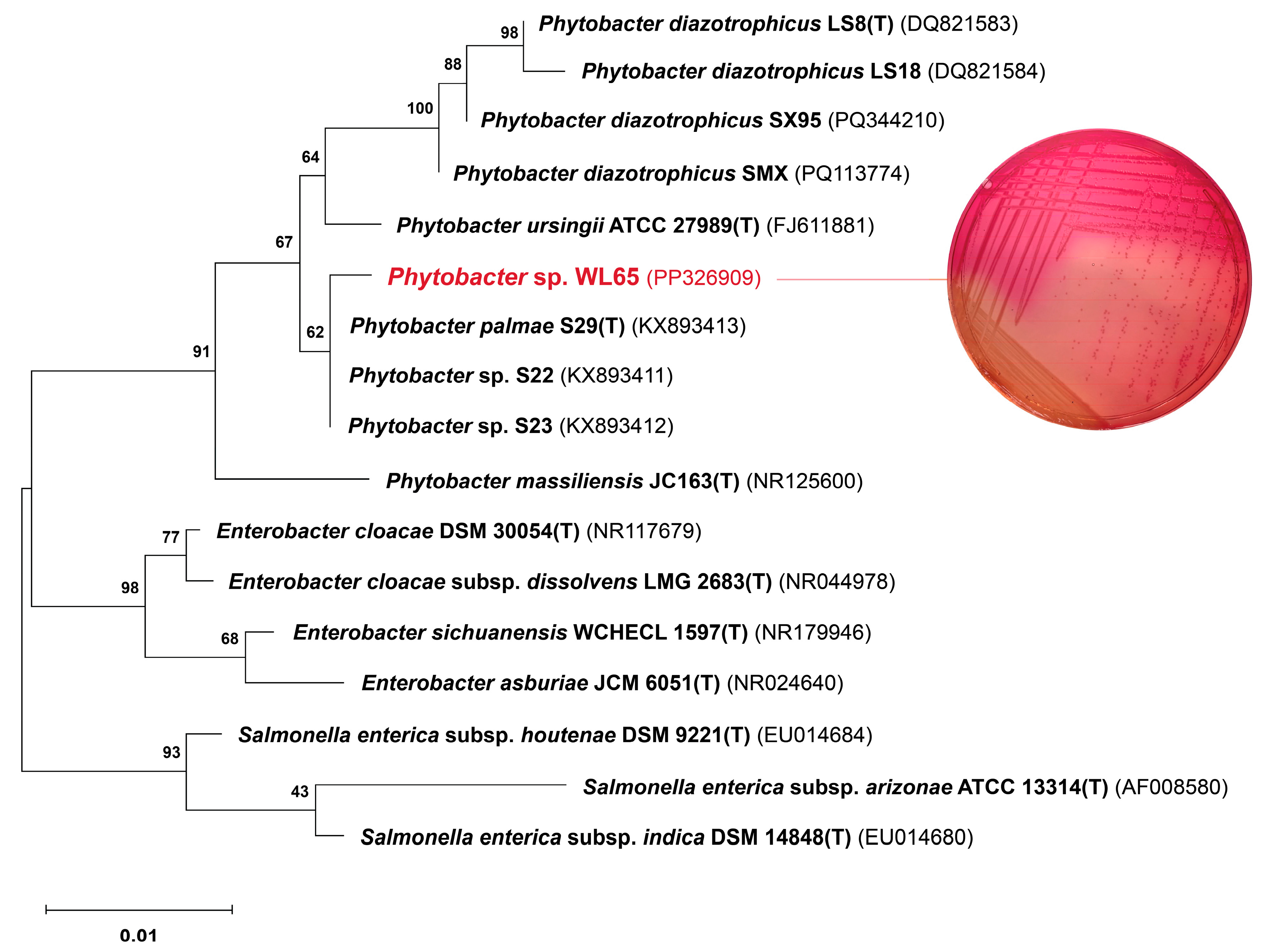
2.3. 16S rRNA Gene Sequences and Phylogenetic Analysis
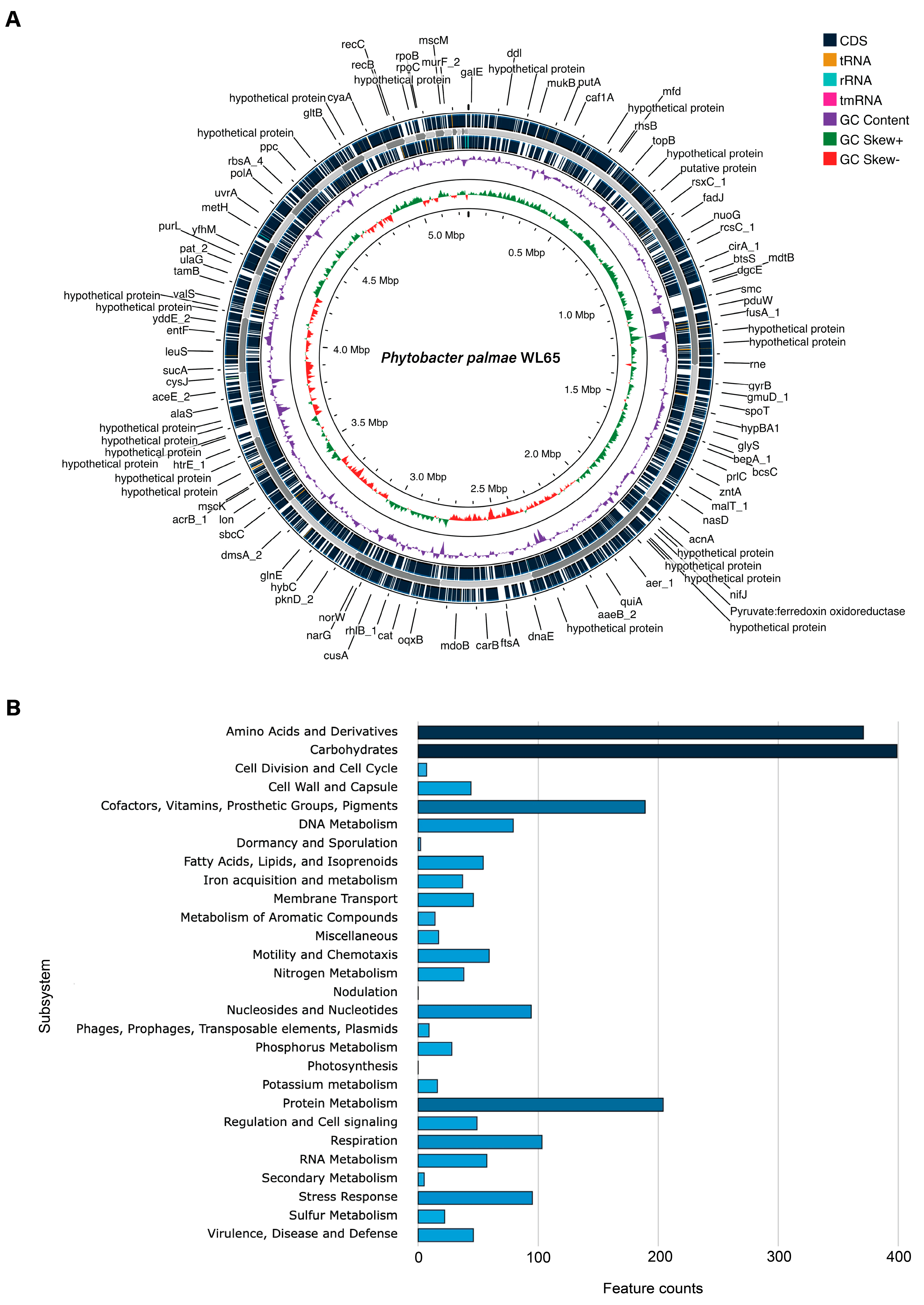
2.4. Genome Sequencing and Genome Annotation
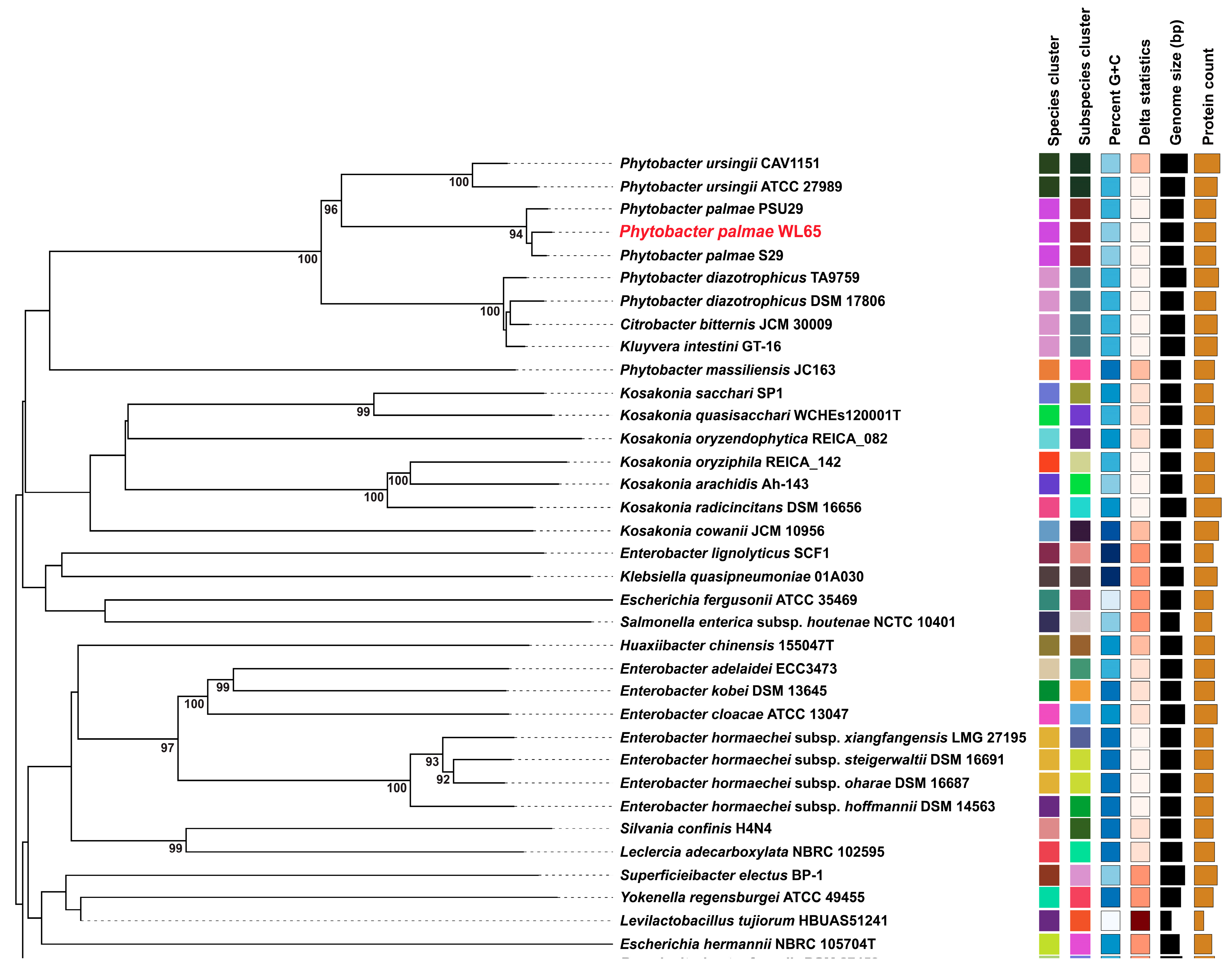
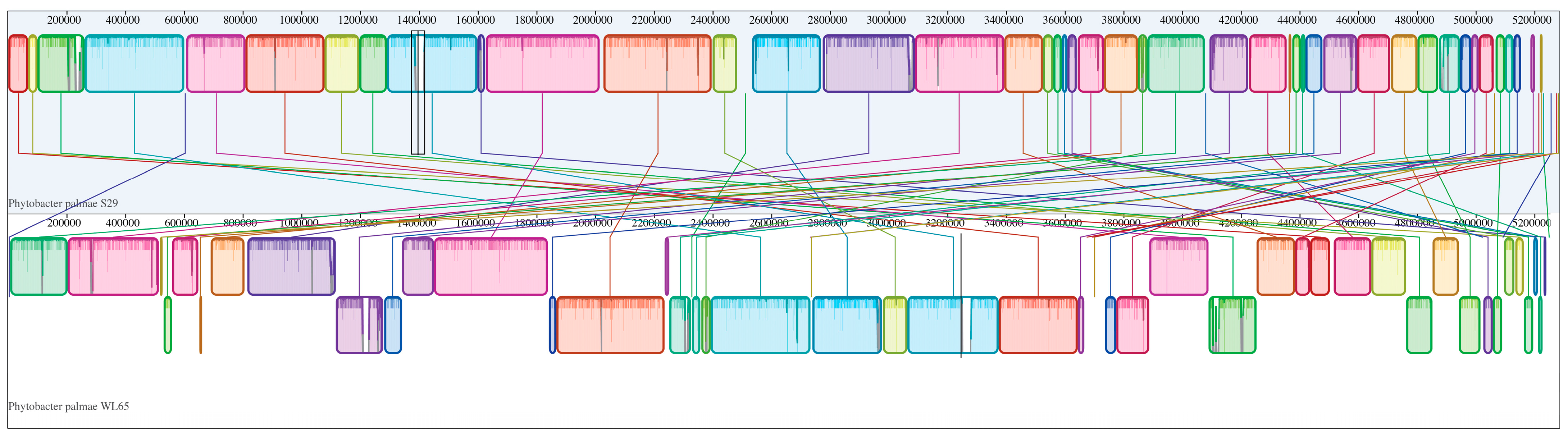
2.5. Genome Mining and Phylogenomic Analysis
2.6. Phytobacter Palmae WL65 as a Plant Growth-Promoting Rhizobacterium for Rice Cultivation in Pot Experiments
2.6.1. Bacterial Colonization on Rice Roots
2.6.2. Effects of P. palmae WL65 on Rice Cultivar RD79 in Pot Experiment
2.7. Statistical Analysis
3. Results
3.1. Biochemical Characteristics and 16S rRNA Gene Sequence Analysis
3.2. Genome Sequencing and Phylogenomics Analysis
3.3. Genome Analysis of P. palmae WL65 as Plant Growth-Promoting Rhizobacterium
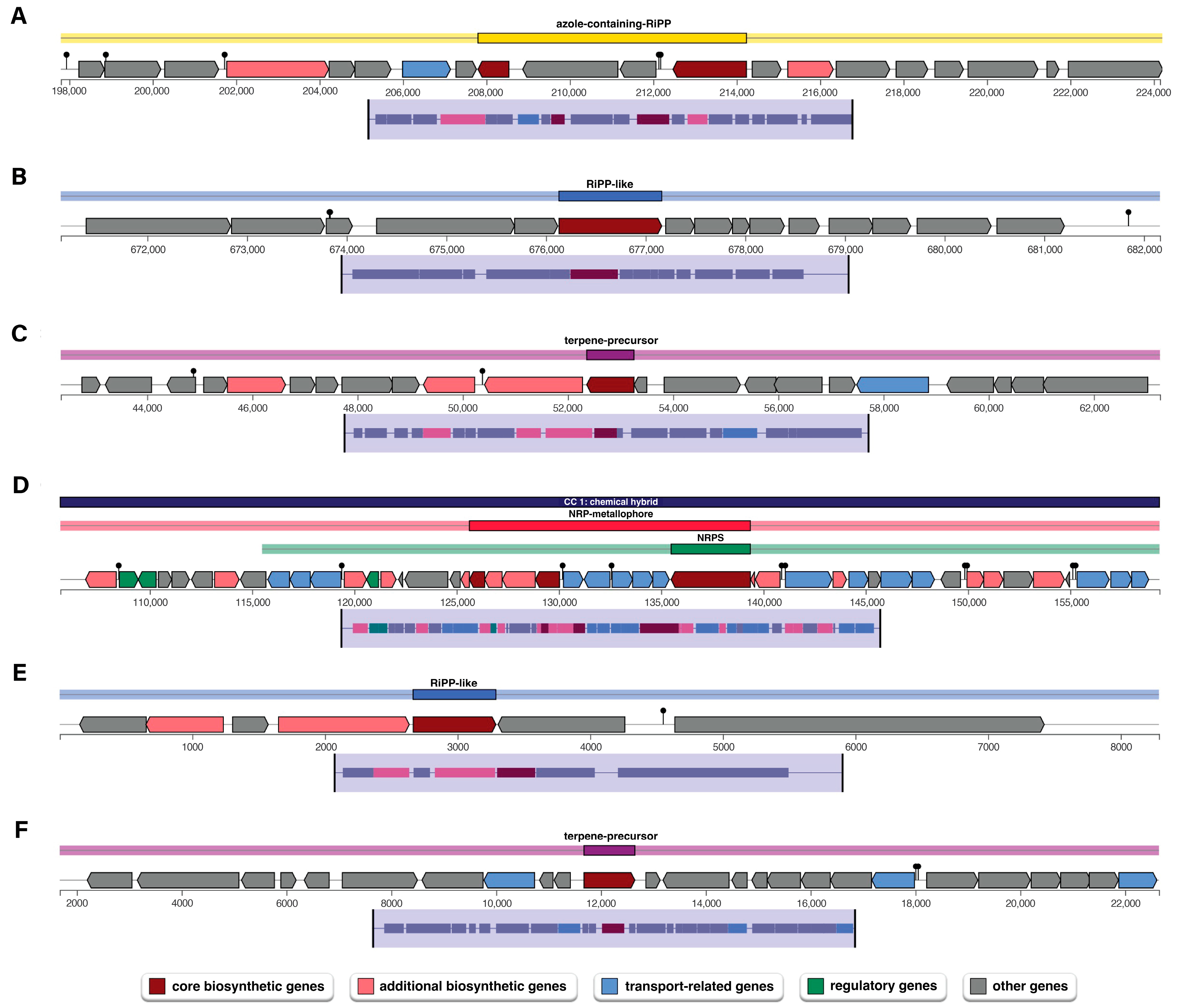
3.4. Genome Analysis of Secondary Metabolism of P. palmae WL65 Using AntiSMASH
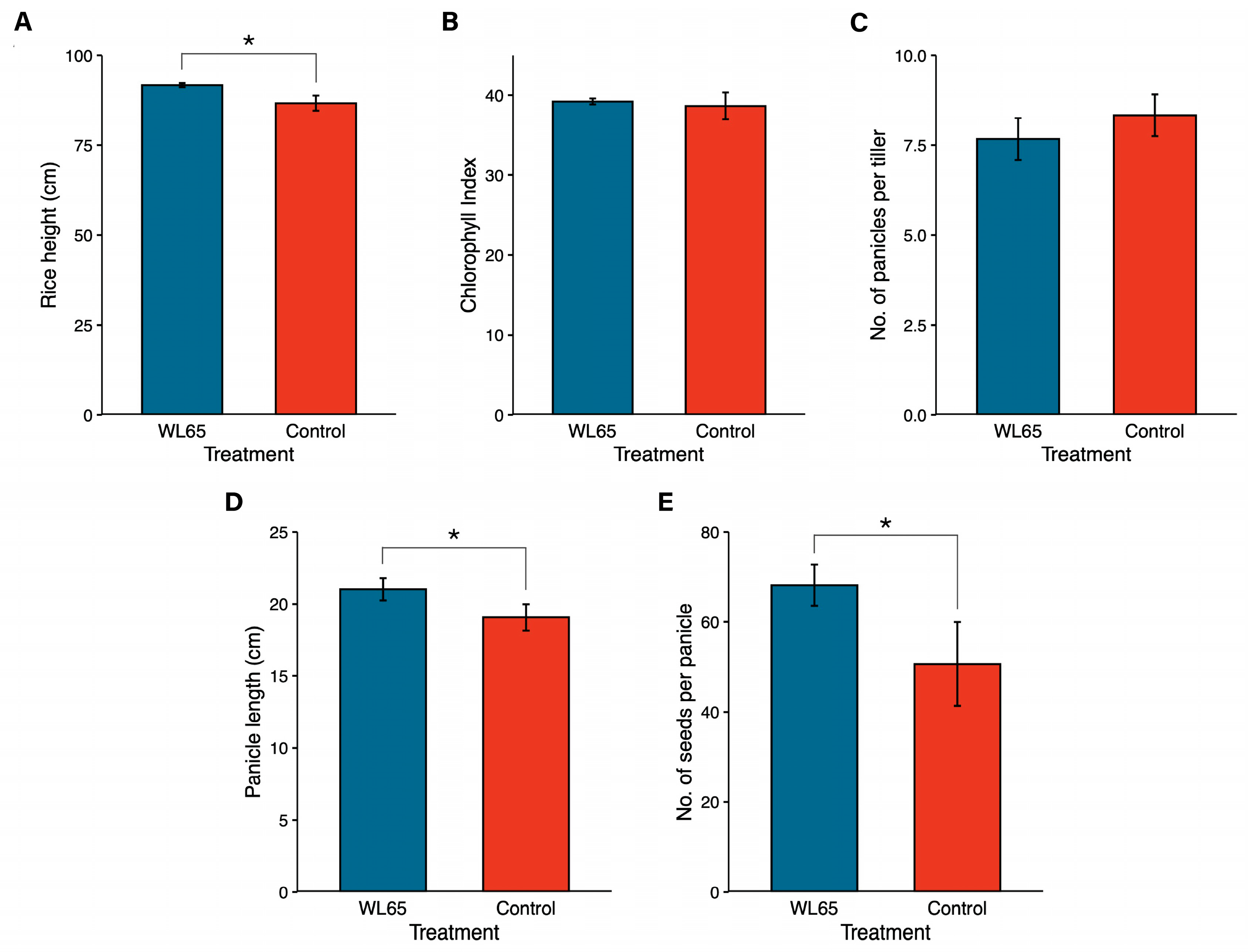
3.5. P. palmae WL65 as a Plant Growth-Promoting Rhizobacterium (PGPR) for Rice Cultivation in Pot Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, G.X.; Peng, G.X.; Wang, E.T.; Yan, H.; Yuan, Q.H.; Zhang, W.; Lou, X.; Wu, H.; Tan, Z.Y. Diverse endophytic nitrogen-fixing bacteria isolated from wild rice Oryza rufipogon and description of Phytobacter diazotrophicus gen. nov. sp. nov. Arch. Microbiol. 2008, 189, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Madhaiyan, M.; Saravanan, V.S.; Blom, J.; Smits, T.H.; Rezzonico, F.; Kim, S.-J.; Weon, H.-Y.; Kwon, S.-W.; Whitman, W.B.; Ji, L. Phytobacter palmae sp. nov., a novel endophytic, N2 fixing, plant growth promoting Gamma proteobacterium isolated from oil palm (Elaeis guineensis Jacq.). Int. J. Syst. Evol. Microbiol. 2020, 70, 841–848. [Google Scholar] [CrossRef]
- Pillonetto, M.; Arend, L.N.; Faoro, H.; D’Espindula, H.R.; Blom, J.; Smits, T.H.; Mira, M.T.; Rezzonico, F. Emended description of the genus Phytobacter, its type species Phytobacter diazotrophicus (Zhang 2008) and description of Phytobacter ursingii sp. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yao, R.; Li, Y.; Wu, X.; Li, S.; An, Q. Proposal for unification of the genus Metakosakonia and the genus Phytobacter to a single genus Phytobacter and reclassification of Metakosakonia massiliensis as Phytobacter massiliensis comb. nov. Curr. Microbiol. 2020, 77, 1945–1954. [Google Scholar] [CrossRef]
- Almuzara, M.; Cittadini, R.; Traglia, G.; Haim, M.S.; De Belder, D.; Alvarez, C.; O’Connor, Z.; Ocampo, C.V.; Barberis, C.; Prieto, M. Phytobacter spp: The emergence of a new genus of healthcare-associated Enterobacterales encoding carbapenemases in Argentina: A case series. Infect. Prev. Pract. 2024, 6, 100379. [Google Scholar] [CrossRef]
- Thamvithayakorn, P.; Phosri, C.; Robinson-Boyer, L.; Limnonthakul, P.; Doonan, J.H.; Suwannasai, N. The synergistic impact of a novel plant growth-promoting rhizobacterial consortium and Ascophyllum nodosum seaweed extract on rhizosphere microbiome dynamics and growth enhancement in Oryza sativa L. RD79. Agronomy 2024, 14, 2698. [Google Scholar] [CrossRef]
- Aslanzadeh, J. Biochemical profile-based microbial identification systems. In Advanced Techniques in Diagnostic Microbiology; Springer: Boston, MA, USA, 2006; pp. 84–116. [Google Scholar]
- Brady, C.; Cleenwerck, I.; Venter, S.; Vancanneyt, M.; Swings, J.; Coutinho, T. Phylogeny and identification of Pantoea species associated with plants, humans and the natural environment based on multilocus sequence analysis (MLSA). Syst. Appl. Microbiol. 2008, 31, 447–460. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes de novo assembler. Curr. Protoc. Bio Inform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.-y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Yaikhan, T.; Suwannasin, S.; Singkhamanan, K.; Chusri, S.; Pomwised, R.; Wonglapsuwan, M.; Surachat, K. Genomic characterization of multidrug-resistant Enterobacteriaceae clinical isolates from Southern Thailand hospitals: Unraveling antimicrobial resistance and virulence mechanisms. Antibiotics 2024, 13, 531. [Google Scholar] [CrossRef]
- Gong, F.; Yanofsky, C. Analysis of tryptophanase operon expression in vitro: Accumulation of TnaC-peptidyl-tRNA in a release factor 2-depleted S-30 extract prevents Rho factor action, simulating induction. J. Biol. Chem. 2002, 277, 17095–17100. [Google Scholar] [CrossRef]
- Moosavian, M.; Khoshkholgh Sima, M.; Ahmad Khosravi, N.; Abbasi Montazeri, E. Detection of OqxAB efflux pumps, a multidrug-resistant agent in bacterial infection in patients referring to teaching hospitals in Ahvaz, Southwest of Iran. Int. J. Microbiol. 2021, 2021, 2145176. [Google Scholar] [CrossRef]
- Bharatham, N.; Bhowmik, P.; Aoki, M.; Okada, U.; Sharma, S.; Yamashita, E.; Shanbhag, A.P.; Rajagopal, S.; Thomas, T.; Sarma, M. Structure and function relationship of OqxB efflux pump from Klebsiella pneumoniae. Nat. Commun. 2021, 12, 5400. [Google Scholar] [CrossRef]
- Raymond, K.N.; Dertz, E.A.; Kim, S.S. Enterobactin: An archetype for microbial iron transport. Proc. Natl. Acad. Sci. USA 2003, 100, 3584–3588. [Google Scholar] [CrossRef]
- Timofeeva, A.M.; Galyamova, M.R.; Sedykh, S.E. Bacterial siderophores: Classification, biosynthesis, perspectives of use in agriculture. Plants 2022, 11, 3065. [Google Scholar] [CrossRef]
- Chuchert, S.; Thongmak, J.; Daungnamkaew, B.; Sudpesrot, P.; Nakorn, W.N.; Marnmad, K.; Khomphet, T. Optimizing Temperature and Drying Conditions to Break Dormancy in Thai RD79 Rice Seeds. Indian J. Agric. Res. 2025, 1–9. [Google Scholar] [CrossRef]
- Itoh, Y.; Rice, J.D.; Goller, C.; Pannuri, A.; Meisner, J.J.; Beveridge, T.J.; Preston III, J.F.; Rom, T. Roles of pgaABCD genes in synthesis, modification, and export of the Escherichia coli biofilm adhesin poly-β-1,6-N-acetyl-D-glucosamine. J. Bacteriol 2008, 190, 3670–3680. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.J.; Tu, I.F.; Tseng, T.S.; Tsai, Y.H.; Wu, S.H. The deficiency of poly-β-1,6-N-acetyl-glucosamine deacetylase trigger A. baumannii to convert to biofilm-independent colistin-tolerant cells. Sci. Rep. 2023, 13, 2800. [Google Scholar] [CrossRef]
- Ajijah, N.; Fiodor, A.; Pandey, A.K.; Rana, A.; Pranaw, K. Plant growth-promoting bacteria (PGPB) with biofilm-forming ability: A multifaceted agent for sustainable agriculture. Diversity 2023, 15, 112. [Google Scholar] [CrossRef]
- Rafique, M.; Naveed, M.; Mumtaz, M.Z.; Niaz, A.; Alamri, S.; Siddiqui, M.H.; Waheed, M.Q.; Ali, Z.; Naman, A.; Rehman, S.U. Unlocking the potential of biofilm-forming plant growth-promoting rhizobacteria for growth and yield enhancement in wheat (Triticum aestivum L.). Sci. Rep. 2024, 14, 15546. [Google Scholar] [CrossRef]
- Lata, D.L.; Abdie, O.; Rezene, Y. IAA-producing bacteria from the rhizosphere of chickpea (Cicer arietinum L.): Isolation, characterization, and their effects on plant growth performance. Heliyon 2024, 10, e39702. [Google Scholar] [CrossRef]
- Shabtai, I.A.; Wilhelm, R.C.; Schweizer, S.A.; Höschen, C.; Buckley, D.H.; Lehmann, J. Calcium promotes persistent soil organic matter by altering microbial transformation of plant litter. Nat. Commun. 2023, 14, 6609. [Google Scholar] [CrossRef]
- Wang, C.; Luan, S. Calcium homeostasis and signaling in plant immunity. Curr. Opin. Plant Biol. 2024, 77, 102485. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Characteristics | WL65 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|---|
| Beta-galactosidase | + | + | + | + | + |
| Arginine dihydrolase | - | - | - | - | - |
| Lysine decarboxylase | - | - | - | - | - |
| Ornithine decarboxylase | - | - | - | - | + |
| Citrate utilization | + | + | + | + | - |
| H2S production | - | - | - | - | - |
| Urease | - | - | - | - | - |
| Tryptophan deaminase | - | + | - | - | - |
| Indole production | + | + | + | + | + |
| Acetoin production (Voges-Proskauer) | - | - | + | + | - |
| Gelatinase | - | - | - | - | + |
| Fermentation-oxidation of: | |||||
| Glucose | + | + | + | + | + |
| Mannitol | + | + | + | + | + |
| Inositol | - | - | - | - | - |
| Sorbitol | + | + | + | + | + |
| Rhamnose | + | + | + | + | + |
| Saccharose | + | + | + | + | - |
| Melibiose | - | - | - | - | - |
| Amygdalin | + | + | + | + | + |
| Arabinose | + | + | + | + | + |
| Cytochrome oxidase | - | - | - | - | - |
| Statistics | P. palmae WL65 | P. palmae S29T 1 |
|---|---|---|
| Genome size (bp) | 5,253,439 | 5,284,330 |
| G + C content (%) | 52.51 | 52.60 |
| Number of contigs | 33 | ND |
| Number of coding sequences | 5210 | ND |
| Number of tRNA | 58 | 70 |
| Number of rRNA | 4 | 13 |
| Number of subsystems | 347 | ND |
| N50 contig size (bp) | 340,721 | ND |
| L50 | 5 | ND |
| Plasmids | 0 | ND |
| Hypothetical proteins | 1071 | ND |
| Proteins with functional assignments | 4139 | ND |
| Proteins with EC number assignments | 1307 | ND |
| Proteins with GO assignments | 1080 | ND |
| Proteins with pathway assignments | 937 | ND |
| PGP Traits | Genes | Functions |
|---|---|---|
| Nitrogen fixation | nifA, nifB, nifD, nifE, nifF, nifH, nifJ, nifK, nifL, nifS, nifU, nifW, nifX | Nitrogenase (molybdenum-iron)-specific transcriptional regulator; nitrogenase molybdenum-iron protein alpha chain; nitrogenase FeMo-cofactor carrier protein, etc. |
| Phosphate solubilization | pstA, pstB, pstC, pstS, phnC, phnD, phnE, phnV | Phosphate transport system permease protein; phosphate import ATP-binding protein; phosphate-binding protein; etc. |
| Siderophore production | fhuA, fhuB, fhuC, fhuD, fhuF, feoA, feoB, feoC, acrA, acrB, acrE, acrR, acrZ | Ferrichrome outer membrane transporter/phage receptor; Iron(3+)-hydroxamate import system permease protein; etc. |
| Indole-3-acetic acid production | trpA, trpB, trpC, trpE, trpGD, trpR, trpS | Tryptophan synthase alpha chain; tryptophan synthase beta chain; tryptophan biosynthesis protein; anthranilate phosphoribosyltransferase; etc. |
| Potassium solubilization | gcd_1, gcd_2 | Glucose dehydrogenase, PQQ-dependent; quinoprotein glucose dehydrogenase |
| ACC deaminase activity | rimI, rimK, rimL, rimM, rimO, rimP, rimJ | ribosomal protein S18 alanine N-acetyltransferase; ribosomal maturation factor; etc. |
| Hydrogen sulfide production | cysA, cysB, cysC, cysD, cysE, cysG, cysH, cysI, cysJ, cysK, cysL, cysM, cysN, cysP, cysQ, cysS, cysT, cysW, cysZ | Adenylylsulfate kinase; sulfate adenylyltransferase subunit 1/2; thioredoxin-dependent 5′-adenylylsulfate reductase; etc. |
| Quorum sensing | luxR, luxS | Two-component transcriptional response regulator; S-ribosylhomocysteine lyase |
| Superoxide dismutase | sodA, sodB, sodC | Superoxide dismutase [Mn/Fe/Cu-Zn] |
| Biofilm production | pgaA, pgaB, pgaC | biofilm PGA outer membrane secretin; biofilm PGA synthesis deacetylase; biofilm PGA synthesis N-glycosyltransferase |
| Peroxidases | osmB, osmC, osmE, osmV, osmW, osmX, osmY, oxyR | Peroxiredoxin; osmotically-inducible lipoprotein; osmoprotectant import permease protein; etc. |
| Acetoin and butanediol synthesis | budC, alsT, poxB | Diacetyl reductase [(S)-acetoin forming]; amino acid carrier protein; pyruvate dehydrogenase |
| Phenazine production | phzF | Trans-2,3-dihydro-3-hydroxyanthranilate isomerase; phenazine biosynthesis protein |
| Glycine-betaine production | proA, proB, proC, proP, proS, proX, proQ, proV, proW, proY, soxR, soxS | L-Proline/glycine-betaine transporter; glycine-betaine/proline betaine transport system ATP-binding protein, etc. |
| Tryptophanase | tnaA | Tryptophanase |
| OqxAB efflux pump | oqxB | OqxAB efflux pump |
| GABA production | gabR | HTH-type transcriptional regulatory protein |
| Region | BGCs Type | From (nt) | To (nt) | Most Similar Known Cluster |
|---|---|---|---|---|
| 1.1 | azole-containing-RiPP | 197,800 | 224,234 | - |
| 1.2 | RiPP-like | 671,131 | 682,162 | - |
| 8.1 | terpene-precursor | 42,358 | 63,257 | - |
| 10.1 | NRP-metallophore, NRPS | 105,627 | 159,362 | Enterobactin, NRP |
| 14.1 | RiPP-like | 1 | 8289 | - |
| 16.1 | terpene-precursor | 1676 | 22,647 | - |
| Soil Properties | WL65 | Control |
|---|---|---|
| Soil texture | Sandy loam | Sandy loam |
| pH (1:1) | 6.08 | 5.99 |
| Organic matter (%) | 3.40 | 2.81 |
| EC 1:5 (dS m−1) | 0.04 | 0.02 |
| Carbon (%) | 2.66 | 2.67 |
| Nitrogen (%) | 0.26 | 0.26 |
| Available phosphorus (mg kg−1) | 13.99 | 10.11 |
| Potassium (mg kg−1) | 12.12 | 9.34 |
| Calcium (mg kg−1) | 365.80 | 288.00 |
| Magnesium (mg kg−1) | 27.94 | 20.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thamvithayakorn, P.; Phosri, C.; Vishal, V.; Suwannasai, N. Characterization and Whole-Genome Sequencing of Phytobacter palmae WL65, a Plant Growth-Promoting Rhizobacterium First Isolated from Rice Rhizosphere Soil in Thailand. Agriculture 2025, 15, 707. https://doi.org/10.3390/agriculture15070707
Thamvithayakorn P, Phosri C, Vishal V, Suwannasai N. Characterization and Whole-Genome Sequencing of Phytobacter palmae WL65, a Plant Growth-Promoting Rhizobacterium First Isolated from Rice Rhizosphere Soil in Thailand. Agriculture. 2025; 15(7):707. https://doi.org/10.3390/agriculture15070707
Chicago/Turabian StyleThamvithayakorn, Pisit, Cherdchai Phosri, Vineet Vishal, and Nuttika Suwannasai. 2025. "Characterization and Whole-Genome Sequencing of Phytobacter palmae WL65, a Plant Growth-Promoting Rhizobacterium First Isolated from Rice Rhizosphere Soil in Thailand" Agriculture 15, no. 7: 707. https://doi.org/10.3390/agriculture15070707
APA StyleThamvithayakorn, P., Phosri, C., Vishal, V., & Suwannasai, N. (2025). Characterization and Whole-Genome Sequencing of Phytobacter palmae WL65, a Plant Growth-Promoting Rhizobacterium First Isolated from Rice Rhizosphere Soil in Thailand. Agriculture, 15(7), 707. https://doi.org/10.3390/agriculture15070707