

Warming Induces High Environmental Pressure in the Lakeshore Wetland, Promoting Interactions Among Carbon-Fixing Microorganisms

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Soil Sampling
2.2. Soil Physicochemical Properties
2.3. DNA Extraction and MiSeq Sequencing
2.4. Statistical Analysis
3. Results
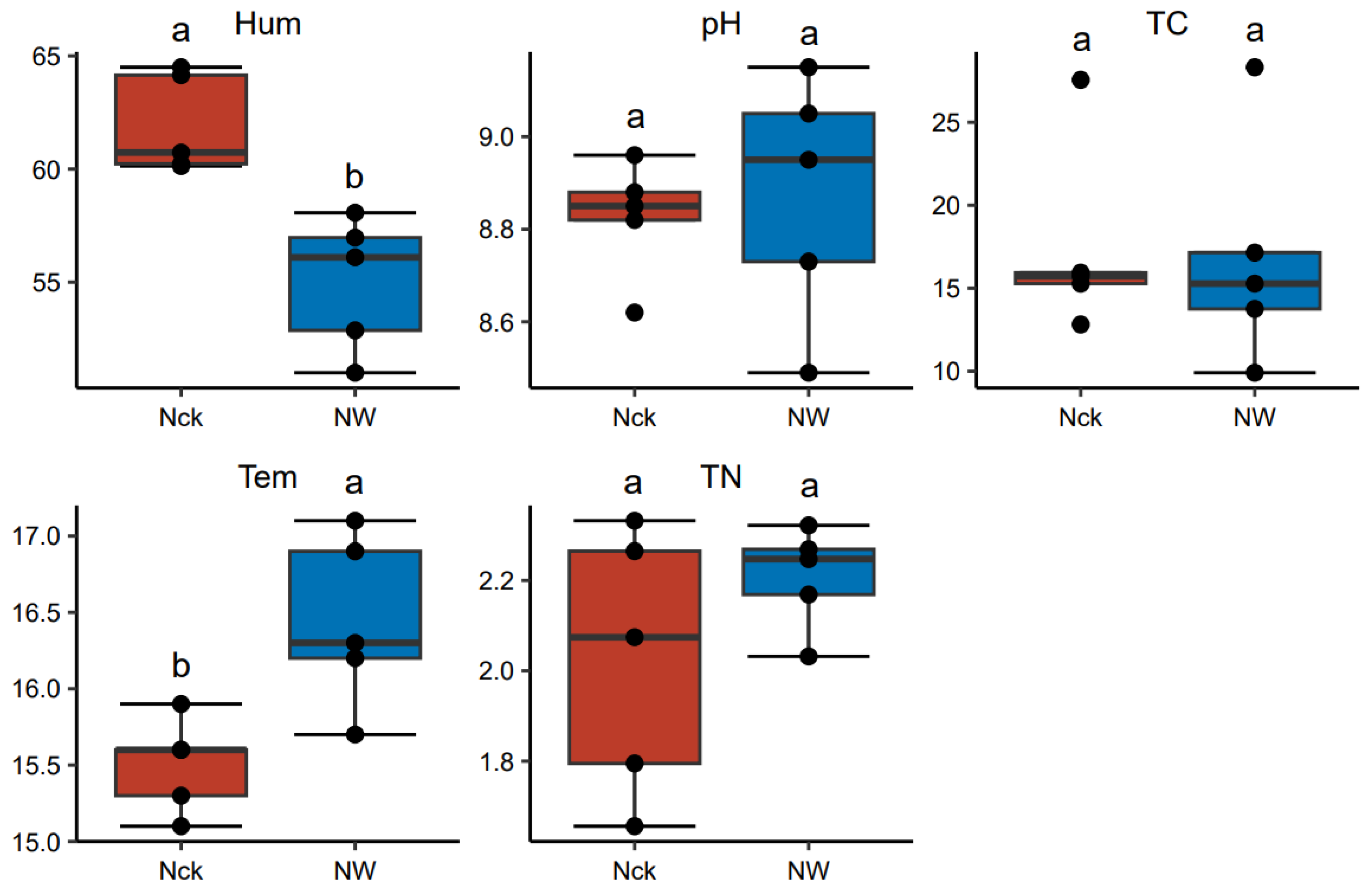
3.1. Soil Physicochemical Properties of the Lakeshore Wetland
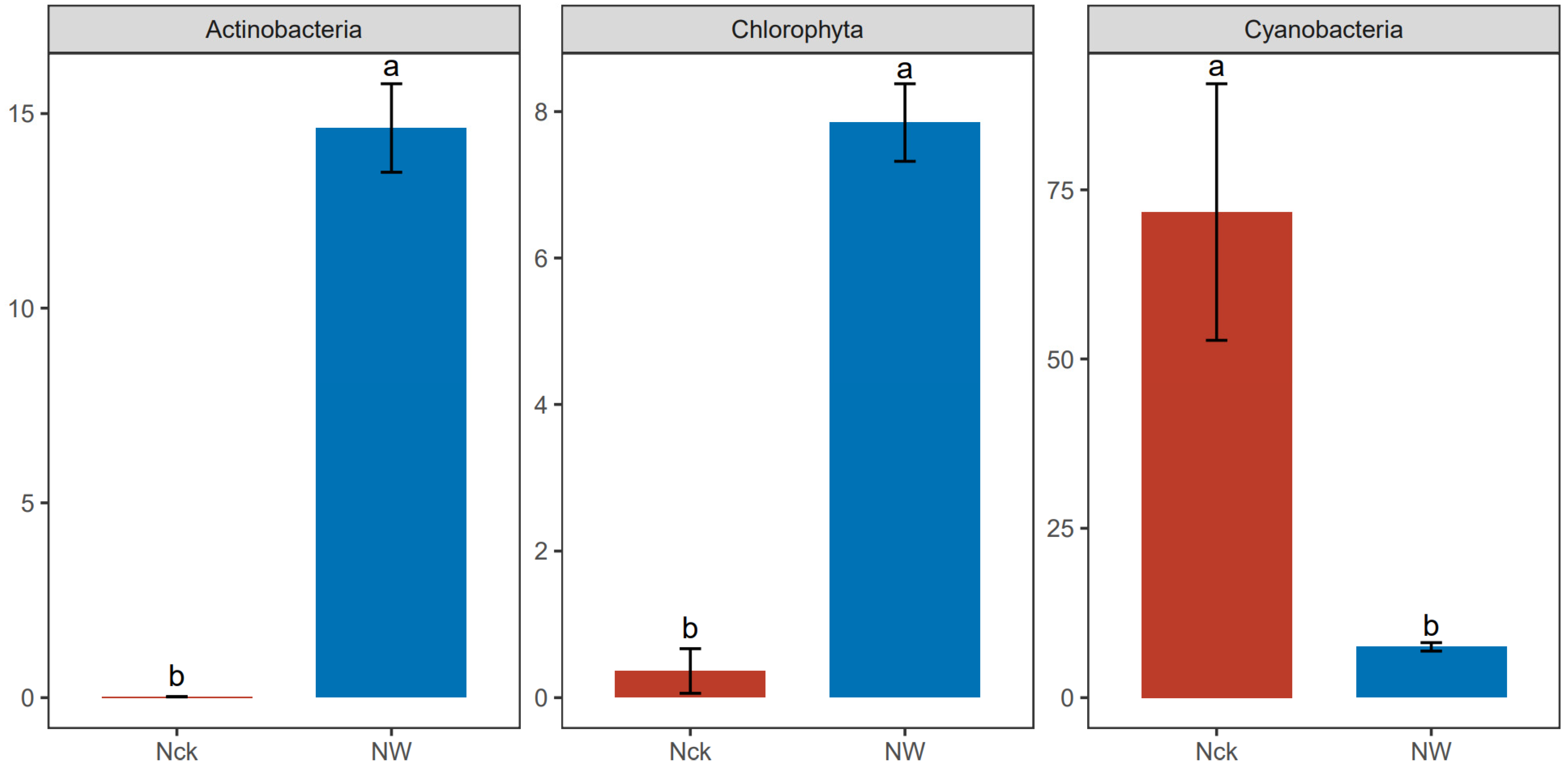
3.2. cbbL Carbon-Fixing Microbial Community Structure
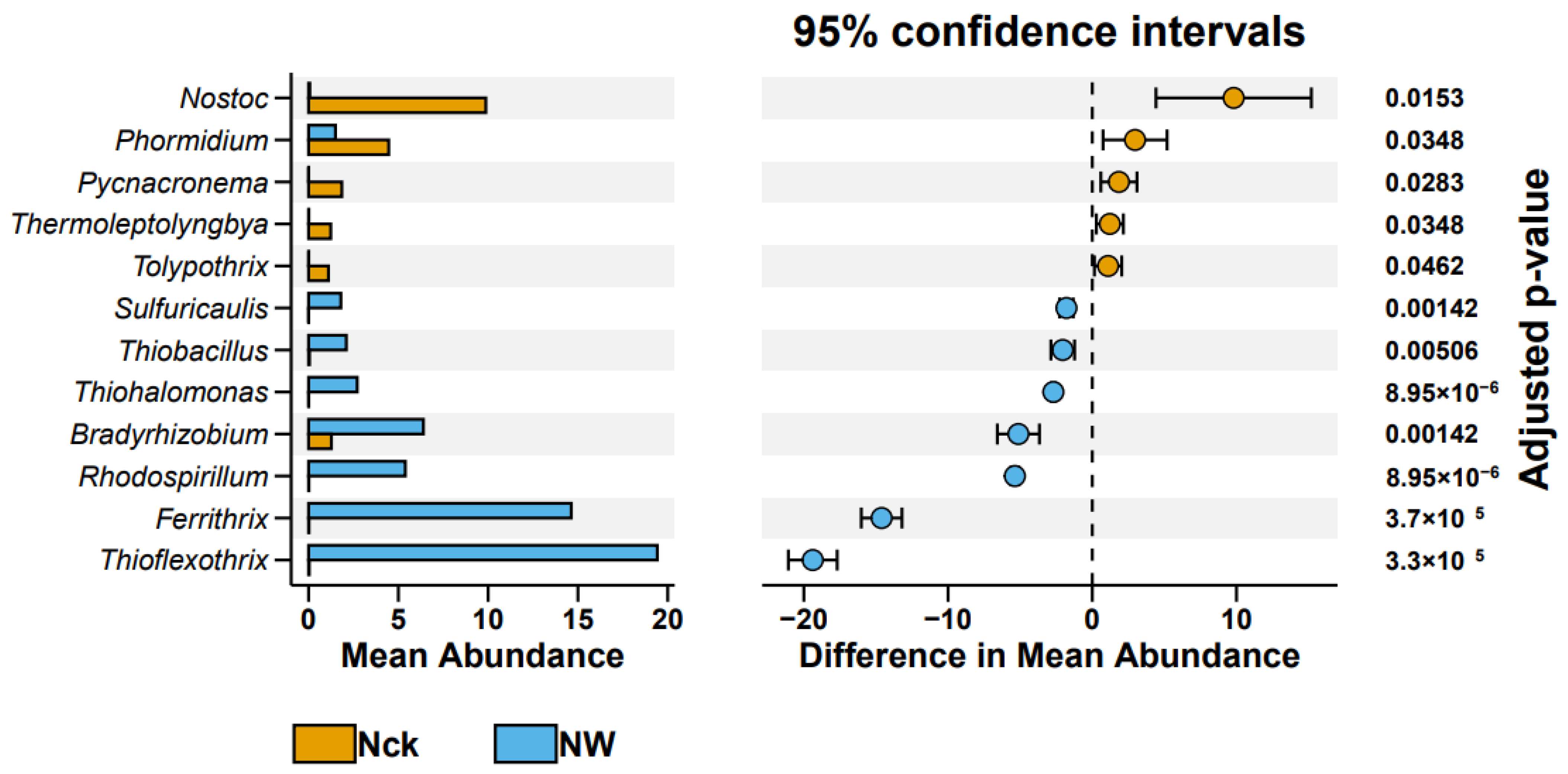
3.3. Community Assembly Mechanisms and the Functional Groups
3.4. Co-Occurrence Network
4. Discussion
4.1. Response of cbbL Carbon-Fixing Microbial Communities in the Lakeshore Wetland to Warming
4.2. Factors Influencing cbbL Carbon-Fixing Microbial Communities and Their Interrelationships in the Lakeshore Wetland
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendia, E., Masson-Delmotte, V., Pörtner, H.O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019; p. 896. [Google Scholar]
- Heimann, M.; Reichstein, M. Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature 2008, 451, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 5th ed.; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar]
- Bloom, A.A.; Bowman, K.W.; Lee, M. A global wetland methane emissions and uncertainty dataset for atmospheric chemical transport models (WetCHARTs version 1.0). Geosci. Model. Dev. 2017, 10, 2141–2156. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.Y.; Jiang, M.; Han, G.X.; Zhang, L.W.; Zhou, J.; Bian, C.Y.; Du, Y.; Yan, L.M.; Xia, J.Y. Experimental warming reduces ecosystem resistance and resilience to severe flooding in a wetland. Sci. Adv. 2022, 8, eabl9526. [Google Scholar] [CrossRef]
- Qiu, J. China: The third pole. Nature 2008, 454, 393–396. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Singh, B.K.; Maestre, F.T. Soil microbial communities drive the resistance of ecosystem multifunctionality to global change in drylands across the globe. Ecol. Lett. 2017, 20, 1295–1305. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- Romero-Olivares, A.; Allison, S.; Treseder, K. Soil microbes and their response to experimental warming over time: A meta analysis of field studies. Soil. Biol. Biochem. 2017, 107, 32–40. [Google Scholar] [CrossRef]
- Che, R.; Wang, S.; Wang, Y.; Xu, Z.; Cui, X. Total and active soil fungal community profiles were significantly altered by six years of warming but not by grazing. Soil. Biol. Biochem. 2019, 139, 107611. [Google Scholar] [CrossRef]
- DeAngelis, K.M.; Pold, G.; Topçuoğlu, B.D.; van Diepen, L.T.; Varney, R.M.; Blanchard, J.L.; Melillo, J.; Frey, S.D. Long-term forest soil warming alters microbial communities in temperate forest soils. Front. Microbiol. 2015, 6, 104. [Google Scholar] [CrossRef]
- Guo, X.; Feng, J.; Shi, Z.; Zhou, X.; Yuan, M.; Tao, X.; Lauren, H.; Yuan, T.; Wang, J.; Qin, Y. Climate warming leads to divergent succession of grassland microbial communities. Nat. Clim. Change 2018, 8, 813–818. [Google Scholar] [CrossRef]
- Kelly, D.P.; Wood, A.P. Reclassification of some species of Thiobacillus to the newly designated genera Acidithiobacillus gen. nov., Halothiobacillus gen. nov. and Thermithiobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Berg, I.A. Ecological aspects of the distribution of different autotrophic CO2 fixation pathways. Appl. Environ. Microbiol. 2011, 77, 1925–1936. [Google Scholar] [CrossRef] [PubMed]
- Tolli, J.; King, G.M. Diversity and structure of bacterial chemolithotrophic communities in pine forest and agroecosystem soils. Appl. Environ. Microbiol. 2005, 71, 8411–8418. [Google Scholar] [CrossRef] [PubMed]
- Selesi, D.; Pattis, I.; Schmid, M.; Kandeler, E.; Hartmann, A. Quantification of bacterial RubisCO genes in soils by cbbL targeted real-time PCR. J. Microbiol. Methods 2007, 69, 497–503. [Google Scholar] [CrossRef]
- Xiao, K.Q.; Bao, P.; Bao, Q.L.; Jia, Y.; Huang, F.Y.; Su, J.Q.; Zhu, Y.G. Quantitative analyses of ribulose-1, 5-bisphosphate carboxylase/oxygenase (RubisCO) large-subunit genes (cbbL) in typical paddy soils. FEMS Microbiol. Ecol. 2014, 87, 89–101. [Google Scholar] [CrossRef]
- Zhang, N.; Chen, K.; Wang, S.; Qi, D.; Zhou, Z.; Xie, C.; Liu, X. Dynamic Response of the cbbL Carbon Sequestration Microbial Community to Wetland Type in Qinghai Lake. Biology 2023, 12, 1503. [Google Scholar] [CrossRef]
- Liang, S.; Deng, J.; Jiang, Y.; Wu, S.; Zhou, Y.; Zhu, W. Functional distribution of bacterial community under different land use patterns based on faprotax function prediction. Pol. J. Env. Stud. 2020, 29, 1245–1261. [Google Scholar] [CrossRef]
- Wu, L.W.; Zhang, Y.; Guo, X.; Ning, D.L.; Zhou, X.S.; Feng, J.J.; Yuan, M.M.; Liu, S.; Guo, J.J.; Gao, Z.P.; et al. Reduction of microbial diversity in grassland soil is driven by long-term climate warming. Nat. Microbiol. 2022, 7, 1054. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.; Chen, F.; Zhang, Z.; Fang, J.; Xing, L.; Zeng, J.; Zhang, Q.; Liu, H.; Liu, W.; et al. Deterministic assembly of grassland soil microbial communities driven by climate warming amplifies soil carbon loss. Sci. Total Environ. 2024, 923, 171418. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, L.; Wang, J.; Zhang, Y.; Xiao, C. Effects of warming on the bacterial community and its function in a temperate steppe. Sci. Total Environ. 2021, 792, 148409. [Google Scholar] [CrossRef]
- Feng, J.J.; Wang, C.; Lei, J.S.; Yang, Y.F.; Yan, Q.Y.; Zhou, X.S.; Tao, X.Y.; Ning, D.L.; Yuan, M.T.M.; Qin, Y.J.; et al. Warming-induced permafrost thaw exacerbates tundra soil carbon decomposition mediated by microbial community. Microbiome 2020, 8, 3. [Google Scholar] [CrossRef]
- Qin, J.; Li, M.; Zhang, H.; Liu, H.; Zhao, J.; Yang, D. Nitrogen Deposition Reduces the Diversity and Abundance of cbbL Gene-Containing CO2-Fixing Microorganisms in the Soil of the Stipa baicalensis Steppe. Front. Microbiol. 2021, 12, 570908. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wang, W.; Liu, T.; Yan, W.; Wu, X.; Lei, J.; Xu, Y.; Chen, Y.; Yao, Y.; Jiang, W.; et al. Midseason drying increases soil dissolved organic carbon and rice yield via soil cbbL bacteria. J. Environ. Manag. 2024, 371, 123131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Ding, J.; Li, T.; Zhang, X. Plant communities are more sensitive than soil microbial communities to multiple environmental changes in the Eurasian steppe. Glob. Ecol. Conserv. 2020, 21, e00779. [Google Scholar] [CrossRef]
- Salcher, M.M.; Pernthaler, J.; Zeder, M.; Psenner, R.; Posch, T. Spatio-temporal niche separation of planktonic Betaproteobacteria in an oligo mesotrophic lake. Environ. Microbiol. 2010, 10, 2074–2086. [Google Scholar] [CrossRef] [PubMed]
- Simek, K.; Kasalicky, V.; Zapomelova, E.; Hornak, K. Alga-derived substrates select for distinct Betaproteobacterial lineages and contribute to niche separation in Limnohabitans strains. Appl. Environ. Microbiol. 2011, 77, 7307–7315. [Google Scholar] [CrossRef]
- Li, C.; Li, X.; Shi, Y.; Yang, Y.; Li, H. Effects of Nitrogen Addition on Soil Carbon-Fixing Microbial Diversity on Different Slopes in a Degraded Alpine Meadow. Front. Plant Sci. 2022, 13, 921278. [Google Scholar] [CrossRef]
- Wang, X.; Li, W.; Cheng, A.; Shen, T.; Xiao, Y.; Zhu, M.; Pan, X.; Yu, L. Community characteristics of autotrophic CO2-fixing bacteria in karst wetland groundwaters with different nitrogen levels. Front. Microbiol. 2022, 13, 949208. [Google Scholar] [CrossRef]
- Xu, J.; Ming, H.; Ren, K.; Li, D.; Huang, H.; Li, J.; Shao, K.; Li, H.; Fan, J. Spatial heterogeneity plays a vital role in shaping the structure and function of estuarine carbon-fixing bacterial communities. Mar. Environ. Res. 2024, 198, 106544. [Google Scholar] [CrossRef]
- Walker, V.K.; Palmer, G.R.; Voordouw, G. Freeze-thaw tolerance and clues to the winter survival of a soil community. Appl. Environ. Microbiol. 2006, 72, 1784–1792. [Google Scholar] [CrossRef]
- Wilson, S.L.; Walker, V.K. Selection of low-temperature resistance in bacteria and potential applications. Environ. Technol. 2010, 31, 943–956. [Google Scholar] [CrossRef]
- Wang, K.; Pan, R.; Fei, H. Changes in soil prokaryotic communities and nitrogen cycling functions along a groundwater table drawdown gradient in desert wetlands. Sci. Total Environ. 2022, 842, 156868. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. Available online: http://www.jstor.org/stable/27651243 (accessed on 8 January 2025). [CrossRef]
- Singh, B.K.; Bardgett, R.D.; Smith, P.; Reay, D.S. Microorganisms and climate change: Terrestrial feedbacks and mitigation options. Nat. Rev. Microbiol. 2010, 8, 779–790. [Google Scholar] [CrossRef]
- Yu, H.; Deng, Y.; He, Z.; Van Nostrand, J.D.; Wang, S.; Jin, D.; Wang, A.; Wu, L.; Wang, D.; Tai, X.; et al. Elevated CO2 and Warming Altered Grassland Microbial Communities in Soil Top-Layers. Front. Microbiol. 2018, 9, 1790. [Google Scholar] [CrossRef]
- Liao, H.; Hao, X.; Qin, F.; Delgado-Baquerizo, M.; Liu, Y.; Zhou, J. Microbial autotrophy explains large-scale soil CO2 fixation. Glob. Chang. Biol. 2022, 29, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Ning, D.; Lu, Z.; Zhang, N.; Hale, L.; Wu, L. Century long fertilization reduces stochasticity controlling grassland microbial community succession. Soil. Biol. Biochem. 2020, 151, 108023. [Google Scholar] [CrossRef]
- Ning, D.L.; Yuan, M.T.; Wu, L.W.; Zhang, Y.; Guo, X.; Zhou, X.S.; Yang, Y.F.; Arkin, A.P.; Firestone, M.K.; Zhou, J.Z. A quantitative framework reveals ecological drivers of grassland microbial community assembly in response to warming. Nat. Commun. 2020, 11, 4717. [Google Scholar] [CrossRef]
- Zhang, M.X.; Zhao, L.Y.; Hu, J.P.; Khan, A.; Yang, X.X.; Dong, Q.M. Different grazers and grazing practices alter the growth, soil properties, and rhizosphere soil bacterial communities of Medicago ruthenica in the Qinghai-Tibetan plateau grassland. Agric. Ecosyst. Environ. 2023, 352, 108522. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D. Climate warming enhances microbial network complexity and stability. Nat. Clim. Change 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Chen, W.J.; Zhou, H.K.; Wu, Y.; Li, Y.Z.; Qiao, L.L.; Wang, J.; Zhai, J.Y.; Song, Y.H.; Zhao, Z.W.; Zhang, Z.H.; et al. Plant-mediated effects of long-term warming on soil microorganisms on the Qinghai-Tibet Plateau. Catena 2021, 204, 105391. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.; et al. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [PubMed]
- Tu, Q.; Yuan, M.; He, Z.; Deng, Y.; Xue, K.; Wu, L.; Hobbie, S.E.; Reich, P.B.; Zhou, J. Fungal communities respond to long-term CO2 elevation by community reassembly. Appl. Environ. Microbiol. 2015, 81, 2445–2454. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef]
- Herren, C.M.; McMahon, K.D. Cohesion: A method for quantifying the connectivity of microbial communities. ISME J. 2017, 11, 2426–2438. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nature reviews. Microbiology 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef]
- Hernandez, D.J.; David, A.S.; Menges, E.S.; Searcy, C.A.; Afkhami, M.E. Environmental stress destabilizes microbial networks. ISME J. 2021, 15, 1722–1734. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.J.; Bufford, J.L.; Barnes, A.D.; Barratt, B.I.P.; Deslippe, J.R.; Dickie, I.A.; Goldson, S.L.; Howlett, B.G.; Hulme, P.E.; Lavorel, S.; et al. A network perspective for sustainable agroecosystems. Trends Plant Sci. 2022, 27, 769–780. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Average Degree | Modularization | Edge | Negative Correlation | Positive Correlation | Point |
|---|---|---|---|---|---|---|
| Nck | 2.217 | 0.73 | 133 | 53 | 80 | 60 |
| NW | 3.985 | 0.583 | 263 | 105 | 158 | 66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Qi, D.; Zhou, S.; Wang, S.; Fu, L.; Sun, J.; Chen, K. Warming Induces High Environmental Pressure in the Lakeshore Wetland, Promoting Interactions Among Carbon-Fixing Microorganisms. Agriculture 2025, 15, 580. https://doi.org/10.3390/agriculture15060580
Zhang N, Qi D, Zhou S, Wang S, Fu L, Sun J, Chen K. Warming Induces High Environmental Pressure in the Lakeshore Wetland, Promoting Interactions Among Carbon-Fixing Microorganisms. Agriculture. 2025; 15(6):580. https://doi.org/10.3390/agriculture15060580
Chicago/Turabian StyleZhang, Ni, Desheng Qi, Shijia Zhou, Siyu Wang, Li Fu, Jianqing Sun, and Kelong Chen. 2025. "Warming Induces High Environmental Pressure in the Lakeshore Wetland, Promoting Interactions Among Carbon-Fixing Microorganisms" Agriculture 15, no. 6: 580. https://doi.org/10.3390/agriculture15060580
APA StyleZhang, N., Qi, D., Zhou, S., Wang, S., Fu, L., Sun, J., & Chen, K. (2025). Warming Induces High Environmental Pressure in the Lakeshore Wetland, Promoting Interactions Among Carbon-Fixing Microorganisms. Agriculture, 15(6), 580. https://doi.org/10.3390/agriculture15060580