Abstract
Rosmarinus officinalis (rosemary) is one of the most cultivated Lamiaceae species because of its medicinal, aromatic and ornamental properties. In the late summer of 2023, symptoms of desiccation were observed in a rosemary field located in southern Italy and the isolation from necrotic roots revealed three different fungal species: Ceratobasidium sp., Fusarium falciforme and Macrophomina phaseolina. All the isolates were morphologically and molecularly identified and Koch’s postulates assays were performed to establish the pathogenicity and the effect of each fungus on the disease incidence and severity. A dual plate method was used to investigate the interaction between Ceratobasidium sp., F. falciforme and M. phaseolina, mixing them in pairs and all together. Similarly, in vivo inoculations with the pathogens in combination were used to verify the role of each isolate in the interaction with the others and the contribution in plant disease expression. The results allowed to identify M. phaseolina and F. falciforme as causal agents of desiccation and root rot of rosemary, with M. phaseolina being the most aggressive between the two. Conversely, Ceratobasidium sp. behavior was influenced by the interaction with the other pathogens and the plant. Notably, this is the first report of M. phaseolina and F. falciforme on rosemary plants worldwide. This disease, caused by multiple pathogens, highlighted the crucial role of the combination of traditional in vivo assays and the use of molecular methods to achieve an accurate diagnosis.
1. Introduction
Rosemary (Rosmarinus officinalis L.) is an evergreen perennial shrub belonging to the Lamiaceae family, characterized with a well-known aromatic odor [1]. Native to the Mediterranean area, rosemary is now grown in Europe and globally due to its wide range of properties, including medicinal, aromatic and ornamental uses [2]. Moreover, the ability of this species to adapt to drought stress and its resistance to biotic adversities make it easy to cultivate. On the other hand, rosemary is sold in several markets; indeed, the leaves are used as common spices and oils as flavors, antioxidants and anti-inflammatory compounds [3]. The versatility of rosemary plants and of its derivates, its strong adaptability to adverse climate conditions and the need to address economic crises led Italian growers to consider rosemary as an alternative agricultural product, attesting a gross output of 27,000 EUR/ha [4]. Official data about the cultivation of rosemary, as for other medicinal species, are unavailable [4], but it is estimated that this plant is cultivated in southern Italy over a surface of about 60 ha, producing 15,000 tons of fresh herb. In the Salerno province (southern Italy), the cultivation of rosemary is recently gaining importance as alternative crop to traditional fruit bearing vegetables and leafy salads. Concurrently, the increasing cultivation of rosemary in open fields and in protected cultivation settings has led to the observation of more frequent biotic disorders that seriously compromise the success of its cultivation. Rosemary has been reported as a host of several agents of powdery mildew [5,6,7], while the literature about root diseases is still quite scarce. In 2007, a mortality of 70% of rosemary plants was reported in Valencia due to crown and root rot caused by Phytophthora nicotianae [8], while the same disorder was observed in Iran in 2012, caused by Fusarium sambucinum [9]. Sclerotinia sclerotium was found to be responsible of stem rot in United States in 2004 [10] and of white mold in Italy in 2017 [11], while Rhizoctonia solani AG1-IA and AG1-IB were reported in Italy, in Korea and in the United Kingdom [12,13] as being responsible for web blight; in addition, binucleate Rhizoctonia fragariae isolates were described in Spain in 2017 [14]. In 2019, Mekonnen et al. [15] reported Fusarium oxysporum as the causal agent of fusarium wilting, while in 2021, stem canker caused by Botrytis cinerea was described in the “Pacific Northwest pest management handbooks, plant disease” [16]. The increase in disease reports on rosemary in the last 15 years could likely be due to a cumulative effect of cultivation spread and the action of pathogens that used to be restricted to warm climates and that, due to rising temperatures, are now spreading all over the globe [17]. In this context, this work reports (i) a desiccation that appeared in rosemary cultivations in southern Italy; (ii) the diagnostic pathway that was followed in order to go deep into the understanding of this new disease, applying either molecular and traditional methods; (iii) the description of a new form of plant wilting caused by multiple fungal strains and the attribution to them to roles in plant tissue infection.
2. Materials and Methods
2.1. Field Inspection and Sampling
In September 2023, rosemary (Rosmarinus officinalis L.) plants of an unknown variety, cultivated in an open field in Eboli (Salerno, southern Italy), showed heavy wilting of the aerial parts, were frequently asymmetric on plant branches and had extended necrosis on the collar. Roots were rotten, taking on a black color, and the cortical part was easily separable from the woody tissues. In the cultivation, there was a 40% incidence of diseased plants, distributed in spots mainly located in concave areas; in December, the incidence reached almost 100%. Eight plant samples with the asymmetric desiccation (Figure 1) were collected and analyzed in the “Laboratory of Plant Diagnostic and Disease Resistance Testing” of the Council for Agricultural Research and Economics—Research centre for Vegetable and Ornamental Crops (CREA) located in Pontecagnano (Salerno, Italy).

Figure 1.
Asymmetrical desiccation on plants from rosemary field in southern Italy.
2.2. Pathogens Isolation and Morphological Characterization
Isolation was performed from the basal part of the plants on Potato Dextrose Agar (PDA) (Oxoid, Thermofisher, Foster City, CA, USA) amended with 50 ppm streptomycin, 50 ppm chloramphenicol and 50 ppm neomycin (antibiotics from Merk, Darmstadt, Germany). Collar and roots were washed in running tap water for 30 min, then sterilized in 1% sodium hypochlorite solution for 20 min and finally rinsed three times with sterile distilled water. Obtained disinfected tissues were put under laminar flow cabinet until drying and then pieces of 2–3 mm were cut from roots and internal tissues of the collar, placed on PDA and incubated at 26 °C. After three days, to avoid overlapping, pieces from multiple colonies were transferred to fresh PDA plates. After one week, three types of colonies were distinguishable and were grouped as Rhizoctonia-type (CS), Macrophomina-type (MP) and Fusarium-type (FS) colonies. Single tip or monosporic cultures were obtained from CS and MP colonies, while monosporic cultures were obtained from FS colonies to be submitted to further tests. All the isolates were coded and stored in 30% glycerol at −80 °C in the MicroHort collection of the CREA laboratories. Three colonies were chosen as representative of the three pathogen genera (Rhizoctonia sp., Macrophomina sp. and Fusarium sp.). They were described observing the main macroscopic characteristics as well as mycelium morphological features by using a Nikon eclipse 90i microscope (Nikon, Nikon Europe B.V, Amstelveen, The Netherlands) with 100×, 200× or 400× magnification in transmitted light configuration; the measurements of fungal structures (conidia and hyphae width) were expressed as the mean of 20 replicates. The Rhizoctonia-like colonies were analyzed by staining the mycelium with safranin (Sigma Aldrich, Merck, Darmstadt, Germany) to highlight the nuclei inside the hyphae [18]. At this point, fungal mycelia were collected and placed on a glass coverslip, incubated in an aqueous solution of 0.1% safranin for 20 min and then washed with sterile phosphate-buffer saline pH 7.4. The specimens were then covered with coverslip and analyzed using a microscope in transmitted light configuration.
2.3. Molecular Identification and Phylogenetic Analysis
Morphological characterization was confirmed by molecular identification carried out by sequencing conservative regions such as the internal transcribed spacer (ITS) and translation elongation factor (TEF) for all the pathogens; for Fusarium species identification, two additional regions were used, namely β-tubulin (TUB) and actin (ACT). Briefly, monosporic and monohyphal colonies were grown in Potato Dextrose Broth (Oxoid, Thermofisher, Foster City, CA, USA) for 48–72 h, filtered with a sterile gauze and grinded in liquid nitrogen. Then, the mycelia were submitted to DNA extraction by using the Genomic DNA Isolation Kit (Norgen, Biotek Corp., Thorold, ON, Canada) and specific gene regions were amplified by using Phusion™ High-Fidelity DNA Polymerase (2 U/µL) (Thermofisher, Foster City, CA, USA) according to manufacturer instructions in a Applied Biosystems™ SimpliAmp™ Thermal Cycler (Applied Biosystem, Fisher Scientific, Rodana, Italy). The primers used in the PCR reactions and the annealing temperatures were reported in Table 1. The GeneJET PCR Purification Kit (Thermofisher, Foster City, CA, USA) was used according to manufacturer’s instructions to purify the amplicons that were subsequently sent to BMR Genomics (Padua, Italy) for Sanger sequencing. Chromatograms were manually inspected using Chromas (http://technelysium.com.au/wp/chromas/), accessed on 1 December 2023. Sequences were identified using a nucleotide Basic Local Alignment Search Tool (BLAST, https://blast.ncbi.nlm.nih.gov) accessed on 29 December 2023 and then deposited in GenBank.

Table 1.
Summary of gene regions and primers used to identify rosemary isolates.
Table 1.
Summary of gene regions and primers used to identify rosemary isolates.
| Pathogen-Type Colony | Region | Primers | Annealing T (°C) | References |
|---|---|---|---|---|
| Rhizoctonia sp. | Internal transcribing spacer | ITS1-4 | 55 | [19] |
| Macrophomina sp. | Internal transcribing spacer | ITS1-4 | 55 | [19] |
| Translation elongation factor 1α | TEF 728-986 | 58 | [20] | |
| Fusarium sp. | Internal transcribing spacer | ITS1-4 | 55 | [19] |
| Translation elongation factor 1α | TEF 688-1251 | 58 | [21] | |
| β-tubulin | T1/β-tub2b | 58 | [22,23] | |
| Actin | ACT512-783 | 59 | [20] |
Phylogenetic analyses were conducted on the representative CS, MP and FS isolates, starting by editing the sequences using MEGA X software (version 10.2.6). After this, annotated ITS sequences of all the amplicons from studied strains, TEF sequences of M. phaseolina and Fusarium solani species complex (FSSC) and β-tubulin and actin of FSSC derived from GenBank were chosen; selecting accessions of reference strains and alignment was performed using the default settings of Muscles and trimmed. The obtained alignment was employed to construct phylogenetic trees using a max likelihood method and Tamura–Nei model with 1000 bootstrap replicates using MEGA X. Agroathelia rolfsii, Botryosphaeria dothidea and Fusarium oxysporum were used as outgroups in Rhizoctonia-type, Macrophomina-type and Fusarium-type trees, respectively.
2.4. In Vitro Interaction Analysis
The interaction among representative CS, MP and FS isolates was evaluated by using dual culture method. Briefly, an agar disk (4 mm) from a specific strain was placed 2 cm from the Petri dish edge and a similar-sized agar disk from a different tested fungus was placed at the opposite side. As a control, each single strain was placed in a similar manner on a fresh PDA plate. All the combinations of two isolates and the three isolates all together were incubated at 28 ± 1 °C for 7 days. Tests were carried out in triplicate. The antagonistic activity was recorded by measuring the radius (Rp1) of the first pathogen (P1) in the direction of the second (P2) and comparing the results with the colony in the control plate (Rcn). In the case of interaction among the three pathogens, the radius of one of them (e.g., radius Rp1vs P2 and Rp1 vs. P3 of pathogen 1) was measured in the direction of other two (e.g., P2 and P3) (Figure 2). The inhibition of radial growth was calculated according to Sonawane et al. [24] by using the following formula:
where PIRG = Percent Inhibition of Radial Growth; Rc = growth of the radius in the control plate; Rp = growth of the radius in the dual plate.
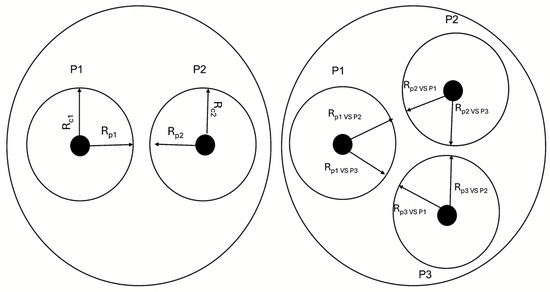
Figure 2.
Schematic representation of dual culture experiments. In detail, Rcn stands for radius of the strain in the control plate; Rpn for radius of the strain in the dual plate; P1, P2 and P3 represent Rhizoctonia-type, Macrophomina-type and Fusarium-type colonies.
2.5. Pathogenicity Tests and Pathogen In Vivo Interaction
In order to establish the causal agents of rosemary wilting and desiccation and to fulfill the Koch’s postulates, pathogenicity tests were performed. Briefly, healthy plants, with roots of 5 cm obtained by rooted cuttings, were transplanted into 10 cm pots filled with sterile peat. To serve as an inoculum source, rice grains were sterilized by autoclaving them two times at 121 °C for 20 min, subsequently contaminated with three mycelium plugs from one-week-old colonies of each strain and then incubated at 26 ± 1 °C for two weeks, until fungi covered all the grains. One week after transplantation, rosemary plants were inoculated with two rice grains for each pathogen (Table 2). The rice grains were located 0.5 cm from the plant stem and were 2.0 cm in depth. Nine plants, arranged in two repetitions, were inoculated with each isolate individually, with each pairwise combination (FS + CS; MP + FS; MP + CS) and with the combination of all three isolates (CS + MP + FS); nine plants were treated with sterile grains serving as negative control.
Table 2.
Summary of inoculations for pathogenicity test.
The symptoms of the disease were recorded two months after the inoculation by uprooting the plants and measuring the symptom level separately for each plant, according to the following disease scale: 0: healthy roots; 1: necrotic lesions on the stem and browned roots for less 50%; 2: extensive necrotic stem rot and roots browned for more than 50%; 3: rotted or absent roots and plant wilting. Symptom values were used to calculate disease incidence (I), disease severity (DS) and the disease index (DI). I was expressed as the percentage of diseased plants on the total plants in a sample. DS was calculated as the mean of the symptom levels of each sample. DI was expressed in percentage and calculated with the McKinney index as
where A, B, C and D are the number of plants that showed the symptom levels 0, 1, 2 and 3, respectively [25]. Plants showing disease level 3 were further focused by calculating the percentage on plants at this level on the total of the plants in the sample. Disease expression on inoculated plants was compared with that of the negative control.
Finally, in order to confirm the pathogenicity of isolates inoculated individually and to evaluate the effect of co-infection on plant tissue colonization, reisolation from inoculated plants was performed. To this purpose, plants were collected and externally sterilized, and small pieces from symptomatic tissues were cut and placed on PDA. For the asymptomatic plants, pieces from roots and collar were randomly cut and placed on fresh PDA.
2.6. Statistical Analysis
All data were tested for normality and homogeneity of variance. If assumptions were met, differences among groups were assessed using a one-way ANOVA followed by Tukey’s HSD post hoc test (p < 0.05). When assumptions were not met, a non-parametric Kruskal–Wallis test was performed, followed by Dunn’s post hoc test (p < 0.05). The χ2 test followed by Dunn’s post hoc test was used to analyze the wilting percentage. Statistical testing was achieved using Past software (version 5.0.2; accessed by https://www.LO4D.com—Free Software Downloads, 18 June 2024).
3. Results
3.1. Isolation and Morphological Identification
Diagnostic analyses carried out on the collar and roots of rosemary plants (Figure 1) enabled the isolation of three types of colonies morphologically similar to Rhizoctonia sp., Macrophomina sp. and Fusarium sp. Rhizoctonia-type (CS) colonies were recognized for their compact creamy-white mycelium that became light brown-orange after 15 days; Macrophomina-type (MP) colonies were fast growing (the surface of 9 cm PDA plates were covered in 7 days), typically black and sometimes dark gray, with sparse mycelium in which abundant black microsclerotia were visible; Fusarium-type (FS) colonies were creamy-white, with fluffy aerial mycelium that developed a yellow-green color after 20 days. MP colonies were isolated from all the tested plants, FS colonies from 50% of them, while CS colonies were isolated only from two plants. Among all the isolates obtained, MP colonies were the most represented (65% of the isolates). The isolates CS MH 1410.10, MP MH 1410.6 and FS MH 1410.3 were chosen as representative of Rhizoctonia-type, Macrophomina-type and Fusarium-type isolates, respectively (Figure 3). All the strains were stored in the McroHort collection of CREA with the prefix MH CREA OF before the numerical code.
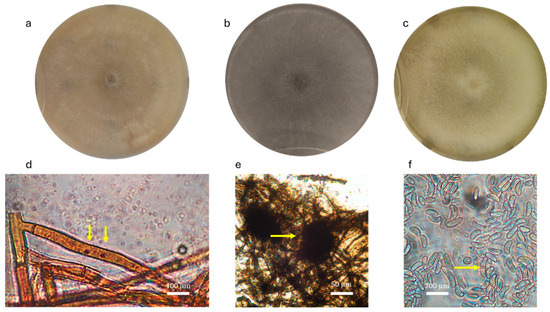
Figure 3.
Morphological characteristics of isolates from rosemary. In the upper row, colony morphology of (a) CS isolate MH CREA OF 1410.10, (b) MP isolate MH CREA OF 1410.6 and (c) FS isolate MH CREA OF 1410.3. In the lower row, are the microscopic structures of the following: (d) MH CREA OF 1410.10 hyphae with the two nuclei, (e) MH CREA OF 1410.6 microsclerotia and (f) conidia of MH CREA OF 1410.3. The arrows indicate the two nuclei in Ceratobasidium sp. hyphae, the microsclerotia of M. phaseolina, and the septum in F. falciforme spores.
After two weeks of development on PDA plates and observation at the optical microscope, CS MH 1410.10 revealed the colony identification as binucleate Rhizoctonia sp. (Ceratobasidium sp.) for its microscopic structures, the latter of which being the following: binucleate hyphae and brown color in mature hyphae with the typical insertion angle of 90° of the main hyphae with the secondary branches. Microscopic features observed in MP MH 1410.6 revealed dark and septate hyphae with branches placed at 90° over the main hypha, with a septum after a constriction in the secondary hypha. The observation highlighted that the isolate had ovoid microsclerotia, with an average main diameter of 97.0 µm (ranging from 93.0 to 99.2 µm). Finally, FS MH 1410.3 produced abundant septate falciform macroconidia that were on average 79.0 × 11.7 μm in size and bicellular microconidia that were on average 18.2 × 7.0 μm in size.
3.2. Molecular Identification and Phylogenetic Analysis
The molecular characterization of CS, MP and FS isolates was carried out by ITS for all the strains, TEF 1α for MP and FS, and TUB and ACT sequences for FS. Sequenced gene regions were aligned in GenBank, allowing to identify the isolates MH 1410.10 as Ceratobasidium sp., MH 1410.4.1, MH 1410.4.2, MH 1410.6, MH 1410.4.14 and MH 1410.15 as M. phaseolina and FS 1410.3 as FSSC (F. solani/F. falciforme). The accession numbers uploaded in GenBank and the results of the alignment are detailed in Table 3.
Table 3.
Summary of the molecular analyses performed on isolates and identification based on multiple alignments performed in BLASTn.
Phylogenetic analyses were performed by using reference accessions for all the rosemary isolates (Figure 4, Figure 5 and Figure 6).

Figure 4.
Maximum likelihood phylogenetic tree of binucleate Ceratobasidium sp. MH 1410.10. ITS sequence of Athelia rolfsii CBS191.62 was used as the outgroup. The bootstrap tree inferred from 1000 replicates is shown. The phylogenetic tree was inferred by using the maximum likelihood method and Tamura–Nei model. The percentage of replicate trees in which the associated taxa are clustered together in the bootstrap test is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). Phylogenetic analyses were conducted in MEGA X.

Figure 5.
Maximum likelihood tree of Macrophomina phaseolina strains from rosemary. ITS and TEF sequences of Botryosphaeria dothidea CREA OF 360.4 were used as the outgroup. The bootstrap tree inferred from 1000 replicates is shown. The phylogenetic tree was inferred by using the maximum likelihood method and Tamura–Nei model. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). Phylogenetic analyses were conducted in MEGA X. The isolates studied in this work were highlighted by arrows and brackets. Regarding FS strain, the combined ITS and TEF phylogenetic analyses showed that MH 1410.3 falls in a clump containing strains of Fusarium falciforme (LC13830 and LC13831), with a 55% percentage of bootstraps (Figure 6).
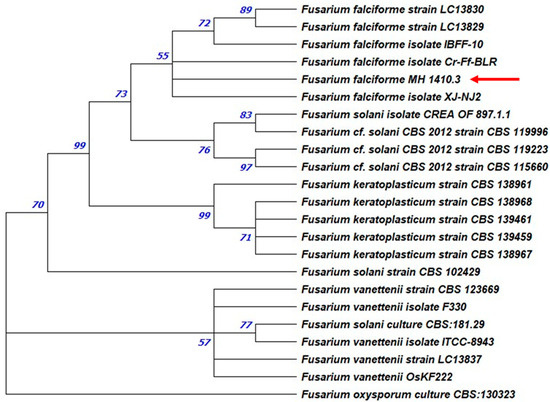
Figure 6.
Maximum likelihood phylogenetic tree. ITS and TEF sequences of Fusarium oxysporum CBS 130323 were used as the outgroup. The bootstrap tree inferred from 1000 replicates is shown. The phylogenetic tree was inferred by using the maximum likelihood method and Tamura–Nei model. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). Phylogenetic analyses were conducted in MEGA X. The isolates studied in this work were highlighted by arrows.
The strain CS MH 1410.10 was analyzed with 49 accessions retrieved from GenBank, including R. solani and Ceratobasidium spp. It grouped with 92% bootstraps with an already reported binucleate Rhizoctonia, isolated in California from grapevine (strain DCHG2B, accession OR052655) and in cluster with Ceratobasidium spp., confirming the molecular identification (Figure 4).
The phylogenetic analysis on MP was performed by combining the ITS and TEF sequences; the concatenated phylogenetic tree confirmed the identification obtained with the alignment performed in BLASTn. Indeed, according to BLASTn comparison, the ITS regions of MH 1410.4.1 and MH1410.4.2 both aligned with a similarity of 93.40%, with the accession OP815340 (strain KE26), while MH 1410.6, MH 1410.14 and MH 1410.15 showed an identity of 97.82%, with the accession MK757624 (isolate BMP16). The alignments in the TEF regions showed that both MH 1410.4.1 and 1410.4.2 aligned with a similarity of 99.22% with the strain KARE243 (accession MN318093) while the strains MH 1410.4.6, MH 1410.4.14 and 1410.4.15 showed an identity of 100.00%, 99.61% and 99.61% with the accessions MN318093 (strain KARE243), PP460507 (isolate Mf1) and KX400853 (strain TP1b), respectively. Moreover, M. phaseolina isolates was shown to cluster with a well-supported bootstrap, with other strains retrieved from the database (Figure 5).
The maximum likelihood analyses based on TUB sequences showed a strongly supported relationship (93%) with the accession KY923278 of F. solani strain CSAEGro-DAAF2 isolated from tomato in Mexico and two accessions (OK087483 and MW534050) of the strains C12 and LC13828 of F. falciforme from India (Figure 7). This grouping partially confirms the identification by BLASTn analysis that allows the sequence alignment with the MN318093 accession of F. solani strain KARE243 retrieved from GenBank.
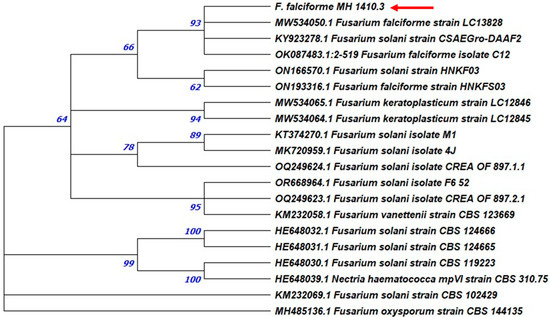
Figure 7.
Maximum likelihood phylogenetic tree of tubulin gene of F. solani. Tubulin sequence of Fusarium oxysporum CBS 144235 strain was used as the outgroup. The bootstrap tree inferred from 1000 replicates is shown. The phylogenetic tree was inferred by using the maximum likelihood method and Tamura–Nei model. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). Phylogenetic analyses were conducted in MEGA X. The isolates studied in this work were highlighted by arrows.
Similarly, the Actin gene sequencing confirmed the relationship between the strain FS MH 1410.3 with F. solani, showing a similarity of 99.27% with the accession MK968888 of strain AVRP-19, after the BLASTn multiple alignment analysis. On the other side, the phylogenetic analyses, performed with the limited number of strains available in the GenBank, allowed the strain MH 1410.3 to allocate in the cluster of F. falciforme. The limited number of accessions used for the comparison did not allow the construction of phylogenetic tree.
3.3. In Vitro Pathogen Interaction
Dual cultures plates were realized to evaluate the in vitro interaction of the three selected isolates. Starting with the combination of FS MH 1410.3 and MP MH 1410.6 (Figure 8a), the test showed that PIRG, in respect to the control, was 57.57% ± 0.10% for the first, while it resulted in 37.53% ± 0.05% for the latter. CS MH 1410.10 growth seemed to be not influenced, in comparison to the control, when tested with FS; indeed, it was not inhibited at all (PIRG = 0.00%), while FS 1410.3 was inhibited of 37.50% ± 0.10%. Moreover, CS and FS colonies were found to overlap by 15.0 mm on average (Figure 8b). On the contrary, in the combination of CS with MP (Figure 8c), the PIRG for CS was attested at 50.10% ± 0.06%, while for MP it was 27.53% ± 0.05%; CS and MP overlapped for 11.0 mm on average. In the mix of the three pathogens (Figure 8d), in the case of MP versus both CS and FS, PIRG was the lowest (37.47% and 37.60%, respectively), while PIRG for FS was about 50% (49.93% and 50.17% versus MP and CS, respectively) and, for CS it was 67.47% versus MP and 50.07% versus FS (Table 4).
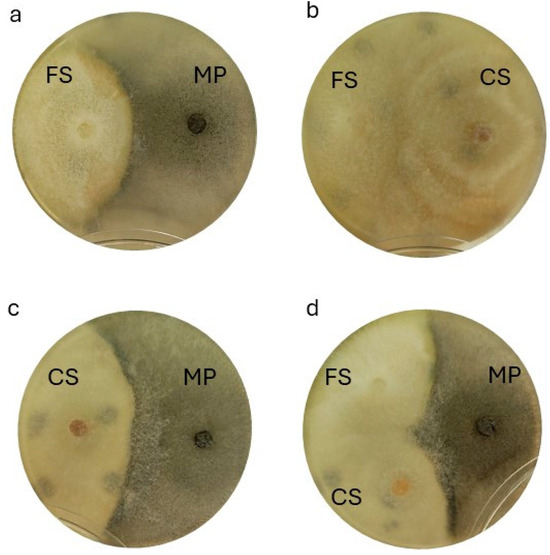
Figure 8.
In vitro dual test plate analyses. (a) F. falciforme MH CREA OF 1410.3 (left) against M. phaseolina MH CREA OF 1410.6 (right); (b) F. falciforme MH CREA OF 1410.3 (left) and Ceratobasidium sp. MH CREA OF 1410.10 (right); (c) Ceratobasidium sp. MH CREA OF 1410.10. (left) against M. phaseolina MH CREA OF 1410.6 (right) and (d) combination of all the three pathogens (MH CREA OF 1410.10 on the left, MH CREA OF 1410.3 in the middle and MH CREA OF 1410.6 on the right).
Table 4.
Percent Inhibition Radius Growth (PIRG) of Macrophomina phaseolina MH CREA OF 1410.3, Fusarium falciforme MH CREA OF 1410.6 and Ceratobasidium sp. MH CREA OF 1410.10 when tested in pairs and all together in dual culture plates.
3.4. Pathogenicity Test and In Vivo Pathogen Interaction
The strains FS MH 1410.3, MP MH 1410.6 and CS MH 1410.10 were analyzed in vivo to verify their role in root rot of rosemary plants. After one month from the inoculation, symptoms on the aerial part were visible in plants inoculated with MP as the wilting of the top leaves. Two months after the inoculation, when most of the plants inoculated with MP were wilted, all the plants were uprooted and the roots were inspected: necrosis was extended to the roots at variable levels, confirming the symptoms observed in the field (Figure 9).

Figure 9.
Symptoms of desiccation and root rot on inoculated plants of rosemary. The picture shows the following: CTRL = control; CS = Ceratobasidium sp. MH 1410.10; MP = M. phaseolina MH 1410.6; FS = F. falciforme MH 1410.3; CS + FS = combination Ceratobasidium sp. and F. falciforme; CS + MP = the combination of Ceratobasidium and M. phaseolina; FS + MP = the combination of F. falciforme and M. phaseolina; CS + MP + FS = the combination of the three isolates.
In detail, the symptoms observed after inoculation with the three selected isolates alone, in pairs and all together, evidenced root discolorations that were absent in the negative control. Indeed, roots showed a color ranging from brown to black: sparse brown spots were observed in plants inoculated with CS while roots inoculated with FS showed confluent and non-confluent brown necrosis, sometimes developing in extended black necrosis. Roots were completely necrotized, resembling the typical charcoal rot in plants inoculated with MP alone. Severe damages (extended necrosis) were also observed after inoculation with FS + CS or with the three isolates. In these cases, a desiccation of leaves was also observed. The symptoms were not specific to the infecting pathogen but differentiable after the comparison with the negative control.
The disease incidence (I) calculated for single strains and for all combinations highlighted that plants inoculated individually with MP and FS and with all combinations of two and three isolates showed an I of 100% with all plants infected and showing symptoms at different levels. Conversely, plants inoculated with CS alone showed an I of 66.7%, a value that was statistically different from the others (p < 0.05). Root rotting was observed in a single plant in the negative control. Reisolation from inoculated roots confirmed the tissue infection with the three isolates and consistently confirmed pathogenicity in the case of MP and FS. CS was not always reisolated from brown necrosis, while it was sometimes recovered from non-symptomatic plants. None of the three isolates was recovered from the symptomatic plant in the negative control, suggesting that the slight rotting could have been attributed to abiotic stresses.
Disease severity (DS) data highlighted the highest DS values for MP (DS = 2.4 ± 0.88), which was statistically different from DS registered in plants inoculated with MP + CS, MP + FS, FS + CS and CS + MP + FS (DS = 1.6 ± 0.86, 1.5 ± 0.88, 2.3 ± 0.70 and 1.7 ± 0.97, respectively) (Figure 10a). DS values in plants inoculated with FS (1.2 ± 0.44) were statistically different from FS + CS and higher than those in plants inoculated with CS alone (0.6 ± 0.5) and than those in the negative control (0.1 ± 0.5). Moreover, DS in plants inoculated with CS was statistically comparable to the negative control (Figure 10a). Figure 10b depicts the distribution of plants based on disease symptom levels in each sample: plants inoculated with MP alone showed most of the plants with a disease level of 3. Similarly, all the combinations including MP showed plants with a disease level of 3 as well as when FS colonies were inoculated together with CS colonies. No plants with a disease level of 3 were observed in the samples inoculated individually with CS or FS. These results are further reported in Figure 10c, where only plants at level 3 were depicted and statistical analysis highlighted differences among plants inoculated with MP (alone or in combination) with the other treatments; in particular, it is shown that FS, although inducing an I of 100%, did not cause heavy symptoms when inoculated alone.
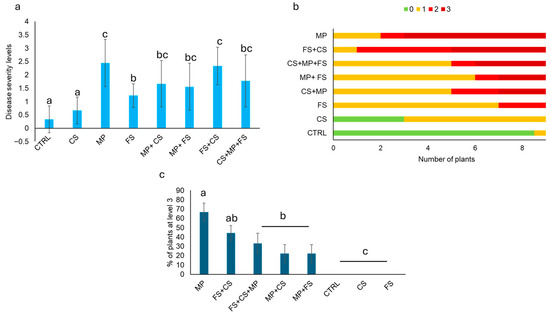
Figure 10.
(a) Disease severity; (b) average number of plants per level of disease; (c) percentage of plants at level 3. The abbreviations stand for the following: CTRL = control; CS = Ceratobasidium sp. MH 1410.10; MP = M. phaseolina MH 1410.6; FS = F. falciforme MH 1410.3; CS + FS = the combined inoculum of Ceratobasidium sp and F. falciforme; CS + MP = the combination of Ceratobasidium and M. phaseolina; FS + MP = the combination of F. falciforme and M. phaseolina; CS + MP + FS = the combination of the three pathogens. Different letters indicate statistically significant differences calculated according to the Kruskal–Wallis test associated with Dunn’s post hoc test (p ≤ 0.05). The wilting percentage was analyzed by using the χ2 test followed by a Dunn’s post hoc test (p ≤ 0.05). The number of plants for the class of symptoms (0, 1, 2 and 3) was expressed as mean between two independent replicates.
The disease index (DI) calculation highlighted that MP was the most virulent pathogen when tested alone (DI = 81.84%), with the ability to infect more plants with the highest disease symptoms level; plants inoculated with FS alone showed a DI of 40.74%, while CS determined a DI of 22.22%. MP in combination with either CS or FS showed a DI of 55.56% and 51.85%, respectively, being significantly lower than MP alone. Conversely, the combination of FS and CS showed a DI of 77.78%, which was higher than that of the two single pathogen infections. The three MP, FS and CS colonies showed a DI of 59.26%, which was comparable to that measured after the inoculation of the three pairs.
In order to evaluate the capability of plant tissue colonization and survival of the three isolates when inoculated in pairs and in three, reisolations were performed from all plants. Table 5 reports the percentage of plants from which the isolates were reisolated: CS was recovered from about 50% of samples when inoculated in pairs (42.8% and 50.0%, when inoculated with MP + CS and FS + CS, respectively), while the CS reisolation rate decreases to 12.5% from plants inoculated with the three isolates. FS was isolated from all plants inoculated with FS + CS and MP + FS, while it was recovered only from 62.5% of plants when inoculated with CS + MP + FS. MP was reisolated from 57.1% and 50.0% of plants when inoculated with CS and FS, respectively, while the percentage increases to 87.5% in CS + MP + FS (Table 5).
Table 5.
Percentage of reisolation of the three isolates from rosemary plants after the inoculation alone and in all the combinations.
4. Discussion
This study reports the outbreak, in 2023, of a severe desiccation of rosemary plants recorded in southern Italy which led to the isolation in the laboratory of three fungi: Ceratobasidium sp., M. phaseolina and F. falciforme. The recognition of the three species was performed complementing the observation of typical morphological features with gene region sequencing, confirming the effectiveness of DNA barcoding, largely employed in plant pathology, as a tool for gaining information about genus and species identification [19,26]. Indeed, the results of molecular analyses allowed the identification of MH 1410.10 as Ceratobasidium sp., the teleomorph of binucleate Rhizoctonia sp., supporting the microscopic observation that had revealed the presence of binucleate cells in the mycelium; moreover, phylogenetic analyses allocated the Rhizoctonia sp. strain in the AG-G anastomosis group, one of most common on cultivated crops [27]. Similarly, BLASTn alignment and concatenated ITS and TEF phylogenetic tree robustly confirmed the identification of MH 1410.6 as M. phaseolina, the most widespread species in the genus [28]. Due to the complexity of Fusarium solani species complex (FSSC) taxonomy, a Multi Locus Sequence Analysis was used to precisely identify the FS strain MH 1410.3; as a result, alignments in BLASTn enabled the recognition of the strain as F. solani for the ITS, TUB and ACT gene regions while TEF1-α identified the strain as F. falciforme. Furthermore, the grouping in the concatenate ITS-TEF phylogenetic tree supported the identification as F. falciforme while the phylogenetic study in the TUB region evidenced minimal genetic distances of rosemary strain from both F. solani and F. falciforme. Overall, the results obtained after the TEF sequencing and the phylogenetic studies of MH 1410.3 backed up the identification as F. falciforme, also a member of FSSC. In fact, FSSC includes 60 phylogenetic species [29] falling into three clades: according to these clusterings, F. falciforme is included in clades 3 + 4 while F. solani sensu stricto is assigned to the clade 5 [30]. The controversial output of BLASTn alignment and phylogenetic analyses for FSSC members was previously reported in diagnostic investigations conducted on root rot of watermelon in Mexico; here, phylogenetic elaboration allowed to identify as F. falciforme a Fusarium strain initially assigned to F. solani [31].
Among the three fungal species detected in rosemary plants, M. phaseolina was the most frequently isolated from necrotic roots, showing the typical charcoal rot caused by this pathogen. Indeed, charcoal rot is largely reported to be associated with M. phaseolina on more than 700 plant species [28], and it was reported on some aromatic species such as catmint in India [32], common sage [33], lemon balm [34] in Turkey and, in 2024, on lavender [35] but, to the best of our knowledge, this is the first report of M. phaseolina on rosemary in Italy and worldwide.
To a lesser extent, F. falciforme and, even less frequently, Ceratobasidium sp., were isolated from the rosemary field examined in this study. FSSC is one of the most widespread pathogen complexes in the globe and it is responsible for the foot and/or root rot of infected plants, evolving in a range of diverse symptomatology in the aerial part (yellowing, stunting, wilting), depending on the host [29]. References about the presence of FSSC members on Lamiaceae family are quite rare: indeed, F. solani had previously been reported only on Lavandula angustifolia (lavender) as a causal agent of wilting and browning of vascular tissues [36], and on S. officinalis [37,38,39]; however, this work describes, for the first time, F. falciforme as a pathogen of rosemary plants. Conversely, Ceratobasidium sp. (also called binucleate Rhizoctonia) is already documented as a plant pathogen of rosemary; in fact, a strain belonging to the anastomosis group AG-G, identified as Ceratorhiza fragariae (binucleate Rhizoctonia), was reported on rosemary in Spain [14], while Rhizoctonia anastomosis groups AG-A, AG-B (o), AG-G and AG-K were identified as causal agents of root rot of strawberry (Fragaria × ananassa) in Albania [40], in India [41], in Kyrgyzstan [42] and in Spain [43]. On the contrary, several binucleate Rhizoctonia strains are reported as mycorrhizal symbionts and biocontrol agents [44,45]; indeed, strains belonging to the anastomosis groups AG-A, AG-B(a), AG-B(o), AG-G, AG-I, AG-K and AG-P were described as antagonistic strains against the causal agents of dumping-off and root rot disease, acting by inducing systemic acquired resistance (SAR) on soybean [46], bean [47] and rice [44]. However, Ceratobasidium is also known to act as mycoparasite of plant pathogens; in fact, mycoparasitism was observed versus Pythium ultimum var. sporangiiferum [48].
Regarding the rosemary isolate’s role in pathogenicity, the Koch’s postulate assays performed in this study confirmed the pathogenic nature of M. phaseolina and F. falciforme strains. In detail, MP was able to induce the highest number of plants with the highest symptom level while F. falciforme FS MH 1410.3, although also pathogenic on rosemary, acts with a weaker virulence. Previously, a similar pathogen complex was reported in studies on Cucumis melo (muskmelon) [49]; in this case, Nascimento and coworkers proved that root rot was associated with multiple pathogens: R. solani, M. phaseolina, M. cannonballus and F. solani. The authors observed that, in natural infestation, M. phaseolina was isolated from plants with the highest level of wilting severity, while F. solani was the most frequently isolated pathogen, causing a lower level of disease [49].
Finally, regarding CS, the effects of inoculation with this isolate proved a low pathogenicity of the strain MH 1410.10. Overall, all the tested strains (CS, MP and FS), caused symptoms in rosemary plants, but their effectiveness in generating disease decreases from MP to FS and from this to CS. It should be noted that CS was reisolated also from non-symptomatic plants, demonstrating that it is able to colonize tissues without manifestation of symptoms.
Once the pathogenic attitude of the strains were verified singularly, the role in the rosemary desiccation and root necrosis of the three isolates was studied by multiple inoculations in all the combinations.
Combining data on disease incidence and severity, the disease index (DI) confirmed the aggressivity of M. phaseolina compared to the other two fungal species when inoculated individually, while DI decreased on plants infected with MP in association with FS or CS, suggesting a competitive behavior of the latter strains versus MP. These results reflected the behavior of the strains in in vitro dual cultures, where the CS and FS colonies showed a certain competition towards MP. The competitive behavior of Ceratobasidium sp. is supported by previous studies conducted by Mosquera-Espinosa et al. [44] on rice: in that study, the authors observed the appearance of slight symptoms of sheath blight due to inoculation of rice leaves with Ceratobasidium strains from orchids. These symptoms were lighter compared to those caused by a R. solani pathogenic strain from rice but, at the same time, plants pre-inoculated with Ceratobasidium were less susceptible to sheath blight caused by the same R. solani strain when inoculated alone [44]. It can be speculated that M. phaseolina (synonymous Rhizoctonia bataticola) and R. solani share the same ecological niche and it is likely that they compete for nutritive resources, which might induce reduction in MP population, with a corresponding depletion in the infection rate and in the attenuation of its aggressiveness [50].
Unlike what observed about MP and the effects of its co-infection with CS and FS, DI value registered after infection of FS + CS was higher than that obtained with FS or CS alone, demonstrating that in this case, the antagonistic activity of CS on FS could not be stated. In contrast, a cumulative effect of FS and CS could be hypothesized, confirming a previous observation on the root rot agents of melon [49] and watermelon [31]. In fact, among the pathogens tested in those studies on melon, F. solani was shown to have an intermediate aggressiveness further powered by Ceratobasidium sp. presence [49], with the incidence of the disease increasing when F. solani was inoculated together with Ceratobasidium sp. [31] in the case of watermelon root rot.
When the selected isolates (CS, MP and FS) from the three species were inoculated all together on rosemary plants, the recorded DI was comparable to those recorded in the pairs, higher than FS and CS separately and lower than that of MP alone. These results confirmed again a reduced level of disease aggressiveness of the three isolates compared to MP alone.
Reisolations after in vivo tests enabled the investigation of the success of the infection process by the pathogens and further confirmed the pathogenicity role of M. phaseolina and F. falciforme in multiple infections. In fact, when inoculated in pairs (FS + CS and FS + MP), FS was reisolated from all the plants, appearing undisturbed by the presence of the two other infecting strains; conversely, CS and MP were reisolated only by about 50% of plants. The percentages of reisolation obtained resembled the results of isolation from the field samples, where the most frequently isolated strain was M. phaseolina; this evidence suggests that, with MP being a necrotrophic pathogen, it could benefit from the presence of rotten tissues caused by multiple infections in the final stages of tissue colonization; in addition, the typical fast growth of MP mycelium on culture media may have contributed to effective isolation from plant tissues.
Unlike MP and FS, CS was poorly reisolated when plants were inoculated with the three isolates and in pairs, although, in vitro, it was not inhibited by FS or partially inhibited by MP. This could be explained by a lower ability of CS to colonize plant tissues compared to MP and FS. Furthermore, it can be hypothesized that, in plant tissue infection, F. falciforme exploits the mutual antagonism of MP and CS, as observed in vitro, managing to overcome the competition of the latter. Therefore, the interaction among strains in planta was partially explained by the interaction of the three pathogens in vitro but both tests confirmed the prevalence of MP.
All these findings allow us to assess that M. phaseolina and F. falciforme are agents of the desiccation and charcoal rot observed in rosemary fields; they can act together, but the symptom severity observed by multiple infections is mitigated in comparison to that observed after MP single infection. The role of Ceratobasidium sp. is controversial, as it can act as a saprophyte (infecting asymptomatic plants), an antagonist (versus M. phaseolina) or a pathogen (in synergy with F. falciforme).
In the literature, multiple pathogen infections in rosemary have already been investigated. In 2010, Ashrafi and co-workers found diseased rosemary plants infected with R. solani, Fusarium spp. and Phytohophthora citrophthora, one of the main causal agents of wilting in rosemary [51]. The authors demonstrated that, among the single pathogen infections, the one caused by P. citrophthora developed the most severe symptoms with the highest number of wilted plants. In contrast to our study, when the pathogens were combined, the occurrence of the disease increased [51], indicating a synergistic effect of the pathogens in all combinations. Multiple infections on rosemary were also reported in Arizona where the co-existence of R. solani, Fusarium oxysporum and Alternaria spp. was proved in a cultivation displaying symptoms of wilting diseases. In order to unravel the development of the disease, the authors performed incubation with the pathogens alone or in all the possible combinations under controlled conditions, but unfortunately, the experiments did not explain the mechanism behind such disease; thus, the plants remained asymptomatic in all the cases [52].
5. Conclusions
This work reports the first description of M. phaseolina and F. falciforme as etiological agents of desiccation and root rot of rosemary. They can act alone or in complex with Ceratobasidium sp., whose role shifts from antagonist to saprophyte or to pathogen, depending on the pathogen–plant interaction. M. phaseolina is responsible for the most severe symptoms when it acts alone while it is partially attenuated by the presence of the other fungal species. On the contrary, F. falciforme and Ceratobasidium sp. amplify their aggressiveness when acting in synergy. This phytopathological observation represents a new case of disease caused by pathogen complex and highlights the crucial role of the employment of appropriate diagnostic tools in pathogen detection; moreover, this case study sheds a light on the importance in the understanding of pathogen roles in the plant infection process, not only for diagnostic purposes but also for adopting proper management strategies. This aspect takes on even greater significance when symptoms are not specific.
Author Contributions
Conceptualization, E.D. and L.S.; methodology, E.D. and L.S.; software, E.D.; validation, L.S.; formal analysis, E.D.; investigation, E.D., G.S., G.R., G.N. and L.S.; resources, L.S.; data curation, E.D.; writing—original draft preparation, E.D. and A.M.; writing—review and editing, L.S.; visualization, E.D. and L.S.; supervision, L.S.; project administration, L.S.; funding acquisition, L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research is funded by Ager Agroalimentare e Ricerca—Grant number: 2022-2985.
Data Availability Statement
Data is contained within the article.
Acknowledgments
The authors thank Boris Basile, Department of Agricultural Sciences, University of Naples Federico II, Portici, Naples, Italy, for comments on the early version of the manuscript and Rosa Marzuillo, freelance agronomist, for collaboration in field inspections and sampling.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mwithiga, G.; Maina, S.; Gitari, J.; Muturi, P. Rosemary (Rosmarinus officinalis L.) growth rate, oil yield and oil quality under differing soil amendments. Heliyon 2022, 8, e09277. [Google Scholar] [CrossRef]
- Datiles, M.J.; Acevedo-Rodríguez, P. Rosmarinus officinalis (rosemary). CABI Compend. 2022. [Google Scholar] [CrossRef]
- Gurbuz, B.; Bagdat, R.B.; Uyanik, M.; Rezaeieh, K.A.P. Rosemary (Rosmarinus officinalis L.) cultivation studies under Ankara ecological conditions. Ind. Crops Prod. 2016, 88, 12–16. [Google Scholar] [CrossRef]
- Macaluso, D.; Licciardo, F.; Carbone, K. Farming of Medicinal and Aromatic Plants in Italy: Structural Features and Economic Results. Agriculture 2024, 14, 151. [Google Scholar] [CrossRef]
- Park, M.J.; Han, J.G.; Shin, H.D. First Korean report of rosemary powdery mildew caused by Golovinomyces biocellatus. Plant Pathol. 2010, 59, 408. [Google Scholar] [CrossRef]
- Wichura, A.; Braun, U.; Weber, R.W.S.; Hildebrands, A. Golovinomyces orontii and other powdery mildews on Rosmarinus officinalis. Plant Pathol. 2012, 2, 162–166. [Google Scholar] [CrossRef]
- Suthaparan, A.; Solhaug, K.A.; Bjugstad, N.; Gislerød, H.R.; Gadoury, D.M.; Stensvand, A. Suppression of powdery Mildews by UV-B: Application frequency and timing, dose, reflectance, and automation. Plant Dis. 2016, 100, 1643–1650. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, L.A.; Pérez-Sierra, A.; Armengol, J.; García-Jiménez, J. Characterization of Phytophthora nicotianae isolates causing collar and root rot of lavender and rosemary in Spain. J. Plant Pathol. 2007, 89, 261–264. [Google Scholar]
- Zarandi, D.M.; Rezaee, S.; Aminaee, M.M.; Sharzei, A. First report of Fusarium sambucinum on rosemary plant in Iran. New Dis. Rep. 2014, 29, 16. [Google Scholar] [CrossRef]
- Putnam, M.L. First report of stem rot of rosemary caused by Sclerotinia sclerotiorum in the United States. Plant Pathol. 2004, 53, 252. [Google Scholar] [CrossRef]
- Garibaldi, A.; Bertetti, D.; Pensa, P.; Matić, S.; Gullino, M.L. First Report of white mould caused by Sclerotinia sclerotiorum on Rosemary (Rosmarinus officinalis L.) in Italy. J. Plant Pathol. 2018, 99, 543. [Google Scholar]
- Garibaldi, A.; Bertetti, D.; Pensa, P.; Poli, A.; Gullino, M.L. First report of web blight on rosemary (Rosmarinus officinalis) caused by Rhizoctonia solani AG-1-IA in Italy. Plant Dis. 2013, 97, 844. [Google Scholar] [CrossRef] [PubMed]
- Azcona, J.S.; Woodhall, J.W.; Perkins, K.; Henderson, D.; Barnes, A.V.; Wharton, P.S.; Henry, C. First report of Rhizoctonia solani AG1-IB on Rosmarinus officinalis in the United Kingdom. New Dis. Rep. 2017, 35, 12. [Google Scholar] [CrossRef]
- González, V.; Garcés-Claver, A.; Ibarra, N. First report of root rot on Rosmarinus officinalis caused by Ceratorhiza fragariae (Binucleate rhizoctonia) in Spain. Plant Dis. 2017, 101, 1542. [Google Scholar] [CrossRef]
- Mekonnen, M.; Wariyo, A.; Hilu, G. Antifungal activities of some essential oils against Fusarium oxysporum of rosemary and sage plants. Adv. Crop Sci. Tech. 2019, 7, 419. [Google Scholar] [CrossRef]
- Pscheidt, J.W. Rosemary (Rosmarinus officinalis)–Botrytis Stem Canker. In Pacific Northwest Pest Management Handbooks, Plant Disease; 2021; Available online: https://pnwhandbooks.org/plantdisease/host-disease/rosemary-rosmarinus-officinalis-botrytis-stem-canker (accessed on 7 June 2025).
- Rachid Lahlali, R.; Taoussi, M.; Laasli, S.E.; Gachara, G.; Ezzouggari, R.; Belabess, Z.; Aberkani, K.; Assouguem, A.; Meddich, A.; El Jarroudi, M.; et al. Effects of climate change on plant pathogens and host-pathogen interactions. Crop Environ. 2024, 3, 159–170. [Google Scholar] [CrossRef]
- Yamamoto, D.T.; Uchida, J.Y. Rapid Nuclear Staining of Rhizoctonia solani and Related Fungi with Acridine Orange and with Safranin O. Mycologia 1982, 74, 145–149. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 9780123721808. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Nosratabadi, M.; Kachuei, R.; Rezaie, S.; Harchegani, A.B. Beta-tubulin gene in the differentiation of Fusarium species by PCR-RFLP analysis. Infez. Med. 2018, 26, 52–60. [Google Scholar] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Sonawane, A.; Mahjan, M.; Renake, S. Antifungal activity of a fungal isolate against pomegranate wilt pathogen Fusarium. Int. J. Curr. Microbiol. Appl. Sci. 2015, 2, 48–57. [Google Scholar]
- McKinney, H.H. A new system of grading plant diseases. Agric. Res. 1923, 26, 95–98. [Google Scholar]
- Anonymous. PM 7/129 (2) DNA barcoding as an identification tool for a number of regulated pests. EPPO Bull. 2021, 51, 100–143. [Google Scholar] [CrossRef]
- Sharon, M.; Kuninaga, S.; Hyakumachi, M.; Naito, S.; Sneh, B. Classification of Rhizoctonia spp. using rDNA-ITS sequence analysis supports the genetic basis of the classical anastomosis grouping. Mycoscience 2008, 49, 93–114. [Google Scholar] [CrossRef]
- Dell’Olmo, E.; Tripodi, P.; Zaccardelli, M.; Sigillo, L. Occurrence of Macrophomina phaseolina on Chickpea in Italy: Pathogen Identification and Characterization. Pathogens 2022, 11, 842. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.J. The Fusarium solani species complex: Ubiquitous pathogens of agricultural importance. Mol. Plant Pathol. 2016, 17, 146–158. [Google Scholar] [CrossRef]
- Sabahi, F.; Banihashemi, Z.; Mirtalebi, M.; Rep, M.; Cacciola, S.O. Molecular Variability of the Fusarium solani Species Complex Associated with Fusarium Wilt of Melon in Iran. J. Fungi. 2023, 9, 486. [Google Scholar] [CrossRef]
- Rentería-Martínez, M.E.; Guerra-Camacho, M.A.; OchoaMeza, A.; Moreno-Salazar, S.F.; Varela-Romero, A.; Gutiérrez-Millán, L.E.; Meza-Moller, A.C. Multilocus phylogenetic analysis of fungal complex associated with root rot watermelon in Sonora, Mexico. Rev. Mex. De Fitopatol. 2018, 36, 233–255. [Google Scholar] [CrossRef]
- Nishad, I.; Srivastava, A.K.; Saroj, A.; Babu, B.K.; Samad, A. First Report of Root Rot of Nepeta cataria Caused by Macrophomina phaseolina in India. Plant Dis. 2018, 102, 2380. [Google Scholar] [CrossRef]
- Koşar, İ.; Güney, İ.G.; Derviş, S.; Kırlı, O.; Özer, G. First report of Macrophomina phaseolina causing charcoal rot on common sage (Salvia officinalis) in Turkey. J. Plant Pathol. 2021, 103, 1371. [Google Scholar] [CrossRef]
- Koşar, I.; Güney, I.G.; Üner, S.E.; Özer, G.; Derviş, S. First report of Macrophomina phaseolina causing charcoal rot on lemon balm (Melissa officinalis) in Turkey. J. Plant Pathol. 2022, 104, 895. [Google Scholar] [CrossRef]
- Palacıoğlu, G.; Ören, E.; Baran, B.; Orak, A.B. First report of Macrophomina phaseolina causing charcoal rot on lavender (Lavandula angustifolia) in Turkey. J. Plant Pathol. 2024, 107, 743. [Google Scholar] [CrossRef]
- Rena, Y.Z.; Tana, H.; Lib, Z.J.; Dua, J.; Lia, H. First report of lavender wilt caused by Fusarium solani in China. Plant Pathol. 2008, 57, 377. [Google Scholar] [CrossRef]
- Sunanda, C.R. Studies on Root Rot of Sage (Salvia officinalis L.) and Its Management. Master’s Thesis, University of Agricultural Science, Bangalore, India, 2000; p. 89. [Google Scholar]
- Omar, A.M.; Ahmed, A.I.S. Antagonistic and Inhibitory Effect of Some Plant Rhizo-Bacteria Against Different Fusarium Isolates on Salvia officinalis. Am.-Eurasian J. Agric. Environ. Sci. 2014, 12, 1437–1446. [Google Scholar]
- Mallesh, S.B.; Narendrappa, T.; Kumari. Management of Root Rot of Sage (Salvia officinalis) Caused by Fusarium solani and Rhizoctonia solani. Int. J. Plant Prot. 2009, 2, 261–264. [Google Scholar]
- Cara, M.; Merkuri, J.; Salliu, A.; Vojvodić, M.; Knežević, I.; Grkinić, M.; Bulajić, A. Binucleate rhizoctonia AG-A causing black root rot of strawberry in Albania. J. Phytopathol. 2024, 172, e13265. [Google Scholar] [CrossRef]
- Baiswar, P.; Ngachan, S.V. First Report of Root and Collar Rot of Strawberry (Fragaria × ananassa) Caused by Ceratobasidium sp. AG-B(o) (Binucleate rhizoctonia) in India. Plant Dis. 2018, 102, 1035. [Google Scholar] [CrossRef]
- Erper, I.; Ozer, G.; Yildirim, E.; Zholdoshbekova, S.; Turkkan, M. First report of root rot on strawberry caused by Binucleate rhizoctonia AG-G and AG-K in Kyrgyzstan. J. Plant Pathol. 2022, 104, 387–388. [Google Scholar] [CrossRef]
- Borrero, C.; Avilés-García, I.; López, N.; Avilés, M. First Report of Root Rot on Strawberry Caused by Binucleate rhizoctonia AG-K in Spain. Plant Dis. 2018, 103, 376. [Google Scholar] [CrossRef]
- Mosquera-Espinosa, A.T.; Bayman, P.; Prado, G.A.; Gómez-Carabalí, A.; Otero, J.T. The double life of Ceratobasidium: Orchid mycorrhizal fungi and their potential for biocontrol of Rhizoctonia solani sheath blight of rice. Mycologia 2013, 105, 141–150. [Google Scholar] [CrossRef]
- Manrique-Barros, S.; Flanagan, N.S.; Ramírez-Bejarano, E.; Mosquera-Espinosa, A.T. Evaluation of Tulasnella and Ceratobasidium as Biocontrol Agents of Fusarium Wilt on Vanilla planifolia. Agronomy 2023, 13, 2425. [Google Scholar] [CrossRef]
- Khan, F.U.; Nelson, B.D.; Helms, T.C. Greenhouse evaluation of Binucleate rhizoctonia for control of R. solani in soybean. Plant Dis. 2005, 89, 373–379. [Google Scholar] [CrossRef]
- Wen, K.; Seguin, P.; St.-Arnaud, M.; Jabaji-Hare, S. Real-Time Quantitative RT-PCR of Defense-Associated Gene Transcripts of Rhizoctonia solani-Infected Bean Seedlings in Response to Inoculation with a Non pathogenic Binucleate rhizoctonia Isolate. phytopathology 2005, 95, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Siwek, K.; Harris, A.R.; Scott, E.S. Mycoparasitism of Pythium ultimum by Antagonistic Binucleate rhizoctonia Isolates in Agar Media and on Capsicum Seeds. J. Phytopathol. 2008, 10, 417–423. [Google Scholar] [CrossRef]
- Nascimento, P.G.M.L.; Ambrósio, M.M.Q.; Freitas, F.C.L.; Cruz, B.L.S.; Dantas, A.M.M.; Sales Junior, R.; da Silva, W.L. Incidence of root rot of muskmelon in different soil management practices. Eur. J. Plant Pathol. 2018, 152, 433–446. [Google Scholar] [CrossRef]
- Dutt, A.; Andrivon, D.; Le May, C. Multi-infections, competitive interactions, and pathogen coexistence. Plant Pathol. 2022, 71, 5–22. [Google Scholar] [CrossRef]
- Ashrafi, S.J.; Rastegar, M.F.; Saremi, H. Rosemary wilting disease and its management by soil solarization technique in Iran. Afr. J. Biotechnol. 2010, 9, 7048–7057. [Google Scholar]
- Dura, S.; Hanson, S.F. Rosemary Decline, an Apparent Disease of Unknown Etiology Affecting the Southwestern US. WJASS 2024, 9, 1–4. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).