
A Review of Supporting Evidence, Limitations and Challenges of Using Cover Crops in Agricultural Systems

Abstract
1. Introduction
2. Methodology
3. Cover Crops and Biological Soil Health
3.1. Improving Soil Organic Matter
3.2. Soil Microbial Activity and Microbial Biomass Enhancement Through the Use of Cover Crops
3.3. Arbuscular Mycorrhizal Fungi (AMF)
3.4. Improving Earthworm Numbers (Macro-Fauna)
4. Physical Aspect of Soil Health
4.1. Enhancing Soil Bulk Density Using Cover Crops
4.2. Enhancing Aggregate Stability
5. Enhancing Soil Fertility
5.1. Increasing Total N Supply to the Soil Using Legumes
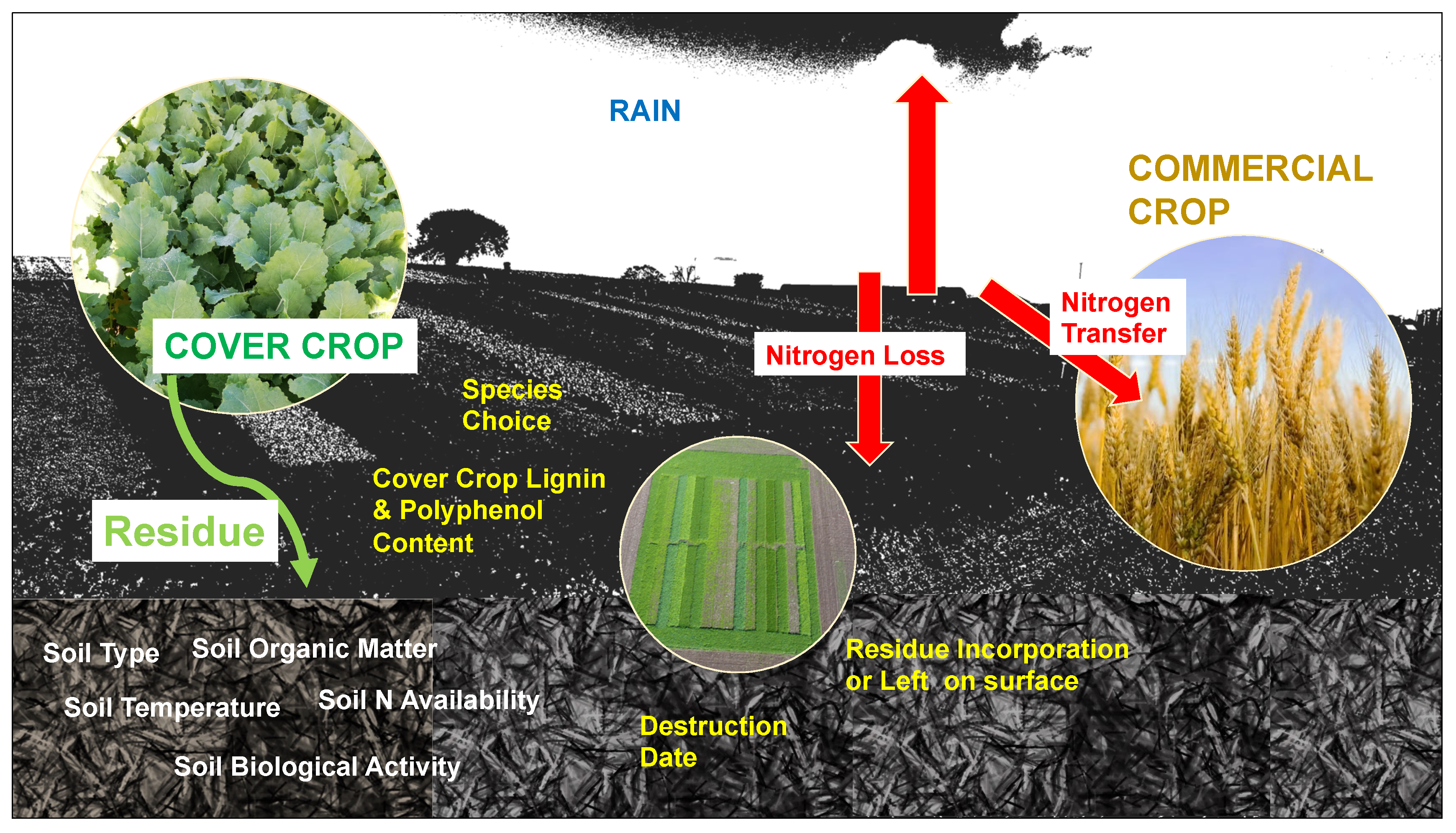
5.2. Increasing N Efficiency Through Reducing N Losses from Leaching
5.3. N Accumulation by Cover Crops
- Species
- Nutrients available (Soil mineral N)
- Light and temperature
- Biomass potential
- Planting date.

{kind=link}
{kind=link}
| Species | Biomass (t/ha) | N Uptake (kg/ha) | % N Leaching Reduction | Notes | Region | Author |
|---|---|---|---|---|---|---|
| Mustard | 2.05 | 36.9 | Switzerland | [102] | ||
| Pea | 2.68 | 76.5 | ||||
| Oat | 2.38 | 31.3 | ||||
| Phacelia | 1.78 | 31.1 | ||||
| Chicory | 1.39 | 25 | 71 | The deep roots were effective in depleting soil N from further down the soil profile | Switzerland | [102] |
| Fodder radish | 2.00 | 45 | 79 | |||
| Ryegrass | 1.05 | 22 | 67 | |||
| Oilseed radish | 2.6 | 57.3 | 96–97 | Starter fertiliser resulted in larger K uptakes 76.8 vs. 90 kg/ha (p < 0.05) | Switzerland | [102] |
| Oilseed radish + starter fertiliser | 2.8 | 79.4 | ||||
| Hairy vetch | 154 | Multiple sources | Switzerland | [102] | ||
| Rye | 30–61 | |||||
| Crimson clover | 28 | |||||
| White mustard | 57–116 | |||||
| Oilseed radish | 70–127 | |||||
| Mustard | 46 | 63 | Initial soil mineral nitrogen (SMN) 20 (kg/ha) | France | [103] | |
| Ryegrass | 44 | 59 | ||||
| Vetch | 61 | 32 | ||||
| Mustard | 62 | 51 | Initial soil mineral nitrogen (SMN) 60 (kg/ha) | |||
| Ryegrass | 56 | 47 | ||||
| Vetch | 56 | 24 | ||||
| Mustard | 80 | 48 | Initial soil mineral nitrogen (SMN) 80 (kg/ha) | |||
| Ryegrass | 70 | 43 | ||||
| Vetch | 54 | 22 | ||||
| Stubble | English site (Telford) | England | [104] | |||
| Oilseed radish | 66 | Equivalent to applying 40–60 kg N/ha | ||||
| White mustard | 57 | |||||
| Winter rye | 48 |
5.4. Integrating Organic Manures to Increase Biomass and Nutrient Assimilation
5.5. Enhancing N Supply from the Cover Crops to Commercial Crops
5.6. Phosphorous (P) Accumulation in Cover Crops and Enhancing Availability to Commercial Crops
5.7. Cover Crops Can Enhance Soil Potassium Cycling
5.8. Using Cover Crops to Enhance Micronutrient Supply to Commercial Crops
6. Weed, Pest and Disease Suppression via Cover Crops
6.1. Weed Suppression by Using Cover Crops
6.2. Soil Pest Suppression by Cover Crops
| Cover Crop | Application | Pest | Result | Location | Author |
|---|---|---|---|---|---|
| Brassica juncea L. (brown mustard) | Green manure | Glodera pallida (Potato cyst nematode) | Reduction in viable egg count | UK | [153] |
| Brassica juncea L. (brown mustard) | Green manure | Glodera pallida (Potato cyst nematode) | 50–95% reduction in juvenile population levels | UK | [154] |
| Brassica napus L. (Fodder rape) | Green manure | Meloidogyne chitwoodi (Root-knot nematode) | 89% reduction in population levels | USA | [162] |
| Raphanus sativus L. (Fodder radish) | Green manure | Glodera pallida (Potato cyst nematode) | Up to 100% in juvenile population levels and >95% reduction in viability of encysted eggs | UK | [154] |
| Raphanus sativus L. (Fodder radish) | Green manure | Glodera pallida (Potato cyst nematode) | 50% reduction in cyst population | USA | [163] |
| Sinapsis alba L. (White Mustard) | Green Manure | Glodera pallida (Potato cyst nematode) | 74% reduction in population levels | UK | [154] |
| Sinapsis alba L. (White Mustard) | Incorporated, fallow and unincorporated | Globodera rotochiensis (Potato cyst nematode) | No decrease | Belgium | [157] |
| Sinapsis alba L. (White Mustard) | Trap crop | Globodera rotochiensis and Glodera pallida (Potato cyst nematode) | 16% reduction in juvenile cyst levels | [156] |
6.3. Disease Suppression in the Commercial Crop by the Previous Cover Crop
| Species | Pot/Greenhouse or Field Trial | Cash Crop | Diseases Observed | Effect | Author |
|---|---|---|---|---|---|
| Brassica Napus Brassica Juncea Linum usitatissimum L. (Linola) | Both | Wheat (cereals) | Gaeumannomyces graminis var. triciti (Take-all) | Greater reduction in inoculum by brassica than by non-brassica (p = 0.05). | [170] |
| Brassica juncea | Field | Sugarbeet (Beta vulgaris L.) | Rhizoctnia solani | Reduction in disease incidence 2006, 2007 and 2008 found to be p < 0.001 | [166] |
| Avena sativa (Oats) Lolium multiflorum (Ryegrass) Brassica napus Raphanus sativa Sinapsis alba Brassica juncea | Pot and field trials | Potatoes | (Field trials) Powdery scab (Spongospora subterranea) | p = 0.05 | [165] |
| Black scurf (Rhizoctnia ultium) | p = 0.05 | ||||
| Common scab (Streptomycies scabiebi) | p = NS | ||||
| Brassica napus B. juncea S. alba Vicia dasycarpa (vetch) Pisum sativum (pea) Avena sativa (oat) | Field | Tomatoes (Solanum lycopersicum L.) | Fusarium wilt (Verticillium dahliae Fusarium spp.) | NS | [167] |
| Fallow was actually better for disease reduction. |
7. Environmental Considerations for Using Cover Crops
7.1. Improvement of Wildlife Diversity in Arable Fields
- Areas to forage
- Shelter from adverse weather
- Areas to breed.
7.2. Mitigating Against Pollution from Soil Erosion, Runoff and Nutrient Leaching Using Cover Crops
7.3. Reducing Nutrient Loss of Phosphorous (P)
8. Cover Crops and Commercial Crop Yield Improvement
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, B. Are Nitrogen Fertilizers Deleterious to Soil Health? Agronomy 2018, 8, 48. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Cheng, K.; Yue, Q.; Chadwick, D.; Espenberg, M.; Truu, J.; Rees, B.; Smith, P. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Change Biol. 2019, 25, 2530–2543. [Google Scholar] [CrossRef]
- Groff, S. The past, present, and future of the cover crop industry. J. Soil Water Conserv. 2015, 70, 130A–133A. [Google Scholar] [CrossRef]
- Boardman, J. Soil Erosion in Britain: Updating the Record. Agriculture 2013, 3, 418–442. [Google Scholar] [CrossRef]
- Stone, J.A.; Buttery, B.R. Nine forages and the aggregation of a clay loam soil. Can. J. Soil Sci. 1989, 69, 165–169. [Google Scholar] [CrossRef]
- Zhang, T.Q.; Tan, C.S.; Zheng, Z.M.; Welacky, T.; Wang, Y.T. Drainage water management combined with cover crop enhances reduction of soil phosphorus loss. Sci. Total Environ. 2017, 586, 362–371. [Google Scholar] [CrossRef] [PubMed]
- White, C.A.; Holmes, H.F.; Morris, N.L.; Stobart, R.M. A Review of the Benefits, Optimal Crop Management Practices and Knowledge Gaps Associated with Different Cover Crop Species; Agriculture and Horticulture Development Board: Cambridge, UK, 2016. [Google Scholar]
- Brust, J.; Weber, J.; Gerhards, R. Do cover crop mixtures have the same ability to suppress weeds as competitive monoculture cover crops? Julius-Kühn-Archiv 2014, 443, 422–430. [Google Scholar] [CrossRef]
- Henderson, D.R.; Riga, E.; Ramirez, R.A.; Wilson, J.; Snyder, W.E. Mustard biofumigation disrupts biological control by Steinernema spp. nematodes in the soil. Biol. Control 2009, 48, 316–322. [Google Scholar] [CrossRef]
- FAO. An international technical workshop. Investing in sustainable crop intensification The case for improving soil health. In Integrated Crop Management; Food and Agricuture Organization of the United Nations: Rome, Italiy, 2008; Volume 6. [Google Scholar]
- Doran, J. Soil health and global sustainability: Translating science into practice. Agric. Ecosyst. Environ. 2002, 88, 119–127. [Google Scholar] [CrossRef]
- Wittwer, R.; Dorn, B.; Jossi, W.; Van der Heijden, M. Cover crops support ecological intensification of arable cropping systems. Sci. Rep. 2017, 7, 41911. [Google Scholar] [CrossRef]
- Cassman, K.G. Ecological intensification of cereal production systems: Yield potential, soil quality, and precision agriculture. Proc. Natl. Acad. Sci. USA 1999, 96, 5952–5959. [Google Scholar] [CrossRef] [PubMed]
- Büchi, L.; Gebhard, C.; Liebisch, F.; Sinaj, S.; Ramseier, H.; Charles, R. Importance of cover crops in alleviating negative effects of reduced soil tillage and promoting soil fertility in a winter wheat cropping system. Agric. Ecosyst. Environ. 2018, 256, 92–104. [Google Scholar] [CrossRef]
- Gurjar, G.; Swami, S.; Telkar, S.; Meena, N.; Kant, K.; Kumar, R. Soil Biological Properties and Their Importance in Agricultural Production. Biomol. Rep. 2017, BR/10/17/03. [Google Scholar]
- FAO. Soil carbon sequestration. In SOLAW Background Thematic Report—TR04B; FAO: Rome, Italy, 2006. [Google Scholar]
- COP 21. 4 pour 1000 4 per 1000 Initiative 2018. Available online: https://www.4p1000.org/ (accessed on 20 February 2025).
- Sainju, U.; Whitehead, W.; Singh, A. Cover crops and nitrogen fertilization effects on soil aggregation and carbon and nitrogen pools. Can. J. Soil Sci. 2003, 83, 155–165. [Google Scholar] [CrossRef]
- Sapkota, T.B.; Askegaard, M.; Lægdsmand, M.; Olesen, J.E. Effects of catch crop type and root depth on nitrogen leaching and yield of spring barley. Field Crops Res. 2012, 125, 129–138. [Google Scholar] [CrossRef]
- Peng, Y.; Van Eerd, L.L. Surface soil sampling underestimates soil carbon and nitrogen storage of long-term cover cropping. Geoderma Reg. 2024, 39, e00885. [Google Scholar] [CrossRef]
- Piotrowska, A.; Wilczewski, E. Effects of catch crops cultivated for green manure and mineral nitrogen fertilization on soil enzyme activities and chemical properties. Geoderma 2012, 189–190, 72–80. [Google Scholar] [CrossRef]
- Haynes, R.J.; Beare, M.H. Influence of six crop species on aggregate stability and some labile organic matter fractions. Soil Biol. Biochem. 1997, 29, 1647–1653. [Google Scholar] [CrossRef]
- Yost, J.L.; Leytem, A.B.; Bjorneberg, D.L.; Dungan, R.S.; Schott, L.R. The use of winter forage crops and dairy manure to improve soil water storage in continuous corn in Southern Idaho. Agric. Water Manag. 2023, 277, 108074. [Google Scholar] [CrossRef]
- Dazzo, F.; Garoutte, A. Rhizosphere. In Reference Module in Life Science, 3rd ed.; Elsevier: Amsterdam, The Netherland, 2017. [Google Scholar]
- Benitez, M.-S.; Taheri, W.I.; Lehman, R.M. Selection of fungi by candidate cover crops. Appl. Soil Ecol. 2016, 103, 72–82. [Google Scholar] [CrossRef]
- Brennan, E.B.; Acosta-Martinez, V. Cover cropping frequency is the main driver of soil microbial changes during six years of organic vegetable production. Soil Biol. Biochem. 2017, 109, 188–204. [Google Scholar] [CrossRef]
- Haney, R.L.; Haney, E.B.; Smith, D.R.; Harmel, R.D.; White, M.J. The soil health tool—Theory and initial broad-scale application. Appl. Soil Ecol. 2018, 125, 162–168. [Google Scholar] [CrossRef]
- In ’t Zandt, D.; Fritz, C.; Wichern, F. In the land of plenty: Catch crops trigger nitrogen uptake by soil microorganisms. Plant Soil 2017, 423, 549–562. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, C.S.; Chauhan, P.S.; Bhatia, C.R. Changes in soil physico-chemical properties and microbial functional diversity due to 14 years of conversion of grassland to organic agriculture in semi-arid agroecosystem. Soil Tillage Res. 2010, 109, 55–60. [Google Scholar] [CrossRef]
- Li, L.; Xu, M.; Eyakub, A.M.; Zhang, W.; Duan, Y.; Li, D. Factors affecting soil microbial biomass and functional diversity with the application of organic amendments in three contrasting cropland soils during a field experiment. PLoS ONE 2018, 13, e0203812. [Google Scholar] [CrossRef]
- Barel, J.M.; Kuyper, T.W.; Paul, J.; de Boer, W.; Cornelissen, J.H.C.; De Deyn, G.B. Winter cover crop legacy effects on litter decomposition act through litter quality and microbial community changes. J. Appl. Ecol. 2019, 56, 132–143. [Google Scholar] [CrossRef]
- Briar, S.S.; Fonte, S.J.; Park, I.; Six, J.; Scow, K.; Ferris, H. The distribution of nematodes and soil microbial communities across soil aggregate fractions and farm management systems. Soil Biol. Biochem. 2011, 43, 905–914. [Google Scholar] [CrossRef]
- Overstreet, L.F.; Hoyt, G.D.; Imbriani, J. Comparing nematode and earthworm communities under combinations of conventional and conservation vegetable production practices. Soil Tillage Res. 2010, 110, 42–50. [Google Scholar] [CrossRef]
- Finney, D.; Buyer, J.S.; Kaye, J.P. Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Conserv. 2017, 72, 361–373. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142, 107701. [Google Scholar] [CrossRef]
- Frasier, I.; Noellemeyer, E.; Figuerola, E.; Erijman, L.; Permingeat, H.; Quiroga, A. High quality residues from cover crops favor changes in microbial community and enhance C and N sequestration. Glob. Ecol. Conserv. 2016, 6, 242–256. [Google Scholar] [CrossRef]
- Bossuyt, H.; Denef, K.; Six, J.; Frey, S.D.; Merckx, R.; Paustian, K. Influence of microbial populations and residue quality on aggregate stability. Appl. Soil Ecol. 2001, 16, 195–208. [Google Scholar] [CrossRef]
- Morgan, J.A.W.; Bending, G.D.; White, P.J. Biological costs and benefits to plant–microbe interactions in the rhizosphere. J. Exp. Bot. 2005, 56, 1729–1739. [Google Scholar] [CrossRef]
- Hao, X.; Abou Najm, M.; Steenwerth, K.L.; Nocco, M.A.; Basset, C.; Daccache, A. Are there universal soil responses to cover cropping? A systematic review. Sci. Total Environ. 2023, 861, 160600. [Google Scholar] [CrossRef]
- Chavarría, D.N.; Verdenelli, R.A.; Serri, D.L.; Restovich, S.B.; Andriulo, A.E.; Meriles, J.M.; Vargas-Gil, S. Effect of cover crops on microbial community structure and related enzyme activities and macronutrient availability. Eur. J. Soil Biol. 2016, 76, 74–82. [Google Scholar] [CrossRef]
- Zelles, L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: A review. Biol. Fertil. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- Torabian, S.; Kim, E.; Qin, R.; Sathuvalli, V.; Gollany, H.T.; Kleber, M. Soil microbial biomass influenced by cover crop after fumigation of potato fields. Sci. Total Environ. 2025, 958, 177910. [Google Scholar] [CrossRef]
- Lehman, R.M.; Taheri, W.I.; Osborne, S.L.; Buyer, J.S.; Douds, D.D. Fall cover cropping can increase arbuscular mycorrhizae in soils supporting intensive agricultural production. Appl. Soil Ecol. 2012, 61, 300–304. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Kabir, Z.; Koide, R.T. Effect of autumn and winter mycorrhizal cover crops on soil properties, nutrient uptake and yield of sweet corn in Pennsylvania, USA. Plant Soil 2002, 238, 205–215. [Google Scholar] [CrossRef]
- Rillig, M.C. Arbuscular mycorrhizae and terrestrial ecosystems processes. Ecology 2004, 7, 740–754. [Google Scholar] [CrossRef]
- Burak, K.; Yanardağ, İ.H.; Gómez-López, M.D.; Faz, Á.; Yalçin, H.; Sakin, E.; Ramazanoğlu, E.; Orak, A.B.; Yanardağ, A. The effect of arbuscular mycorrhizal fungi on biological activity and biochemical properties of soil under vetch growing conditions in calcareous soils. Heliyon 2024, 10, e24820. [Google Scholar] [CrossRef]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular Mycorrhizal Fungi Alleviate Drought Stress in C(3) (Leymus chinensis) and C(4) (Hemarthria altissima) Grasses via Altering Antioxidant Enzyme Activities and Photosynthesis. Front. Plant Sci. 2019, 10, 499. [Google Scholar] [CrossRef]
- García-González, I.; Quemada, M.; Gabriel, J.L.; Hontoria, C. Arbuscular mycorrhizal fungal activity responses to winter cover crops in a sunflower and maize cropping system. Appl. Soil Ecol. 2016, 102, 10–18. [Google Scholar] [CrossRef]
- Stobart, R.; Morris, N.L.; Fielding, H.; Leake, A.; Egan, J.; Burkinshaw, R. Developing the use of cover crops on farm through the Kellogg’s OriginsTM grower programme. Asp. Appl. Biol. 2015, 129, 27–34. [Google Scholar]
- Stroud, J.L.; Irons, D.E.; Watts, C.W.; Storkey, J.; Morris, N.L.; Stobart, R.M.; Fielding, H.A.; Whitmore, A.P. Population collapse of Lumbricus terrestris in conventional arable cultivations and response to straw applications. Appl. Soil Ecol. 2016, 108, 72–75. [Google Scholar] [CrossRef]
- Stroud, J.L.; Irons, D.E.; Watts, C.W.; Storkey, J.; Morris, N.L.; Stobart, R.M.; Fielding, H.A.; Whitmore, A.P. Cover cropping with oilseed radish (Raphanus sativus) alone does not enhance deep burrowing earthworm (Lumbricus terrestris) midden counts. Soil Tillage Res. 2017, 165, 11–15. [Google Scholar] [CrossRef]
- Curry, P. Some effects of animal manures on earthworms in grassland. Pedobiologia 1976, 16, 425–438. [Google Scholar] [CrossRef]
- Roarty, S.; Hackett, R.; Schmidt, O. Earthworm populations in twelve cover crop and weed management combinations. Appl. Soil Ecol. 2017, 114, 142–151. [Google Scholar] [CrossRef]
- Schmidt, O.; Clements, R.O.; Donaldson, G. Why do cereal–legume intercrops support large earthworm populations? Appl. Soil Ecol. 2003, 22, 181–190. [Google Scholar] [CrossRef]
- Chen, G. Alleviation of Soil Compaction by Brassica Cover Crops. In Geography; University of Maryland: College Park, MD, USA, 2009. [Google Scholar]
- Servadio, P.; Marsili, A.; Vignozzi, A.; Pellegrini, S.; Pagliai, M. Effects on some clay soil qualities following the passage of rubber-tracked and wheeled tractors in central Italy. Soil Tillage Res. 2001, 61, 143–155. [Google Scholar] [CrossRef]
- Chen, G.H.; Weil, R.R. Root growth and yield of maize as affected by soil compaction and cover crops. Soil Tillage Res. 2010, 117, 17–27. [Google Scholar] [CrossRef]
- Soil Science Society of America. Glossary of Soil Science Terms; Soil Science Society of America: Madison, WI, USA, 1997. [Google Scholar]
- Vaz, C.M.P.; Manieri, J.M.; de Maria, I.C.; Tuller, M. Modeling and correction of soil penetration resistance for varying soil water content. Geoderma 2011, 166, 92–101. [Google Scholar] [CrossRef]
- Lund, Z.F.; Elkins, C.B. Creating soil conditions favourable to rooting. Proc. Beltwide Cotton Prod. 1978, 91–92. [Google Scholar]
- Ren, L.; Nest, T.V.; Ruysschaert, G.; D’Hose, T.; Cornelis, W.M. Short-term effects of cover crops and tillage methods on soil physical properties and maize growth in a sandy loam soil. Soil Tillage Res. 2019, 192, 76–86. [Google Scholar] [CrossRef]
- Materechera, S.A.; Alston, A.M.; Kirby, J.M.; Dexter, A.R. Influence of root diameter on the penetration of seminal roots into a compacted subsoil. Plant Soil 1991, 144, 297–303. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Foloni, J.S.S.; Tiritan, C.S. Root growth and nutrient accumulation in cover crops as affected by soil compaction. Soil Tillage Res. 2002, 85, 109–115. [Google Scholar] [CrossRef]
- Chen, G.; Weil, R.R.; Hill, R.L. Effects of compaction and cover crops on soil least limiting water range and air permeability. Soil Tillage Res. 2014, 136, 61–69. [Google Scholar] [CrossRef]
- Chen, G.; Weil, R. Penetration of cover crop roots through compacted soil. Plant Soil 2010, 331, 31–43. [Google Scholar] [CrossRef]
- Welch, R.Y.; Behnke, G.D.; Davis, A.S.; Masiunas, J.; Villamil, M.B. Using cover crops in headlands of organic grain farms: Effects on soil properties, weeds and crop yields. Agric. Ecosyst. Environ. 2016, 216, 322–332. [Google Scholar] [CrossRef]
- Rabot, E.; Wiesmeier, M.; Schlüter, S.; Vogel, H.J. Soil structure as an indicator of soil functions: A review. Geoderma 2018, 314, 122–137. [Google Scholar] [CrossRef]
- Abiven, S.; Menasseri, S.; Chenu, C. The effects of organic inputs over time on soil aggregate stability—A literature analysis. Soil Biol. Biochem. 2009, 41, 1–12. [Google Scholar] [CrossRef]
- Oades, J.M. Mucilages at the root surface. J. Soil Sci. 1972, 29, 1–16. [Google Scholar] [CrossRef]
- Ali, W.; Hussain, S.; Chen, J.; Hu, F.; Liu, J.; He, Y.; Yang, M. Cover crop root-derived organic carbon influences aggregate stability through soil internal forces in a clayey red soil. Geoderma 2023, 429, 116271. [Google Scholar] [CrossRef]
- Haynes, R.J.; Beare, M.H. Aggregation and organic matter storage in meso-thermal, humid soils. In Advances in Soil Science: Structure and Organic Matter Storage in Agricultural Soils; Carter, B.A.S., Ed.; CRC Lewis: Boca Raton, FL, USA, 1995; pp. 213–263. [Google Scholar]
- Wright, S.F.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 1998, 198, 97–107. [Google Scholar] [CrossRef]
- Dai, W.; Feng, G.; Huang, Y.; Adeli, A.; Jenkins, J.N. Influence of cover crops on soil aggregate stability, size distribution and related factors in a no-till field. Soil Tillage Res. 2024, 244, 106197. [Google Scholar] [CrossRef]
- Celik, I.; Ortas, L.; Kilic, S. Effects of compost, mycorrhiza, manure and fertiliser on some physical properties of a chromoxerert soil. Soil Tillage Res. 2004, 78, 59–67. [Google Scholar] [CrossRef]
- Hartemink, A. Soil Fertility Decline: Definitions and Assessment. In Encyclopedia of Soil Science, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Ladha, J.; Padre, A.; K Reddy, C.; Cassman, K.; Verma, S.; Powlson, D.S.; Kessel, C.; Richter, D.; Chakraborty, D.; Pathak, S. Global nitrogen budgets in cereals: A 50-year assessment for maize, rice, and wheat production systems. Sci. Rep. 2016, 6, 19355. [Google Scholar] [CrossRef]
- Silgram, M.; Williams, D.; Wale, S.; Griffin-Walker, R.; Baddeley, J.A.; Jones, S.; Topp, C.F.E.; Watson, C.A.; Helming, J.; Stoddard, F.L. Biological Nitrogen Fixation (BNF) by Legume Crops in Europe. Legume Futures Report 1.5. 2014. Available online: https://www.legumehub.eu/wp-content/uploads/2021/06/Legume-Futures-Report-1.5.pdf (accessed on 20 February 2025).
- Thomas, R.B.; Van Bloem, S.J.; Schlesinger, W.H. Climate change and symbiotic nitrogen fixation in agroecosystems. In Agroecosystems in a Changing Climate; CRC Press: Boca Raton, FL, USA, 2006; pp. 85–116. [Google Scholar] [CrossRef]
- Büchi, L.; Gebhard, C.; Liebisch, F.; Sinaj, S.; Ramseier, H.; Charles, R. Accumulation of biologically fixed nitrogen by legumes cultivated as cover crops in Switzerland. Plant Soil 2015, 393, 273–287. [Google Scholar] [CrossRef]
- Li, X.; Sorensen, P.; Li, F.; Petersen, S.O.; Olesen, J.E. Quantifying biological nitrogen fixation of different catch crops, and residual effects of roots and tops on nitrogen uptake in barley using in-situ 15N labelling. Plant Soil 2015, 395, 273–287. [Google Scholar] [CrossRef]
- Carlsson, G.; Huss-Danell, K. Nitrogen fixation in perennial forage legumes in the field. Plant Soil 2003, 253, 353–372. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Aronsson, H.; Hansen, E.; Thomsen, I.K.; Liu, J.; Øgaard, A.; Kankanen, H.; Ulén, B. The ability of cover crops to reduce nitrogen and phosphorus losses from arable land in southern Scandinavia and Finland. J. Soil Water Conserv. 2016, 71, 41–55. [Google Scholar] [CrossRef]
- Gabriel, J.L.; Alonso-Ayuso, M.; García-González, I.; Hontoria, C.; Quemada, M. Nitrogen use efficiency and fertiliser fate in a long-term experiment with winter cover crops. Eur. J. Agron. 2016, 79, 14–22. [Google Scholar] [CrossRef]
- Couëdel, A.; Alletto, L.; Tribouillois, H.; Justes, É. Cover crop crucifer-legume mixtures provide effective nitrate catch crop and nitrogen green manure ecosystem services. Agric. Ecosyst. Environ. 2018, 254, 50–59. [Google Scholar] [CrossRef]
- De Notaris, C.; Peixoto, L.; Mortensen, E.Ø.; Rasmussen, J. Cover crop biomass production as a predictor of nitrogen fertilizer replacement value—Legumes secure positive effects. Agric. Ecosyst. Environ. 2025, 381, 109446. [Google Scholar] [CrossRef]
- Moreno-Cadena, P.; Salmeron, M.; Canisares, L.P.; Poffenbarger, H.J. Productivity benefits of cereal-legume cover crop mixtures under variable soil nitrogen and termination times. Eur. J. Agron. 2024, 155, 127114. [Google Scholar] [CrossRef]
- Couëdel, A.; Alletto, L.; Justes, E. Crucifer-legume cover crop mixtures provide effective sulphate catch crop and sulphur green manure services. Plant Soil 2018, 426, 61–76. [Google Scholar] [CrossRef]
- Cooper, R.J.; Hama-Aziz, Z.; Hiscock, K.M.; Lovett, A.A.; Dugdale, S.J.; Sünnenberg, G.; Noble, L.; Beamish, J.; Hovesen, P. Assessing the farm-scale impacts of cover crops and non-inversion tillage regimes on nutrient losses from an arable catchment. Agric. Ecosyst. Environ. 2017, 237, 181–193. [Google Scholar] [CrossRef]
- Teixeira, E.I.; Johnstone, P.; Chakwizira, E.; Ruiter, J.d.; Malcolm, B.; Shaw, N.; Zyskowski, R.; Khaembah, E.; Sharp, J.; Meenken, E.; et al. Sources of variability in the effectiveness of winter cover crops for mitigating N leaching. Agric. Ecosyst. Environ. 2016, 220, 226–235. [Google Scholar] [CrossRef]
- Meisinger, J.J.; Hargrove, W.; Mikkelsen, R.; Williams, J.; Benson, V. Effects of cover crops on groundwater quality. In Cover Crops for Clean Water; 1991; Available online: https://www.swcs.org/static/media/cms/CCCW4ground_D30E9A7F53D56.pdf (accessed on 20 February 2025).
- Hooker, K.; Coxon, C.; Hackett, R.; Kirwan, L.; O’Keeffe, E.; Richards, K. Evaluation of Cover Crop and Reduced Cultivation for Reducing Nitrate Leaching in Ireland. J. Environ. Qual. 2008, 37, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Premrov, A.; Coxon, C.E.; Hackett, R.; Kirwan, L.; Richards, K.G. Effects of over-winter green cover on soil solution nitrate concentrations beneath tillage land. Sci. Total Environ. 2014, 470–471, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Premrov, A.; Coxon, C.E.; Hackett, R.; Kirwan, L.; Richards, K.G. Effects of over-winter green cover on groundwater nitrate and dissolved organic carbon concentrations beneath tillage land. Sci. Total Environ. 2012, 438, 144–153. [Google Scholar] [CrossRef]
- Hashemi, M.; Farsad, A.; Sadeghpour, A.; Weis, S.; Herbert, S. Cover-crop seeding-date influence on fall nitrogen recovery. J. Plant Nutr. Soil Sci. 2013, 176, 69–75. [Google Scholar] [CrossRef]
- Kumar, U.; Thomsen, I.; Eriksen, J.; Vogeler, I.; Mäenpää, M.; Hansen, E. Delaying sowing of cover crops decreases the ability to reduce nitrate leaching. Agric. Ecosyst. Environ. 2023, 355, 108598. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K. Are differences in root growth of nitrogen catch crops important for their ability to reduce soil nitrate-N content, and how can this be measured? Plant Soil 2001, 230, 185–195. [Google Scholar] [CrossRef]
- Tribouillois, H.; Fort, F.; Cruz, P.; Charles, R.; Flores, O.; Garnier, E.; Justes, E. A Functional Characterisation of a Wide Range of Cover Crop Species: Growth and Nitrogen Acquisition Rates, Leaf Traits and Ecological Strategies. PLoS ONE 2015, 10, e0122156. [Google Scholar] [CrossRef]
- Kanders, M.J.; Berendonk, C.; Fritz, C.; Watson, C.; Wichern, F. Catch crops store more nitrogen below-ground when considering Rhizodeposits. Plant Soil 2017, 417, 287–299. [Google Scholar] [CrossRef]
- Wendling, M.; Büchi, L.; Amossé, C.; Jeangros, B.; Walter, A.; Charles, R. Specific interactions leading to transgressive overyielding in cover crop mixtures. Agric. Ecosyst. Environ. 2017, 241, 88–99. [Google Scholar] [CrossRef]
- Constantin, J.; Le Bas, C.; Justes, E. Large-scale assessment of optimal emergence and destruction dates for cover crops to reduce nitrate leaching in temperate conditions using the STICS soil–crop model. Eur. J. Agron. 2015, 69, 75–87. [Google Scholar] [CrossRef]
- Silgram, M.; Williams, D.; Wale, S.; Griffin-Walker, R. Managing Cultivations and Cover Crops for Improved Profitability and Envionmental Benefits in Potatoes; Agriculture and Horticulture Development Board: Warwickshire, UK, 2015. [Google Scholar]
- Frost, J.P.; Bailey, J.S.; Stevens, R.J. Making Best on-Farm Use of Plant Nutrients in Livestock Manures; 78th Annual Report; Agricultural Research Institute of Northern Ireland: Hillsborough, UK, 2004. [Google Scholar]
- Cambardella, C.A.; Moorman, T.B.; Singer, J.W. Soil nitrogen response to coupling cover crops with manure injection. Nutr. Cycl. Agroecosyst. 2010, 87, 383–393. [Google Scholar] [CrossRef]
- Parkin, T.B.; Kaspar, T.; Singer, J.W. Cover crop effects on the fate of N following soil application of swine manure. Plant Soil 2006, 289, 141–152. [Google Scholar] [CrossRef]
- Thilakarathna, M.; Serran, S.; Lauzon, J.; Janovicek, K.; Deen, B. Management of Manure Nitrogen Using Cover Crops. Agron. J. 2015, 107, 1595–1607. [Google Scholar] [CrossRef]
- Cottney, P.; Black, L.; White, E.; Williams, P.N. The Correct Cover Crop Species Integrated with Slurry Can Increase Biomass, Quality and Nitrogen Cycling to Positively Affect Yields in a Subsequent Spring Barley Rotation. Agronomy 2020, 10, 1760. [Google Scholar] [CrossRef]
- Corti, M.; Bechini, L.; Cavalli, D.; Ben Hassine, M.; Michelon, L.; Cabassi, G.; Pricca, N.; Perego, A.; Marino Gallina, P. Early sowing dates and pre-plant nitrogen affect autumn weed control and nitrogen content of winter cover crops in rotation with spring crops. Eur. J. Agron. 2024, 155, 127140. [Google Scholar] [CrossRef]
- Everett, L.; Wilson, M.; Pepin, R.; Coulter, J. Winter Rye Cover Crop with Liquid Manure Injection Reduces Spring Soil Nitrate but Not Maize Yield. Agronomy 2019, 9, 852. [Google Scholar] [CrossRef]
- Singer, J.W.; Cambardella, C.A.; Moorman, T.B. Enhancing Nutrient Cycling by Coupling Cover Crops with Manure Injection. Agron. J. 2008, 100, 1735–1739. [Google Scholar] [CrossRef]
- Janusauskaite, D.; Arlauskienė, A.; Maikštėnienė, S. Soil mineral nitrogen and microbial parameters as influenced by catch crops and straw management. Zemdirb. Agric. 2013, 100, 9–18. [Google Scholar] [CrossRef]
- Rosenfield, A.; Raynes, F. Sort out your soil: A practical guide to Green Manures. In Cotswold Grass Seeds Direct; Cotswold Seeds: Gloucestershire, UK, 2011. [Google Scholar]
- Justes, E.; Beaudoin, N.; Bertuzzi, P.; Charles, R.; Constantin, J.; Dürr, C.; Hermon, C.; Joannon, C.; Le Bas, B.; Mary, C.; et al. The use of cover crops to reduce nitrate leaching: Effect on the water and nitrogen balance and other ecosystem services. Synop. Study Rep. 2012, 68, 1–8. [Google Scholar]
- Fillery, I.R.P. The fate of biologically fixed nitrogen in legume-based dryland farming systems: A review. Aust. J. Exp. Agric. 2001, 41, 361–381. [Google Scholar] [CrossRef]
- Justes, E.; Mary, B.; Nicolardot, B. Quantifying and modelling C and N mineralization kinetics of catch crop residues in soil: Parameterization of the residue decomposition module of STICS model for mature and non mature residues. Plant Soil 2009, 325, 171–185. [Google Scholar] [CrossRef]
- Nevins, C.J.; Nakatsu, C.; Armstrong, S. Characterization of microbial community response to cover crop residue decomposition. Soil Biol. Biochem. 2018, 127, 39–49. [Google Scholar] [CrossRef]
- Kremen, A.; Weil, R. Nitrogen Mineralization from Brassica Cover Crops. Master’s Thesis, University of Maryland, College, College Park, MD, USA, 2006; p. 115. [Google Scholar]
- Silgram, M.; Harrison, R. The Mineralisation of Nitrogen in Cover Crops: A Review; NT1508; Department of Environment and Rural Affairs: London, UK, 1998.
- Melkonian, J.; Poffenbarger, H.J.; Mirsky, S.; Ryan, M.R.; Moebius-Clune, B. Estimating Nitrogen Mineralization from Cover Crop Mixtures Using the Precision Nitrogen Management Model. Agron. J. 2017, 109, 1944–1959. [Google Scholar] [CrossRef]
- Poffenbarger, H.; Mirsky, S.; Kramer, M.; Weil, R.; Meisinger, J.J.; Cavigelli, M.; Spargo, J. Cover Crop and Poultry Litter Management Influence Spatiotemporal Availability of Topsoil Nitrogen. Soil Sci. Soc. Am. J. 2015, 79, 1660–1673. [Google Scholar] [CrossRef]
- Wagger, M.G. Time of desiccation effects on plant composition and subsequent nitrogen release from several winter annual cover crops. Agron. J. 1989, 81, 236–241. [Google Scholar] [CrossRef]
- Ranells, N.N.; Wagger, M.G. Nitrogen Release from Grass and Legume Cover Crop Monocultures and Bicultures. Agron. J. 1996, 88, 777–882. [Google Scholar] [CrossRef]
- Wendling, M.; Büchi, L.; Amosse, C.; Sinaj, S.; Walter, A.; Charles, R. Nutrient accumulation by cover crops with different root systems. Asp. Appl. Biol. 2015, 129, 91–95. [Google Scholar]
- Hallama, M.; Pekrun, C.; Lambers, H.; Kandeler, E. Hidden miners—The roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 2018, 434, 7–45. [Google Scholar] [CrossRef]
- Dube, E.; Chiduza, C.; Muchaonyerwa, P. High biomass yielding winter cover crops can improve phosphorous availability in soil. S. Afr. J. Sci. 2014, 110, 1–4. [Google Scholar]
- Reynolds, S.H.; Ritz, K.; Crotty, F.H.; Stoate, C.; West, H.; Neal, A.L. Effects of cover crops on phosphatase activity in a clay arable soil in the UK. Asp. Appl. Biol. 2017, 136, 215–220. [Google Scholar]
- Stutter, M.I.; Shand, C.A.; George, T.S.; Blackwell, M.S.; Bol, R.; Mackay, R.L.; Richardson, A.E.; Condron, L.M.; Turner, B.L.; Haygarth, P.M. Recovering phosphorous from soil: A root solution? Environ. Sci. Technol. 2012, 46, 1977–1978. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Williams, P.; Zhang, H.; Yang, Y.; Yin, D.; Liu, Z.; Sun, H.; Luo, J. Combining Multiple High-Resolution In Situ Techniques to Understand Phosphorous Availability Around Rice Roots. Environ. Sci. Technol. 2021, 55, 13082–13092. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Zhou, Y.; Gu, Z.; Zhu, H.; Fu, S.; Yao, Q. The combined effects of cover crops and symbiotic microbes on phosphatase gene and organic phosphorus hydrolysis in subtropical orchard soils. Soil Biol. Biochem. 2015, 82, 119–126. [Google Scholar] [CrossRef]
- Cavigelli, M.A.; Thien, S.J. Phosphorous bioavailability following incorporation of green manure crops. Soil Sci. Soc. Am. 2003, 67, 1186–1194. [Google Scholar] [CrossRef]
- White, C.; Weil, R. Forage Radish Cover Crops Increase Soil Test Phosphorus Surrounding Radish Taproot Holes. Soil Sci. Soc. Am. J. 2011, 75, 121. [Google Scholar] [CrossRef]
- Crespo, C.; Wyngaard, N.; Sainz Rozas, H.R.; Pizzuto, A.; Barbagelata, P.; Barraco, M.; Gudelj, V.; Barbieri, P.A. Cover crops affect phosphorus fractions in soybean-based sequences with different phosphorus availability in Mollisols. Soil Tillage Res. 2024, 240, 106096. [Google Scholar] [CrossRef]
- Valadares, R.; Ávila-Silva, L.; Teixeira, R.; de Sousa, R.; Vergutz, L. Green Manures and Crop Residues as Source of Nutrients in Tropical Environment. In Organic Fertilizers—From Basic Concepts to Applied Outcomes; IntechOpen: London, UK, 2016; pp. 51–84. [Google Scholar]
- Murrell, T.S.; Mikkelsen, R.; Sulewski, G.; Norton, R.; Thompson, M. Improving Potassium Recommendations for Agricultural Crops; Springer: Cham, Swizerland, 2021. [Google Scholar]
- Rosolem, C.; Mallarino, A.; Nogueira, T. Considerations for Unharvested Plant Potassium; Springer: Cham, Swizerland, 2021; pp. 147–162. [Google Scholar]
- Rosolem, C.A.; Steiner, F. Effects of soil texture and rates of K input on potassium balance in tropical soil. Eur. J. Soil Sci. 2017, 68, 658–666. [Google Scholar] [CrossRef]
- Echer, F.; Peres, V.; Rosolem, C. Potassium application to the cover crop prior to cotton planting as a fertilization strategy in sandy soils. Sci. Rep. 2020, 10, 20404. [Google Scholar] [CrossRef]
- Wessells, K.R.; Brown, K.H. Estimating the global prevalence of zinc deficiency: Results based on zinc availability in national food supplies and the prevalence of stunting. Sci. Total Environ. 2012, 7, 1330–1343. [Google Scholar] [CrossRef]
- Jones, D.L.; Dennis, P.G.; Owen, A.G.; van Hees, P.A.W. Organic acid behavior in soils—Misconceptions and knowledge gaps. Plant Soil 2003, 248, 31–41. [Google Scholar] [CrossRef]
- Grüter, R.; Costerousse, B.; Bertoni, A.; Mayer, J.; Thonar, C.; Frossard, E.; Schulin, R.; Tandy, S. Green manure and long-term fertilization effects on soil zinc and cadmium availability and uptake by wheat (Triticum aestivum L.) at different growth stages. Sci. Total Environ. 2017, 599–600, 1330–1343. [Google Scholar] [CrossRef] [PubMed]
- Altomare, C.; Tringovska, L. Beneficial soil microorganisms, an ecological alternative for soil fertility management. Sustain. Agric. Res. Educ. 2011, 7, 161–214. [Google Scholar]
- Aghili, F.; Gamper, H.A.; Eikenberg, J.; Khoshgoftarmanesh, A.H.; Afyuni, M.; Schulin, R.; Jansa, J. Green manure addition to soil increases grain zinc concentration in bread wheat. PLoS ONE 2014, 9, 101487. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Khoshgoftarmanesh, A.H.; Afyuni, M.; Shrivani, M.; Schulin, R. The effect of preceding crop on wheat grain zinc concentration and its relationship to total amino acids and dissolved organic carbon in rhizosphere soil solution. Biol. Fertil. Soils 2014, 50, 239–247. [Google Scholar] [CrossRef]
- Habiby, H.; Afyuni, M.; Khoshgoftarmanesh, A.H.; Schulin, R. Effect of preceding crops and their residues on availability of zinc in a calcareous Zn-deficient soil. Biol. Fertil. Soils 2014, 50, 1061–1067. [Google Scholar] [CrossRef]
- Baraibar, B.; Hunter, M.; Schipanski, M.; Hamilton, A.; Mortensen, D. Weed Suppression in Cover Crop Monocultures and Mixtures. Weed Sci. 2017, 66, 121–133. [Google Scholar] [CrossRef]
- Bezuidenhout, S.R.; Reinhardt, C.F.; Whitwell, M.I. Cover crops of oats, stooling rye and three annual ryegrass cultivars influence maize and Cyperus esculentus growth. Weed Res. 2012, 52, 153–160. [Google Scholar] [CrossRef]
- Masilionyte, L.; Maiksteniene, S.; Kriauciuniene, Z.; Jablonskyte-Rasce, D.; Zou, L.; Sarauskis, E. Effect of cover crops in smothering weeds and volunteer plants in alternative farming systems. Crop Prot. 2017, 91, 74–81. [Google Scholar] [CrossRef]
- Gosling, P.; Nicholls, C. Managing Weeds in an Arable Rotation; Agriculture and Horticulture Development Board: Warwickshire, UK, 2014. [Google Scholar]
- U.S. Environmental Protection Agency. Ozone Layer Depletion—Regulatory Programs, Methyl Bromide Alternatives; U.S. Environmental Protection Agency: Washington, DC, USA, 2008.
- Fourie, H.; Ahuja, P.; Lammers, J.; Daneel, M. Brassicacea-based management strategies as an alternative to combat nematode pests: A synopsis. Crop Prot. 2016, 80, 21–41. [Google Scholar] [CrossRef]
- Ngala, B.M.; Haydock, P.P.J.; Woods, S.; Back, M.A. Biofumigation with Brassica juncea, Raphanus sativus and Eruca sativa for the management of field populations of the potato cyst nematode Globodera pallida. Pest Manag. Sci. 2014, 71, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.S.; Lazzeri, L.; Atkinson, A.H.; Urwin, P.E. Biofumigation for control of pale potato cyst nematodes: Activity of Brassica leaf extracts and green manures on Globodera pallida in vitro and in soil. J. Agric. Food Chem. 2011, 59, 7882–7890. [Google Scholar] [CrossRef]
- Kruger, D.; Fourie, J.; Malan, A. Cover Crops with Biofumigation Properties for the Suppression of Plant-Parasitic Nematodes: A Review. S. Afr. J. Enol. Vitic. 2013, 34, 287. [Google Scholar] [CrossRef]
- Scholte, K.; Vos, J. Effects of potential trap crops and planting date on soil infestation with potato cyst nematodes and root-knot nematodes. Ann. Appl. Biol. 2000, 137, 153–164. [Google Scholar] [CrossRef]
- Valdes, Y.; Viaene, N.; Moens, M. Effects of yellow mustard amendments on the soil nematode community in a potato field with focus on Globodera rostochiensis. Appl. Soil Ecol. 2012, 59, 39–47. [Google Scholar] [CrossRef]
- Gruver, L.S.; Weil, R.R.; Zasada, A.I.; Sardanelli, S.; Momen, B. Brassicaceae and rye cover crops altered free-living soil nematode community composition. Appled Soil Ecol. 2010, 10, 1–12. [Google Scholar]
- Ramirez, R.A.; Henderson, D.R.; Riga, E.; Lacey, L.A.; Snyder, W.E. Harmful effects of mustard bio-fumigants on entomopathogenic nematodes. Biol. Control 2009, 48, 147–154. [Google Scholar] [CrossRef]
- Jaffuel, G.; Blanco-Pérez, R.; Büchi, L.; Mäder, P.; Fließbach, A.; Charles, R.; Degen, T.; Turlings, T.C.J.; Campos-Herrera, R. Effects of cover crops on the overwintering success of entomopathogenic nematodes and their antagonists. Appl. Soil Ecol. 2017, 114, 62–73. [Google Scholar] [CrossRef]
- Everts, K. Soil Biology: Cover Crops and Disease Suppression; Sustainable Agriculture Research and Education: Griffin, GA, USA, 2016. [Google Scholar]
- Mojtahedi, H.; Santo, G.S.; Wilson, J.H.; Hang, A.N. Managing Meloidogyne chitwoodi on potato with rapeseed as green manure. Plant Dis. 1993, 77, 42–46. [Google Scholar] [CrossRef]
- Dossey, Z. Potential of Green Manure Biofumigants and Seed Exudates in the Control of Globodera pallida, the White Potato Cyst Nematode. Master’s Thesis, Washington State University, Pullman, WA, USA, 2010. [Google Scholar]
- Martinez, M.S. Influence of Cover Crops on the Development of Some Soil-Borne Plant Pathogens; Swedish University of Agricultural Science of Crop Production Ecology: Uppsala, Sweden, 2009; Volume 30. [Google Scholar]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop Prot. 2007, 26, 1067–1077. [Google Scholar] [CrossRef]
- Motisi, N.; Montfort, F.; Faloya, V.; Lucas, P.; Doré, T. Growing Brassica juncea as a cover crop, then incorporating its residues provide complementary control of Rhizoctonia root rot of sugar beet. Field Crops Res. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Hartz, T.K.; Johnstone, P.R.; Miyao, E.M.; Davis, R.M. Mustard Cover Crops Are Ineffective in Suppressing Soilborne Disease or Improving Processing Tomato Yield. Hortic. Sci. 2005, 40, 2016–2019. [Google Scholar] [CrossRef]
- Hansen, Z.R.; Keinath, A.P. Increased pepper yields following incorporation of biofumigation cover crops and the effects on soilborne pathogen populations and pepper diseases. Appl. Soil Ecol. 2013, 63, 67–77. [Google Scholar] [CrossRef]
- Falk, K.L.; Tokuhisa, J.G.; Gershenzon, J. The Effect of Sulfur Nutrition on Plant Glucosinolate Content: Physiology and Molecular Mechanisms. Plant Biol. 2007, 9, 573–581. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Sarwar, M.; Wong, P.T.W.; Mead, A.; Howe, G.; Newell, M. Field studies on the biofumigation of take-all by Brassica break crops. Aust. J. Agric. Res. 2000, 51, 445–456. [Google Scholar] [CrossRef]
- Holland, J.M.; Storkey, J.; Wutman, P.J.W.; Henderson, I.; Orson, J. The Farm4Bio project: Managing uncropped land for biodiversity. Asp. Appl. Biol. 2013, 118, 89–99. [Google Scholar]
- Heaney, I.; Foy, F.B.; Kennedy, J.A.; Crozier, G.; Walter, K.; O’ Connor, W.C.K. Impacts of agriculture on aquatic systems: Lessons learnt and new unknowns in Northern Ireland. Biol. Mar. Freshw. Res. 2001, 52, 151–163. [Google Scholar] [CrossRef]
- Wilcoxen, C.A.; Walk, J.W.; Ward, M.P. Use of cover crop fields by migratory and resident birds. Agric. Ecosyst. Environ. 2018, 252, 42–50. [Google Scholar] [CrossRef]
- Storr, T.; Simmons, R.W.; Hannam, J.A. A UK survey of the use and management of cover crops. Ann. Appl. Biol. 2019, 174, 179–189. [Google Scholar] [CrossRef]
- Gómez, J.A.; Sobrinho, T.A.; Giráldez, J.V.; Fereres, E. Soil management effects on runoff, erosion and soil properties in an olive grove of Southern Spain. Soil Tillage Res. 2009, 102, 5–13. [Google Scholar] [CrossRef]
- Granfors, D.A.; Church, K.E.; Smith, L.M. Eastern meadowlarks nesting in rangelands and conservation reserve program fields in Kansas. J. Field Ornithol. 1996, 67, 222–235. [Google Scholar]
- Hubbard, R.D.; Althoff, D.P.; Blecha, K.A.; Bruvold, B.A.; Japuntich, R.D. Nest site characteristics of eastern meadowlarks and grasshopper sparrows in tallgrass prairie at the Fort Riley military installation, Kansas. Trans. Kans. Acad. Sci. 2006, 109, 168–174. [Google Scholar] [CrossRef]
- Bryan, G.G.; Best, L.B. Avian nest density and success in grassed waterways in Iowa rowcrop fields. Wildl. Soc. Bull. 1994, 22, 583–592. [Google Scholar]
- EC. Soil Erosion Costs European Farmers €1.25 Billion a Year. EU Science Hub. 2018. Available online: https://european-union.europa.eu/index_en (accessed on 13 February 2020).
- Eurostat. Agri-Environmental Indicators—Soil Cover. 2012. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Agri-environmental_indicator_-_soil_cover (accessed on 20 February 2025).
- Fleskens, L.; Stroosnijder, L. Is soil erosion in olive groves as bad as often claimed? Geoderma 2007, 141, 260–271. [Google Scholar] [CrossRef]
- Joyce, B.A.; Wallender, W.W.; Mitchell, J.P.; Huyck, L.M.; Temple, S.R.; Brostrom, P.N. Infiltration and soil water storage under winter cover cropping in California’s Sacramento Valley. Am. Soc. Agric. Eng. 2002, 45, 315–326. [Google Scholar]
- Smith, R.V.; Jordan, C.; Annett, J.A. A phosphorus budget for Northern Ireland: Inputs to inland and coastal waters. J. Hydrol. 2005, 304, 193–202. [Google Scholar] [CrossRef]
- Foy, B.; Bailey, J.; Lennox, S.D. Mineral balances for the use of phosphorus and other nutrients by agriculture in Northern Ireland from 1925 to 2000—Methodology, trends and impacts of losses to water. Ir. J. Agric. Food Res. 2002, 41, 247–263. [Google Scholar]
- Kataoka, R.; Nagasaka, K.; Tanaka, Y.; Yamamura, H.; Shinohara, S.; Haramoto, E.; Hayakawa, M.; Sakamoto, Y. Hairy vetch (Vicia villosa), as a green manure, increases fungal biomass, fungal community composition, and phosphatase activity in soil. Appl. Soil Ecol. 2017, 117–118, 16–20. [Google Scholar] [CrossRef]
- Teboh, J.M.; Franzen, D.W. Buckwheat (Flagopyrum esculentum Monch) potential to contribute solubilized soil phosphorous to subsequent crops. Commun. Soil Sci. Plant Anal. 2011, 42, 1544–1550. [Google Scholar] [CrossRef]
- Kamh, M.; Horst, W.J.; Amer, F. Mobilization of soil fertilizer phosphate by cover crops. Plant Soil 1999, 211, 19–27. [Google Scholar] [CrossRef]
- AHDB. Nutrient Management Guide (RB209); AHDB: Warwickshire, UK, 2017. [Google Scholar]
- Smith, R.G.; Atwood, L.W.; Warren, N.D. Increased Productivity of a Cover Crop Mixture Is Not Associated with Enhanced Agroecosystem Services. PLoS ONE 2014, 9, e97351. [Google Scholar] [CrossRef] [PubMed]
- Marcillo, G.S.; Miguez, F.E. Corn yield response to winter cover crops: An updated meta-analysis. J. Soil Water Conserv. 2017, 72, 226–239. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, L.; Jacinthe, P.-A.; Ren, W. Global synthesis of cover crop impacts on main crop yield. Field Crops Res. 2024, 310, 109343. [Google Scholar] [CrossRef]
- Miguez, F.E.; Bollero, G.A. Review of Corn Yield Response under Winter Cover Cropping Systems Using Meta-Analytic Methods. Crop Sci. 2005, 45, 2318–2329. [Google Scholar] [CrossRef]
- Qin, Z.; Guan, K.; Zhou, W.; Peng, B.; Villamil, M.B.; Jin, Z.; Tang, J.; Grant, R.; Gentry, L.; Margenot, A.J.; et al. Assessing the impacts of cover crops on maize and soybean yield in the U.S. Midwestern agroecosystems. Field Crops Res. 2021, 273, 108264. [Google Scholar] [CrossRef]
- Toom, M.; Tamm, S.; Talgre, L.; Tamm, I.; Tamm, Ü.; Narits, L.; Hiiesalu, I.; Mae, A.; Lauringson, E. The Effect of Cover Crops on the Yield of Spring Barley in Estonia. Agriculture 2019, 9, 172. [Google Scholar] [CrossRef]
- Holland, J.; Brown, J.L.; MacKenzie, K.; Neilson, R.; Piras, S.; McKenzie, B.M. Over winter cover crops provide yield benefits for spring barley and maintain soil health in northern Europe. Eur. J. Agron. 2021, 130, 126363. [Google Scholar] [CrossRef]
- Stobart, M.R.; Morris, N.L. The impact of cover crops on yield and soils in the New Farming Systems programme. Asp. Appl. Biol. 2014, 127, 223–232. [Google Scholar]
- Hunter, M.C.; Schipanski, M.E.; Burgess, M.H.; LaChance, J.C.; Bradley, B.A.; Barbercheck, M.E.; Kaye, J.P.; Mortensen, D.A. Cover Crop Mixture Effects on Maize, Soybean, and Wheat Yield in Rotation. Agric. Environ. Lett. 2019, 4, 180051. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K. The effect of nitrogen catch crop species on the nitrogen nutrition of succeeding crops. Fertil. Res. 1994, 37, 227–234. [Google Scholar] [CrossRef]
- Lawson, A.; Cogger, C.; Bary, A.; Fortuna, A.-M. Influence of Seeding Ratio, Planting Date, and Termination Date on Rye-Hairy Vetch Cover Crop Mixture Performance under Organic Management. PLoS ONE 2015, 10, e0129597. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cottney, P.; Black, L.; White, E.; Williams, P.N. A Review of Supporting Evidence, Limitations and Challenges of Using Cover Crops in Agricultural Systems. Agriculture 2025, 15, 1194. https://doi.org/10.3390/agriculture15111194
Cottney P, Black L, White E, Williams PN. A Review of Supporting Evidence, Limitations and Challenges of Using Cover Crops in Agricultural Systems. Agriculture. 2025; 15(11):1194. https://doi.org/10.3390/agriculture15111194
Chicago/Turabian StyleCottney, Paul, Lisa Black, Ethel White, and Paul N. Williams. 2025. "A Review of Supporting Evidence, Limitations and Challenges of Using Cover Crops in Agricultural Systems" Agriculture 15, no. 11: 1194. https://doi.org/10.3390/agriculture15111194
APA StyleCottney, P., Black, L., White, E., & Williams, P. N. (2025). A Review of Supporting Evidence, Limitations and Challenges of Using Cover Crops in Agricultural Systems. Agriculture, 15(11), 1194. https://doi.org/10.3390/agriculture15111194