
Genetic Variability and Interrelationships of Grain, Cooking, and Nutritional Quality Traits in Cowpea: Implications for Cowpea Improvement


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area Description
2.2. Plant Materials
2.3. Experimental Design and Field Layout
2.4. Determination of Grain Quality Traits
2.5. Determination of Cooking Quality Traits
2.5.1. Cooking Time
2.5.2. Broth Viscosity
2.6. Determination of Nutritional Quality Traits
2.6.1. Digestion Procedure
2.6.2. Determination of Protein
2.6.3. Determination of Iron and Zinc
2.7. Data Analysis
3. Results
3.1. Frequency Distribution of Quantitative Traits
3.2. Genetic Variability and Mean Performance for Grain Quality Traits
3.3. Genetic Variability and Mean Performance for Cooking Quality Traits
3.4. Genetic Variability and Mean Performance for Nutritional Quality Traits
3.5. Correlation Coefficients among Grain, Cooking, and Nutritional Quality Traits of Cowpea Genotypes
3.6. Genetic Diversity among the Cowpea Genotypes
3.6.1. Principal Component Analysis for Grain Quality, Cooking Quality, and Nutritional Quality Traits
3.6.2. Hierarchical Clustering of Genotypes Based on Grain Quality, Cooking Quality and Nutritional Quality Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boukar, O.; Belko, N.; Chamarthi, S.; Togola, A.; Batieno, J.; Owusu, E.; Haruna, M.; Diallo, S.; Umar, M.L.; Olufajo, O.; et al. Cowpea (Vigna unguiculata): Genetics, genomics and breeding. Plant Breed. 2018, 138, 415–424. [Google Scholar] [CrossRef]
- Lonardi, S.; Muñoz-Amatriain, M.; Liang, Q.; Shu, S.; Wanamaker, S.; Lo, S.L.; Tanskanen, J.; Schulman, A.H.; Zhu, T.; Luo, M.C.; et al. The genome of cowpea (Vigna unguiculata [L.] Walp.). Plant J. Cell Mol. Biol. 2019, 98, 767–782. [Google Scholar] [CrossRef]
- Timko, M.P.; Rushton, P.J.; Laudeman, T.W.; Bokowiec, M.T.; Chipumuro, E.; Cheung, F.; Town, C.D.; Chen, X. Sequencing and analysis of the gene-rich space of cowpea. BMC Genom. 2008, 9, 103. [Google Scholar] [CrossRef]
- Badiane, F.A.; Diouf, M.; Diouf, D. Cowpea. In Broadening the Genetic Base of Grain Legumes; Singh, M., Bisht, I., Dutta, M., Eds.; Springer: New Delhi, India, 2014. [Google Scholar]
- Dakora, F.D.; Belane, A.K. Evaluation of protein and micronutrient levels in edible cowpea (Vigna unguiculata L. Walp.). Front. Sustain. Food Syst. 2019, 3, 70. [Google Scholar] [CrossRef]
- Boukar, O.; Massawe, F.; Muranaka, S.; Franco, J.; Maziya-Dixon, B.; Singh, B.; Fatokun, C. Evaluation of cowpea germplasm lines for protein and mineral concentrations in grains. Plant Genet. Resour. 2011, 9, 515–522. [Google Scholar] [CrossRef]
- Kirse, A.; Karklina, D. Integrated evaluation of cowpea (Vigna unguiculata (L.) Walp.) and maple pea (Pisum sativum var. Arvense L.) spreads. Agron. Res. 2015, 13, 956–968. [Google Scholar]
- Blair, M.; Astudillo, C.; Grusak, M.; Graham, R.; Beebe, S. Inheritance of seed iron and zinc content in common bean (Phaseolus vulgaris L.). Mol. Breed. 2009, 23, 197–207. [Google Scholar] [CrossRef]
- Affrifah, N.S.; Phillips, R.D.; Saalia, F.K. Cowpeas: Nutritional profile, processing methods and products—A review. Legume Sci. 2022, 4, e131. [Google Scholar] [CrossRef]
- Belane, A.; Dakora, F. Elevated concentrations of dietarily-important trace elements and macronutrients in edible leaves and grain of 27 cowpea (Vigna unguiculata L. Walp.) genotypes: Implications for human nutrition and health. Food Nutr. Sci. 2012, 3, 377–386. [Google Scholar] [CrossRef]
- Gerrano, A.S.; van Rensburg, W.S.J.; Adebola, P.O. Nutritional composition of immature pods in selected cowpea (Vigna unguiculata’(L.) Walp.) genotypes in South Africa. Aust. J. Crop Sci. 2017, 11, 134–141. [Google Scholar] [CrossRef]
- Gerrano, A.; Adebola, P.; van Rensburg, W.J.; Venter, S. Genetic variability and heritability estimates of nutritional composition in the leaves of selected cowpea genotypes [Vigna unguiculata (L.) Walp.]. HortScience A Publ. Am. Soc. Hortic. Sci. 2015, 50, 1435–1440. [Google Scholar] [CrossRef]
- FAOSTAT. Statistical Database. Food and Agriculture Organization of the United Nations. 2023. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 1 March 2023).
- Coulibaly, O.; Alene, A.D.; Abdoulaye, T.; Chianu, C.; Manyong, V.; Aitchedji, C.; Fatokun, D.; Kamara, A.; Ousmane, B.; Tefera, H. Baseline Assessment of Cowpea Breeding and Seed Delivery Efforts to Enhance Poverty Impacts in Sub-Saharan Africa; ICRISAT: Telengana, India, 2010. [Google Scholar]
- Chipeta, M.M.; Kampanje-Phiri, J.; Moyo, D.; Colial, H.; Tamba, M.; Belarmino, D.; Hella, J.; Yohane, E.; Mvula, N.; Kafwambira, J. Understanding specific gender dynamics in the cowpea value chain for key traits to inform cowpea breeding programs in Malawi, Mozambique and Tanzania. Front. Sociol. 2024, 9, 1254292. [Google Scholar] [CrossRef] [PubMed]
- Nkongolo, K.; Bokosi, J.; Malusi, M.; Vokhiwa, Z.; Mphepo, M. Agronomic, culinary, and genetic characterization of selected cowpea elite lines using farmers’ and breeder’s knowledge: A case study from Malawi. Afr. J. Plant Sci. 2009, 3, 147–156. [Google Scholar]
- Traore, K.; Sawadogo, P.; Batiéno, T.B.J.; Sawadogo, N.; Zongo, H.; Poda, S.L.; Tignegré, J.B.D.S.; Ouédraogo, T.J.; Sawadogo, M. Physical and cooking characteristics of six cowpea varieties seeds cultivated in Burkina Faso. Am. J. Plant Sci. 2022, 13, 7. [Google Scholar] [CrossRef]
- Egbadzor, K.F.; Dadoza, M.; Danquah, E.; Yeboah, M.; Offei, S.; Ofori, K. Genetic control of seed size in cowpea (Vigna unguiculata (L.) Walp). Int. J. Agric. Sci. 2013, 5, 367–371. [Google Scholar] [CrossRef]
- Egbadzor, K.F.; Danquah, E.Y.; Ofori, K.; Yeboah, M.; Offei, S.K. Diversity in 118 cowpea [Vigna unguiculate (L.) Walp] accessions assessed with 16 morphological traits. Int. J. Plant Breed. Genet. 2014, 8, 13–24. [Google Scholar] [CrossRef][Green Version]
- Gondwe, T.; Alamu, E.O.; Mdziniso, P.; Maziya-Dixon, B.B. Cowpea (Vigna unguiculata (L.) Walp) for food security: An evaluation of end-user traits of improved varieties in Swaziland. Sci. Rep. 2019, 9, 15991. [Google Scholar] [CrossRef]
- Hella, J.P.; Chilongo, T.; Mbwag, A.M.; Bokosi, J.; Kabambe, V.; Riches, C.; Massawe, C.L. Participatory market-led cowpea breeding in Sub- Saharan Africa: Evidence pathway from Malawi and Tanzania. Merit Res. J. Agric. Sci. Soil Sci. 2013, 1, 11–18. [Google Scholar]
- Herniter, I.A.; Lo, R.; Muñoz-Amatriaín, M.; Lo, S.; Guo, Y.-N.; Huynh, B.-L.; Lucas, M.; Jia, Z.; Roberts, P.A.; Lonardi, S.; et al. coat pattern QTL and development in cowpea (Vigna unguiculata [L.] Walp.). Front. Plant Sci. 2019, 10, 1346. [Google Scholar] [CrossRef]
- Kuldeep Tripathi, P.G.G.; Ahlawat, S.P.; Tyagi, V.; Semwal, D.P.; Gautam, N.K.; Rana, J.C.; Kumar, A. Cowpea genetic resources and its utilization: Indian perspective-A review. Legume Res. 2019, 42, 437–446. [Google Scholar] [CrossRef]
- Moses, D.R.; Zibokere, D.S. Effect of moisture content on some physical and mechanical properties of three varieties of cowpea (Vigna Unguiculata (L.) Walp). Agric. Eng. Int. CIGR J. 2011, 13, 1. [Google Scholar]
- de Oliveira Moura, J.; Rocha, M.D.M.; Gomes, R.L.F.; Freire, F.R.F.; Silva, K.J.D.E.; Ribeiro, V.Q. Path analysis of iron and zinc contents and others traits in cowpea. Crop Breed. Appl. Biotechnol. 2012, 12, 245–252. [Google Scholar] [CrossRef]
- Nalawade, A.D.; Patil, S.M.; Rajwade, P.R.; Kauthale, V.K. Evaluation of cowpea germplasm by using agro-morphological characters. Indian J. Agric. Res. 2020, 55, 364–368. [Google Scholar] [CrossRef]
- Rife, T.; Poland, J. Field Book: An open-source application for field data collection on android. Crop Sci. 2014, 54, 1624–1627. [Google Scholar] [CrossRef]
- IPGRI. Cowpea Descriptors; International Board for Plant Genetic Resources, FAO: Rome, Italy, 1983. [Google Scholar]
- Akinyele, I.O.; Onigbinde, A.O.; Hussain, M.A.; Omololu, A. Physicochemical characteristics of 18 cultivars of Nigerian cowpeas (V. uniguiculafa) and their cooking properties. J. Food Sci. 1986, 51, 1483–1485. [Google Scholar] [CrossRef]
- Kinyanjui, P.; Njoroge, D.; Makokha, A.; Christiaens, S.; Sila, D.; Hendrickxm, M. Quantifying the effects of postharvest storage and soaking pretreatments on the cooking quality of common beans (Phaseolus vulgaris). J. Food Process. Preserv. 2017, 41, e13036. [Google Scholar] [CrossRef]
- Vindiola, O.L.; Seib, O.A.; Hoseney, R.C. Accelerated development of hard-to-cook state in beans. Cereal Foods World 1986, 31, 538–552. [Google Scholar]
- Munthali, J.; Nkhata, S.G.; Masamba, K.; Mguntha, T.; Fungo, R.; Chirwa, R. Soaking beans for 12 h reduces split percent and cooking time regardless of type of water used for cooking. Heliyon 2022, 8, e10561. [Google Scholar] [CrossRef]
- Gama, A.P.; Hung, Y.-C.; Adhikari, K. Optimization of Emulsifier and Stabilizer Concentrations in a Model Peanut-Based Beverage System: A Mixture Design Approach. Foods 2019, 8, 116. [Google Scholar] [CrossRef]
- Sahrawat, K.; Kumar, G.; Murthy, K. Sulfuric acid-selenium digestion for multi-element analysis in a single digest. Commun. Soil Sci. Plant Anal. 2002, 33, 3757–3765. [Google Scholar] [CrossRef]
- Okalebo, J.R.; Gathua, K.W. Laboratory methods of soil and plant analysis. A Work. Man. 2002, 2, 29–68. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 2 January 2023).
- Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. In R Package Version; R Foundation for Statistical Computing: Vienna, Austria, 2010; Volume 1, pp. 1–8. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions; R Package Version 2.1.6; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Boukar, O.; Muranaka, S.; Franco, J.; Fatokun, C.C. Protein and mineral composition in grains of elite cowpea lines. In Proceedings of the Fifth World Cowpea Conference—Improving Livelihoods in the Cowpea Value Chain through Advancement in Science, 26 September–1 October 2010, Saly, Senegal; Boukar, O., Coulibaly, O., Fatokun, C.A., Lopez, K., Tamo, M., Eds.; IITA: Ibadan, Nigeria, 2012; pp. 88–100. [Google Scholar]
- Muranaka, S.; Shono, M.; Myoda, T.; Takeuchi, J.; Franco, J.; Nakazawa, Y.; Boukar, O.; Takagi, H. Genetic diversity of physical, nutritional and functional properties of cowpea grain and relationships among the traits. Plant Genet. Resour. 2015, 1, 1–10. [Google Scholar] [CrossRef]
- Ishikawa, H.; Drabo, I.; Joseph, B.B.; Muranaka, S.; Fatokun, C.; Boukar, O. Characteristics of farmers’ selection criteria for cowpea (Vigna unguiculata) varieties differ between north and south regions of Burkina Faso. Exp. Agric. 2020, 56, 94–103. [Google Scholar] [CrossRef]
- Quaye, W.; Adofo, K.; Buckman, S.E.; Frempong, G.; Jongerden, J.; Ruivenkamp, G. A socio-economic assessment of cowpea diversity on the Ghanaian market: Implications for breeding. Int. J. Consum. Stud. 2011, 35, 679–687. [Google Scholar] [CrossRef]
- Kim, D.K.; Son, D.M.; Choi, J.K.; Shin, H.R.; Choi, K.J.; Lee, J.R.; Lee, K.D.; Rim, Y.S. Agronomic characteristics and seed quality of cowpea (Vigna unguiculata L.) Germplasm. Korean J. Crop Sci. 2013, 58, 1–7. [Google Scholar] [CrossRef][Green Version]
- Olapade, A.A.; Okafor, G.I.; Ozumba, A.U.; Olatunji, O. Characterization of common Nigerian cowpea (Vigna unguiculata L. Walp) varieties. J. Food Eng. 2002, 55, 101–105. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Brandt, W.E.; Singh, B.B. Genetic variability for nutritional composition and cooking time of improved cowpea lines. Crop Sci. 1993, 33, 469–472. [Google Scholar] [CrossRef]
- Jean Baptiste, N.T.; Bell, J.M.; Nassourou, A.M.; Njintang, N.Y.; Youmbi, E. Genetic analysis of seed proteins contents in cowpea (Vigna unguiculata L. Walp.). Afr. J. Biotechnol. 2011, 10, 3077–3086. [Google Scholar] [CrossRef]
- Sanni, S.A.; Adebowale, A.-R.; Yusuf, A. Physical, proximate, and cooking properties of selected Nigerian cowpea varieties. Niger. Food J. 2006, 24, 34–41. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source of Variation | DF | Seed Length | Seed Width | HSDWT |
|---|---|---|---|---|
| Rep | 2 | 124.68 *** | 9.49 *** | 543.50 *** |
| Block:Rep | 99 | 4.67 *** | 2.14 *** | 10.90 ** |
| Genotype | 305 | 18.55 *** | 6.31 *** | 38.30 *** |
| Residuals | 4183 | 1.18 | 0.54 | 7.00 |
| Genotype | Seed Length (mm) | Seed Width (mm) | HSDWT (g) | Seed Coat Color | Seed Shape |
|---|---|---|---|---|---|
| Top 10 genotypes | |||||
| MWcp36 | 9.33 ± 1.29 | 6.53 ± 1.06 | 26.13 ± 0.67 | Brown | Rhomboid |
| MWcp03 | 10.33 ± 0.98 | 6.67 ± 1.05 | 23.70 ± 1.56 | Purple | Ovoid |
| MWcp67 | 7.80 ± 0.94 | 5.53 ± 0.52 | 22.57 ± 3.32 | Reddish-brown | Rhomboid |
| MWcp60 | 8.13 ± 1.13 | 6.13 ± 0.74 | 22.23 ± 1.53 | Reddish-brown | Rhomboid |
| UAM14-126-19-2 | 8.60 ± 1.24 | 5.80 ± 0.68 | 21.70 ± 7.01 | White | Rhomboid |
| MWcp45 | 8.67 ± 1.35 | 6.00 ± 1.07 | 21.30 ± 0.70 | Reddish-brown | Rhomboid |
| MWcp24 | 8.87 ± 0.74 | 6.20 ± 0.86 | 21.03 ± 1.24 | Reddish-brown | Rhomboid |
| MWcp53 | 8.73 ± 1.28 | 5.93 ± 0.59 | 21.00 ± 4.75 | Reddish-brown | Ovoid |
| MWcp04 | 8.00 ± 1.77 | 5.27 ± 0.70 | 20.97 ± 2.78 | Reddish-brown | Rhomboid |
| TVu-14004 | 9.80 ± 0.77 | 5.73 ± 0.70 | 20.93 ± 4.16 | White | Kidney |
| Bottom 10 genotypes | |||||
| TVu-3526 | 5.40 ± 0.74 | 4.07 ± 0.70 | 7.87 ± 0.50 | Purple | Ovoid |
| TVu-1177 | 5.67 ± 0.62 | 3.73 ± 0.46 | 7.77 ± 0.40 | Brown | Ovoid |
| TVu-3063 | 6.87 ± 1.06 | 4.33 ± 0.90 | 7.30 ± 0.79 | White | Ovoid |
| TVu-81 | 5.4 ± 0.63 | 3.73 ± 0.46 | 7.23 ± 0.95 | White | Rhomboid |
| TVu-3228 | 6.27 ± 1.10 | 4.73 ± 0.70 | 7.00 ± 0.40 | Purplish-brown | Ovoid |
| TVu-3229 | 5.20 ± 0.68 | 4.07 ± 0.46 | 7.00 ± 1.11 | Purple | Ovoid |
| TVu-972 | 5.80 ± 0.94 | 3.87 ± 0.64 | 6.97 ± 0.32 | Brown | Ovoid |
| TVu-17060 | 5.07 ± 0.80 | 3.87 ± 0.64 | 6.53 ± 0.65 | Purple | Ovoid |
| TVu-3217 | 6.13 ± 0.83 | 4.27 ± 0.59 | 6.13 ± 0.70 | Brown | Rhomboid |
| TVu-10169 | 6.27 ± 1.28 | 3.8 ± 0.56 | 6.00 ± 1.42 | Black | Rhomboid |
| Grand mean | 7.17 | 4.93 | 12.86 | ||
| Pr (>F) | 0.00 | 0.00 | 0.00 | ||
| CV (%) | 15.14 | 14.97 | 20.51 | ||
| Source of Variation | DF | Broth Viscosity | Cooking Time |
|---|---|---|---|
| Genotype | 303 | 1.17 *** | 1274.80 *** |
| Residuals | 304 | 0.45 | 10.70 |
| Entry | Cooking Time (min) | Broth Viscosity (cP) | SDCCOL | SDSHP |
|---|---|---|---|---|
| Top 10 genotypes | ||||
| TVu-15631 | 39.00 ± 1.41 | 4.60 ± 0.28 | Red | Kidney |
| TVu-328 | 45.50 ± 2.12 | 4.20 ± 0.14 | Mottled | Ovoid |
| IT98K-131-2 | 46.50 ± 4.95 | 3.75 ± 1.34 | Brown | Rhomboid |
| TVu-22 | 47.50 ± 3.54 | 3.90 ± 0.14 | Mottled | Kidney |
| TVu-3550 | 48.50 ± 0.71 | 5.00 ± 0.00 | Reddish-brown | Kidney |
| IT00k-126-3 | 58.50 ± 2.12 | 4.60 ± 0.28 | Brown | Ovoid |
| TVu-14004 | 49.50 ± 2.12 | 3.60 ± 1.41 | White | Kidney |
| MWcp40 | 49.50 ± 2.12 | 3.85 ± 1.34 | Purple | Ovoid |
| TVu-13265 | 49.50 ± 2.12 | 3.85 ± 1.34 | White | Kidney |
| TVu-2661 | 49.50 ± 3.54 | 2.75 ± 0.64 | Brown | Ovoid |
| Bottom 10 genotypes | ||||
| TVu-11674 | 137.50 ± 0.71 | 2.95 ± 0.64 | Brown | Rhomboid |
| MWcp31 | 145.00 ± 2.83 | 3.20 ± 0.28 | Brown | Ovoid |
| MWcp17 | 159.50 ± 0.71 | 4.20 ± 0.14 | White | Rhomboid |
| MWcp61 | 160.50 ± 3.54 | 4.60 ± 0.28 | Mottled | Ovoid |
| MWcp64 | 167.50 ± 0.71 | 3.30 ± 0.28 | Brown | Ovoid |
| MWcp54 | 171.00 ± 5.66 | 4.10 ± 0.14 | Mottled | Rhomboid |
| MWcp29 | 171.50 ± 6.36 | 4.00 ± 0.14 | Purple | Ovoid |
| MWcp46 | 180.50 ± 0.71 | 2.40 ± 0.0 | Mottled | Rhomboid |
| MWcp43 | 181.00 ± 10.41 | 3.65 ± 0.21 | White | Rhomboid |
| MWcp07 | 191.00 ± 7.07 | 2.65 ± 0.64 | Mottled | Ovoid |
| Grand mean | 81.52 | 3.62 | ||
| Pr (>F) | 0.00 | 0.00 | ||
| CV (%) | 4.01 | 18.48 | ||
| Source of Variation | DF | Protein | Iron | Zinc |
|---|---|---|---|---|
| Genotype | 305 | 34.42 *** | 13.58 *** | 0.10 *** |
| Residuals | 306 | 1.05 | 0.05 | 0.00 |
| Entry | Protein (%) | Iron (mg/kg) | Zinc (mg/kg) | SDCCOL | SDSHP |
|---|---|---|---|---|---|
| Top 10 genotypes | |||||
| TVu-3243 | 22.92 ± 0.25 | 8.39 ± 0.00 | 0.45 ± 0.00 | Brown | Ovoid |
| MWcp305 | 22.21 ± 0.85 | 6.56 ± 0.00 | 0.34 ± 0.00 | Brown | Ovoid |
| TVu-2706 | 21.32 ± 0.10 | 3.20 ± 0.00 | 0.39 ± 0.00 | Reddish-brown | Kidney |
| TVu-3094 | 21.29 ± 0.15 | 1.29 ± 0.00 | 0.36 ± 0.00 | White | Kidney |
| MWcp69 | 21.21 ± 0.75 | 8.67 ± 0.00 | 0.39 ± 0.00 | Purple | Ovoid |
| MZcp 024 | 20.97 ± 0.60 | 5.00 ± 0.00 | 0.34 ± 0.00 | Brown | Ovoid |
| TVu-3524 | 20.40 ± 0.10 | 1.19 ± 0.00 | 0.29 ± 0.00 | Brown | Rhomboid |
| IT 99K-529-1 | 20.29 ± 0.15 | 7.15 ± 0.00 | 0.45 ± 0.00 | Brown | Ovoid |
| TVu-10169 | 20.18 ± 0.70 | 1.93 ± 0.00 | 0.33 ± 0.00 | Black | Rhomboid |
| TVu-1483 | 20.04 ± 0.20 | 1.64 ± 0.00 | 0.43 ± 0.00 | Purplish- brown | Ovoid |
| Bottom 10 genotypes | |||||
| TVu-1015 | 4.29 ± 0.10 | 1.58 ± 0.00 | 0.27 ± 0.00 | Brown | Ovoid |
| MWcp601 | 4.19 ± 0.03 | 1.89 ± 0.00 | 0.22 ± 0.00 | Brown | Ovoid |
| MWcp48 | 4.17 ± 0.15 | 1.90 ± 0.00 | 1.77 ± 0.02 | Purple | Ovoid |
| MWcp46 | 4.09 ± 0.03 | 2.00 ± 0.00 | 0.23 ± 0.00 | Mottled | Rhomboid |
| IT90K-76 | 4.06 ± 0.10 | 1.57 ± 0.01 | 0.31 ± 0.00 | Brown | Ovoid |
| TVx-3236 | 4.00 ± 0.03 | 2.67 ± 0.00 | 0.27 ± 0.00 | White | Rhomboid |
| TZcp67 | 3.78 ± 0.20 | 1.94 ± 0.00 | 0.18 ± 0.00 | Brown | Ovoid |
| MWcp50 | 3.64 ± 0.10 | 1.67 ± 0.00 | 0.24 ± 0.00 | Purple | Ovoid |
| MZcp 004 | 3.63 ± 0.07 | 1.98 ± 0.00 | 0.17 ± 0.00 | Brown | Ovoid |
| TVu-3263 | 3.60 ± 0.15 | 1.24 ± 0.00 | 0.28 ± 0.00 | Purple | Rhomboid |
| Grand mean | 11.05 | 3.21 | 0.36 | ||
| Pr (>F) | 0.00 | 0.00 | 0.00 | ||
| CV% | 9.27 | 7.10 | 1.23 | ||
| Trait 1 | Trait 2 | r | 95% CI | t | df | p |
|---|---|---|---|---|---|---|
| Protein | Iron | 0.28 | [0.17, 0.38] | 5.05 | 304 | <0.001 *** |
| Protein | Zinc | 0.21 | [0.10, 0.31] | 3.72 | 304 | 0.005 ** |
| Protein | Broth | −0.07 | [−0.18, 0.04] | −1.23 | 302 | >0.999 |
| Protein | CT | −0.03 | [−0.14, 0.08] | −0.53 | 302 | >0.999 |
| Protein | Seed.L | −0.06 | [−0.17, 0.05] | −1.09 | 304 | >0.999 |
| Protein | Seed.W | −0.03 | [−0.15, 0.08] | −0.59 | 304 | >0.999 |
| Protein | HSDWT | −0.06 | [−0.17, 0.05] | −1.08 | 304 | >0.999 |
| Iron | Zinc | 0.24 | [0.13, 0.34] | 4.34 | 304 | <0.001 *** |
| Iron | Broth | −0.04 | [−0.15, 0.08] | −0.61 | 302 | >0.999 |
| Iron | CT | −0.08 | [−0.19, 0.03] | −1.45 | 302 | >0.999 |
| Iron | Seed.L | −7.24 × 10−03 | [−0.12, 0.10] | −0.13 | 304 | >0.999 |
| Iron | Seed.W | 0.03 | [−0.09, 0.14] | 0.47 | 304 | >0.999 |
| Iron | HSDWT | −0.06 | [−0.17, 0.05] | −1.08 | 304 | >0.999 |
| Zinc | Broth | 0.03 | [−0.08, 0.14] | 0.5 | 302 | >0.999 |
| Zinc | CT | −0.04 | [−0.15, 0.07] | −0.75 | 302 | >0.999 |
| Zinc | Seed.L | 0.07 | [−0.04, 0.18] | 1.29 | 304 | >0.999 |
| Zinc | Seed.W | 0.06 | [−0.05, 0.17] | 1.06 | 304 | >0.999 |
| Zinc | HSDWT | 0.03 | [−0.09, 0.14] | 0.48 | 304 | >0.999 |
| Broth | CT | −0.04 | [−0.16, 0.07] | −0.76 | 302 | >0.999 |
| Broth | Seed.L | 6.74 × 10−03 | [−0.11, 0.12] | 0.12 | 302 | >0.999 |
| Broth | Seed.W | −0.03 | [−0.14, 0.08] | −0.54 | 302 | >0.999 |
| Broth | HSDWT | −0.09 | [−0.20, 0.02] | −1.61 | 302 | >0.999 |
| CT | Seed.L | 0.21 | [0.10, 0.31] | 3.72 | 302 | 0.005 ** |
| CT | Seed.W | 0.26 | [0.15, 0.36] | 4.67 | 302 | <0.001 *** |
| CT | HSDWT | 0.19 | [0.08, 0.30] | 3.44 | 302 | 0.013 * |
| Seed.L | Seed.W | 0.89 | [0.86, 0.91] | 33.49 | 304 | <0.001 *** |
| Seed.L | HSDWT | 0.71 | [0.65, 0.76] | 17.57 | 304 | <0.001 *** |
| Seed.W | HSDWT | 0.75 | [0.70, 0.80] | 19.88 | 304 | <0.001 *** |
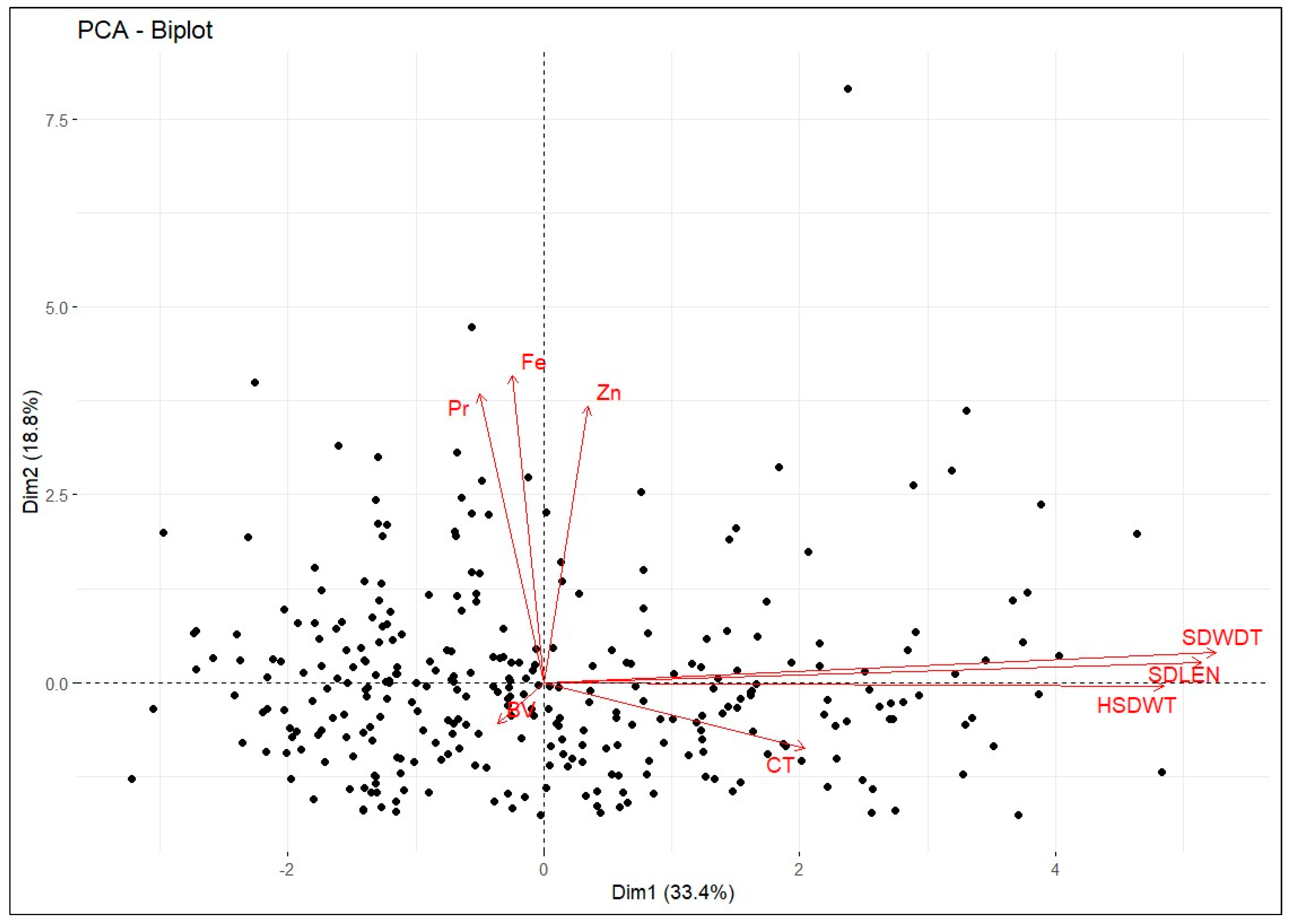
| Traits | Principal Components | Trait Contributions (%) | ||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| Seed length | 0.57 | 0.04 | 0.11 | 32.13 | 0.16 | 1.30 |
| Seed width | 0.58 | 0.06 | 0.05 | 33.59 | 0.34 | 0.26 |
| 100 seed weight | 0.53 | −0.00 | −0.02 | 28.51 | 0.00 | 0.05 |
| Cooking time | 0.22 | −0.13 | −0.29 | 5.08 | 1.62 | 8.65 |
| Broth viscosity | −0.04 | −0.08 | 0.89 | 0.16 | 0.65 | 79.84 |
| Protein | −0.06 | 0.56 | −0.20 | 0.31 | 31.92 | 3.84 |
| Iron | −0.03 | 0.60 | 0.01 | 0.08 | 36.04 | 0.01 |
| Zinc | 0.04 | 0.54 | 0.25 | 0.14 | 36.04 | 6.05 |
| Eigenvalue | 2.67 | 1.51 | 1.04 | |||
| % of variance | 33.36 | 18.81 | 12.98 | |||
| Cumulative % of variance | 33.37 | 52.19 | 65.17 | |||
| Cluster | N | Statistic | Pr (%) | Fe (mg/kg) | Zn (mg/kg) | CT (min) | BV (cP) | Seed.L (mm) | Seed.W (mm) | HSDWT (g) |
|---|---|---|---|---|---|---|---|---|---|---|
| I | 234 | M | 10.95 | 3.26 | 0.35 | 76.98 | 3.68 | 6.74 | 4.66 | 11.43 |
| Min | 3.60 | 0.32 | 0.08 | 39.00 | 1.60 | 3.73 | 2.93 | 6.00 | ||
| Max | 22.92 | 21.44 | 1.23 | 145.00 | 5.50 | 8.93 | 5.73 | 18.80 | ||
| II | 72 | M | 11.39 | 3.06 | 0.40 | 96.03 | 3.43 | 8.57 | 5.79 | 17.58 |
| Min | 3.64 | 0.12 | 0.09 | 49.50 | 2.20 | 5.67 | 4.53 | 12.80 | ||
| Max | 21.21 | 14.64 | 2.63 | 191.00 | 4.90 | 10.33 | 7.13 | 26.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chipeta, M.M.; Yohane, E.; Kafwambira, J.; Kampanje-Phiri, J. Genetic Variability and Interrelationships of Grain, Cooking, and Nutritional Quality Traits in Cowpea: Implications for Cowpea Improvement. Agriculture 2024, 14, 633. https://doi.org/10.3390/agriculture14040633
Chipeta MM, Yohane E, Kafwambira J, Kampanje-Phiri J. Genetic Variability and Interrelationships of Grain, Cooking, and Nutritional Quality Traits in Cowpea: Implications for Cowpea Improvement. Agriculture. 2024; 14(4):633. https://doi.org/10.3390/agriculture14040633
Chicago/Turabian StyleChipeta, Michael M., Esnart Yohane, John Kafwambira, and Jessica Kampanje-Phiri. 2024. "Genetic Variability and Interrelationships of Grain, Cooking, and Nutritional Quality Traits in Cowpea: Implications for Cowpea Improvement" Agriculture 14, no. 4: 633. https://doi.org/10.3390/agriculture14040633
APA StyleChipeta, M. M., Yohane, E., Kafwambira, J., & Kampanje-Phiri, J. (2024). Genetic Variability and Interrelationships of Grain, Cooking, and Nutritional Quality Traits in Cowpea: Implications for Cowpea Improvement. Agriculture, 14(4), 633. https://doi.org/10.3390/agriculture14040633