
Effects and Underlying Mechanisms of Rice-Paddy-Upland Rotation Combined with Bacterial Fertilizer for the Abatement of Radix pseudostellariae Continuous Cropping Obstacles
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
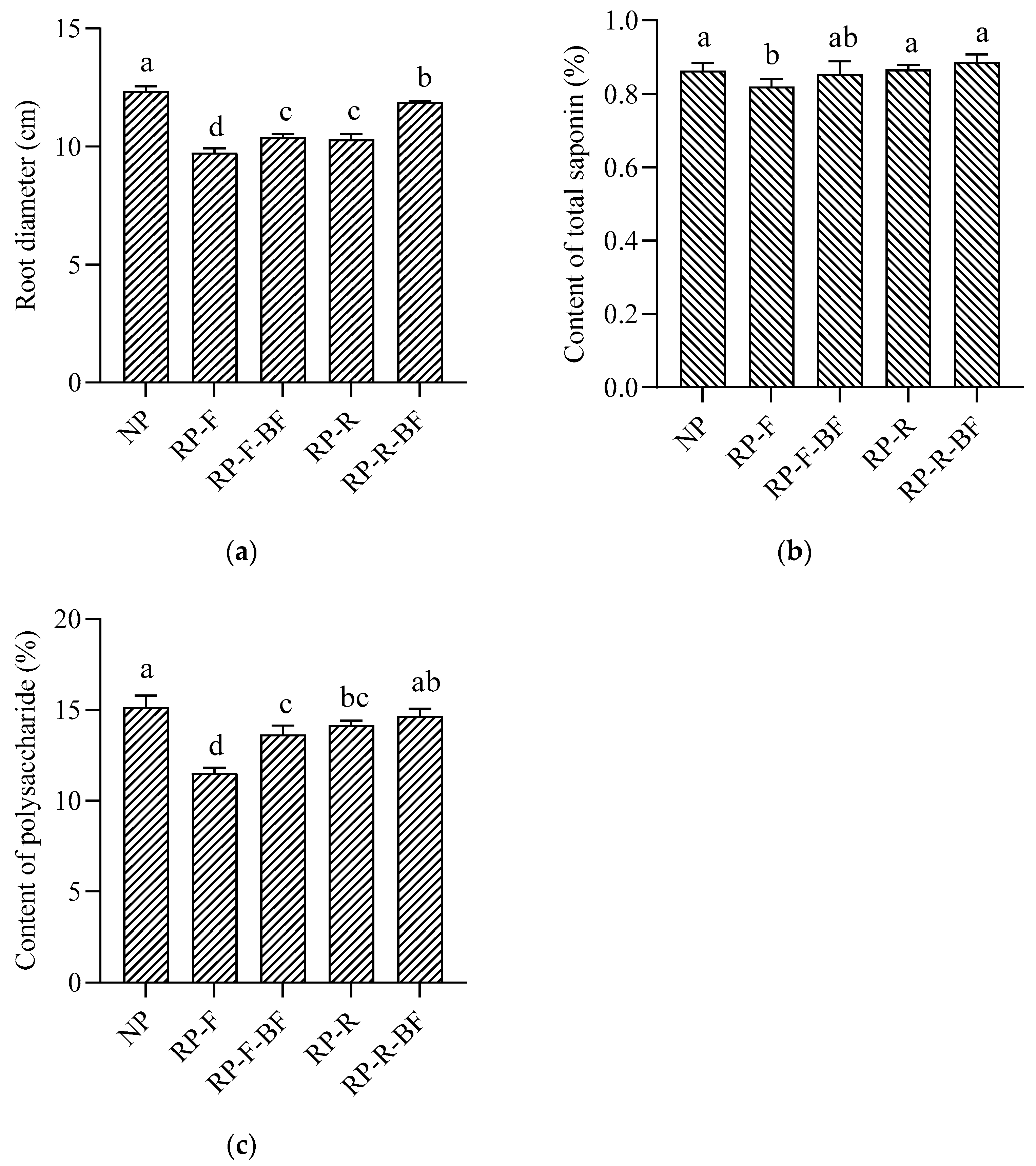
2.2. Yield Measurement and Quality Detection
2.3. Determination of Soil Physicochemical Properties and Enzyme Activity
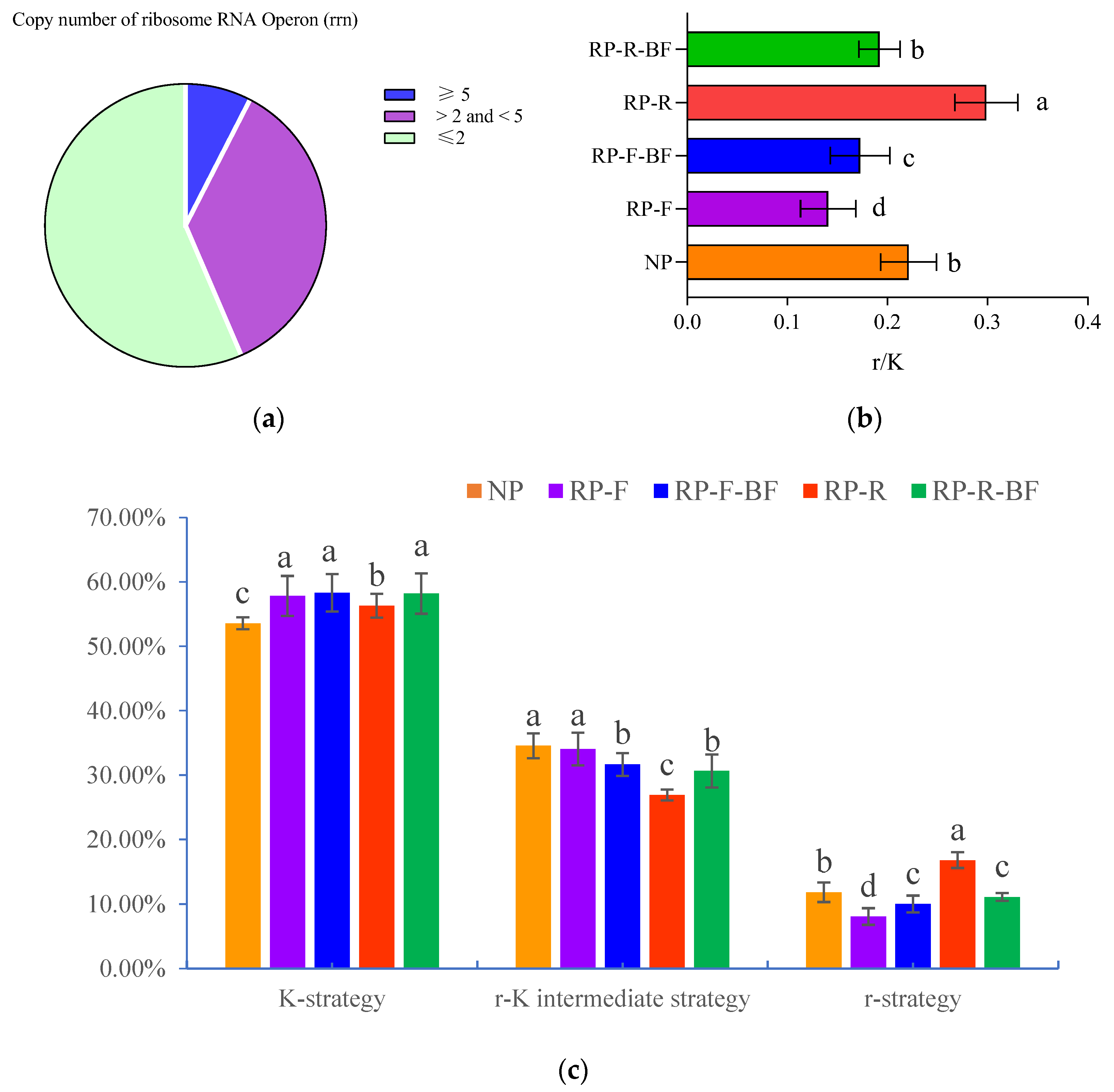
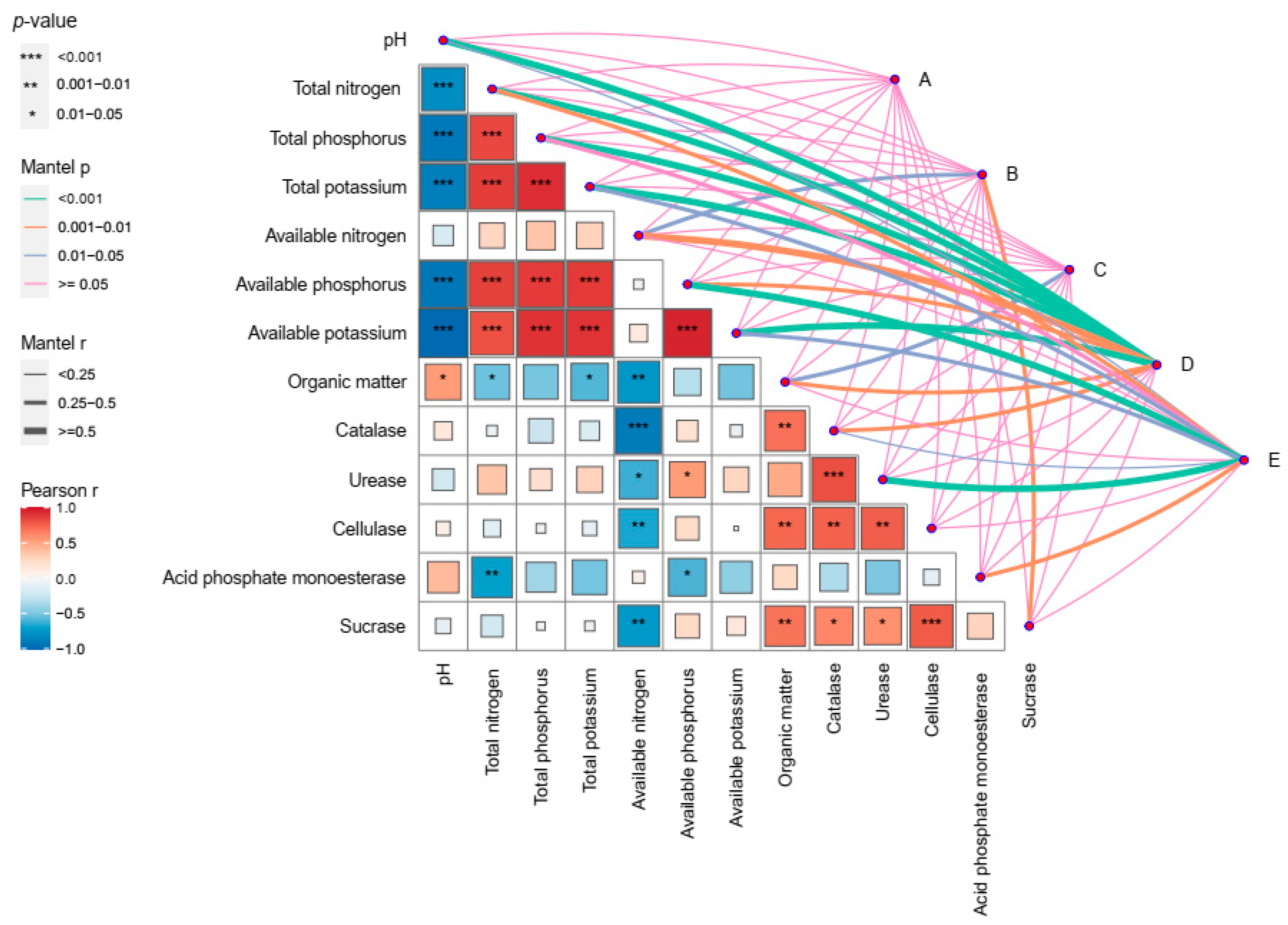
2.4. High-Throughput Sequencing Analysis of Rhizosphere Soil Microorganisms
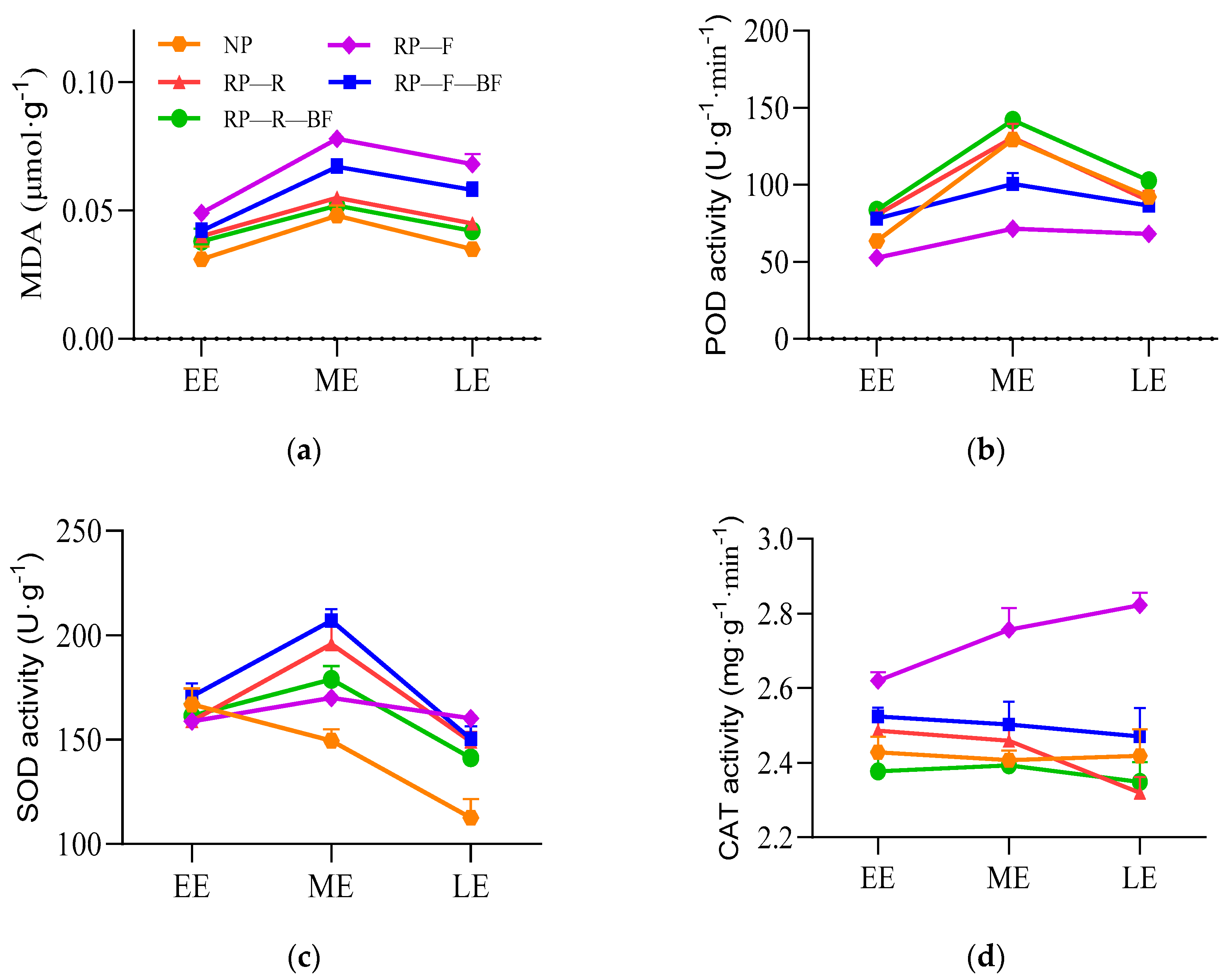
2.5. Determination of Protective Enzyme Activity and Gene Expression Related to Disease Resistance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chao1 | Observed_Species | Shannon | Simpson | ||
|---|---|---|---|---|---|
| Fungi | NP | 1067.65 ± 256.77 a | 824 ± 205.79 a | 5.28 ± 0.92 ab | 0.88 ± 0.08 ab |
| RP-F | 670.16 ± 272.04 b | 475.67 ± 169.47 b | 4.05 ± 0.92 c | 0.81 ± 0.11 b | |
| RP-F-BF | 917.54 ± 84.79 ab | 597 ± 35.34 ab | 5.03 ± 0.24 abc | 0.93 ± 0.01 a | |
| RP-R | 1048.44 ± 141.76 a | 760.97 ± 107.96 a | 5.91 ± 0.42 a | 0.96 ± 0.01 a | |
| RP-R-BF | 810.22 ± 165.94 ab | 514.9 ± 85.17 b | 4.37 ± 0.45 bc | 0.87 ± 0.04 ab | |
| Bacteria | NP | 3664.64 ± 95.05 ab | 2762.2 ± 21.64 a | 9.27 ± 0.01 a | 0.9947 ± 0.0005 a |
| RP-F | 3249.78 ± 24.59 c | 2381.93 ± 58.01 c | 9.01 ± 0.10 c | 0.9944 ± 0.0004 a | |
| RP-F-BF | 3793.65 ± 200.31 a | 2754.3333 ± 58.2789 a | 9.21 ± 0.05 ab | 0.995 ± 0.0002 a | |
| RP-R | 3475.52 ± 57.11 b | 2491.59 ± 68.73 b | 8.80 ± 0.12 d | 0.9902 ± 0.0015 b | |
| RP-R-BF | 3628.55 ± 126.86 ab | 2588.5 ± 68.61 b | 9.10 ± 0.09 bc | 0.9946 ± 0.0003 a |
2.6. Data Analysis
3. Results
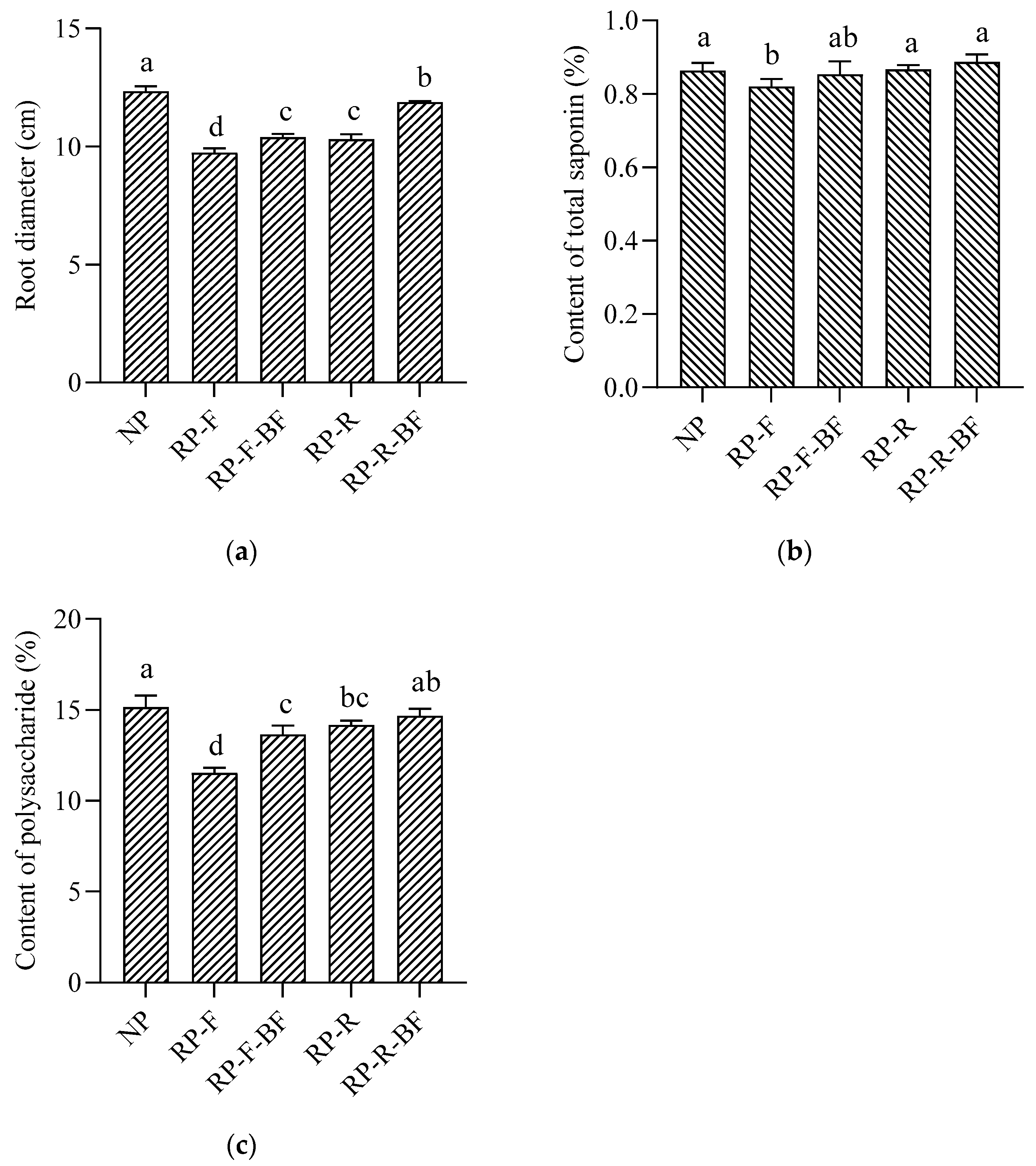
3.1. Yield and Quality of R. pseudostellariae under Different Treatments
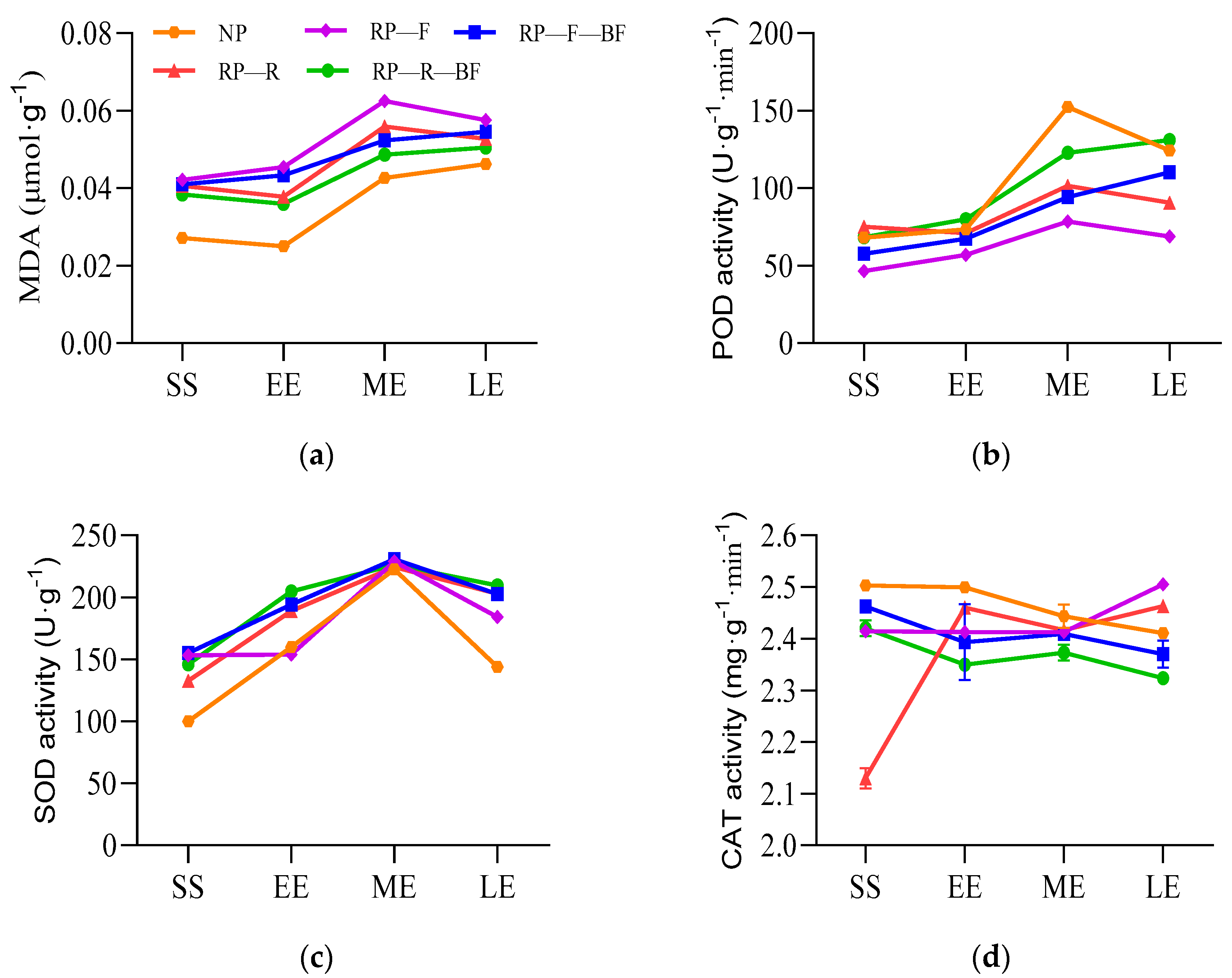
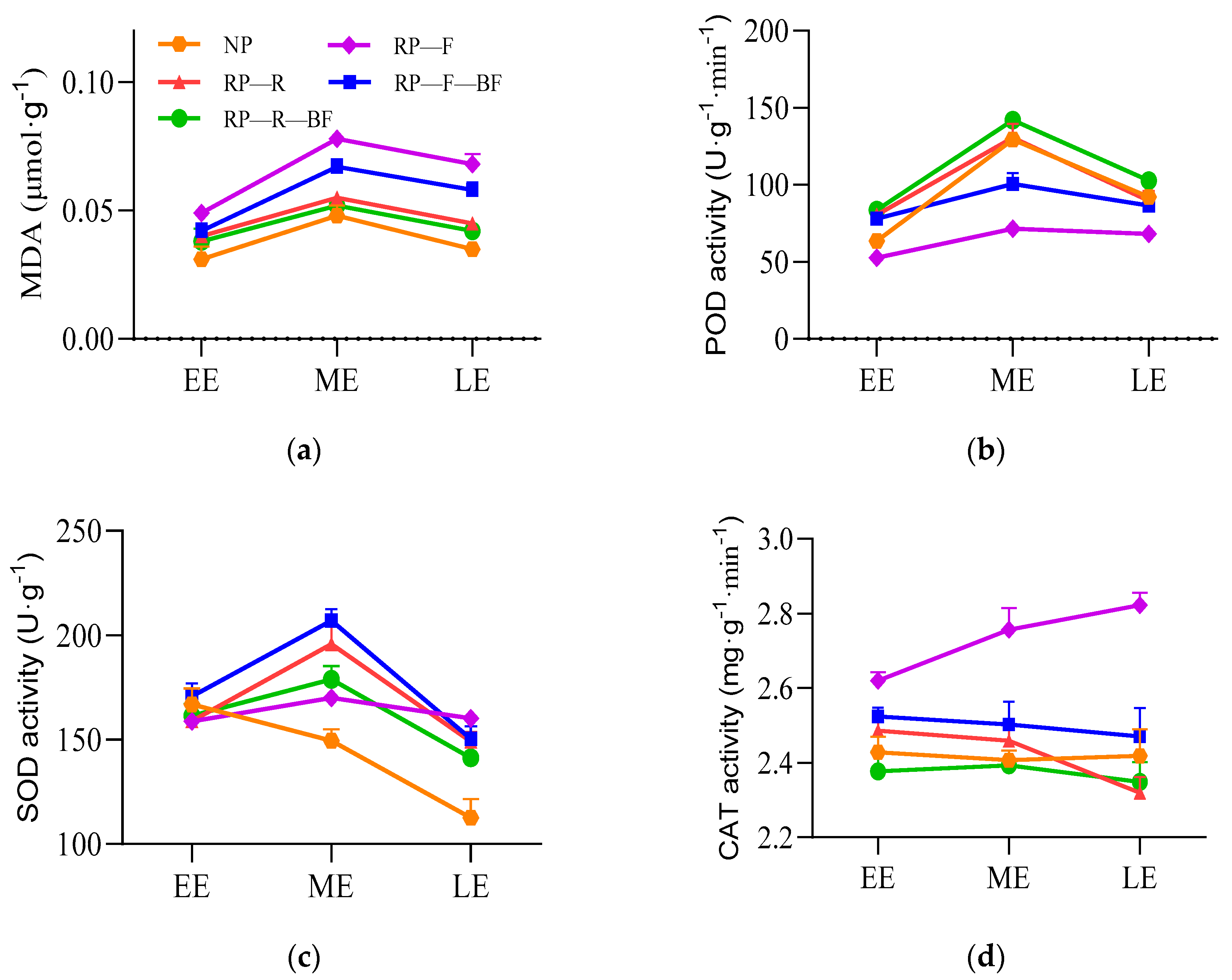
3.2. Differential Physiological Responses of Leaves and Root Tubers under Different Treatments
3.3. Differential Expression of Disease Resistance Genes in Leaves and Tuberous Roots
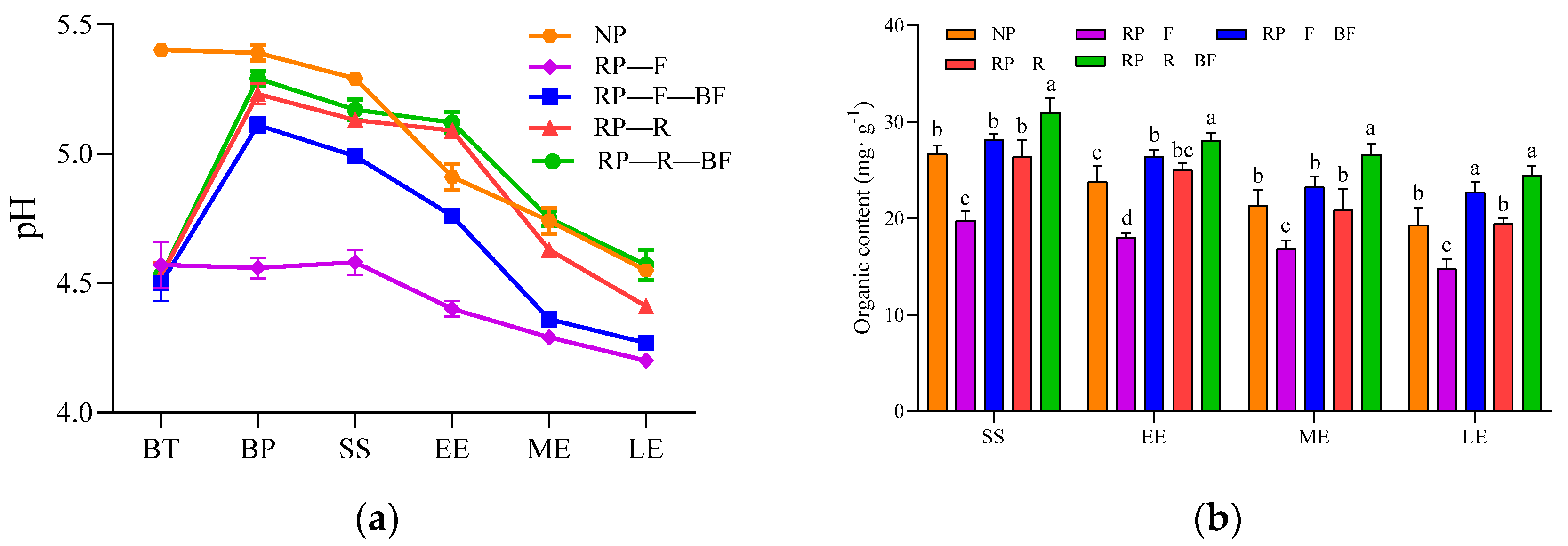
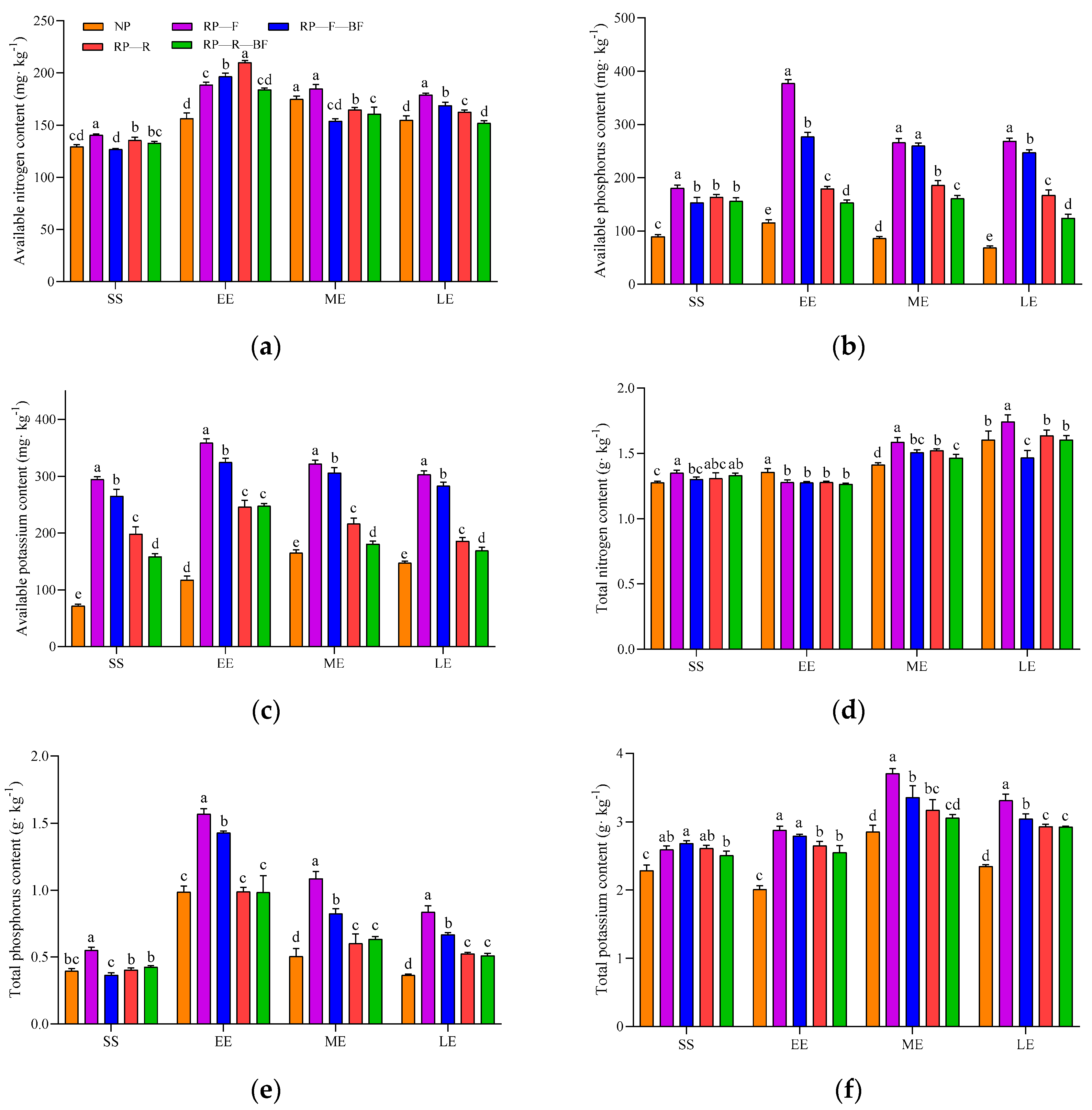
3.4. Differences in Soil Physicochemical Properties under Different Treatments
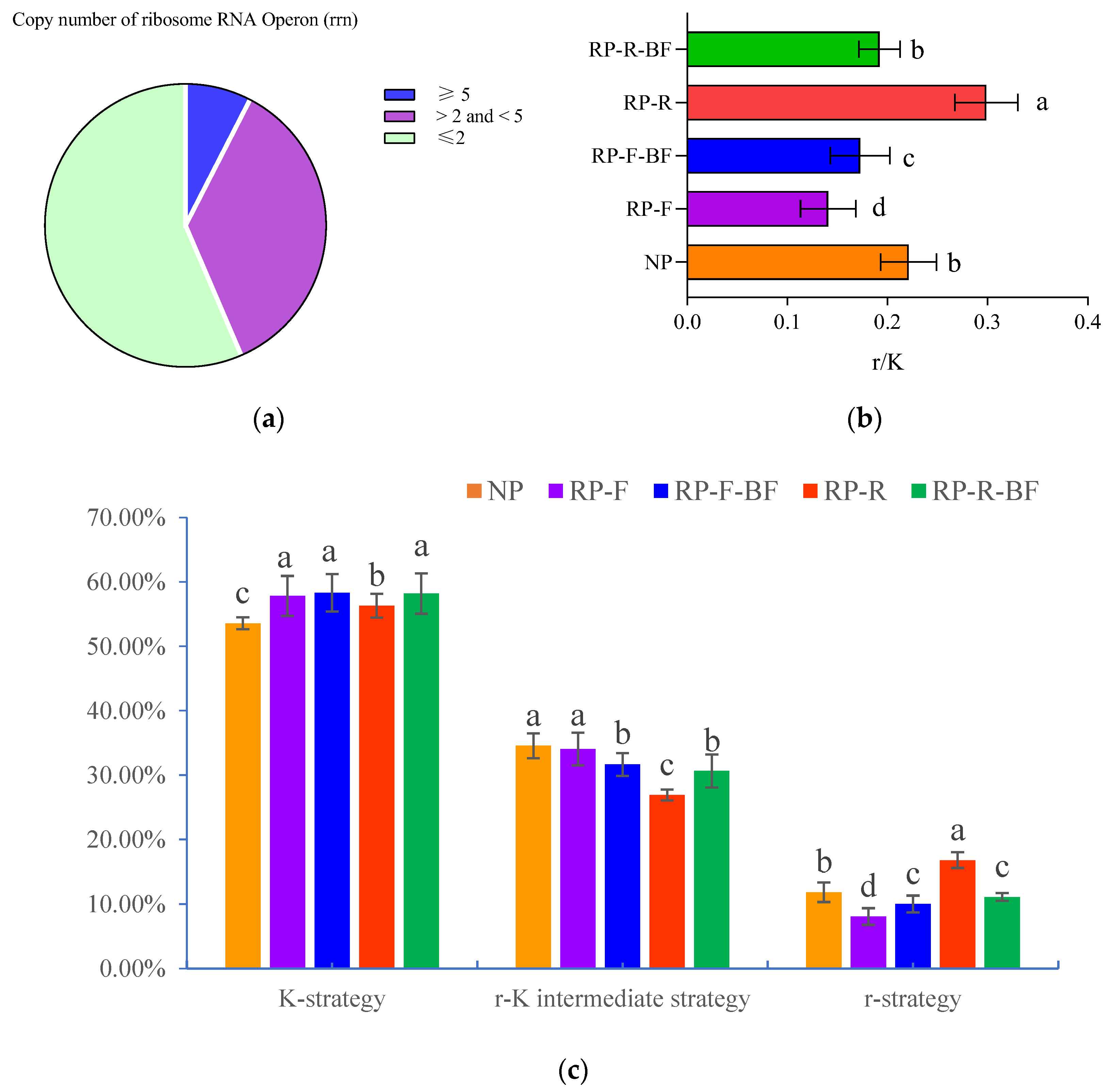
3.5. Differences in Microbial Community Structure in the Rhizosphere Soil of R. pseudostellariae under Different Treatments
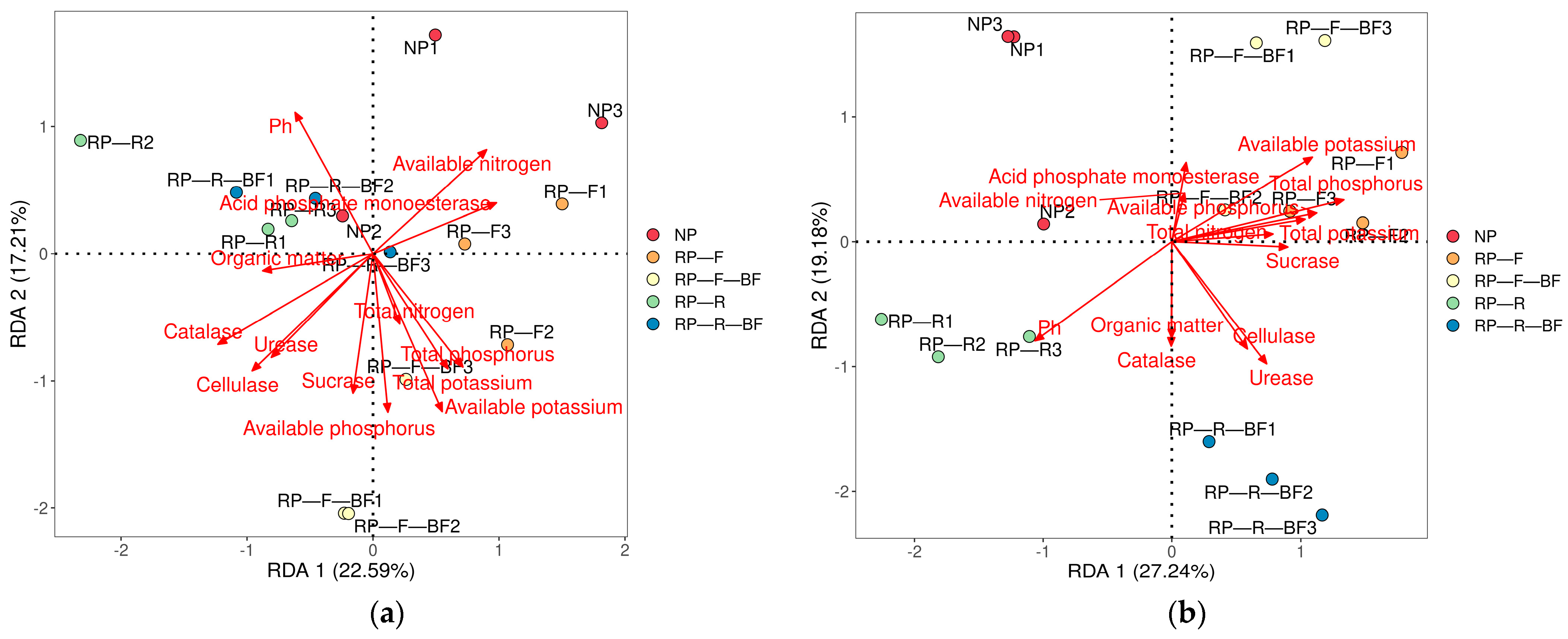
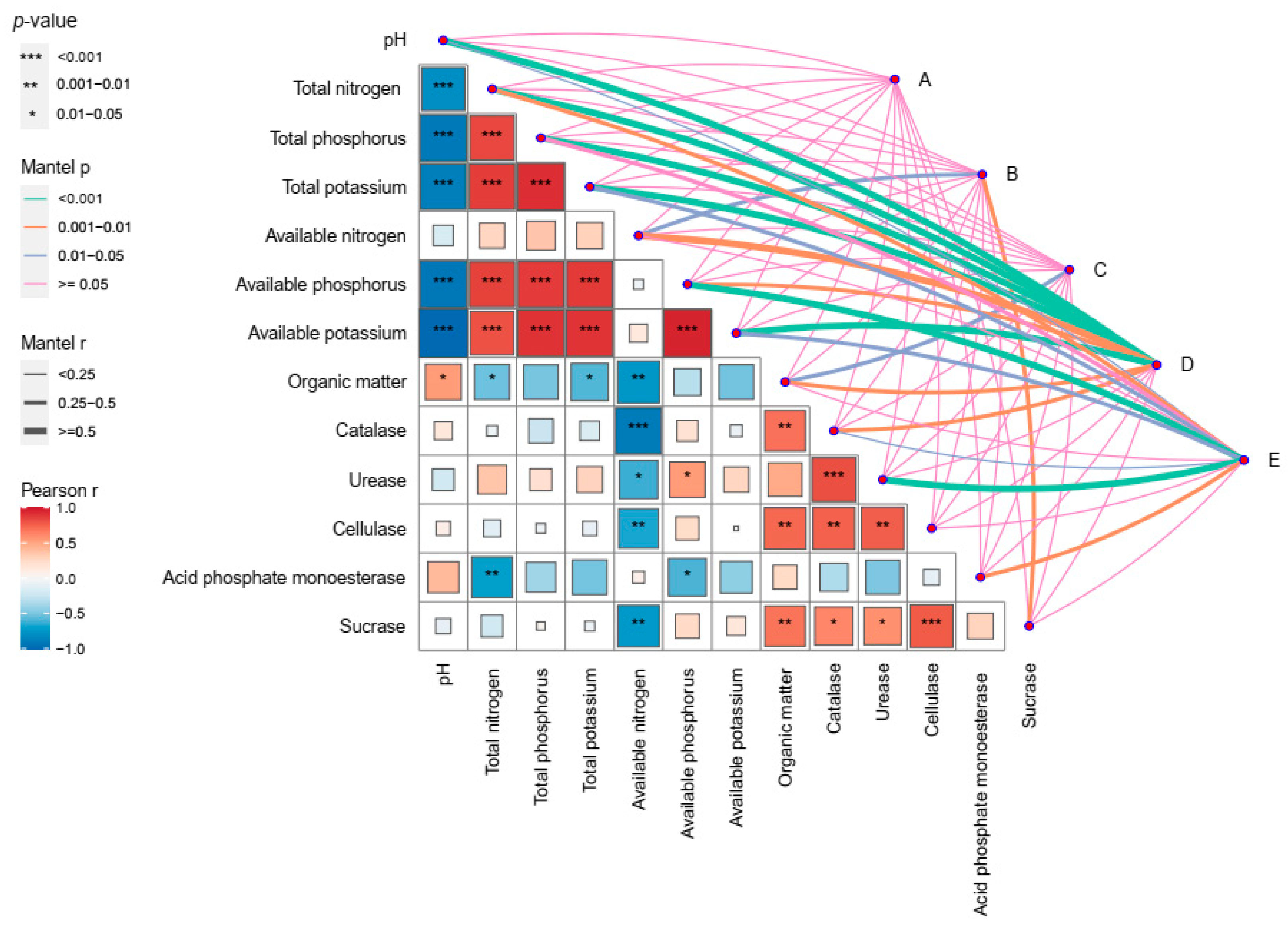
3.6. Correlation Analysis between Rhizosphere Microbial Community and Environmental Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Eisenstein, M. Natural solutions for agricultural productivity. Nature 2020, 588, 58–59. [Google Scholar] [CrossRef]
- Crippa, M.; Solazzo, E.; Guizzardi, D.; Monforti-Ferrario, F.; Tubiello, F.-N.; Leip, A. Food systems are responsible for a third of global anthropogenic GHG emissions. Nat. Food. 2021, 2, 198–209. [Google Scholar] [CrossRef]
- Lin, W.-X.; Chen, T. Transition of agricultural systems to ecologicalizaton and new vision of modern eco-agriculture development in China. Chinese J. Eco-Agri. 2019, 27, 169–176. [Google Scholar]
- Huang, L.-F.; Song, L.-X.; Xia, X.J.; Mao, W.-H.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q. Plant-soil feedbacks and soil sickness: From mechanisms to application in agriculture. J. Chem. Ecol. 2013, 39, 232–242. [Google Scholar] [CrossRef]
- Wang, J.-H.; Chen, T.; Lin, W.-X. Plant allelopathy types and their application in agriculture. Chinese J. Eco-Agri. 2013, 21, 1173–1183. [Google Scholar] [CrossRef]
- Chen, T.; Lin, S.; Wu, L.-K.; Lin, W.-X.; Sampietro, D.-A. Soil Sickness: Current Status and Future Perspectives. Allelopathy J. 2015, 36, 167–195. [Google Scholar]
- Guo, L.-P.; Huang, L.-Q.; Jiang, Y.-X.; Lyu, D.-M. Soil deterioration during cultivation of medicinal plants and ways to prevent it. China J. Chin. Mater. Medica 2006, 31, 714–717. [Google Scholar]
- Gao, W.-W.; Zhao, Y.-J.; Wang, Y.-P.; Chen, S.-L. Review of research on sustainable use of medicinal plants cropland in China. China J. Chin. Mater. Medica 2006, 31, 1665–1669. [Google Scholar]
- Lin, M.-Z.; Zhang, Z.-X.; Lin, Z.-C.; Long, C.-H.; Zeng, L.-J.; Lin, W.-X. Analysis of differential expression of proteins in replanting disease of Pseudostellaria heterophylla. Acta Prataculturae Sin. 2010, 19, 197–207. [Google Scholar]
- Xia, P.-H.; Liu, Y. Study on Obstacle Effect of Succession Cropping on Pseudostellaria heterophylla. Acta Bot. Boreali-Occident. Sin. 2010, 30, 2240–2246. [Google Scholar]
- Zhang, F.-S.; Shen, J.-B.; Feng, G. Rhizosphere Ecology: Processes & Management; China Agricultural University Press: Beijing, China, 2009. [Google Scholar]
- Wu, H.-M.; Qin, X.-J.; Wang, J.-Y.; Wu, L.-K.; Chen, J.; Fan, J.-K.; Zheng, L.; Tangtai, H.-P.; Arafat, Y.; Lin, W.-W.; et al. Rhizosphere chemical language in the Radix pseudostellariae rhizosphere under continuous monoculture regimes. Agr. Ecosyst. Environ. 2019, 270, 19–31. [Google Scholar] [CrossRef]
- van der Putten, W.-H.; Bardgett, R.-D.; Bever, J.-D.; Bezemer, T.-M.; Casper, B.-B.; Fukami, T.; Kardol, P.; Klironomos, J.-N.; Kulmatiski, A.; Schweitzer, J.-A.; et al. Plant-soil feedback: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.-E.; Tringe, S.-G.; Sa, T.-M.; Singh, B.-K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Zhao, M.; Zhao, J.; Yuan, J.; Hale, L.; Tao, W.; Huang, Q.-W.; Vivanco, J.-M.; Zhou, J.-Z.; Kowalchuk, G.-A.; Shen, Q.-R. Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant Cell Environ. 2021, 44, 613–628. [Google Scholar] [CrossRef]
- Tittonell, P. Ecological intensification of agriculture—Sustainable by nature. Curr. Opin. Env. Sust. 2014, 8, 53–61. [Google Scholar] [CrossRef]
- Kleijn, D.; Bommarco, R.; Fijen, T.-P.-M.; Garibaldi, L.-A.; Potts, S.-G.; van der Putten, W.-H. Ecological Intensification: Bridging the Gap between Science and Practice. Trends Ecol. Evol. 2019, 34, 154–166. [Google Scholar] [CrossRef]
- Jing, J.-Y.; Cong, W.-F.; Bezemer, T.-M. Legacies at work: Plant–soil–microbiome interactions underpinning agricultural sustainability. Trends Plant Sci. 2022, 8, 781–792. [Google Scholar] [CrossRef]
- Li, Y.-Z.; Zhang, J.-Z.; Jia, J.-Y.; Fan, F.; Zhang, F.-S.; Zhang, J.-L. Research progresses on farmland soil ecosystem multifunctionality. Acta Pedol. Sin. 2022, 59, 1177–1189. [Google Scholar]
- Mao, L.-L.; Zhang, L.-Z.; ZHANG, S.-P.; Evers, J.-B.; van der Werf, W.; Wang, J.-J.; Sun, H.-Q.; Su, Z.-C.; Spiertz, H. Resource use efficiency, ecological intensification and sustainability of intercropping systems. J. Integr. Agr. 2015, 14, 1542–1550. [Google Scholar] [CrossRef]
- Wu, Q.-M.; Zhang, J.-M.; Li, Y.-Y.; Zhang, Y. Recent advances on the mechanism of beneficial microbial fertilizers in crops. Biotech. Bulletin. 2021, 37, 221–230. [Google Scholar]
- Bergstrand, K.-J. Organic fertilizers in greenhouse production systems-a review. Sci. Hortic. 2022, 295, 110855. [Google Scholar] [CrossRef]
- Cao, A.-C.; Fang, W.-S.; Li, Y.; Yan, D.-D.; Wang, Q.-X.; Guo, M.-X.; Huang, B.; Song, Z.-X.; Jin, X. Review on 60 years of soil fumigation and disinfestation in China. J. Plant Protect. 2022, 49, 325–335. [Google Scholar]
- Yadav, S.-P.-S.; Bhandari, S.; Bhatta, D.; Poudel, A.; Bhattarai, S.; Yadav, P.; Ghimire, N.; Paudel, P.; Paudel, P.; Shrestha, J.; et al. Biochar application: A sustainable approach to improve soil health. J. Agr. Food Res. 2023, 11, 100498. [Google Scholar]
- Dore, T.; Makowski, D.; Malezieux, E.; Munier, N.; Tchamitchian, M.; Tittonell, P. Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. Eur. J. Agron. 2011, 34, 197–210. [Google Scholar] [CrossRef]
- Li, L. Intercropping enhances agroecosystem services and functioning: Current knowledge and perspectives. Chinese J. Eco.-Agri. 2016, 24, 403–415. [Google Scholar]
- Li, Q.J.; Zhang, D.Q.; Song, Z.X.; Ren, L.R.; Jin, X.; Fang, W.-S.; Yan, D.-D.; Li, Y.; Wang, Q.-X.; Cao, A.-C. Organic fertilizer activates soil beneficial microorganisms to promote strawberry growth and soil health after fumigation. Environ. Pollut. 2022, 295, 118653. [Google Scholar] [CrossRef]
- Pu, R.-F.; Wang, P.-P.; Guo, L.-P.; Li, M.-H.; Cui, X.-M.; Wang, C.-X.; Liu, Y.; Yang, Y. The remediation effects of microbial organic fertilizer on soil microorganisms after chloropicrin fumigation. Ecotox. Environ. Safe. 2022, 231, 113188. [Google Scholar] [CrossRef]
- Bai, S.-H.; Omidvar, N.; Gallart, M.; Kämper, W.; Tahmasbian, I.; Farrar, M.-B.; Singh, K.; Zhou, G.-Y.; Muqadass, B.; Xu, C.-Y.; et al. Combined effects of biochar and fertilizer applications on yield: A review and meta-analysis. Sci. Total Environ. 2022, 808, 152073. [Google Scholar] [CrossRef]
- Reyes-Cabrera, J.; Leon, R.-G.; Erickson, J.-E.; Rowland, D.-L.; Silveira, M.-L.; Morgan, K.-T. Differences in biomass and water dynamics between a cotton-peanut rotation and a sweet sorghum bioenergy crop with and without biochar and vinasse as soil amendments. Field Crop Res. 2017, 214, 123–130. [Google Scholar] [CrossRef]
- Hu, Y.-J.; Sun, B.-H.; Wu, S.-F.; Feng, H.; Gao, M.-X.; Zhang, B.-B.; Liu, Y.-Y. After-effects of straw and straw-derived biochar application on crop growth, yield, and soil properties in wheat (Triticum aestivum L.)-maize (Zea mays L.) rotations: A four-year field experiment. Sci. Total Environ. 2021, 780, 146560. [Google Scholar] [CrossRef]
- Wang, Y.-G.; Huang, D.-S. Efficient cultivation technique of Radix pseudostellariae and rice in rotation. Agric. Technol. Commun. 2009, 12, 171–172. [Google Scholar]
- Yang, Y. Techniques of rice rotation cultivation in Radix pseudostellariae. Mod. Rural Sci. Technol. 2017, 4, 15. [Google Scholar]
- Wei, X.-Y. Insight into the Mechanism of Panax notoginseng under Continuous Monoculture and Evaluation of Microial Fertilizer Remediation; Fujian Agriculture and Forestry University: Fuzhou, China, 2018. [Google Scholar]
- Xu, Q.; Wang, H.-F.; Zhou, X.-Y. Optimization of extraction technology of saponins from Radix pseudostellariae. Chin. Tradit. Herb. Drugs 2001, 32, 34–35. [Google Scholar]
- Liu, D.; Zhang, C.-H.; Chen, X.; Ling, Q.-Y.; Han, B.-X.; Chen, N.-F. Optimization of extraction process and contrastive study of polysoccharides from Radix Pseudostellarial. J. Anhui Agri. Sci. 2014, 42, 12469–12471. [Google Scholar]
- Pansu, M.; Gautheyrou, J. Handbook of Soil Analysis: Mineralogical, Organic and Inorganic Methods; Springer Science & Business Media: Berlin, Germany, 2007. [Google Scholar]
- Guan, S.-Y. Soil Enzyme and Study Method; Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Wu, H.-M.; Lin, M.-H.; Rensing, C.; Qin, X.-J.; Zhang, S.-K.; Chen, J.; Wu, L.-K.; Zhao, Y.-L.; Lin, S.; Lin, W.-X. Plant-mediated rhizospheric interactions in intraspecific intercropping alleviate the replanting disease of Radix pseudostellariae. Plant Soil 2020, 454, 411–430. [Google Scholar] [CrossRef]
- Edgar, R.-C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Wang, X.-K. Principles and Techniques of Plant Physiology and Biochemistry Experiment; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Qin, X.-J.; Wu, H.-M.; Chen, J.; Wu, L.-K.; Lin, S.; Khan, M.-U.; Boorboori, M.-R.; Lin, W.-X. Transcriptome analysis of Pseudostellaria heterophylla in response to the infection of pathogenic Fusarium oxysporum. BMC Plant Biol. 2017, 17, 155. [Google Scholar] [CrossRef]
- Stoddard, S.-F.; Smith, B.-J.; Hein, R.; Roller, B.-R.-K.; Schmidt, T.-M. rrnDB: Improved tools for interpreting rRNA gene abundance in bacteria and archaea and a new foundation for future development. Nucleic Acids Res. 2015, 43, 593–598. [Google Scholar] [CrossRef]
- Nemergut, D.-R.; Knelman, J.-E.; Ferrenberg, S.; Bilinski, T.; Melbourne, B.; Lin, J.; Violle, C.; Darcy, J.-L.; Prest, T.; Schmidt, S.-K. Decreases in average bacterial community rRNA operon copy number during succession. Isme J. 2016, 10, 1147–1156. [Google Scholar] [CrossRef]
- Chen, L.; Jiang, Y.; Liang, C.; Luo, Y.; Xu, Q.; Han, C.; Zhao, Q.; Sun, B. Competitive interaction with keystone taxa induced negative priming under biochar amendments. Microbiome 2019, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.-A.; Hilton, A. Statistical Analysis in Microbiology: Statnotes; Wiley: Hoboken, NJ, USA, 2010. [Google Scholar]
- Shou, N.-S.; Huang, D.-W.; Wu, Y.; Chen, R.-F.; Zhang, L.-Q.; Wu, X.-Y. Effects of Silicon on Microbial Community Structure of Rhizosphere Soil of Rice at Different Nitrogen Levels. Chin. J. Soil Sci. 2021, 52, 903–911. [Google Scholar]
- Etesami, H.; Schaller, J. Improving phosphorus availability to rice through silicon management in paddy soils: A review of the role of silicate-solubilizing bacteria. Rhizosphere 2023, 27, 1007. [Google Scholar] [CrossRef]
- Kang, C.-Z.; Lyu, C.-G.; Huang, L.-Q.; Wang, S.; Wang, H.-Y.; Zhang, W.-J.; Wang, R.-S.; Wang, T.-L.; Sun, J.-H.; Zhou, T.; et al. Pattern of ecological planting for Chinese materia medica based on regional distribution. China J. Chin. Mater. Medica 2020, 45, 1982–1989. [Google Scholar]
- Yuan, Y.-D.; Zuo, J.-J.; Zhang, H.-Y.; Zu, M.-T.; Liu, S.-A. The Chinese medicinal plants rhizosphere: Metabolites, microorganisms, and interaction. Rhizosphere 2022, 22, 100540. [Google Scholar] [CrossRef]
- Perez Brandan, C.; Arzeno, J.L.; Huidobro, D.J.; Conforto, E.C.; Grumberg, B.C.; Hilton, S.; Vargas Gil, S. The effect of crop sequences on soil microbial, chemical and physical indicators and its relationship with soybean sudden death syndrome (complex of Fusarium species). Spanish J. Agri. Res. 2014, 12, 252–264. [Google Scholar] [CrossRef]
- Pérez-Brandán, C.; Huidobro, J.; Grümberg, B.; Scandiani, M.M.; Luque, A.G.; Meriles, J.M.; Vargas-Gil, S. Soybean fungal soil-borne diseases: A parameter for measuring the effect of agricultural intensification on soil health. Canadian J. Microbiol. 2014, 60, 73–84. [Google Scholar] [CrossRef]
- Dong, Y.-F.; Lyu, X.-Z.; Zhang, Z.-K.; He, H.-J.; Yu, J.-Q.; Zhou, Y.-H. Effects of different cultivation patterns on soil microbial community and enzyme activity in continuous cropped pepper field. Acta Agri. Zhejiangensis 2019, 31, 1485–1492. [Google Scholar]
- Meena, V.-S.; Meena, S.-K.; Verma, J.-P.; Kumar, A.; Aeron, A.; Mishra, P.-K.; Bisht, J.-K.; Pattanayak, A.; Naveed, M.; Dotaniya, M.-L. Plant beneficial rhizospheric microorganism (PBRM) strategies to improve nutrients use efficiency: A review. Ecol. Eng. 2017, 107, 8–32. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.-L.; Yadav, A.-N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.-S.; Saxena, A.-K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatal. Agri. Biotech. 2020, 23, 101487. [Google Scholar] [CrossRef]
- Shahwar, D.; Mushtaq, Z.; Mushtaq, H.; Alqarawi, A.-A.; Park, Y.; Alshahrani, T.-S.; Faizan, S. Role of microbial inoculants as bio fertilizers for improving crop productivity: A review. Heliyon 2023, 9, e16134. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.-F.; Zhu, Y.-J.; Wang, Z.-R.; Zhang, H.-F.; Chen, M.-C.; Chen, Y.-P.; Wang, J.-P.; Liu, B. Effects of a novel bio-organic fertilizer on the composition of rhizobacterial communities and bacterial wilt outbreak in a continuously mono-cropped tomato field. Appl. Soil Ecol. 2020, 156, 103717. [Google Scholar] [CrossRef]
- Liu, J.; Li, H.; Yuan, Z.-Y.; Feng, J.-J.; Chen, S.-H.; Sun, G.-Z.; Wei, Z.-H.; Hu, T.-T. Effects of microbial fertilizer and irrigation amount on growth, physiology and water use efficiency of tomato in greenhouse. Sci. Horticul. 2024, 323, 112553. [Google Scholar] [CrossRef]
- Ali, A.; Ghani, M.-I.; Ding, H.-Y.; Iqbal, M.; Cheng, Z.-H.; Cai, Z.-C. Arbuscular mycorrhizal inoculum coupled with organic substrate induces synergistic effects for soil quality changes, and rhizosphere microbiome structure in long-term monocropped cucumber planted soil. Rhizosphere 2021, 20, 100428. [Google Scholar] [CrossRef]
- Zhao, Y.-N.; Mao, X.-X.; Zhang, M.-S.; Yang, W.; Di, H.-J.; Ma, L.; Liu, W.-J.; Li, B.-W. The application of Bacillus Megaterium alters soil microbial community composition, bioavailability of soil phosphorus and potassium, and cucumber growth in the plastic shed system of North China. Agr. Ecosyst. Environ. 2021, 307, 107236. [Google Scholar] [CrossRef]
- Ling, N.; Deng, K.-Y.; Song, Y.; Wu, Y.-C.; Zhao, J.; Raza, W.; Huang, Q.-W.; Shen, Q.-R. Variation of rhizosphere bacterial community in watermelon continuous mono-cropping soil by long-term application of a novel bioorganic fertilizer. Microbiol. Res. 2014, 169, 570–578. [Google Scholar] [CrossRef]
- Ding, S.; Zhou, D.-P.; Wei, H.-W.; Wu, S.-H.; Xie, B. Alleviating soil degradation caused by watermelon continuous cropping obstacle: Application of urban waste compost. Chemosphere 2021, 262, 128387. [Google Scholar] [CrossRef]
- Zhang, D.-Q.; Cheng, H.-Y.; Hao, B.-Q.; Li, Q.-J.; Fang, W.-S.; Ren, L.-R.; Yan, D.-D.; Ouyang, C.-B.; Li, Y.; Wang, Q.-X.; et al. Effect of fresh chicken manure as a non-chemical soil fumigant on soil-borne pathogens, plant growth and strawberry fruit profitability. Crop Prot. 2021, 146, 105653. [Google Scholar] [CrossRef]
- He, Z.-K. Chitinolyticbacter Meiyuanensis Isolation, Identification, and Fermentation for Chintinase Production; Jiangnan University: Wuxi, China, 2011. [Google Scholar]
- Luo, Y. Effect of Low Temperature Stress and Cold Hardening on Physiology Character of Strawberry and Molecular Cloning of Class II Chitinase Gene; Sichuan Agricultural University: Chengdu, China, 2007. [Google Scholar]




| Fungi | Bacteria | |||||
|---|---|---|---|---|---|---|
| Environmental Factor | R2 | p Value | R2 | p Valure | ||
| pH | 0.4989 | 0.016 | * | 0.544 | 0.014 | * |
| Total nitrogen | 0.1079 | 0.499 | 0.2161 | 0.233 | ||
| Total phosphorus | 0.4013 | 0.052 | 0.1832 | 0.279 | ||
| Total potassium | 0.3612 | 0.073 | 0.128 | 0.426 | ||
| Available nitrogen | 0.4544 | 0.032 | * | 0.0496 | 0.73 | |
| Available phosphorus | 0.4847 | 0.023 | * | 0.3328 | 0.096 | |
| Available potassium | 0.569 | 0.007 | ** | 0.4612 | 0.021 | * |
| Organic matter | 0.2398 | 0.202 | 0.508 | 0.019 | * | |
| Catalase | 0.6204 | 0.004 | ** | 0.5835 | 0.007 | ** |
| Urease | 0.4039 | 0.033 | * | 0.4054 | 0.05 | * |
| Cellulase | 0.5428 | 0.007 | ** | 0.339 | 0.084 | |
| Acid phosphate monoesterase | 0.3427 | 0.079 | 0.1918 | 0.278 | ||
| Sucrase | 0.3805 | 0.062 | 0.2511 | 0.167 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.; Yang, Y.; Chen, T.; Jiao, Y.; Yang, J.; Cai, Z.; Lin, W. Effects and Underlying Mechanisms of Rice-Paddy-Upland Rotation Combined with Bacterial Fertilizer for the Abatement of Radix pseudostellariae Continuous Cropping Obstacles. Agriculture 2024, 14, 326. https://doi.org/10.3390/agriculture14020326
Lin S, Yang Y, Chen T, Jiao Y, Yang J, Cai Z, Lin W. Effects and Underlying Mechanisms of Rice-Paddy-Upland Rotation Combined with Bacterial Fertilizer for the Abatement of Radix pseudostellariae Continuous Cropping Obstacles. Agriculture. 2024; 14(2):326. https://doi.org/10.3390/agriculture14020326
Chicago/Turabian StyleLin, Sheng, Yuanyuan Yang, Ting Chen, Yanyang Jiao, Juan Yang, Zhaoying Cai, and Wenxiong Lin. 2024. "Effects and Underlying Mechanisms of Rice-Paddy-Upland Rotation Combined with Bacterial Fertilizer for the Abatement of Radix pseudostellariae Continuous Cropping Obstacles" Agriculture 14, no. 2: 326. https://doi.org/10.3390/agriculture14020326
APA StyleLin, S., Yang, Y., Chen, T., Jiao, Y., Yang, J., Cai, Z., & Lin, W. (2024). Effects and Underlying Mechanisms of Rice-Paddy-Upland Rotation Combined with Bacterial Fertilizer for the Abatement of Radix pseudostellariae Continuous Cropping Obstacles. Agriculture, 14(2), 326. https://doi.org/10.3390/agriculture14020326