
The Main Compounds of Bio-Fumigant Plants and Their Role in Controlling the Root-Knot Nematode Meloidogyne incognita (Kofoid and White) Chitwood




 ,
,  ,
, 
 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Bio-Fumigant Plants
2.2. Identification of Chemical Compounds
2.3. In Vitro Studies
2.4. Chemical Compound Studies
2.5. Parameter Measurement
2.6. Data Analysis
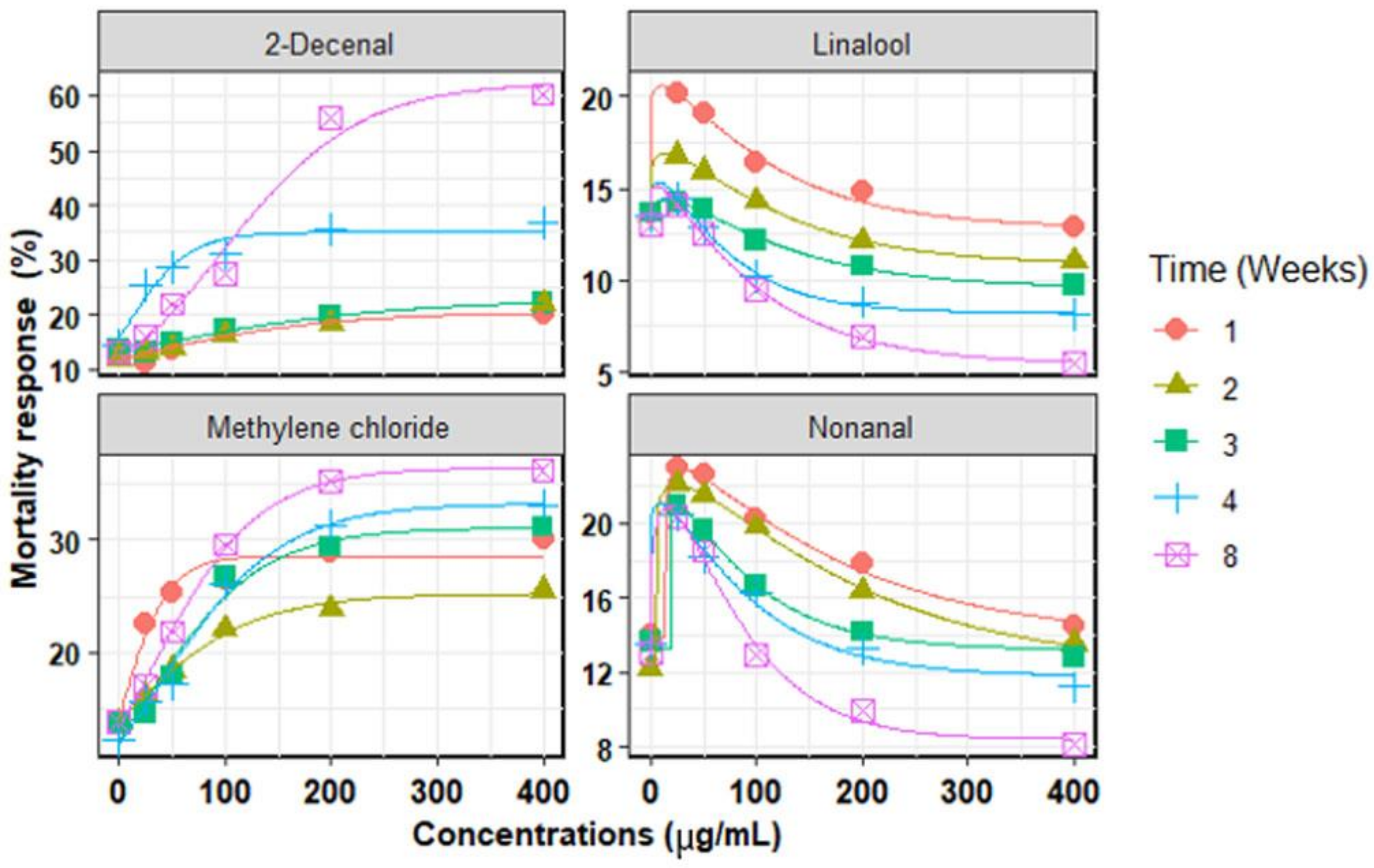
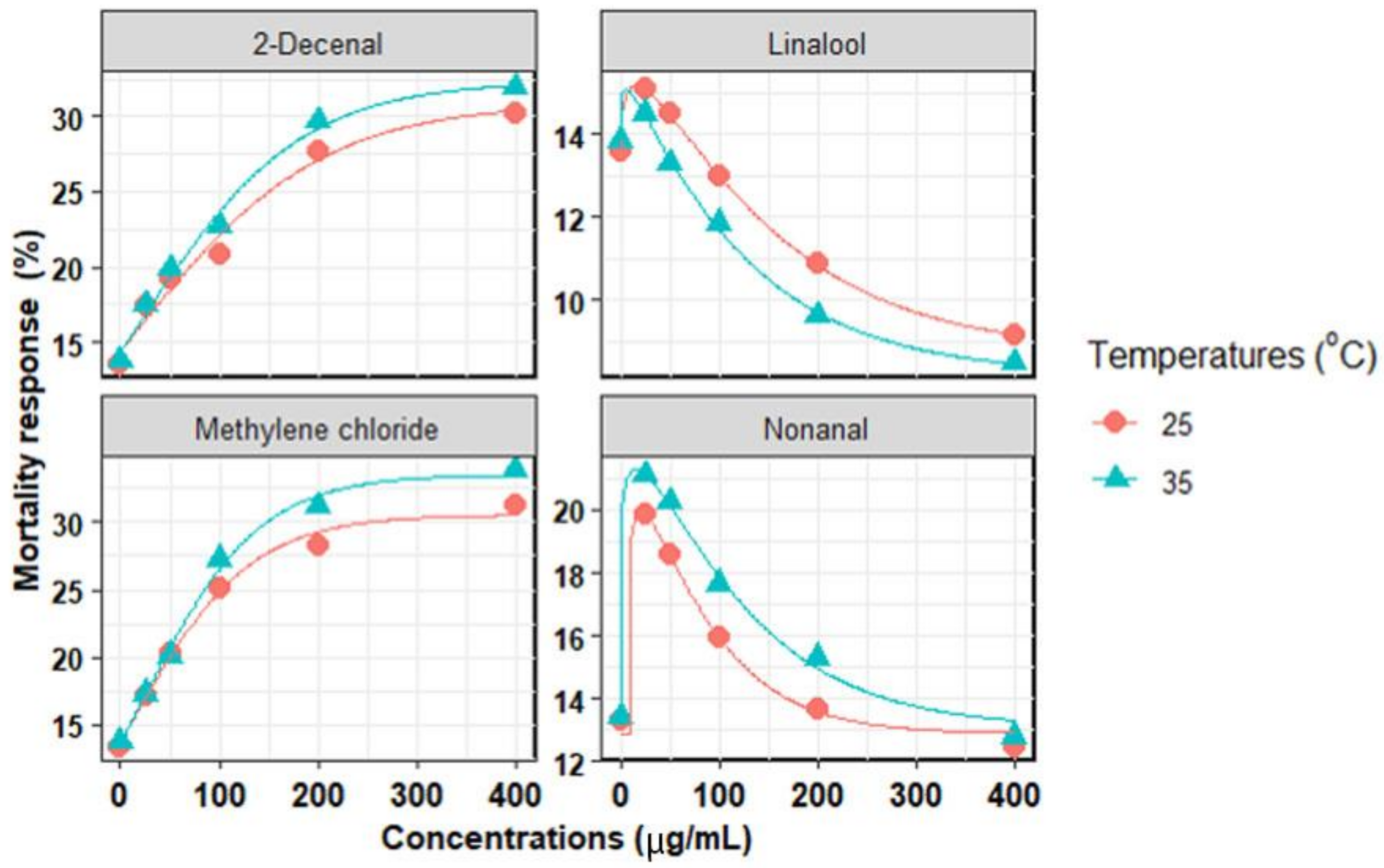
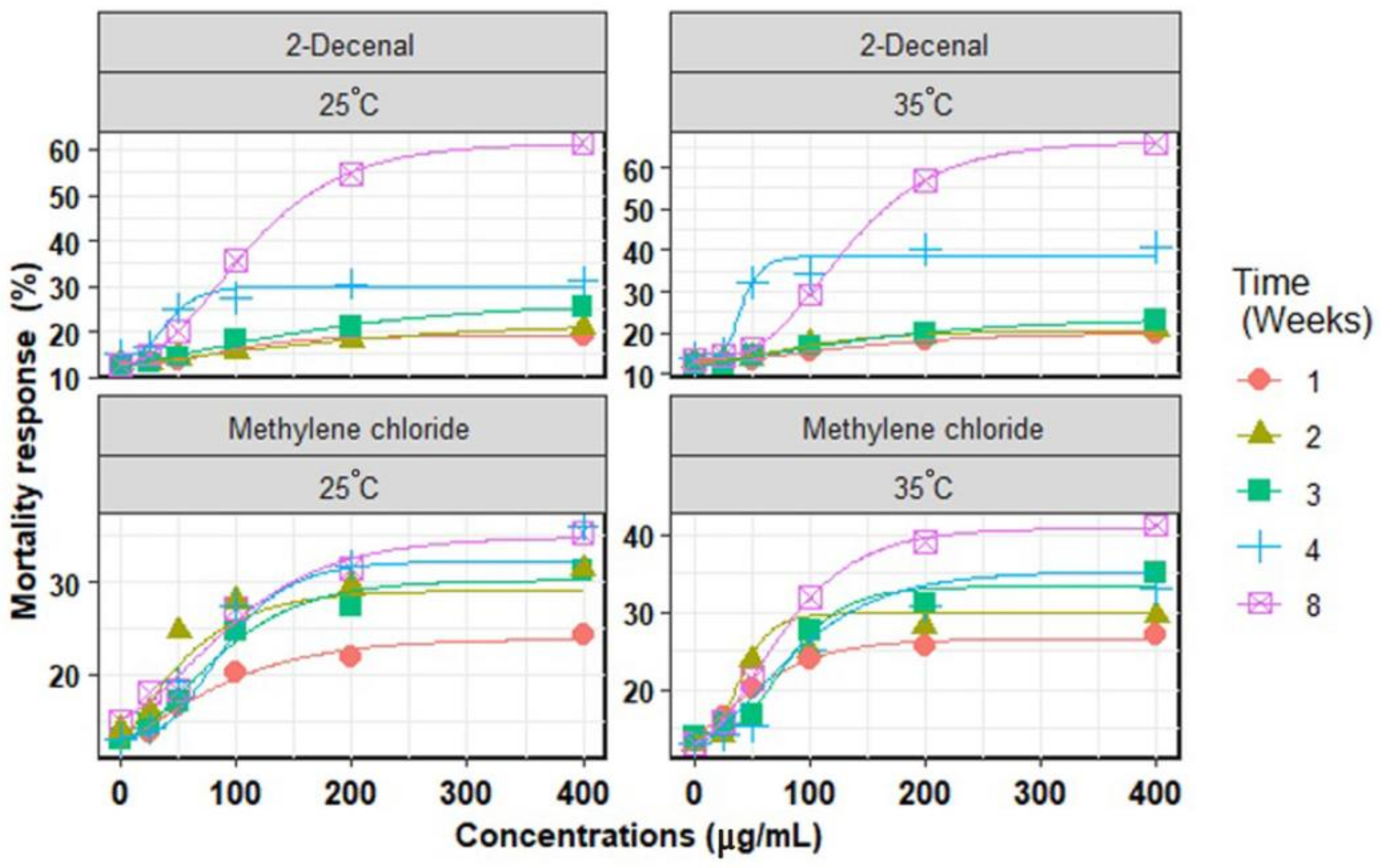
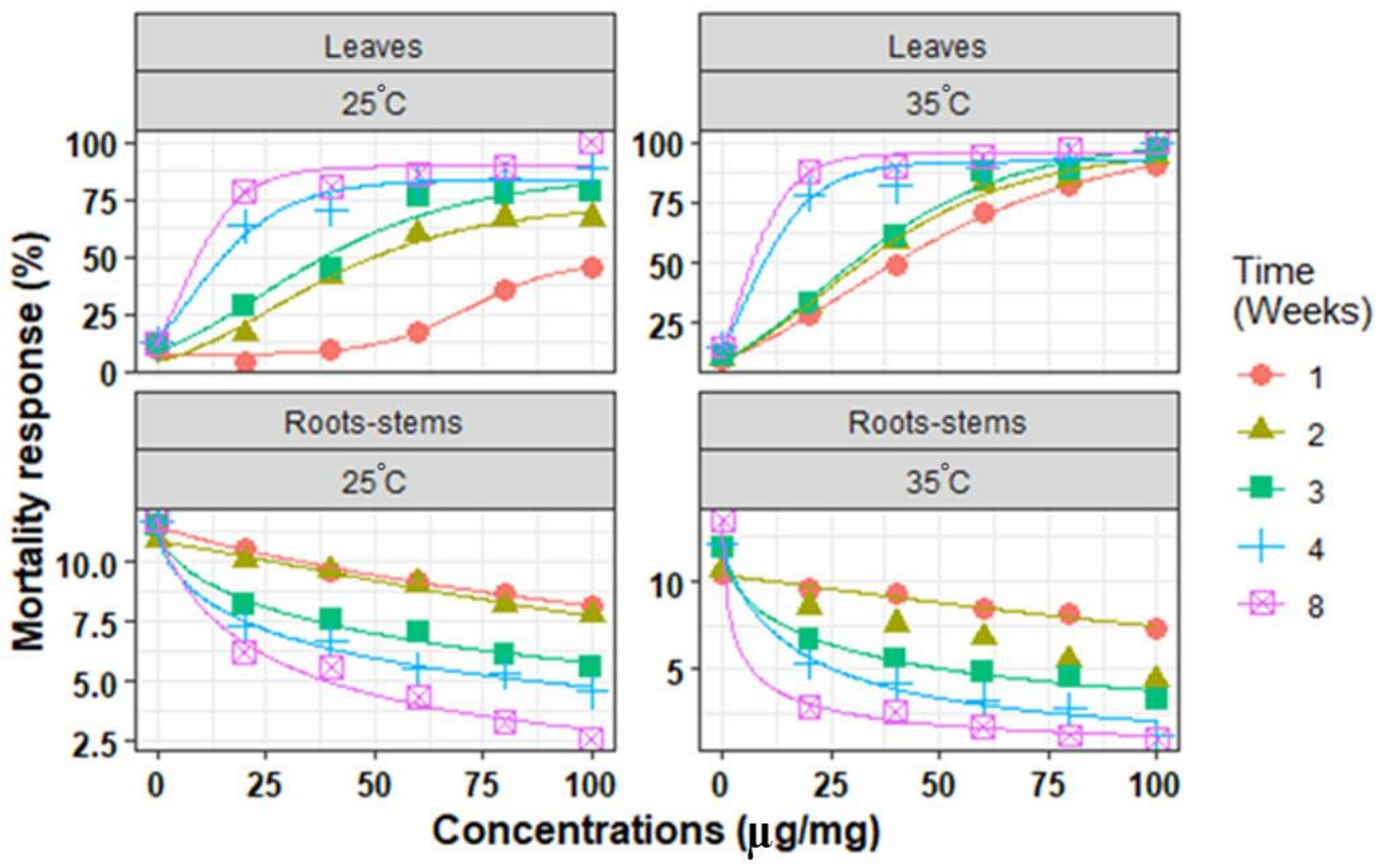
3. Results
3.1. Chemical Compounds
3.2. In Vitro Mortality of Second-Stage Juveniles
3.3. Chemical Compounds Extracted
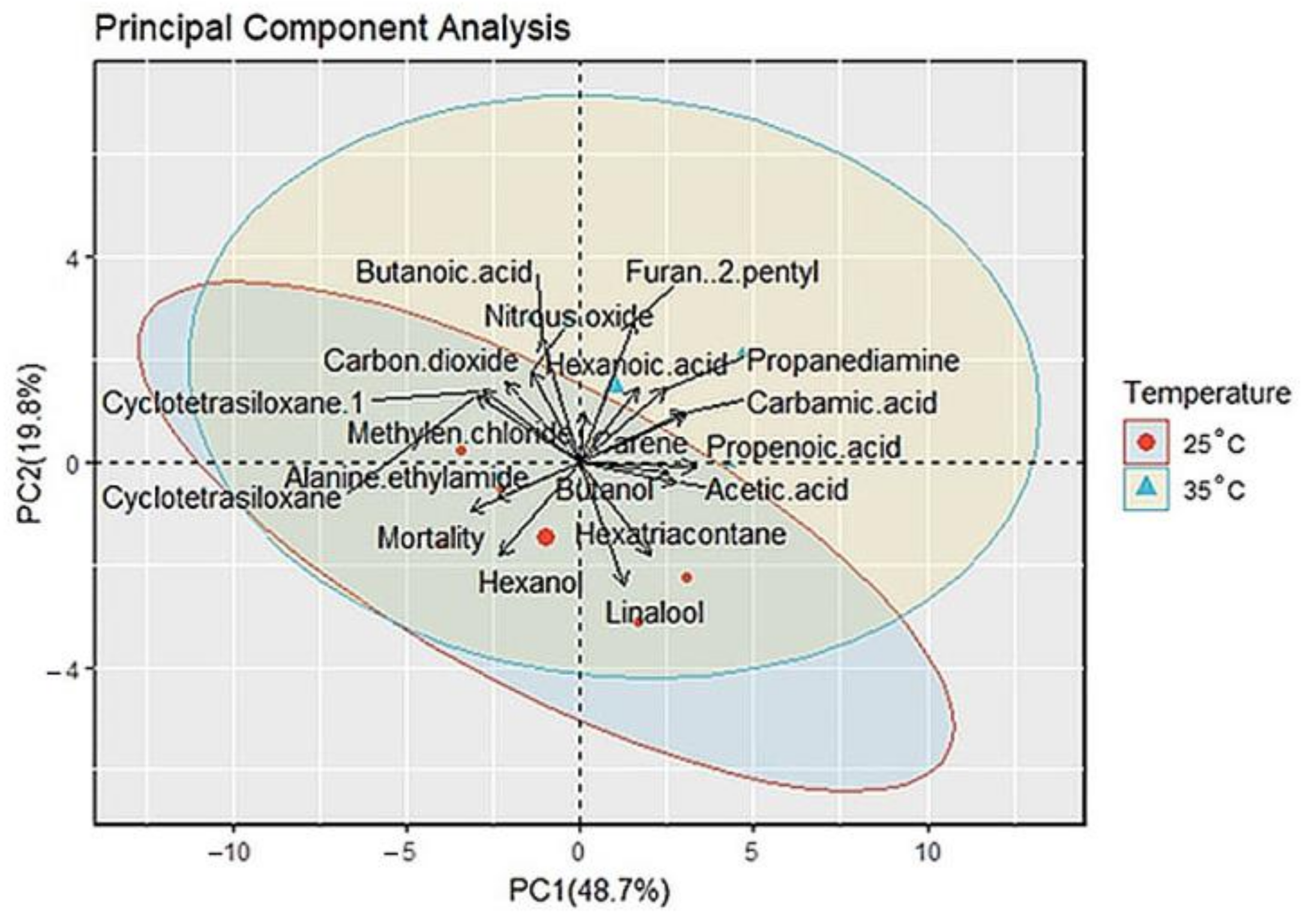
3.4. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kimura, S.; Sinha, N. Tomato (Solanum lycopersicum): A model fruit-bearing crop. Cold Spring Harb. Protoc. 2008, 3, pdb.emo105. [Google Scholar] [CrossRef]
- Seid, A.; Fininsa, C.; Mekete, T.; Decraemer, W.; Wesemael, W.M.L. Tomato (Solanum lycopersicum) and root-knot nematodes (Meloidogyne spp.)–A century-old battle. Nematology 2015, 17, 995–1009. [Google Scholar] [CrossRef]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The good, the bad, and the ugly of rhizosphere microbiome. In Probiotics and Plant Health; Springer: Singapore, 2017; pp. 253–290. [Google Scholar]
- Nicol, J.M.; Turner, S.J.; Coyne, D.L.; den Nijis, L.; Hockland, S.; Maafi, Z.T. Current Nematode Threats to World Agriculture in Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 21–43. [Google Scholar]
- Maqsood, A.; Wu, H.; Kamran, M.; Altaf, H.; Mustafa, A.; Ahmar, S.; Hong, N.T.T.; Tariq, K.; He, Q.; Chen, J.T. Variations in growth, physiology, and antioxidative defense responses of two tomato (Solanum lycopersicum) cultivars after co-infection of Fusarium oxysporum and Meloidogyne incognita. Agronomy 2020, 10, 159. [Google Scholar] [CrossRef]
- Barbary, A.; Djian-Caporalino, C.; Palloix, A.; Castagnone-Sereno, P. Host genetic resistance to root-knot nematodes, Meloidogyne spp., in Solanaceae: From genes to the field. Pest Manag. Sci. 2015, 71, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Sharma, I.P.; Sharma, A. Physiological and biochemical changes in tomato cultivar PT-3 with dual inoculation of mycorrhiza and PGPR against root-knot nematode. Symbiosis 2017, 71, 175–183. [Google Scholar] [CrossRef]
- Fuller, V.L.; Lilley, C.J.; Urwin, P.E. Nematode resistance. New Phytol. 2008, 180, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Matthiessen, J.N.; Kirkegaard, J.A. Biofumigation and enhanced biodegradation: Opportunity and challenge in soilborne pest and disease management. CRC Crit. Rev. Plant Sci. 2006, 25, 235–265. [Google Scholar] [CrossRef]
- Desmedt, W.; Mangelinckx, S.; Kyndt, T.; Vanholme, B. A phytochemical perspective on plant defense against nematodes. Front. Plant Sci. 2020, 11, 602079. [Google Scholar] [CrossRef]
- Seigler, D.S. Plant Secondary Metabolism, 1st ed.; Springer Science & Business Media: New York, NY, USA, 1998; p. 711. [Google Scholar]
- Hartmann, T. From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef]
- Parsiaaref, S.; Cao, A.; Li, Y.; Ebadollahi, A.; Parmoon, G.; Wang, Q.; Yan, D.; Fang, W.; Zhang, M. Nematicidal and toxicity effects of Eupatorium adenophorum Spreng against the root-knot nematode Meloidogyne incognita in soil producing cucumber. Agriculture 2023, 13, 1109. [Google Scholar] [CrossRef]
- Sun, W.; Liu, S.S.; Zhao, C.C. Biological properties of active compounds from Ageratina adenophora. SAGE Open Med. 2023, 11, 7964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.L.; Wu, M.; Zhang, J.J.; Irwin, D.; Gu, Y.C.; Shi, Q.W. Chemical constituents of plants from the genus Eupatorium. Chem. Biodivers. 2008, 5, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Curtis, R.H. Plant-nematode interactions: Environmental signals detected by the nematode’s chemosensory organs control changes in the surface cuticle and behaviour. Parasite 2008, 15, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Rasmann, S.; Hiltpold, I.; Ali, J. The role of root–produced volatile secondary metabolites in mediating soil interactions. In Advances in Selected Plant Physiology Aspects; Montanaro, G., Dichio, B., Eds.; InTech Open Access: London, UK, 2012; pp. 269–290. [Google Scholar] [CrossRef]
- Samara, R. Evaluation of 11 potential trap crops for root-knot nematode (RKN) control under glasshouse conditions. Open Agric. 2022, 7, 61–68. [Google Scholar] [CrossRef]
- Zhang, P.; Wei, J.N.; Zhao, C.; Zhang, Y.F.; Li, C.Y.; Li, S.S.; Dicke, M.; Yu, X.P.; Turlings, T.C.J. Airborne host-plant manipulation by whiteflies via an inducible blend of plant volatiles. Proc. Natl. Acad. Sci. USA 2019, 116, 7387–7396. [Google Scholar] [CrossRef] [PubMed]
- Santhosh, J.E.; Beena, B.; Ramana, K.V. Tropical soil microflora of spice-based cropping systems as potential antagonists of root-knot nematodes. J. Invert. Pathol. 2005, 88, 218–225. [Google Scholar]
- Hartman, K.M.; Sasser, J.N. Identification of Meloidogyne species on the basis of differential host test and perineal—Pattern morphology. In An Advanced Treatise on Meloidogyne, Methodology; Barker, K.R., Carter, C.C., Sasser, J.N., Eds.; North Carolina State University Graphics: Raleigh, NC, USA, 1985; Volume 2, pp. 69–77. [Google Scholar]
- Hussey, R.S.; Barker, K. A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Baermann, G. Eine einfache methode zur auffindung von ankylostomum (Nematoden) larven in Erdproben. Geneeskd. Tijdschr. Ned. Indie 1917, 57, 131–137. [Google Scholar]
- De Ley, I.T.; Karssen, G.; De Ley, P.; Vierstraete, A.; Waeyenberge, L.; Moens, M.; Vanfleteren, J. Phylogenetic analyses of internal transcribed spacer region sequences within Meloidogyne. J. Nematol. 1999, 31, 530–531. [Google Scholar]
- Mao, L.; Yan, D.; Wang, Q.; Li, Y.; Ouyang, C.; Liu, P.; Shen, J.; Guo, M.; Cao, A. Evaluation of the combination of dimethyl disulfide and dazomet as an efficient methyl bromide alternative for cucumber production in China. J. Agric. Food Chem. 2014, 62, 4864–4869. [Google Scholar] [CrossRef]
- Conklin, A.R. Introduction to Soil Chemistry. Analysis and Instrumentation, 3rd ed.; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2005; p. 218. [Google Scholar]
- Kaur, K.; Mahajan, R.; Deepak, B. A review of various soil moisture measurement techniques. Int. J. Innov. Res. Sci. Eng. Technol. 2016, 5, 5774–5778. [Google Scholar]
- Ahmadi Mansourabad, M.; Kargar Bideh, A.; Abdollahi, M. Effects of some micronutrients and macronutrients on the root—Knot nematode, Meloidogyne incognita, in greenhouse cucumber (Cucumis sativus cv. Negin). J. Crop. Prot. 2016, 5, 507–517. [Google Scholar] [CrossRef]
- Andrade-Garda, J.M.; Carlosena-Zubieta, A.; Maria Paz Gomez-Carracedo, M.P.; Maestro-Saavedra, M.; Prieto-Blanco, M.C.; Soto-Ferreiro, R.M. Fundamental Calculations in Analytical Chemistry. In Problems of Instrumental Analytical Chemistry: A Hands-on Guide, Kindle ed.; World Scientific Publishing Europe Ltd.: London, UK, 2017; pp. 1–40. [Google Scholar]
- Kundu, A.; Saha, S.; Walia, S.; Dutta, T.K. Antinemic potentiality of chemical constituents of Eupatorium adenophorum Spreng leaves against Meloidogyne incognita. Natl. Acad. Sci. Lett. 2016, 39, 145–149. [Google Scholar] [CrossRef]
- Schneider, P.; Orelli, O. Entomological internship. In Entomologisches Praktikum; Verlag, H.R., Sauerlander Co.: Aarau, Switzerland, 1947; p. 237. [Google Scholar]
- Parmoon, G.; Moosavi, S.A.; Ataollah Siadat, S. Descriptions of okra seed longevity loss behavior using nonlinear regression models. Adv. Hort. Sci. 2019, 33, 301–312. [Google Scholar]
- Deng, X.; Wang, X.; Li, G. Nematicidal effects of volatile organic compounds from microorganisms and plants on plant-parasitic nematodes. Microorganisms 2022, 10, 1201. [Google Scholar] [CrossRef] [PubMed]
- Teufack, S.E.N.; Tayo, G.N.; Alidou, M.N.; Yondo, J.; Djiomene, A.F.; Poné, J.W.; Mbida, F.M. Anthelminthic properties of Methylene chloride-methanol (1:1) extracts of two Cameroonians medicinal plants on Heligmosomoides bakeri (Nematoda: Heligmosomatidea). BMC Complement. Altern. Med. 2017, 17, 400. [Google Scholar] [CrossRef]
- Ntalli, N.; Ratajczak, M.; Oplos, C.; Menkissoglu-Spiroudi, U.; Adamski, Z. Acetic acid, 2-Undecanone, and (E)-2-Decenal ultrastructural malformations on Meloidogyne incognita. J. Nematol. 2016, 48, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, Y.; Ge, J.; Xie, B.; Zhu, S.; Cheng, X. Effects of α-pinene on the pinewood nematode (Bursaphelenchus xylophilus) and its symbiotic bacteria. PLoS ONE 2019, 14, e0221099. [Google Scholar] [CrossRef] [PubMed]
- Duschatzky, C.B.; Martinez, A.N.; Almeida, N.V.; Bonivardo, S.L. Nematicidal activity of the essential oils of several Argentina plants against the root-knot nematode. Essent. Oil Res. 2004, 16, 626–628. [Google Scholar] [CrossRef]
- Liarzi, O.; Bucki, P.; Miyara, S.B.; Ezra, D. Bioactive volatiles from an endophytic Daldinia cf. concentrica isolate affect the viability of the plant parasitic nematode Meloidogyne javanica. PLoS ONE 2016, 11, e0168437. [Google Scholar] [CrossRef]
- Buda, V.; Cepulyte-Rakauskiene, R. The effect of linalool on second-stage juveniles of the potato cyst nematodes Globodera rostochiensis and G. pallida. J. Nematol. 2011, 43, 149–151. [Google Scholar]
- Hirano, N.; Sakamoto, K. Linalool odor stimulation improves heat stress tolerance and decreases fat accumulation in nematodes. Biosci. Biotechnol. Biochem. 2019, 83, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Ndomo-Moualeu, A.F.; Ulrichs, C.; Adler, C. Behavioral responses of Callosobruchus maculatus to volatile organic compounds found in the headspace of dried green pea seeds. J. Pest. Sci. 2016, 89, 107–116. [Google Scholar] [CrossRef]
- Sari, D.I.P.; Oemry, S.; Safni, I.; Lubis, K.; Tantawi, A.R. Use of organic waste as biofumigant for controlling root-knot nematodes (Meloidogyne spp.) on potato. IOP Conf. Ser. Earth Environ. Sci. 2018, 122, 012065. [Google Scholar] [CrossRef]
- da Silva Rocha, F.; Campos, V.P.; Fernandes, M.D.F.G.; Muniz, M.D.F.S. Migration and reproduction of Meloidogyne in-cognita in two soil textures. Nematropica 2016, 46, 162–171. [Google Scholar]
- Horiuchi, J.; Prithiviraj, B.; Bais, H.P.; Kimball, B.A.; Vivanco, J.M. Soil nematodes mediate positive interactions between legume plants and rhizobium bacteria. Planta 2005, 222, 848–857. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | DF | Means Square | |||
|---|---|---|---|---|---|
| Linalool | Nonanal | Methylene Chloride | 2-Decenal | ||
| Time | 4 | 375.10 ** | 220.13 ** | 190.19 ** | 2449.24 ** |
| Temperature | 1 | 60.87 ** | 28.12 ** | 86.08 ** | 51.18 ** |
| Concentration | 5 | 137.89 ** | 426.18 ** | 1665.10 ** | 1340.18 ** |
| Time × temperature | 4 | 27.96 ** | 27.73 ** | 37.57 ** | 87.70 ** |
| Time × concentration | 20 | 20.39 ** | 14.84 ** | 41.42 ** | 515.5 ** |
| Temp × concentration | 5 | 7.25 ** | 5.84 ** | 14.35 ** | 6.00 ** |
| Time × temperature × concentration | 20 | 1.52 ns | 4.38 ns | 7.39 * | 9.64 ** |
| Error | 120 | 2.44 | 4.41 | 4.24 | 3.45 |
| Coefficient of Variation (%) | - | 12.4 | 12.7 | 8.8 | 8.4 |
| Chemical Compounds | Time (Weeks) | Model | Parameters Model | R2 | RMSE | AICc | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MRmax (%) | slope | c | EC50 (µg/mL) | ECmax (µg/mL) | MRmin (%) | ||||||
| Linalool | 1 | W | 20.6 ± 3.8 | 103.6 ± 8.3 | 1.02 ± 1.3 | - | 16.4 ± 1.9 | 12.8 ± 1.4 | 0.951 | 1.59 | −6.8 |
| 2 | W | 16.9 ± 1.2 | 109.1 ± 6.2 | 1.04 ± 0.3 | - | 14.0 ± 3.4 | 11.8 ± 0.2 | 0.999 | 0.39 | −29.7 | |
| 3 | W | 16.0 ± 0.7 | 111.0 ± 6.8 | 1.33 ± 0.2 | - | 14.3 ± 2.0 | 9.6 ± 0.4 | 0.995 | 0.28 | −14.1 | |
| 4 | W | 15.3 ± 1.5 | 66.3 ± 9.4 | 1.21 ± 0.2 | - | 7.7 ± 2.3 | 8.2 ± 0.3 | 0.998 | 0.28 | −14.2 | |
| 8 | W | 15.1 ± 1.3 | 94.6 ± 11.9 | 1.01 ± 0.1 | - | 6.1 ± 1.4 | 5.4 ± 0.3 | 0.999 | 0.23 | −16.3 | |
| Nonanal | 1 | W | 23.0 ± 0.9 | 156.1 ± 37.3 | 1.07 ± 0.2 | - | 27.4 ± 7.8 | 13.8 ± 0.6 | 0.995 | 0.60 | −5.0 |
| 2 | W | 22.0 ± 0.7 | 170.0 ± 12.2 | 1.06 ± 0.1 | - | 24.5 ± 6.3 | 12.2 ± 0.2 | 0.999 | 0.21 | −17.5 | |
| 3 | W | 21.9 ± 1.1 | 76.3 ± 22.5 | 1.05 ± 0.5 | - | 25.8 ± 8.6 | 11.2 ± 0.5 | 0.993 | 0.64 | −4.1 | |
| 4 | W | 20.1 ± 3.4 | 85.2 ± 39.3 | 1.01 ± 0.5 | - | 18.9 ± 1.4 | 10.9 ± 1.3 | 0.978 | 1.14 | 2.8 | |
| 8 | W | 20.8 ± 2.3 | 75.2 ± 16.6 | 1.03 ± 0.2 | - | 15.6 ± 6.0 | 8.4 ± 0.9 | 0.993 | 0.92 | 0.2 | |
| Methylene chloride | 1 | G | 28.5 ± 0.9 | 21.8 ± 5.6 | - | 10.2 ± 4.3 | - | 0.961 | 1.51 | 48.8 | |
| 2 | G | 25.2 ± 0.8 | 61.9 ± 6.4 | - | 11.9 ± 8.5 | - | 0.994 | 0.44 | 34.0 | ||
| 3 | G | 31.0 ± 1.4 | 54.3 ± 9.6 | - | 23.1 ± 2.4 | - | 0.972 | 1.66 | 49.9 | ||
| 4 | G | 36.4 ± 0.5 | 51.3 ± 3.3 | - | 28.1 ± 6.0 | - | 0.998 | 0.57 | 37.2 | ||
| 8 | G | 33.2 ± 1.0 | 58.1 ± 8.3 | - | 33.9 ± 6.9 | - | 0.989 | 1.15 | 30.0 | ||
| 2-Decenal | 1 | G | 20.6 ± 1.7 | 99.1 ± 7.2 | - | 36.8 ± 6.5 | - | 0.944 | 2.48 | 46.3 | |
| 2 | G | 22.8 ± 1.1 | 70.2 ± 30.2 | - | 35.4 ± 3.7 | - | 0.909 | 1.37 | 47.6 | ||
| 3 | G | 23.4 ± 1.1 | 78.2 ± 30.2 | - | 31.0 ± 3.7 | - | 0.988 | 0.60 | 35.4 | ||
| 4 | G | 35.1 ± 1.6 | 68.4 ± 9.0 | - | 15.6 ± 1.2 | - | 0.982 | 0.62 | 38.1 | ||
| 8 | G | 62.3 ± 4.1 | 64.5 ± 12.8 | - | 95.7 ± 5.4 | - | 0.979 | 3.82 | 42.1 | ||
| Chemical Compounds | Temperature (°C) | Model | Parameters Model | R2 | RMSE | AICc | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MRmax (%) | slope | c | EC50 (µg/mL) | ECmax (µg/mL) | MRmin (%) | ||||||
| Linalool | 25 | W | 15.2 ± 0.2 | 141.1 ± 8.1 | 1.18 ± 0.1 | - | 15.6 ± 2.8 | 9.7 ± 0.2 | 0.999 | 0.06 | −33.0 |
| 35 | W | 14.0 ± 1.4 | 119.5 ± 7.0 | 1.16 ± 0.2 | - | 10.1 ± 2.0 | 8.2 ± 0.4 | 0.998 | 0.22 | −17.1 | |
| Nonanal | 25 | W | 19.9 ± 2.5 | 76.5 ± 9.4 | 1.03 ± 0.9 | - | 20.4 ± 9.7 | 12.9 ± 0.5 | 0.991 | 0.65 | −4.0 |
| 35 | W | 21.3 ± 2.5 | 113.6 ± 12.6 | 1.02 ± 0.1 | - | 15.6 ± 7.6 | 13.1 ± 1.0 | 0.991 | 0.73 | −2.6 | |
| Methylene chloride | 25 | G | 30.5 ± 9.6 | 59.8 ± 7.3 | - | 18.1 ± 2.3 | - | - | 0.990 | 0.44 | 42.2 |
| 35 | G | 33.5 ± 7.0 | 58.9 ± 6.1 | - | 21.9 ± 4.0 | - | - | 0.994 | 0.78 | 40.8 | |
| 2-Decenal | 25 | G | 30.9 ± 1.5 | 95.4 ± 10.6 | - | 12.3 ± 1.6 | - | - | 0.979 | 1.18 | 45.8 |
| 35 | G | 32.4 ± 1.0 | 81.6 ± 11.3 | - | 18.7 ± 2.8 | - | - | 0.991 | 0.88 | 42.3 | |
| Chemical Compounds | Time (Weeks) | Temperature (°C) | Parameters Model | R2 | RMSE | AICc | |||
|---|---|---|---|---|---|---|---|---|---|
| MRmax (%) | Slope | EC50 (µg/mL) | MRmin (%) | ||||||
| Methylene chloride | 1 | 25 | 25.3 ± 1.6 | 68.2 ± 35.0 | 28.0 ± 7.5 | 9.5 ± 8.4 | 0.984 | 0.90 | −6.8 |
| 35 | 27.0 ± 1.0 | 44.3 ± 11.1 | 26.2 ± 9.6 | 11.4 ± 3.5 | 0.998 | 0.36 | −9.6 | ||
| 2 | 25 | 29.7 ± 2.0 | 12.4 ± 6.8 | 36.1 ± 8.4 | 12.9 ± 2.2 | 0.960 | 2.23 | −2.2 | |
| 35 | 32.4 ± 7.6 | 16.7 ± 6.3 | 35.6 ± 6.0 | 13.9 ± 1.6 | 0.980 | 1.63 | −6.2 | ||
| 3 | 25 | 30.9 ± 8.9 | 53.0 ± 23.3 | 56.6 ± 14.3 | 11.8 ± 3.8 | 0.981 | 1.62 | −6.2 | |
| 35 | 35.6 ± 5.4 | 31.7 ± 12.9 | 70.6 ± 12.2 | 14.5 ± 1.7 | 0.979 | 2.08 | −5.8 | ||
| 4 | 25 | 33.3 ± 4.4 | 37.3 ± 8.6 | 75.4 ± 7.2 | 13.3 ± 1.0 | 0.994 | 1.09 | −8.8 | |
| 35 | 36.0 ± 5.9 | 56.1 ± 20.8 | 58.7 ± 12.2 | 10.7 ± 4.8 | 0.987 | 1.70 | −7.4 | ||
| 8 | 25 | 34.9 ± 9.6 | 63.6 ± 29.1 | 62.5 ± 15.5 | 13.6 ± 5.0 | 0.981 | 1.79 | −6.2 | |
| 35 | 43.1 ± 7.6 | 46.1 ± 4.0 | 54.6 ± 4.0 | 11.8 ± 1.0 | 0.998 | 0.55 | −9.6 | ||
| 2-Decenal | 1 | 25 | 19.6 ± 1.9 | 40.8 ± 5.4 | 81.9 ± 4.4 | 13.1 ± 0.2 | 0.998 | 0.18 | −9.6 |
| 35 | 21.4 ± 1.9 | 82.3 ± 18.9 | 93.2 ± 16.6 | 12.4 ± 0.7 | 0.995 | 0.32 | −9.0 | ||
| 2 | 25 | 21.1 ± 3.0 | 139.9 ± 16.2 | 65.0 ± 15.5 | 10.4 ± 4.3 | 0.995 | 0.36 | −9.1 | |
| 35 | 22.7 ± 1.6 | 54.9 ± 19.8 | 74.5 ± 15.3 | 13.0 ± 1.3 | 0.969 | 0.89 | −3.8 | ||
| 3 | 25 | 22.6 ± 1.0 | 130.4 ± 15.8 | 48.3 ± 17.5 | 8.0 ± 5.7 | 0.989 | 0.85 | −7.8 | |
| 35 | 24.8 ± 2.5 | 111.9 ± 12.3 | 53.5 ± 11.2 | 9.0 ± 6.1 | 0.991 | 0.64 | −8.2 | ||
| 4 | 25 | 31.6 ± 6.3 | 17.1 ± 7.7 | 35.5 ± 7.2 | 14.8 ± 1.9 | 0.971 | 1.84 | −4.2 | |
| 35 | 41.6 ± 18.6 | 11.1 ± 6.5 | 37.0 ± 8.6 | 13.8 ± 3.5 | 0.967 | 3.49 | −3.4 | ||
| 8 | 25 | 62.4 ± 4.6 | 58.3 ± 3.2 | 83.0 ± 2.6 | 11.8 ± 0.8 | 0.999 | 0.60 | −9.8 | |
| 35 | 70.5 ± 8.8 | 54.2 ± 1.4 | 110.5 ± 1.1 | 13.7 ± 0.2 | 0.999 | 0.33 | −9.8 | ||
| Source of Variation | DF | Means Square | |
|---|---|---|---|
| Leaves | Root-Stems | ||
| Time | 4 | 9293.31 ** | 215.3 ** |
| Temperature | 1 | 10,298.5 ** | 249.3 ** |
| Concentration | 5 | 22,683.2 ** | 176.7 ** |
| Time × temperature | 4 | 1034.9 ** | 12.63 ** |
| Time × concentration | 20 | 687.3 ** | 17.73 ** |
| Temp × concentration | 5 | 409.4 ** | 16.51 ** |
| Time × temperature × concentration | 20 | 95.042 ** | 9.18 * |
| Error | 120 | 38.15 | 3.561 |
| Coefficient of Variation (%) | - | 10.4 | 25.6 |
| Parts | Time (Weeks) | Temperature (°C) | Parameters Model | R2 | RMSE | AICc | ||
|---|---|---|---|---|---|---|---|---|
| MRmax (%) | Slope | EC50 (µg/mg) | ||||||
| Leaves | 1 | 25 | 47.6 ± 6.2 | 10.5 ± 3.7 | 71.6 ± 4.6 | 0.987 | 2.92 | 16.3 |
| 35 | 91.9 ± 3.7 | 32.7 ± 2.4 | 28.0 ± 1.5 | 0.999 | 1.50 | 48.7 | ||
| 2 | 25 | 73.1 ± 7.1 | 24.9 ± 7.1 | 25.7 ± 4.5 | 0.975 | 5.14 | 63.5 | |
| 35 | 95.4 ± 2.1 | 25.3 ± 3.9 | 21.9 ± 2.5 | 0.992 | 3.84 | 60.0 | ||
| 3 | 25 | 83.8 ± 3.4 | 26.5 ± 8.4 | 22.3 ± 5.2 | 0.968 | 6.70 | 66.7 | |
| 35 | 97.5 ± 5.1 | 28.8 ± 4.3 | 21.5 ± 2.7 | 0.990 | 4.44 | 61.7 | ||
| 4 | 25 | 86.1 ± 9.5 | 12.1 ± 2.7 | 7.1 ± 2.3 | 0.975 | 5.82 | 65.0 | |
| 35 | 99.9 ± 3.3 | 8.8 ± 2.3 | 5.2 ± 2.1 | 0.970 | 7.00 | 67.2 | ||
| 8 | 25 | 89.5 ± 4.1 | 7.8 ± 2.5 | 5.2 ± 2.4 | 0.963 | 7.78 | 68.5 | |
| 35 | 100.2 ± 5.7 | 6.5 ± 1.3 | 4.0 ± 1.1 | 0.991 | 4.04 | 60.6 | ||
| Root -stems | 1 | 25 | 10.8 ± 0.2 | 0.9 ± 0.1 | 267.8 ± 8.3 | 0.994 | 0.12 | 18.6 |
| 35 | 10.3 ± 0.2 | 1.1 ± 0.2 | 231.2 ± 24.9 | 0.988 | 0.27 | 25.7 | ||
| 2 | 25 | 11.7 ± 0.5 | 1.2 ± 0.2 | 214.2 ± 17.3 | 0.976 | 0.22 | 2.8 | |
| 35 | 10.3 ± 0.2 | 1.1 ± 0.2 | 211.2 ± 14.9 | 0.976 | 0.22 | 25.7 | ||
| 3 | 25 | 11.5 ± 0.3 | 0.6 ± 0.2 | 103.2 ± 5.3 | 0.987 | 0.31 | 29.6 | |
| 35 | 11.9 ± 0.4 | 0.7 ± 0.1 | 31.7 ± 4.8 | 0.991 | 0.38 | 32.1 | ||
| 4 | 25 | 11.6 ± 0.3 | 0.6 ± 0.1 | 55.0 ± 6.4 | 0.992 | 0.29 | 29.1 | |
| 35 | 12.1 ± 0.7 | 0.9 ± 0.2 | 17.4 ± 4.8 | 0.983 | 0.65 | 38.7 | ||
| 8 | 25 | 10.5 ± 0.1 | 0.8 ± 0.2 | 27.5 ± 5.3 | 0.984 | 0.54 | 36.4 | |
| 35 | 13.5 ± 0.3 | 0.7 ± 0.1 | 3.0 ± 1.7 | 0.997 | 0.32 | 30.1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parsiaaref, S.; Cao, A.; Li, Y.; Ebadollahi, A.; Parmoon, G.; Wang, Q.; Yan, D.; Fang, W.; Huang, B.; Zhang, M. The Main Compounds of Bio-Fumigant Plants and Their Role in Controlling the Root-Knot Nematode Meloidogyne incognita (Kofoid and White) Chitwood. Agriculture 2024, 14, 261. https://doi.org/10.3390/agriculture14020261
Parsiaaref S, Cao A, Li Y, Ebadollahi A, Parmoon G, Wang Q, Yan D, Fang W, Huang B, Zhang M. The Main Compounds of Bio-Fumigant Plants and Their Role in Controlling the Root-Knot Nematode Meloidogyne incognita (Kofoid and White) Chitwood. Agriculture. 2024; 14(2):261. https://doi.org/10.3390/agriculture14020261
Chicago/Turabian StyleParsiaaref, Shiva, Aocheng Cao, Yuan Li, Asgar Ebadollahi, Ghasem Parmoon, Qiuxia Wang, Dongdong Yan, Wensheng Fang, Bin Huang, and Min Zhang. 2024. "The Main Compounds of Bio-Fumigant Plants and Their Role in Controlling the Root-Knot Nematode Meloidogyne incognita (Kofoid and White) Chitwood" Agriculture 14, no. 2: 261. https://doi.org/10.3390/agriculture14020261
APA StyleParsiaaref, S., Cao, A., Li, Y., Ebadollahi, A., Parmoon, G., Wang, Q., Yan, D., Fang, W., Huang, B., & Zhang, M. (2024). The Main Compounds of Bio-Fumigant Plants and Their Role in Controlling the Root-Knot Nematode Meloidogyne incognita (Kofoid and White) Chitwood. Agriculture, 14(2), 261. https://doi.org/10.3390/agriculture14020261