1. Introduction
Brassicogethes aeneus Fabricius, 1775 is one of the most significant pests of spring and winter oilseed rape. Treatment against this pest has been used regularly for several decades and is causing problems with resistance to the insecticide's active ingredients. Insecticide resistance testing is carried out in most European countries, including the Czech Republic (CZ). Pollen beetle populations were highly resistant to the pyrethroid lambda-cyhalothrin, according to 2014–2015 test results from CZ [
1]. Relatively high proportions of the populations also showed resistance to the pyrethroid tau-fluvalinate [
2]. At present, especially if the number of registered active ingredients is reduced, a reduction in the level of resistance cannot be expected. Many other pollen beetle species were present in the oilseed rape in addition to
Brassicogethes spp. (Coleoptera: Nitidulidae). The larvae of all
Brassicogethes species develop on Brassicaceae plants, and adults feed on the pollen of various family plants [
3,
4]. Usually, the pollen beetle (
Brassicogethes aeneus) is considered a pest of oilseed rape, although there are several other pollen beetle species that may inflict damage as well [
5].
The pollen beetles, which are the oilseed rape pests, formerly belonged to the genus
Meligethes Stephens, 1830. Until recently, the genus
Meligethes was one of the most numerous of the Nitidulidae family. Just in the Palearctic region, there are about 250 polinivorous species associated with plants of the families Brassicaceae, Lamiaceae, Fabaceae, and Rosaceae [
6]. The classification of this genus, as well as the subfamily Meligethinae, has been revised. Currently, this genus contains many groups of species that are morphologically similar and hardly distinguishable [
6]. Some species were excluded from the genus
Meligethes and newly classified as separate genera:
Boragogethes,
Brassicogethes,
Clypeogethes,
Fabogethes,
Genistogethes,
Lamiogethes,
Meligethes,
Pria, and
Sagittogethes [
7].
The beetles of the Nitidulidae family overwinter as adults, in early spring they leave the wintering ground and there is a possibility to find them feeding on the pollen of different species of flowering plants [
8]. There are many species of pollen beetles associated with Brassicaceae plants present in the Czech Republic. Namely:
Brassicogethes aeneus,
B. viridescens (Fabricius, 1787),
B. anthracinus (C. Brisout de Barneville, 1863),
B. coeruleovirens (Forster, 1849),
B. coracinus (Sturm, 1845),
B. czwalinai (Reitter, 1871),
B. subaeneus (Sturm, 1845),
B. matronalis (Audisio & Spornraft, 1990), and
Clypeogethes lepidii (Miller, 1852). Other species whose larval development is not associated with Brassicaceae plants but whose adults feed on the pollen of oilseed rape are
Boragogethes symphyti (Heer, 1841),
Cychramus luteus (Fabricius 1787),
Epurea aestiva (Linnaeus, 1758),
Fabogethes nigrescens (Stephens, 1830),
Genistogethes carinulatus (Förster, 1849),
Lamiogethes atramentarius (Förster, 1849),
Meligethes atratus (Olivier, 1790),
Pria dulcamare (Scopoli, 1763), and
Sagitogethes maurus (Sturm, 1845).
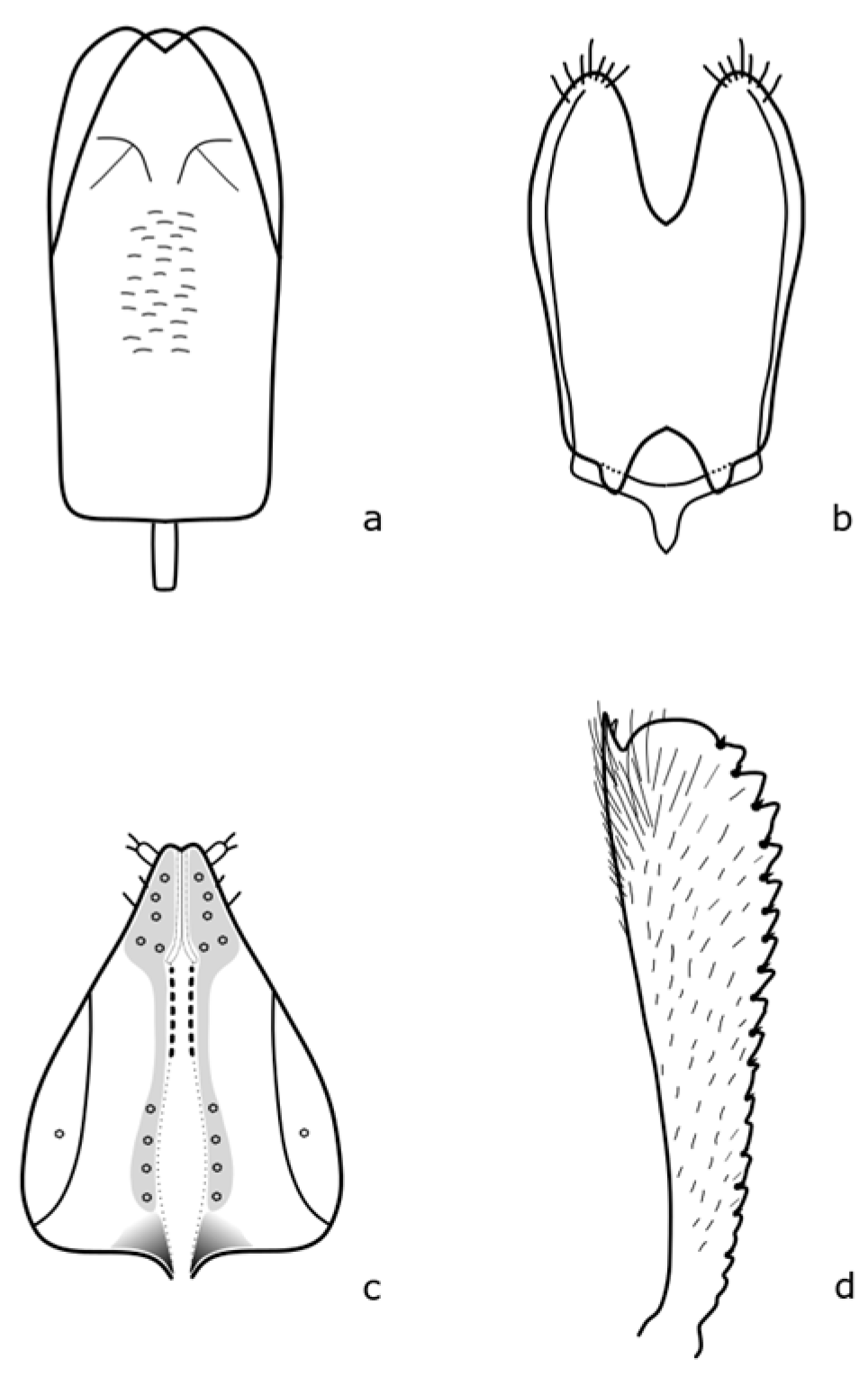
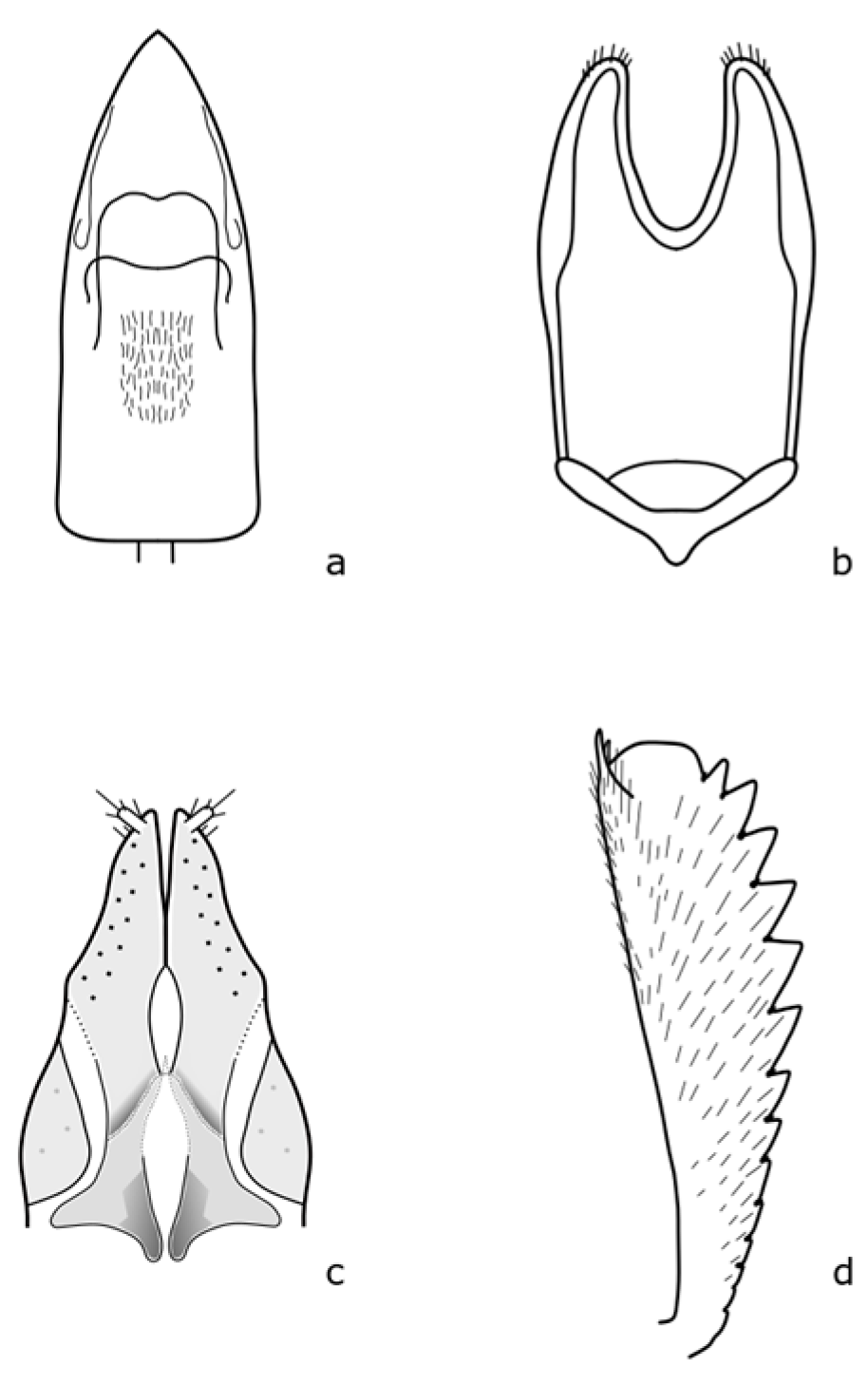
Brassicogethes aeneus biology is the best described. All developing stages are described according to Osborn [
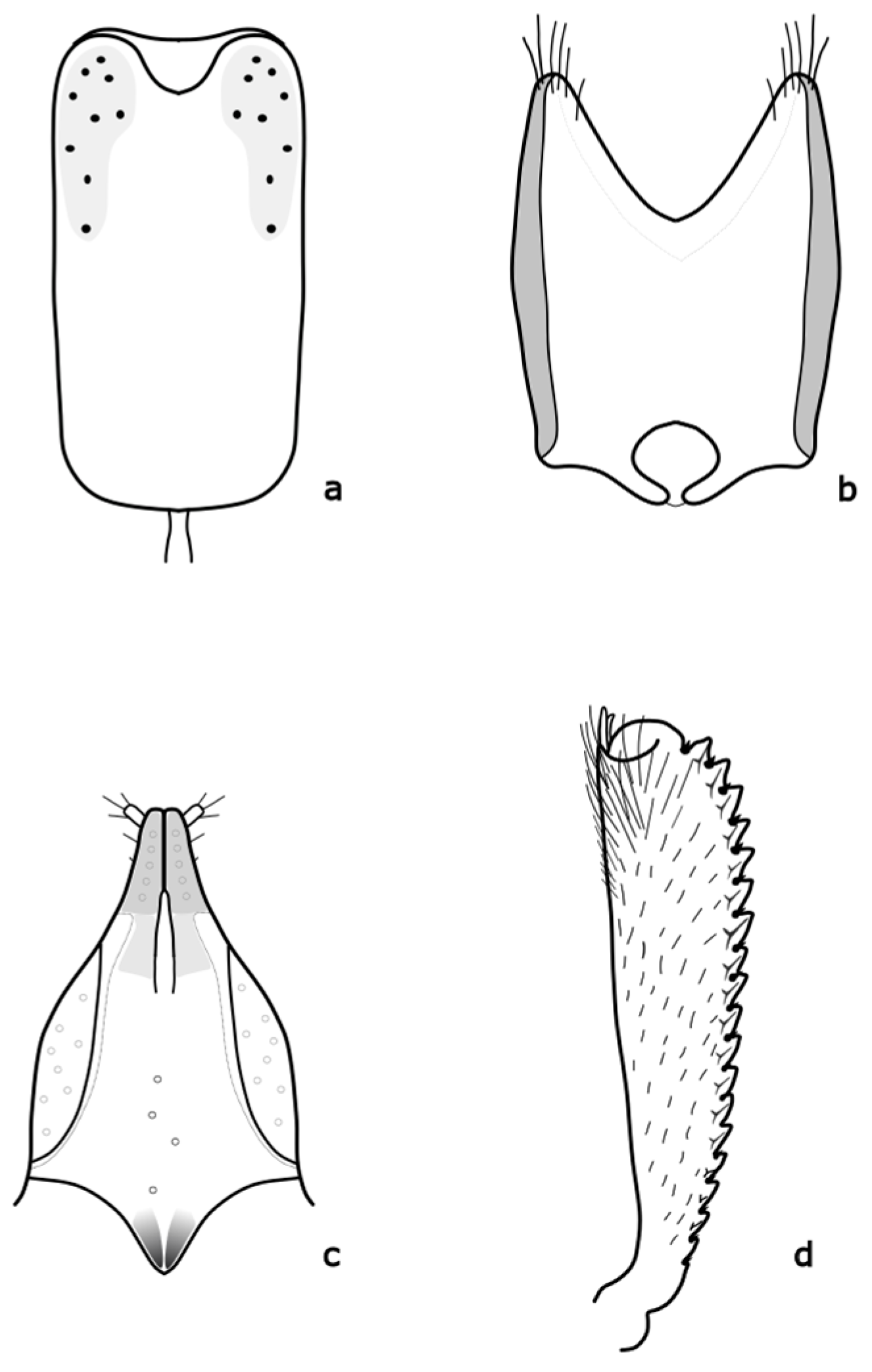
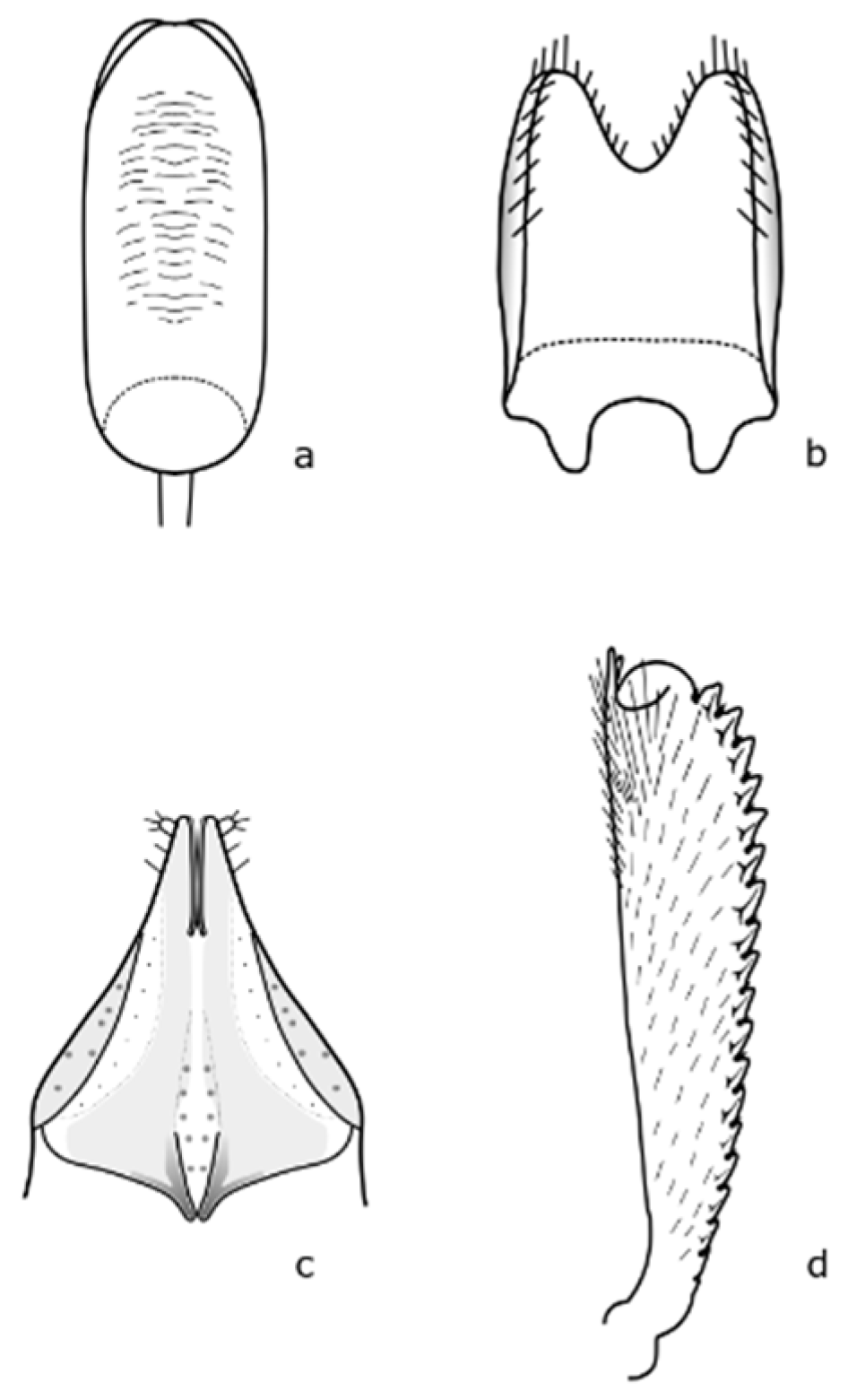
9]. Adults are 1.9–2.7 mm long, oval in shape, broadly rounded, and have a relatively broad head. The head and body are black with a metallic shine or green, blue, or purple. The legs are lighter, especially the front tibiae, up to dark yellow. The length of the antennae is almost as long as the width of the head. The anterior tibiae are finely toothed on the outer margin (
Figure A1 in
Appendix A). The middle femur lacks a tooth in the apical third. The eggs are cylindrical, 0.81 mm long, 0.29 mm wide, and whitish gray. The first instar larva is up to 4.4 mm long, and whitish with a brown to black scutellum. The pupa is about 2.35 mm long, oval-shaped, and creamy white colored.
B. aeneus occurs in Europe, North Africa, and West Asia in fields and forests [
10]. This species is oligophagous, their larvae are associated with species of the Brassicaceae family, especially the subfamilies Arabidae, Brassicae, and Hesperidae. It is a significant pest on all Brassicaceae plants, and its larvae develop on
Brassica oleracea var.
Capitata L.,
B. nigra (L.) W. D. J. Koch,
Sinapis alba L., and
S. arvensis L. [
11]. They also develop on
Barbarea verna (Mill.) Asch. and
B. vulgaris W. T. Aiton [
12] and the genera
Cakile and
Biscutella [
10].
B. aeneus prefers
Brassica rapa L. to
B. napus L. after overwintering due to its earlier flowering, as stated by Cook et al. [
13]. Borg [
14] studied pollen beetle preferences to the host plants and shows this sequence:
B. juncea (L.) Czern. >
B. napus >
S. alba. Kaasik et al. [
15] found that beetles are more attracted to
Sinapis alba than
B. napus. According to Veroman et al. [
16], females of the common pollen beetle prefer species of genus
Brassica for laying eggs over other species of this family, and if the larvae develop on
Raphanus sativus L., up to 35% of them remain undeveloped. Noronha and Mason [
17], said that
B. aeneus in Europe infests particularly winter oilseed rape, while spring oilseed rape is infested mainly by
B. viridescens.
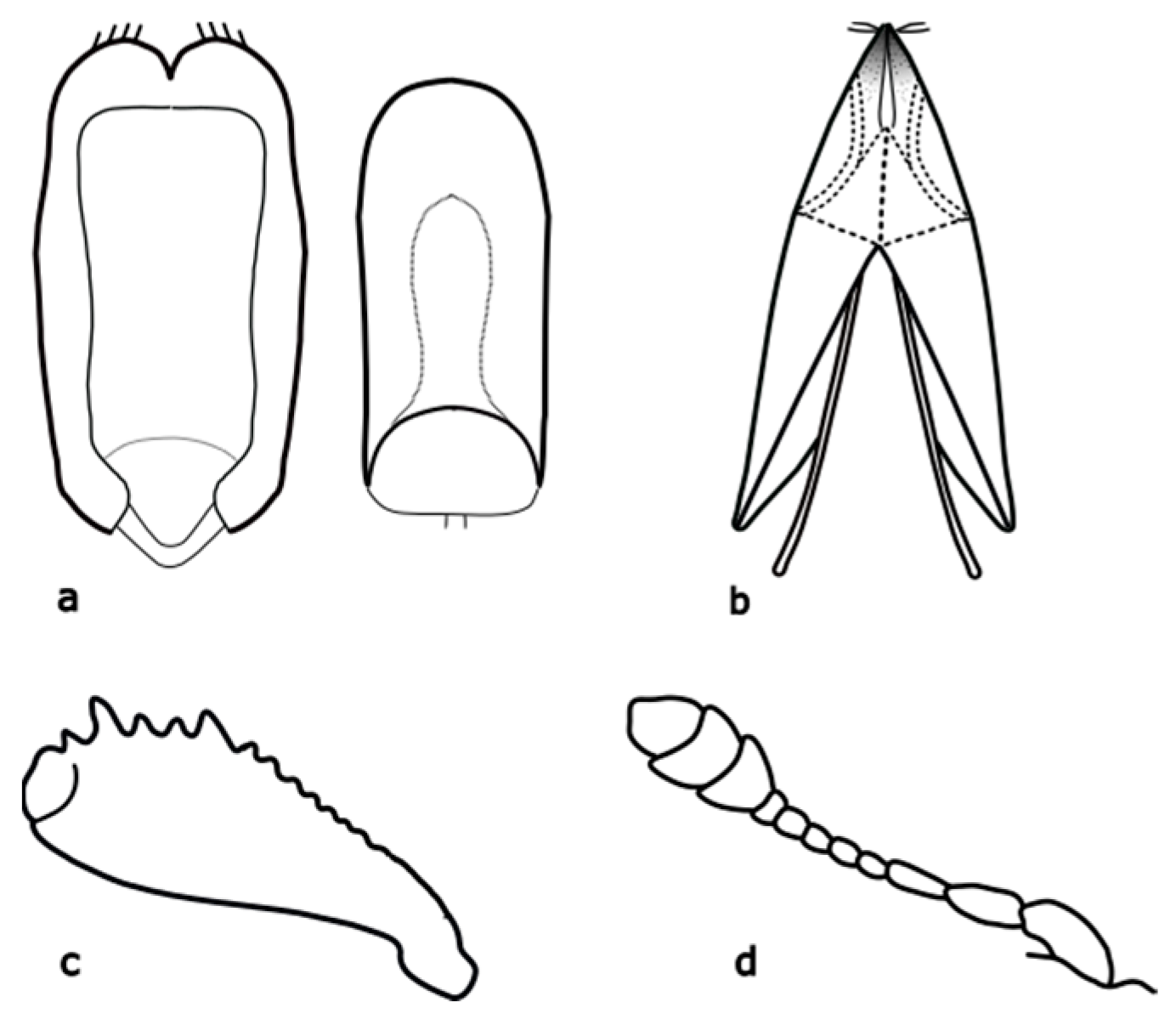
Brassicogethes viridescens (Fabricius, 1787) is 2.4–2.9 mm long. This species is similar to
B. aeneus, and is metallic shine colored with yellowish legs and finer punctuation at the dorsal part [
17]. Hoebeke & Wheeler [
18] pointed out that the legs, mouthpart, and proximal antennal segments are yellowish or orange-yellow. The body is elongated-oval, and the color is variable, usually dark green, often with bluish, violet, or less often black, bronze, or copper shining. Pubescence is fine, sparse, silver or yellow, and uniformly distributed. This species is slightly distinguished from the others by the presence of a subapical tooth along the posterior margin of the middle femur [
18]. Male and female genitalia bear important distinguishing marks (
Figure A2). This species is oligophagous on plants of the Brassicaceae family, mainly on the genus
Brassica spp. and
Sinapis spp. Borg [
14] shows the order of preferences for nutrient plants:
B. napus >
S. alba >
B. juncea. The beetles were found on flowers of different plant species—
Acer pseudoplatanus L.,
Tripleurospermum inodorum (L.) Sch.-Bip.,
Crataegus sp., and on plants of the family Ranunculaceae as well. Audisio et al. [
19] present
Diplotaxis as a host plant for larvae. Its larval biology is similar to
B. aeneus [
17,
20] and it is associated with oilseed rape during its larval development [
11] and also with species of the Brassicaceae family [
21]. Females lay eggs on flower bud bases [
22]. As stated by Mason et al. [
23], this pollen beetle species is a significant pest of oilseed rape in the Palearctic region and an invasive pest on this plant in Canada. Osborne [
9] and Sorauer [
24] considered this species to be the second most important pest of oilseed rape, with
B. aeneus in the first place. This species is widespread [
8], especially in disturbed habitats [
19].
B. viridescens (
Figure A2) causes damage in North America [
17].
B. aeneus and
B. viridescens are considered to be the most damaging species to cruciferous crops [
25]. Zimmer et al. [
26] explain that the pollen beetles select pollen-rich plants in both periods—adults in the spring before they lay their eggs and new beetles in the summer after they have hatched—and also feed on pollen from plants outside the Brassicaceae family. For overwintering, they migrate to places outside the field, especially to forests or similar habitats that they prefer [
27]. Although
B. aeneus is considered to be a pest of oilseed rape, Alford [
28] states that it can also damage the flowers of fruit species in large numbers in an attempt to obtain pollen. In contrast, the benefits of
B. aeneus and
B. viridescens for raspberry pollination are reported by Levesque and Levesque [
29].
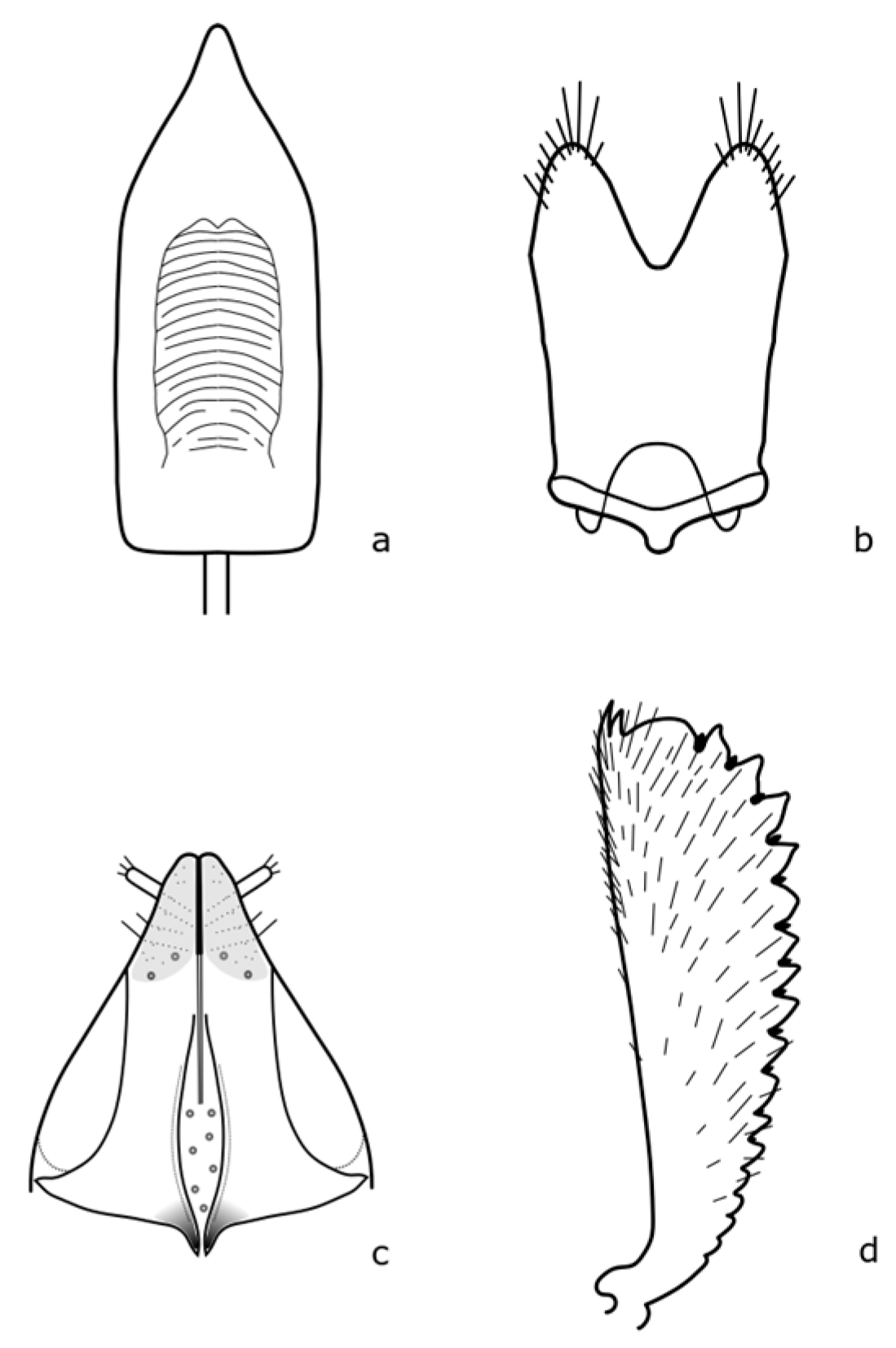
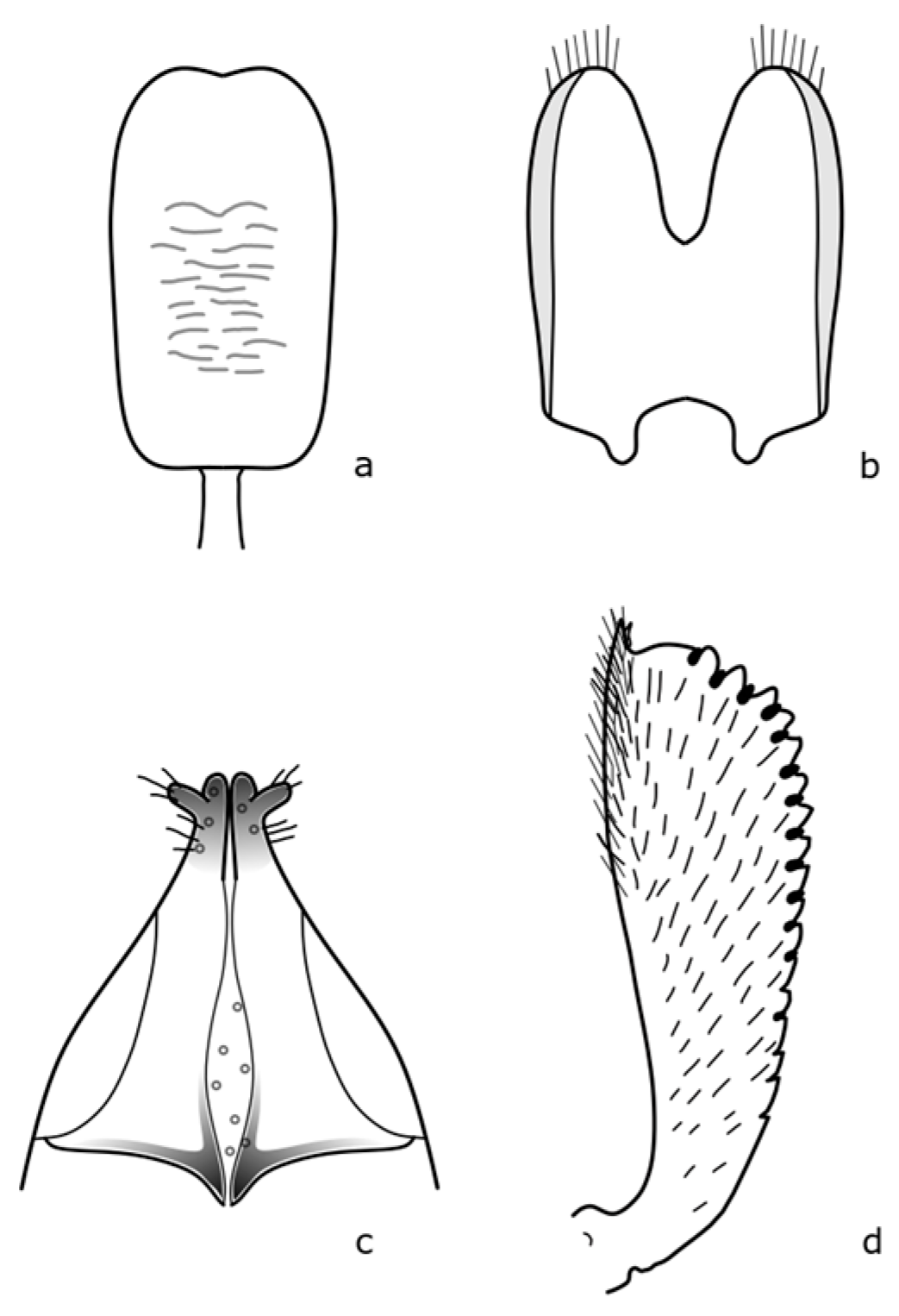
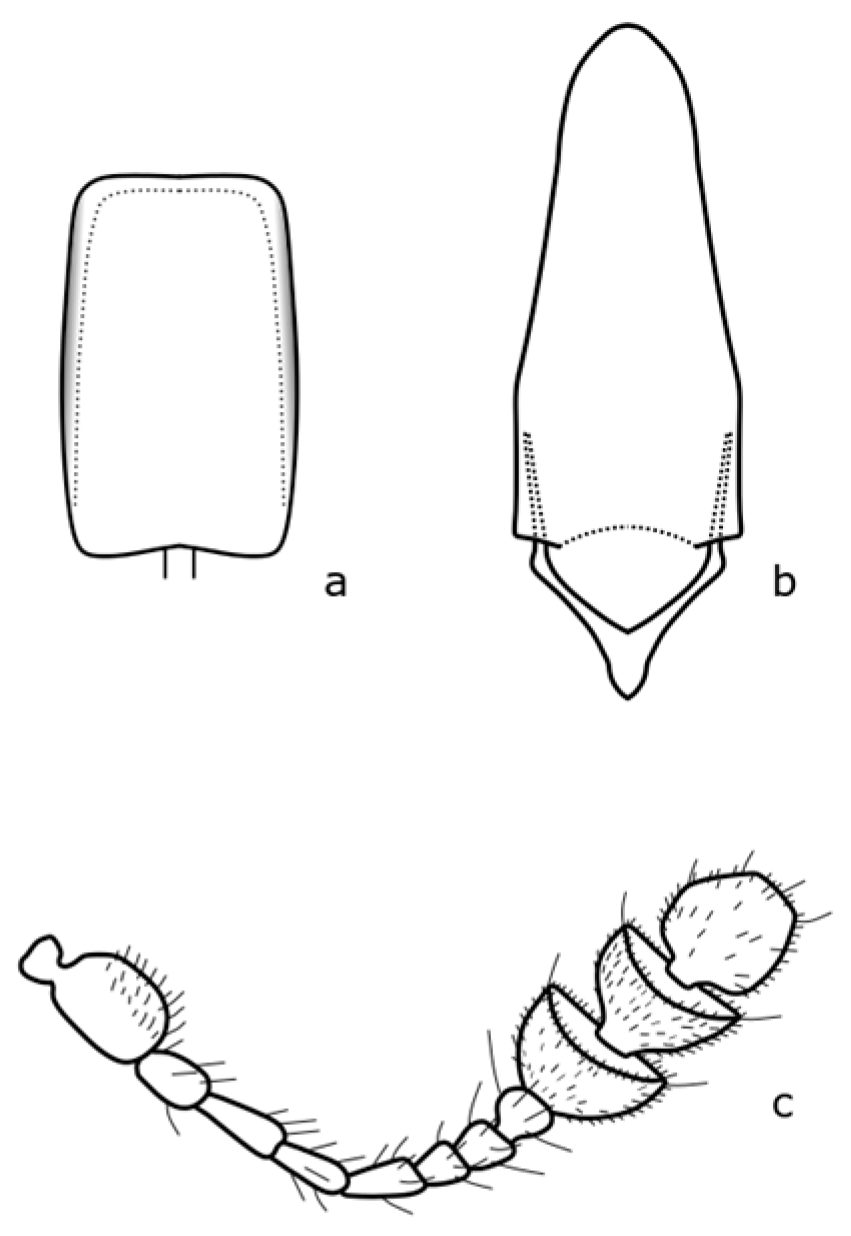
Brassicogethes anthracinus (C. Brisout de Barneville, 1863) adults are 1.6–2.6 mm long, black, with dense fine punctuation on the pronotum and elytra, rather matte [
8]. The distinguishing features are shown in
Figure A3. This species is associated with
Isatis tinctoria L. by its larval development [
8,
21]. This species occurs mainly in dry grassland stands, dry meadows, and also in disturbed habitats [
19].
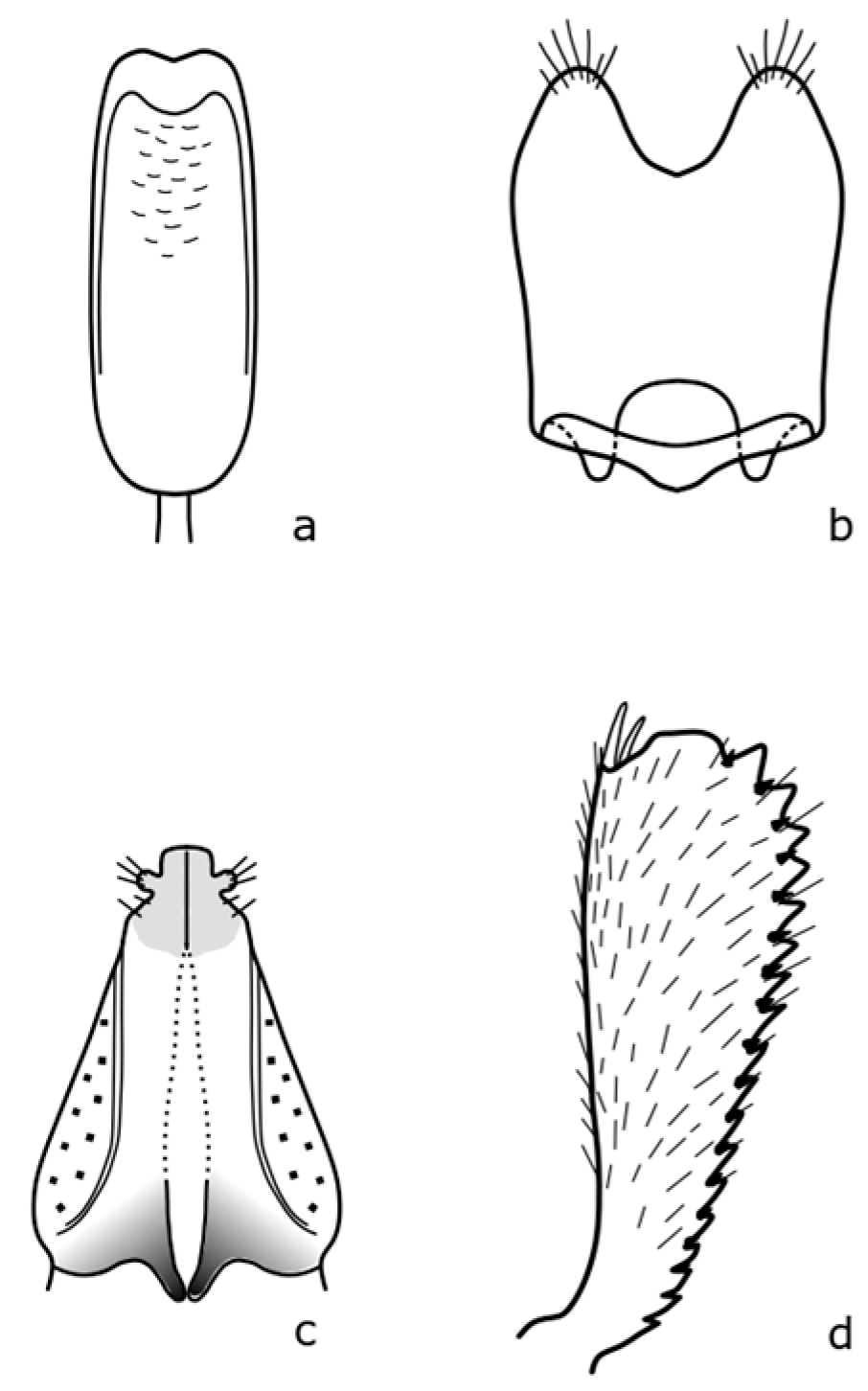
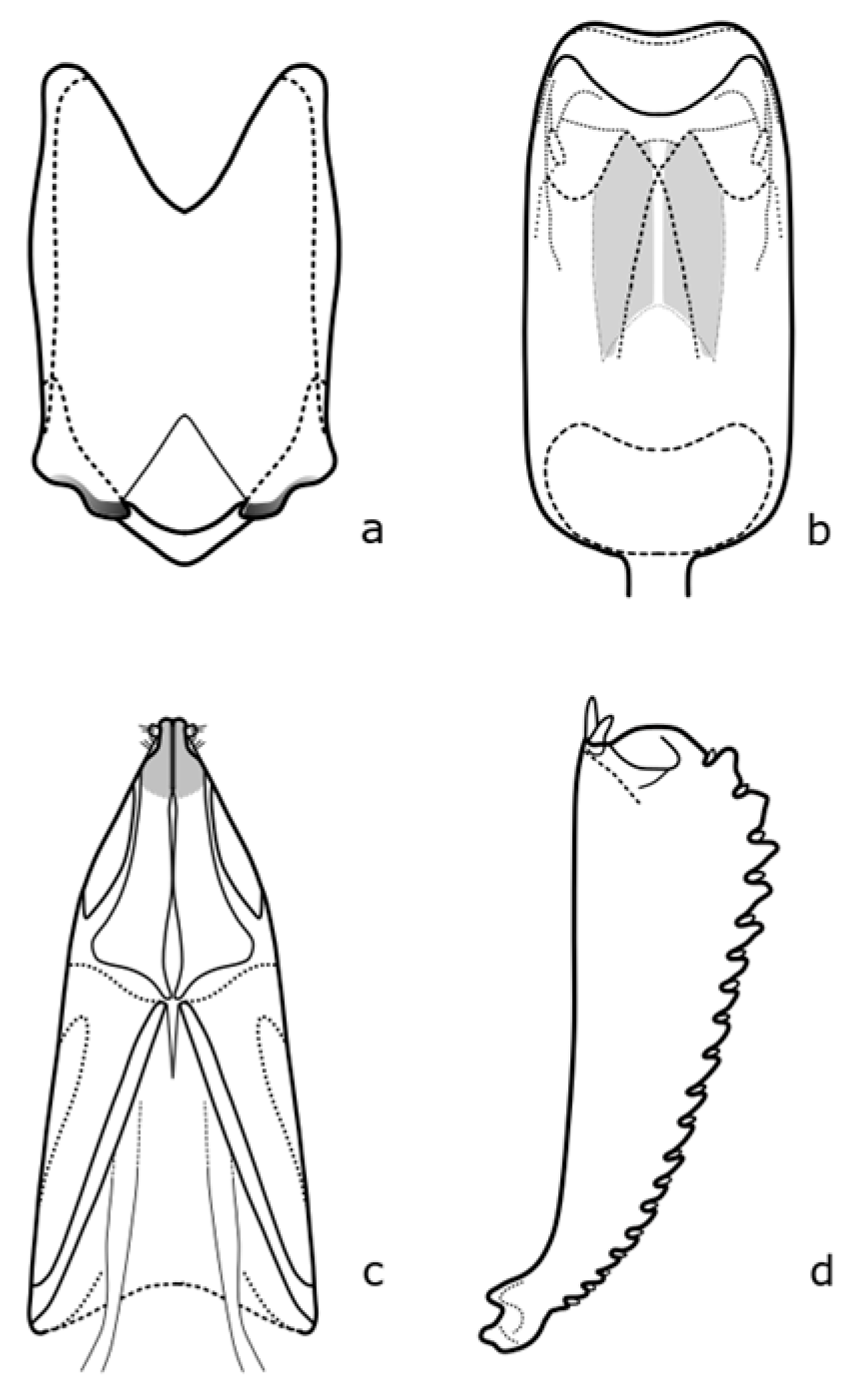
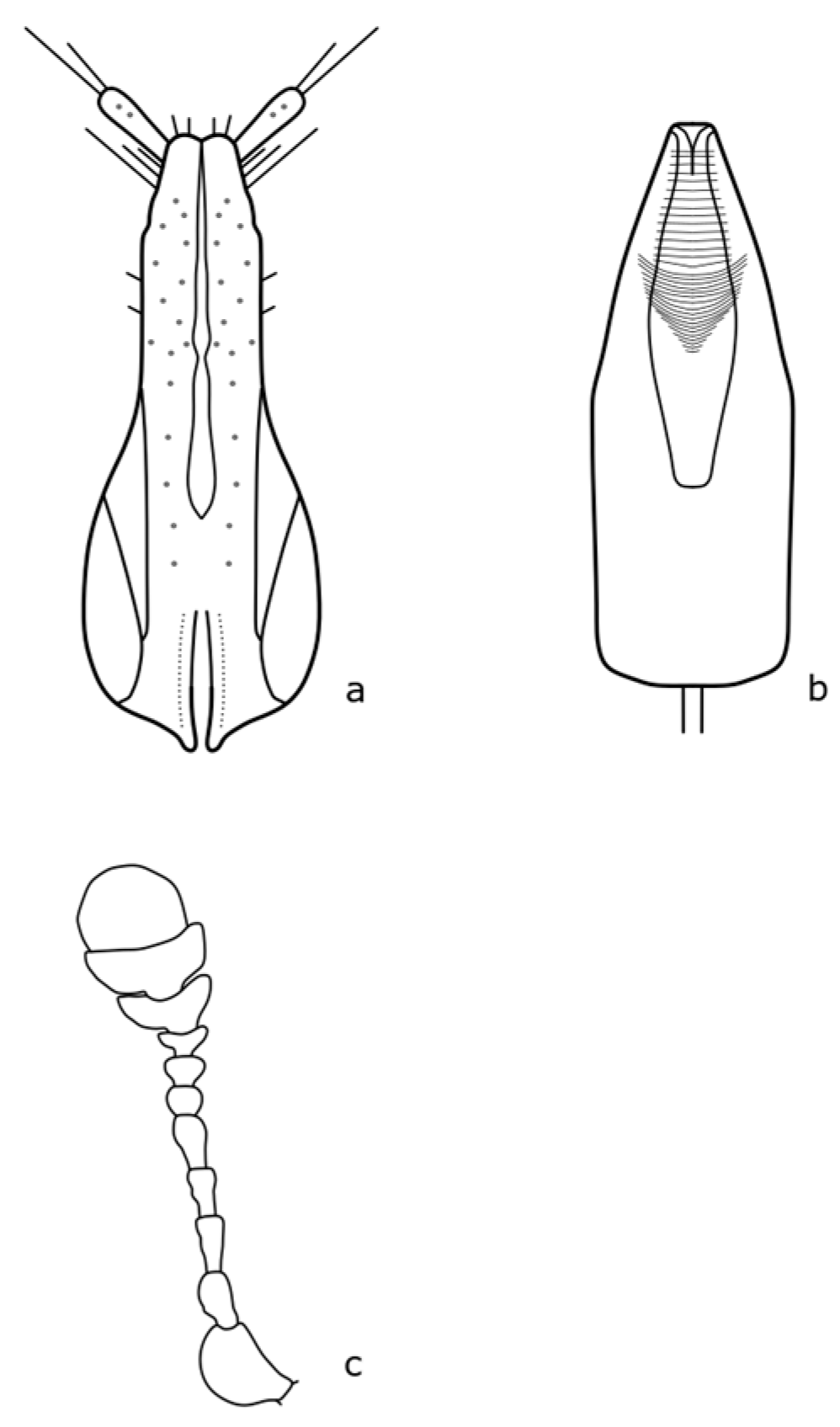
Brassicogethes coeruleovirens (Forster, 1849) is a species whose bionomy is not completely known yet. The adult is 1.8–2.5 mm long, oval-shaped, and black with a blue or blue-green metallic shine [
8]. Distinguishing features are shown in
Figure A4.
Brassicogethes coracinus (Sturm, 1845) occurs in xerothermic habitats [
30], fields, meadow edges, river banks, and shady stands [
10]. Adults are 1.5–2.7 mm long [
8]. The body is black or black-brown, never green, blue, or violet metallic shine [
10]. The distinguishing features are shown in
Figure A5. In the Brassicaceae family, it feeds on
Barbarea spp.,
Sisymbrium spp.,
Brassica spp., and
Sinapis spp. [
10,
11]. The adults were found on
Sisymbrium loeselli L.,
Rorippa austriaca (Crantz) Besser, and also on species of the Apiaceae family [
30]. Larval development takes place on
Brassica spp.,
Sinapis spp., and
Sisymbrium spp. Audisio et al. [
31] and Kurochkin [
32] show only
S. loeselli as the larval food plant. The simultaneous presence of
B. coracinus and
B. aeneus adults on plants of the Rosaceae family is reported by Kurochkin [
32].
B. coracinus is common in central and northern Europe [
10], and Mancini et al. [
10] report that it occurs throughout Europe, Turkey, Central Asia, and Siberia. Differentiation between
B. coracinus and
B. subaeneus is difficult due to the genital similarity of both species [
31].
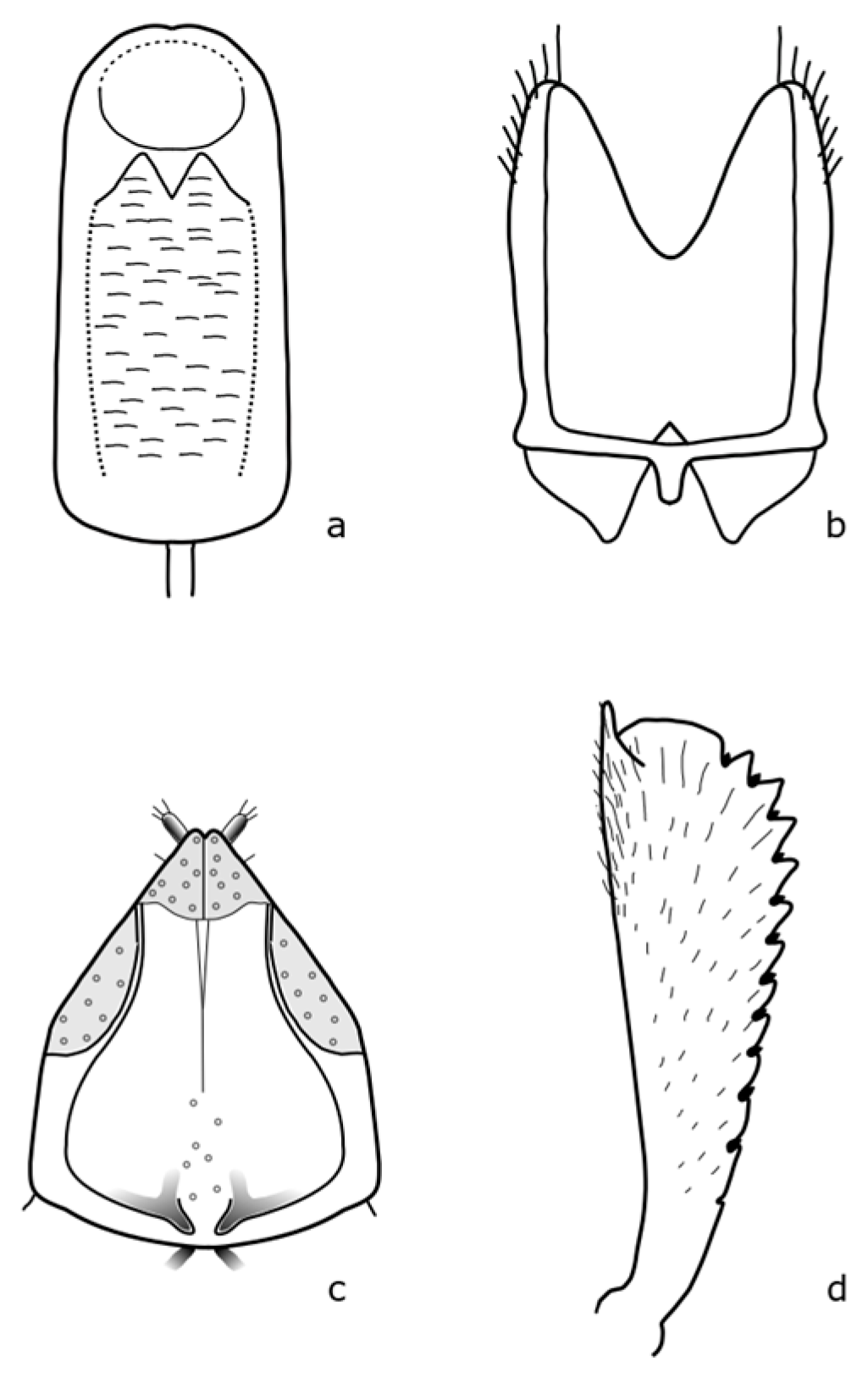
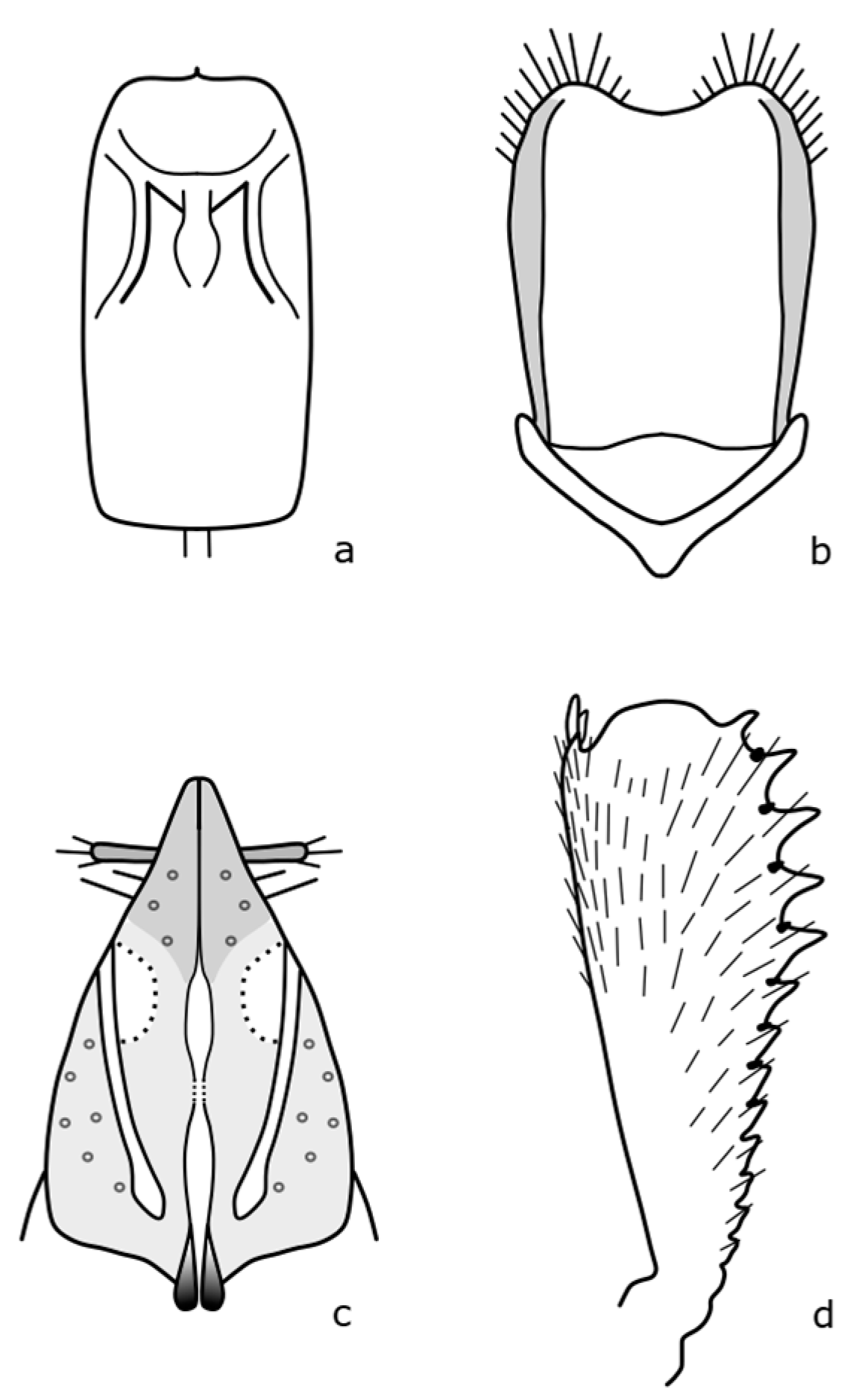
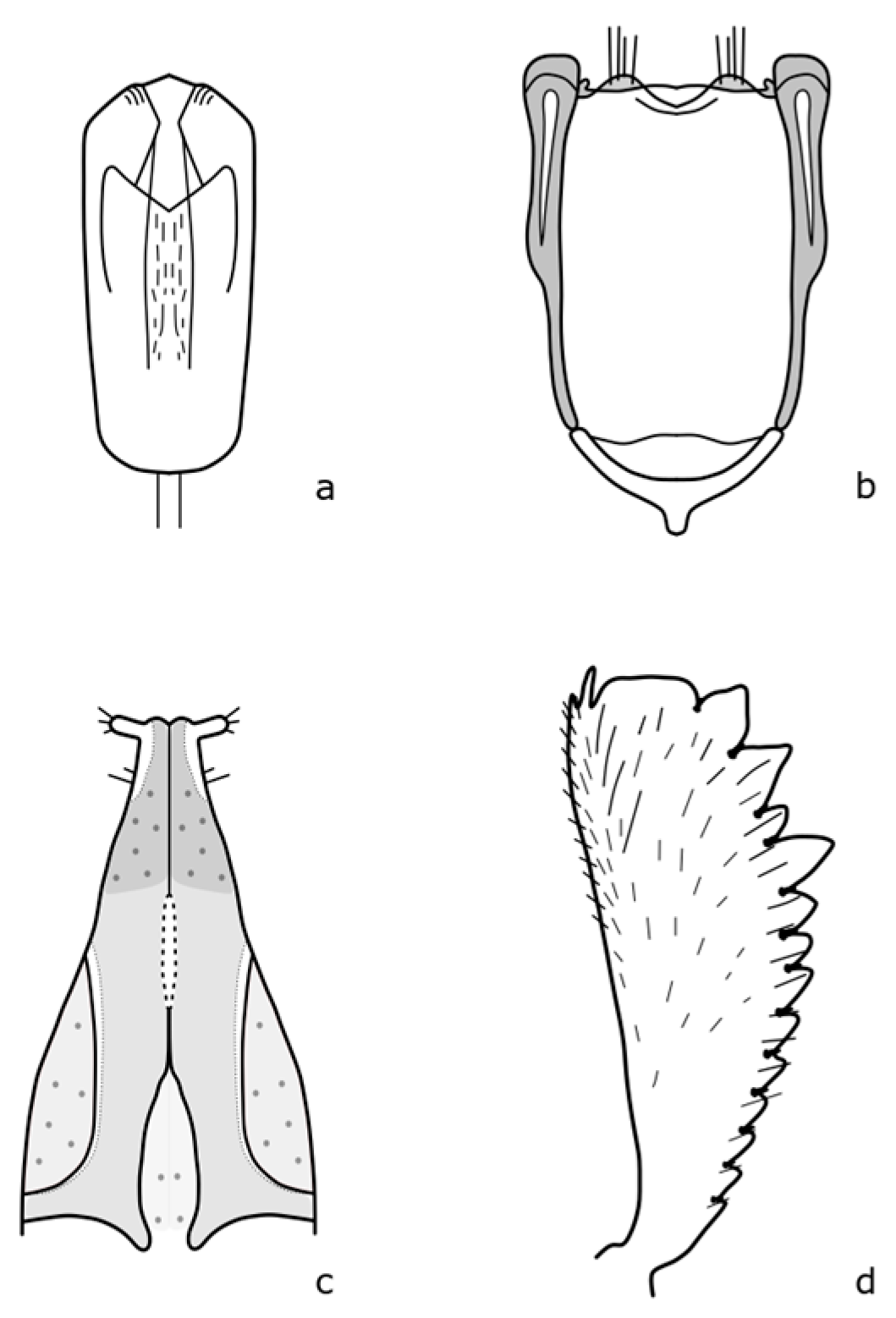
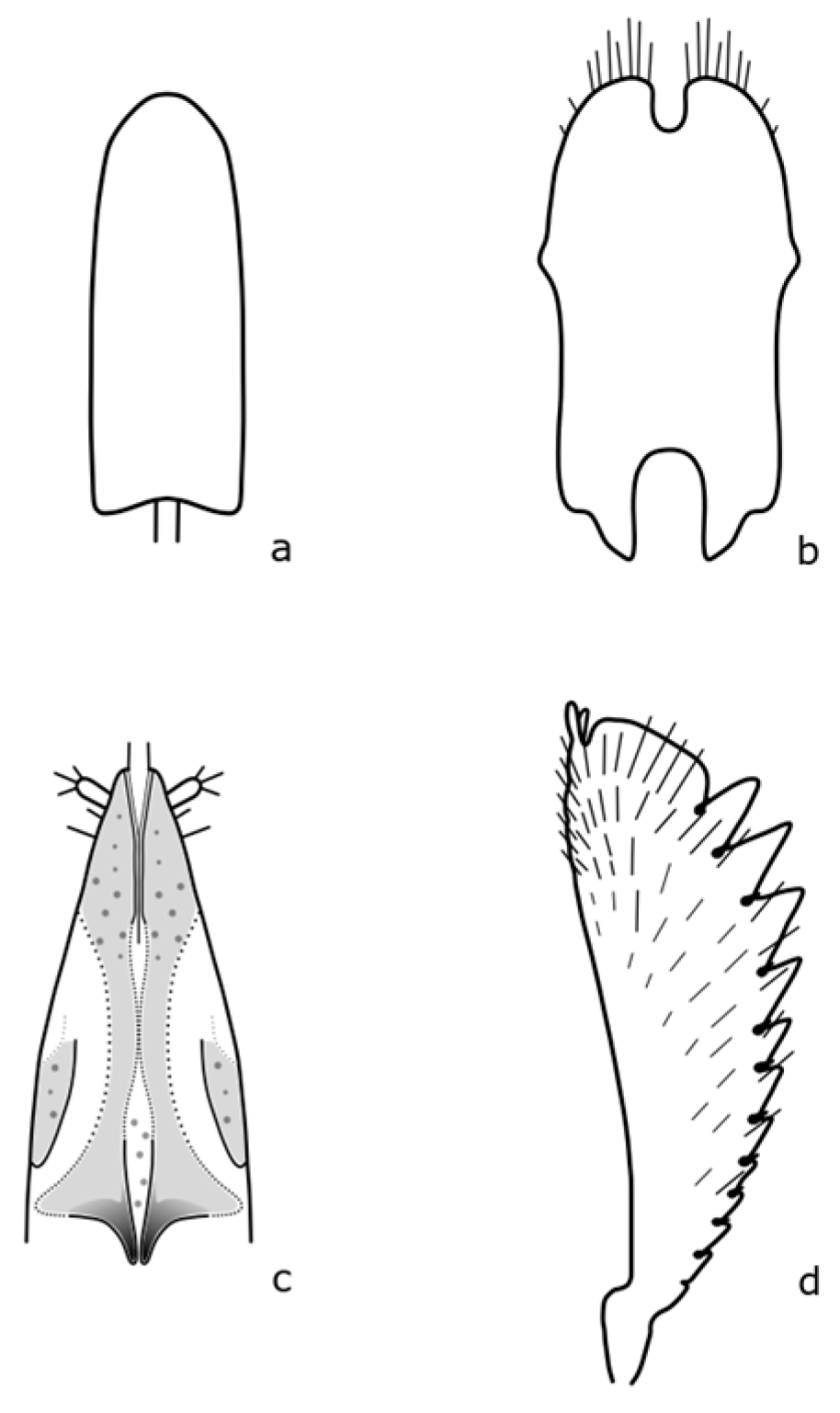
Brassicogethes czwalinai (Reitter, 1871) adults are 2.3–3.1 mm long and habitually very similar to
B. viridescens. The anterior tibia is widest at the level of the third tooth [
8]. The distinguishing features are shown in
Figure A6. This species occurs in mountain and foothill areas and the larvae develop on
Lunaria rediviva L. [
8].
Brassicogethes subaeneus (Sturm, 1845) adults are 1.8–2.2 mm long. The body is oval-shaped, and black with a green metallic shine. The legs are dark brown, with fine teeth on the front leg's tibia [
8]. The distinguishing features are shown in
Figure A7. Larvae feed on
Cardamine species [
31],
Cardaminopsis species [
33], especially
C. arenosa (L.) Hayek [
10,
34]. Adults are found on the flowers of various plants. According to Lasoń [
30], they are found on
Armoracia rusticana P. Gaertn., B. Mey. & Scherb.,
Geranium spp.,
Prunus spp.,
Filipendula ulmaria (L.) Maxim.,
Sorbus aucuparia L.,
Oenothera biennis L., and flowers of the Apiaceae family. It is abundant in lower and middle altitudes [
34]. This species is considered to inhabit xerothermic habitats and is widespread, occurring in northern, central, and eastern Europe [
10].
Brassicogethes matronalis (Audisio & Spornraft, 1990). The imago of this species is 2.6 mm long and dark brown [
35]. It is similar to
B. subaeneus [
36]. During its development, larvae are associated with
Hesperis matronalis L. [
31]. It is similar in appearance to
B. subaeneus [
36]. The distinguishing features are shown in
Figure A8.
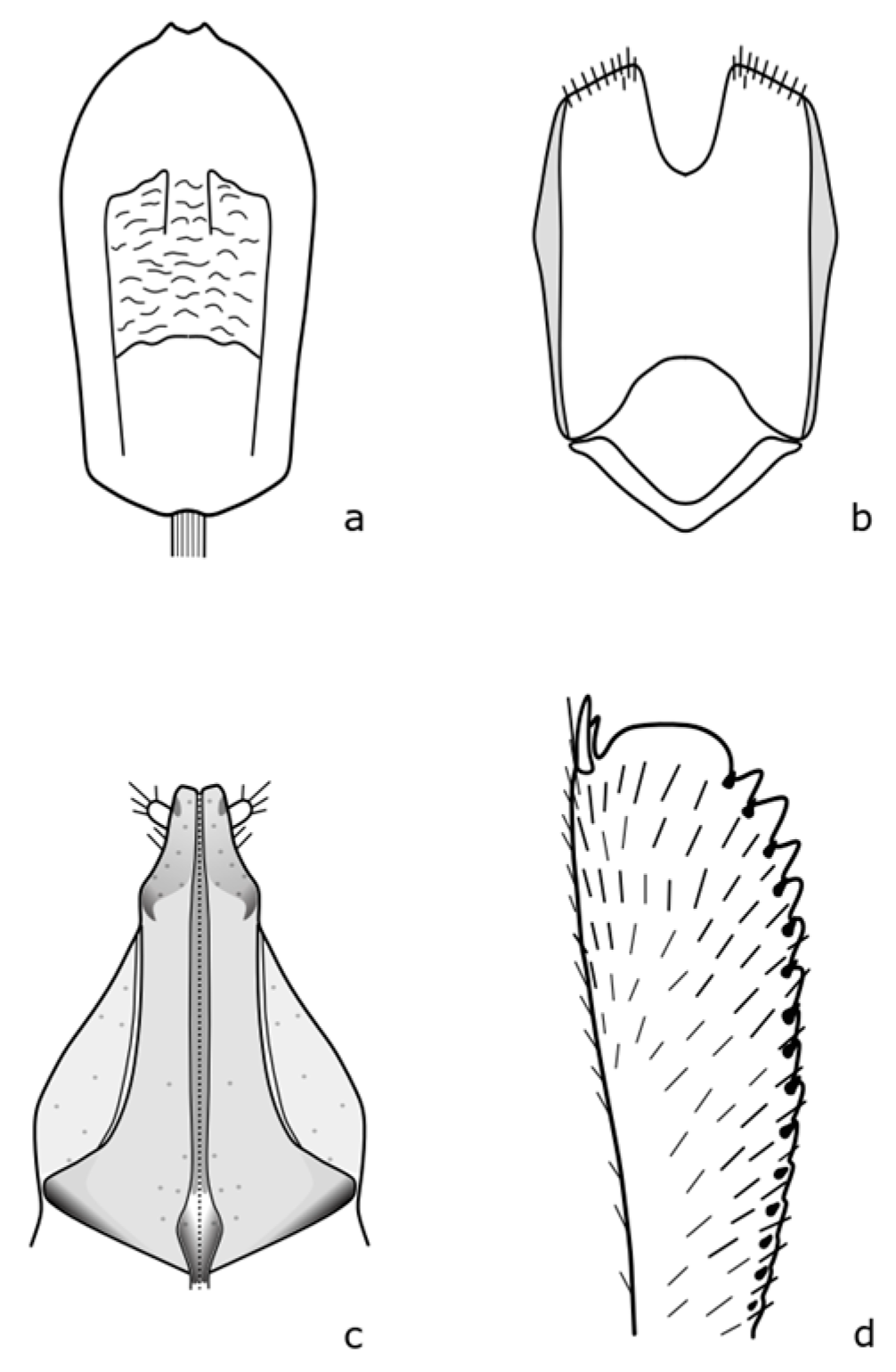
Clypeogethes lepidii (Miller, 1852) is a fine-haired black beetle with a body length of 1.2–2.2 mm; antennae and legs are dark brown to black [
8]. The distinguishing features are shown in
Figure A9. The larvae develop on
Lepidium draba L. [
8];
Alyssum spp.,
Aurinia spp.,
Moricandia spp.,
Hesperis spp., and
Mathiola spp. Baviera and Audisio [
21] show it on
Isatis tinctoria L. It occurs mainly in lowland meadows [
19].
Boragogethes symphyti (Heer, 1841) adults are 2.2–3.0 mm long, the body is convex, oval-shaped, dark blue, rarely dark green colored, with brown legs, the front legs are lighter [
8]. The distinguishing features are shown in
Figure A10. This species is associated with plants of the Boraginaceae family, particularly
Symphytum [
7]. According to Nunberg [
8], larvae develop on
Symphytum officinale L.
The
Cychramus luteus (Fabricius 1787) adult body is rusty brown; the pronotum and elytra with hairs in visible rows. [
35]. The distinguishing features are shown in
Figure A11. The larvae of this species develop in fungi of the Agaricaceae and Lycoperdaceae families, they are often present in species of the genus
Armillaria and
Lycoperdon, and less often, in fungi of the class Ascomycetes [
37]. According to Rimšaité [
38] and Schigel [
39], adults are raised from larvae present in
Armillaria spp. Jelínek [
34] states that larvae develop in the fruiting bodies of softer wood-destroying fungi and adults are present on flowers, especially of
Aruncus dioicus (Walter) Fernald. It is abundant in Europe, adults feed on the pollen on the flowers of plants around the beehives [
40]. Neumann and Ritter [
41] reported finding adults in hives. However,
C. luteus (Fabricius, 1787) did not leave holes in the cells of the honeycomb, and neither oviposition nor the presence of larvae on the combs was detected [
41].
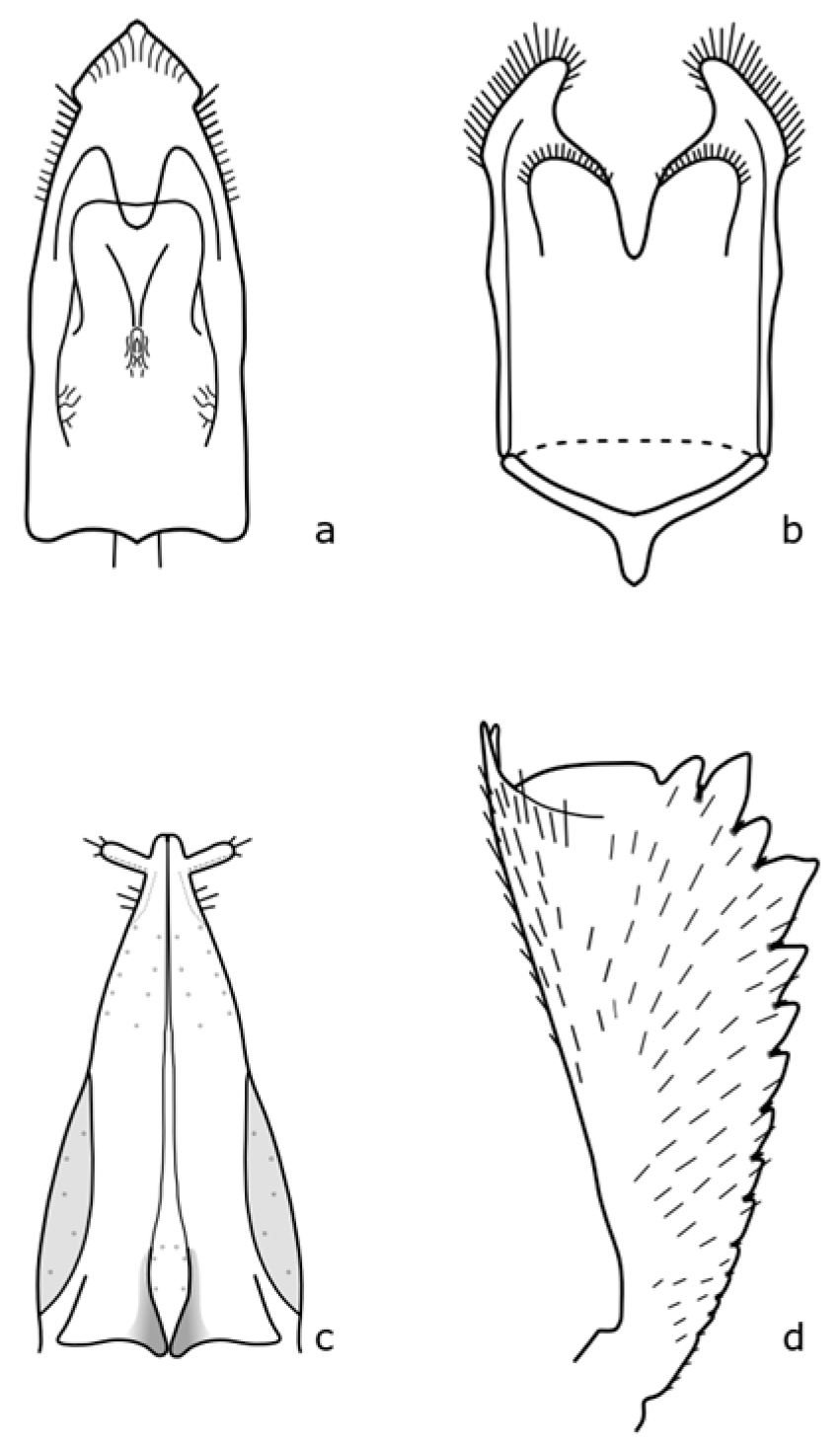
Epurea aestiva (Linnaeus, 1758) is 3 mm long, and rusty yellow [
35]. The distinguishing features are shown in
Figure A12. This species is widespread throughout Europe, where adults are found on plant flowers at the edge of forests [
21] and they are abundant in deciduous forests [
34]. Kurochkin [
32] observed adults most frequently on woody plants of the family Rosaceae—
Cerasus fruticosa Pall.,
Malus domestica (Suckow) Borkh.,
Prunus avium L.,
Rosa majalis Herrm.,
Filipendula ulmaria and
F. vulgaris Moench, rarely on
Knautia arvensis (L.) J.M. Coult. Majka and Klimaszewski [
42] consider adults to be sap-feeders. Larvae develop in the nests of bumblebees and small mammals [
19,
34] and probably also on moldy substrates [
34]. As noted by Arbogast et al. [
40], the larvae are mycetophagous. This species is widespread in Europe, Asia, and the USA, and it has also been found in Canada [
42].
Fabogethes nigrescens (Stephens, 1830) adults are 1.7–2.5 mm long; black with fine punctuation on pronotum and elytra; antennae and the legs are pitch brown; and the first, second, and fourth teeth of forelegs tibia are bigger [
8,
11]. The distinguishing features are shown in
Figure A13. The larvae are associated with plants of the Fabaceae family, in particular
Trifolium spp. [
43],
Onobrychis spp.,
Ononis spp.,
Lotus spp. [
5],
Medicago spp. [
44]. Niezgodziński [
45] considers it to be an insignificant pest of
Vicia faba L. in Poland. It is found in open habitats [
46]. It is an important pollinator of
Primula [
47,
48] and
Rubus [
29]. Mifsud and Audisio [
44] consider it to be a pest of medicinal plants in North America.
Genistogethes carinulatus (Förster, 1849) is 1.4–2.2 mm long, with black antennae and brown legs, and the forelegs tibia are relatively narrow [
8]. The distinguishing features are shown in
Figure A14. This species is associated with
Lotus corniculatus L. by their larval development [
11,
21]. It is mainly found in meadows [
19]. Niezgodziński [
45] considers it to be an insignificant bean pest. Duke [
48] considers it to be a pest of
Onobrychis viciifolia Scop.
Lamiogethes atramentarius (Förster, 1849). The imago is 2.0–2.8 mm long, oval-shaped, and black with metallic blue or green shine. Pronotum and elytra are finely punctuated. Antennae and legs are reddish-yellow. Tibiae of the foreleg are finely toothed [
8]. The distinguishing features are shown in
Figure A15. The nutrient plants of larvae are
Galeobdolon luteum Huds.,
Lamium album L., and
Galeopsis spp., adults are present on the flowers of various woody plants, e.g.,
Crataegus spp. and
Prunus padus L. [
49].
Meligethes atratus (Olivier, 1790) is 2.4–3.8 mm long and brownish-black. The first antennal segment is darker [
8]. The distinguishing features are shown in
Figure A16. It occurs at the edges of forests, road ditches, and grassy areas. Audisio et al. [
19] stated its presence in disturbed habitats. Its larvae develop on species of the genus
Rosa, including
R. canina L. [
11]. According to Audisio et al. [
7], its larval development is strictly associated with the genus
Rosa spp. and
Rubus spp. It most likely also develops on
Prunus spp.,
Pyrus spp., and
Crataegus spp. [
6]. Adults are polyphagous [
11] and it is widespread throughout Europe [
8,
50].
The
Pria dulcamare (Scopoli, 1763) imago is 1.6–1.8 mm long and rusty brown. Its body is egg-shaped or oval, less convex, elytra are shortened and 1–3 abdominal tergites are visible. The fourth tarsal segment is always shorter than the others [
10,
11]. This species is associated with the
Solanum dulcamara L. [
8,
11,
51,
52], as well as with
S. nigrum L. [
11] during its larval development. Adults are also found on flower species other than Solanaceae [
44].
The
Sagitogethes maurus (Sturm, 1845) imago is 2.0–3.0 mm long, black colored, the legs and antennae are dark brown, and the distal part of the tibia of the front legs is distinctly widened [
8]. The distinguishing features are shown in
Figure A17. All species of this genus are associated with plants of the family Lamiaceae [
7]. This species is associated with
Salvia spp. [
43]. Adults are found on the flower of
Salvia pratensis L.,
S. verticillata L.,
Sambucus,
Taraxacum, and
Potentilla [
49].
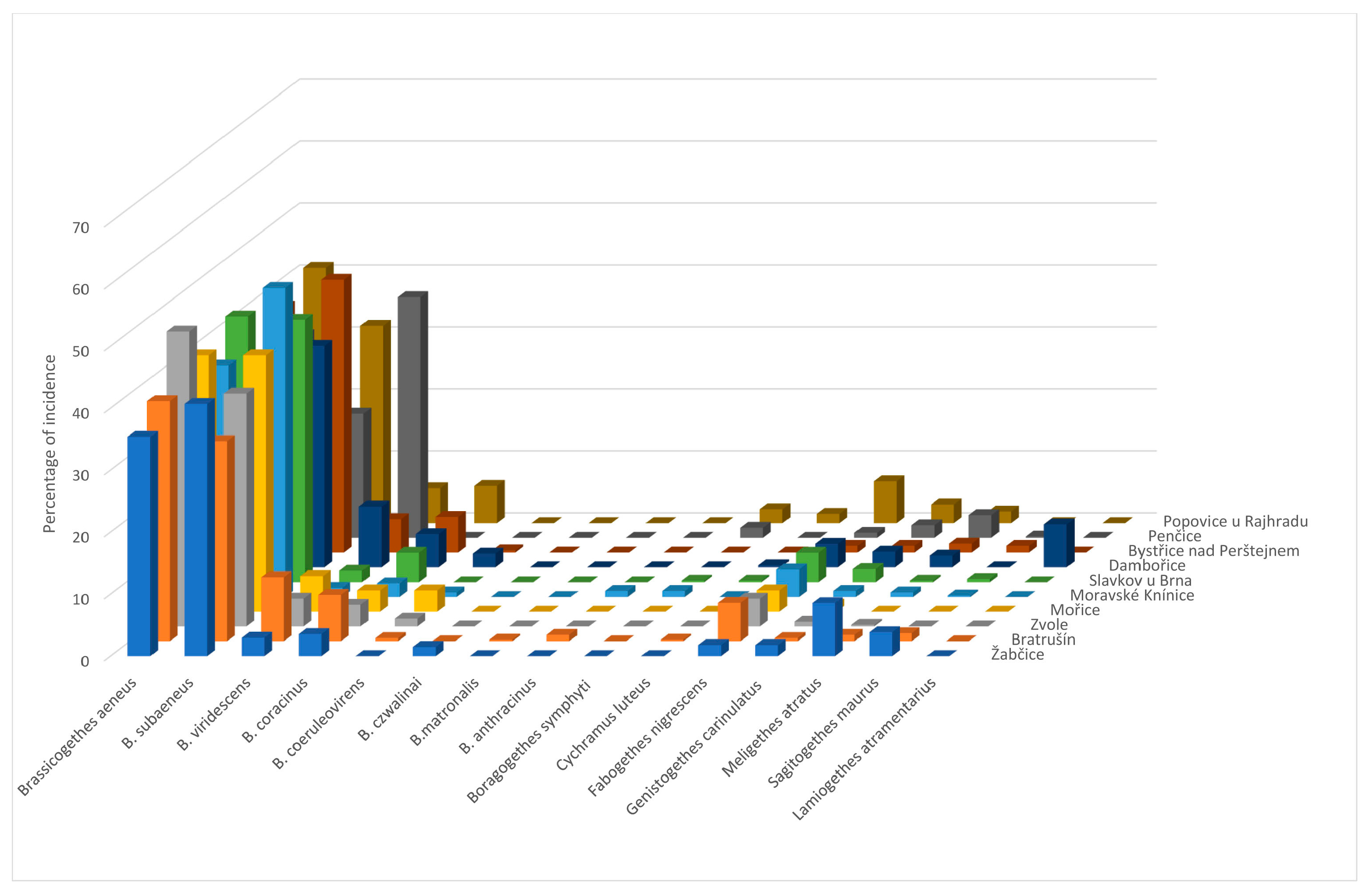
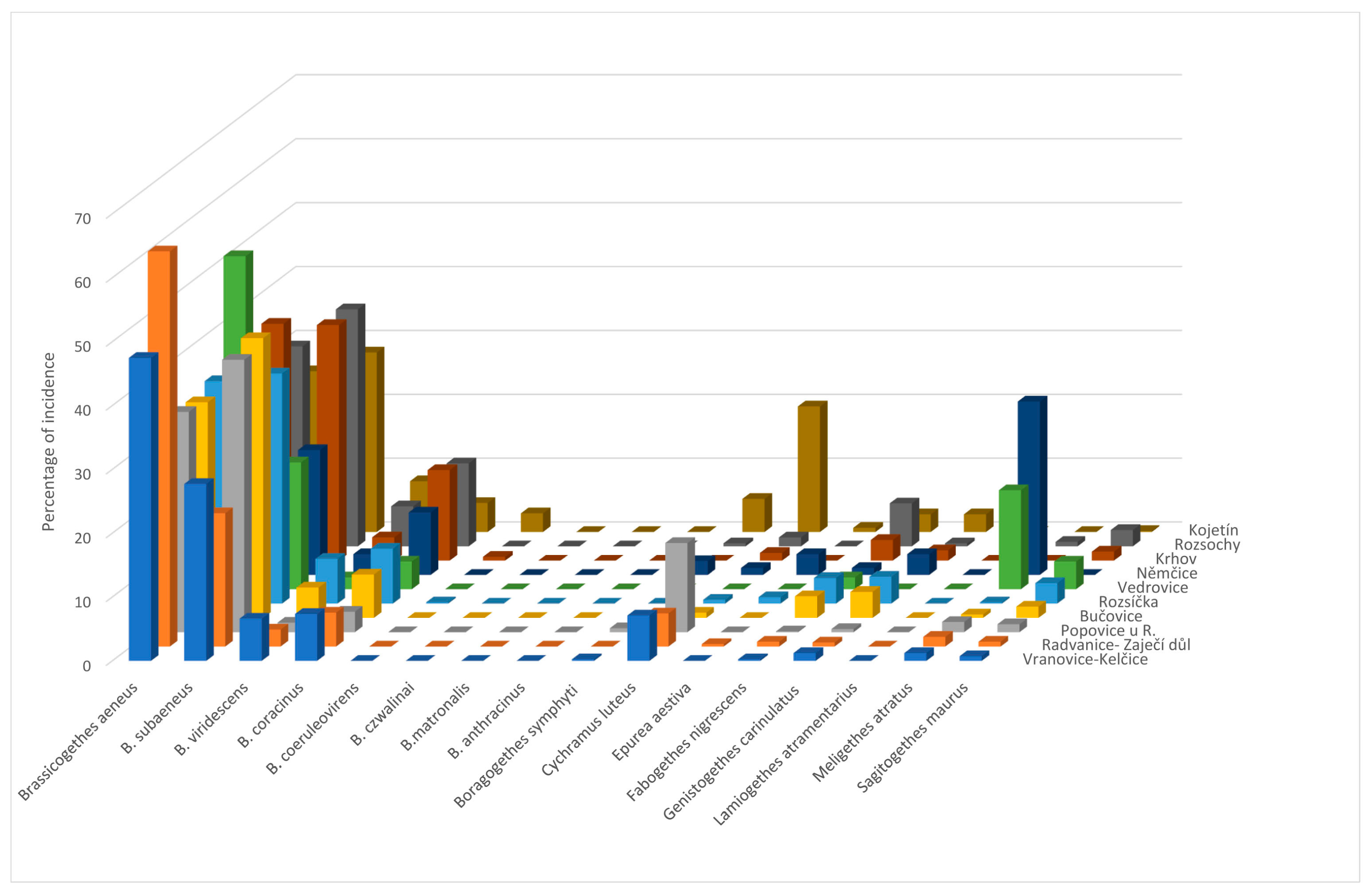
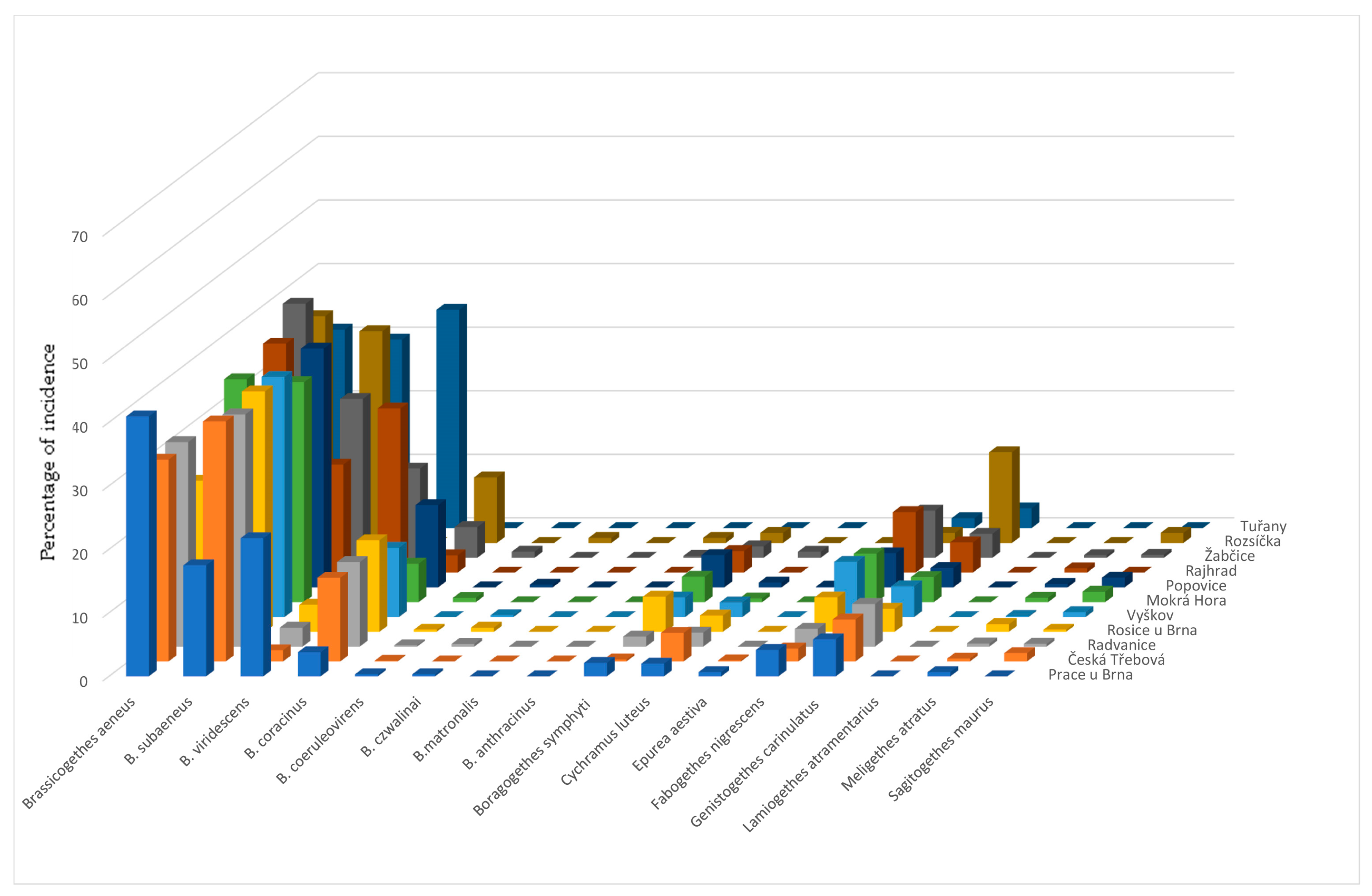
The main contribution of this work was the identification of the species spectrum of the genus Brassicogethes present in oilseed rape stands and its representation in the samples. Determining the proportion of B. aeneus and B. viridescens in the samples was particularly important. The results could help to refine the control of pollen beetles in OSR crops with insecticides, increase their effectiveness, and reduce the selection pressure inducing resistance to insecticides in pollen beetle populations. Until now, only a few authors have dealt with this issue and the available information is not very comprehensive.
4. Discussion
Brassicogethes aeneus (Fabricius, 1775) is considered a key pest of oilseed rape and turnip rape (
B. rapa) in Europe [
4]; another significant species is
B. viridescens (Fabricius, 1787), which is harmful in both Europe and North America. Not only is
B. aeneus present in oilseeds, but other species are also common [
62,
63,
64]. To our knowledge, few studies have focused on the species spectrum and abundance of individual pollen beetle species in oilseed rape. In Croatia Gotlin Čuljak and Juran [
5] dealt with the species spectrum of pollen beetles in oilseed rape, they reported the presence of
Brassicogethes aeneus,
B. viridescens,
B. coracinus (Sturm, 1845),
Clypeogethes lepidii (L. Miller, 1851), and
Fabogethes nigrescens (Stephens, 1830).
B. aeneus was the eudominant species [
5], the same results we obtained in 2011 and 2012. Interestingly,
Clypeogethes lepidii was not present in any of the samples we analyzed. The reason for this remains unknown as we were unable to find any relevant information.
B. aeneus and
B. viridescens are among the most common species in the UK [
3,
9], both these species are also considered to be important in Sweden [
65]. In Estonia,
B. viridescens is very common in oilseed rape, more so in winter oilseed rape [
66]. As stated by Finch et al. [
67], both species mentioned above can also damage cauliflower and broccoli, reducing their market value. On the other hand, none of the listed pollen beetle species are known to be a pest of cruciferous vegetables in the Czech Republic. The species composition of pollen beetle populations present in oilseed rape fields after overwintering was studied in Hungary by Marczali [
68], who found
Brassicogethes aeneus in all collections,
B. coracinus was in 78%,
B. viridescens in 50%,
M. picipes (Sturm, 1845) in 58%,
M. nigrescens (Stephens, 1830) in 29%,
Sagitogethes maurus (Sturm, 1845) in 43%,
M. atratus (Olivier, 1790) in 21%,
Odontogethes denticulatus (Heer, 1841) in 14% and
Genistogethes carinulatus (Förster, 1849) in 7% of collections. Although
M. picipes and
M. nigrescens have been incorrectly reported as two species [
68], they are in fact only one species—
Fabogethes nigrescens [
59]. As in the work of Marczali [
68],
B. aeneus was present in 100% of our collections in 2011–2013. The situation is slightly different for other species.
B. coracinus was present in 90% of the samples in 2011 and 2013 and in 100% in 2012.
B. viridescens was present in 100% of samples in 2011 and 2012 and in 90% of samples in 2013,
Fabogethes nigrescens was present in 100% of samples in all the years observed.
Sagitogethes maurus (Sturm, 1845) was present in 60% of samples in 2011, 90% in 2012, and 80% in 2013;
M. atratus was present in 90% of samples in 2011, 80% in 2012, and 90% in 2013.
Genistogethes carinulatus was present in 100% of samples in 2011 and 2013, and in 90% of samples in 2012.
Meligethes denticulatus was absent in all samples during subsequent years. In their next study, Marczali and Keszthelyi [
69] reported the presence of four pollen beetle species—
M. aeneus,
M. coracinus,
M. viridescens and
F. nigrescens in oilseed rape in Hungary. Thieme et al. [
70] reported the presence of
B. viridescens,
F. nigrescens,
B. coeruleovirens (Förster, 1849), and
B. coracinus (Sturm, 1845). When studying the migration of pollen beetles from hibernacula to oilseed rape fields, Juhel et al. [
71] found 99%
B. aeneus and the presence of only two individuals of
B. viridescens, they did not specify the species affiliation of the remaining proportion.
In other European countries, several pollen beetle species are found in oilseed rape fields like in the Czech Republic [
62,
63,
64].
Brassicogethes aeneus is the eudominant species in the Czech Republic, but in some localities,
B. subaeneus is also eudominant [
62]. The treatment of oilseed rape against pollen beetles is based on reaching some of the economic threshold values. All specimens of pollen beetles present in oilseed rape are considered to be
Brassicogethes aeneus (Fabricius, 1775). However, a whole complex of pollen beetles present in oilseed rape has been identified.
One of the first evaluations of the species spectrum of pollen beetles was carried out by us in 2010 [
71], where
B. aeneus (Fabricius, 1775) was found to be a eudominant species.
B. subaeneus (Sturm, 1845),
B. viridescens (Fabricius, 1787), and
B. erythropus (Marsham, 1802) were also eudominant species in several localities as well. In most cases,
B. subaeneus and
B. viridescens were dominant or subdominant species. In the following years, pollen beetle populations were again analyzed to determine their species composition. Several pollen beetle species were present in the samples collected from 2011 to 2013 in oilseed rape in different regions of the Czech Republic. The eudominant species were usually
B. aeneus,
B. subaeneus, and
B. viridescens. These results largely correspond to those obtained in subsequent years [
62,
64].
In reality, the presence of individual species is not taken into account, although the common occurrence of several pollen beetle species in oilseed rape fields has long been known, e.g., [
72,
73]. The determination of individual species is very time-consuming, and, with a few exceptions, requires the preparation of male and female genitalia. Knowledge of the species spectrum of pollen beetles and their bionomy could be one of the components of anti-resistance strategies. The degree of resistance of pollen beetles to insecticide active ingredients is investigated for population samples containing different species. Some authors claimed that it is not essential to know the level of resistance of other species [
74]. Others, however, have considered this issue in more detail.
B. viridescens is not resistant to insecticides [
72]. According to Spaar et al. [
22],
B. aeneus is the most common species in cruciferous oilseeds fields, followed by
B. viridescens,
B. coracinus (Sturm, 1845), and
Fabogethes nigrescens (Stephens, 1830). According to this author, only a few single specimens of
B. coeruleovirens (Förster, 1849),
Meligethes atratus (Olivier, 1790),
Lamiogethes bidens (C. Brisout de Barneville, 1882),
Sagittogethes maurus (Sturm, 1849),
Meligethes flavimanus (Stephens, 1830), and
Lamiogethes pedicularius (Gyllenhal, 1808) were found.
Brassicogethes aeneus is the dominant species together with
B. subaeneus and
B. viridescens. The genus
Brassicogethes was represented in the collections from 81.8 to 90.5% of the pollen beetle population. A similar representation is also mentioned by some authors, e.g., Ouvrard et al. [
3], where the representation of
B. aeneus is 95%. In Lithuania, the representation is 98.2% to 98.8% [
75]. In Romania, Talmaciu and Talmaciu [
76] examined the fauna of pollen beetles on oilseed rape, and they reported only
B. aeneus. Up to 1% of
Brassicogethes coracinus and
Fabogethes nigrescens were present in growth, as observed by Ouvrard et al. [
3]. The presence of these species in spring and winter oilseed rape growths in Lithuania is reported—0.15%
B. coracinus and 0.04%
F. nigrescens [
77]. In the populations collected from winter oilseed rape fields in the Czech Republic between 2011 and 2013,
B. coracinus was represented by 4.21%, 7.72%, and 8.41%, (sorted by years ascending) and
F. nigrescens was 3.75%, 2.42%, and 5.11%, 5.45 (sorted by years ascending).
Brassicogethes viridescens occurs on all yellow-flowering cruciferous plants, but it is not as abundant as
B. aeneus [
11]. As stated by Derron et al. [
72], its relative representation in the stands is highly variable. Ouvrard et al. [
3] stated its representation in oilseed rape stands at 4%. Makūnas [
77] showed its representation (average from winter and spring oilseed rape) at 0.35%, but later Makūnas [
75] performed a genetic analysis of pollen beetles in Lithuania and stated the representation of this species in spring oilseed rape fields at 1.8%. The proportion of
B. viridescens in our collections was 8.66% in 2011, 4.55% in 2012, and 10.51% in 2013. In 2012, it is close to the value that is reported by Derron et al. [
72]. Overall, however, our results confirm the variability reported by Derron et al. [
72]. In Spain, the presence of
B. aeneus,
B. viridescens,
B. coeruleovirens, and
B. coracinus in oilseed rape fields was reported by Moradillo [
78].
The presence of adult
Meligethes atratus in oilseed rape stands is probably due to the frequent occurrence of rose hips, a nutrient plant for larvae of this species, at field margins. In our collections,
M. atratus was represented by 1.97% in 2011, 4.85% in 2012, and 0.48% in 2013. Pollen beetles actively fly into oilseed rape stands, but they also use the wind to occupy fields [
79]. The pollen beetles can fly up to 13.5 km and can actively reach a distance of up to 300 m [
80]. They are often more abundant at the edges of stands. They are also the first to be colonized by pollen beetles [
81]. Pollen beetles are more abundant on the windward side of the field [
82] and near overwintering sites [
71]. The main reason for the presence of pollen beetle species associated with their development on non-cruciferous plants in oilseed stands is sufficient pollen as a food source for adults. Furthermore, as described by Ouvrard et al. [
3], they can also occur here randomly if they can be transported over relatively long distances by air currents.
5. Conclusions
Several Brassicogethes species occur in oilseed rape stands. Among them, B. aeneus (Fabricius, 1775) is the eudominant species in all the sampling sites during all the years monitored. Other eudominant species, but with isolated fluctuations in abundance in some localities and some years, are B. subaeneus (Sturm, 1845), B. viridescens (Fabricius, 1787), and B. coracinus (Sturm, 1845). All species of this genus are associated with cruciferous plants by their larval development.
Thus, as our collection analysis shows, the pollen beetle species associated with cruciferous plants clearly predominated in individual years (representation in the collections was 83.7–93.4%), of which more than half were B. aeneus and B. viridescens. The pollen beetle species, whose larvae are not associated with the Brassicaceae family by their development, were represented in the collections between 9.6% and 16.3%.
From the point of view of protection of oilseed rape against pollen beetles, it is possible to ignore the fact that there are several species of pollen beetles and to work with this complex genus Brassicogethes as one species. To determine the critical number of beetles in the stand for protection purposes, it is not necessary to distinguish between pollen beetle species. On the other hand, when pollen beetle populations sampled in distinct localities are tested on susceptibility to insecticides, it would be good to know the differences in the species composition among them.
The presence of accompanying species can be beneficial, as they host some antagonists of harmful pollen beetle species, such as parasitoids, and can also be a reservoir for insect pathogens, thus indirectly contributing to bioregulation.
At the same time, as B. aeneus is treated with insecticides, other pollen beetle species present in the stand are also affected. However, as in the case of B. aeneus or the Brassicogethes species complex, it can be assumed that resistance to the insecticides used is also selected in the associated species.
Nowadays, it is important to monitor the spectrum of pollen beetle species in oilseed rape fields, but also in mustard or poppy fields. In particular, it provides early detection of harmful species of the Nitidulidae family, such as Aethina tumida Murray, 1867, an invasive species harmful to beehives. Species monitoring is also important from an ecological point of view, as it allows us to detect the effects of insecticides on non-target species and to take timely measures against the negative effects of pesticides.



 ,
, 




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}