
Assessment of Bioactive Profile of Sorghum Brans under the Effect of Growing Conditions and Nitrogen Fertilization

 ,
, 
 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
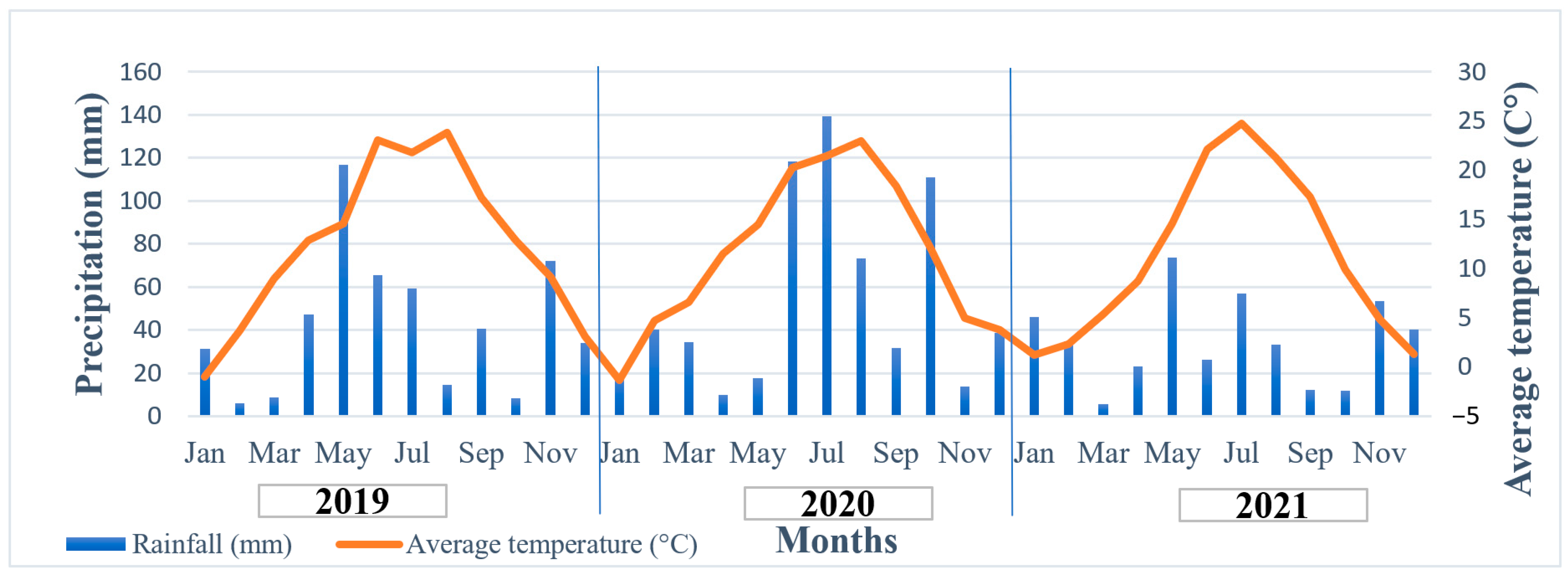
2.2. Properties of Experimental Site and Weather Conditions
2.3. Chemical Reagents
2.4. Preparation of Sorghum Bran
2.5. Measurement of Total Phenolic Content
2.6. Measurement of Condensed Tannins
2.7. Antioxidant Content and Capacity
2.8. Statistical Analysis
3. Results
3.1. Statistical Analysis
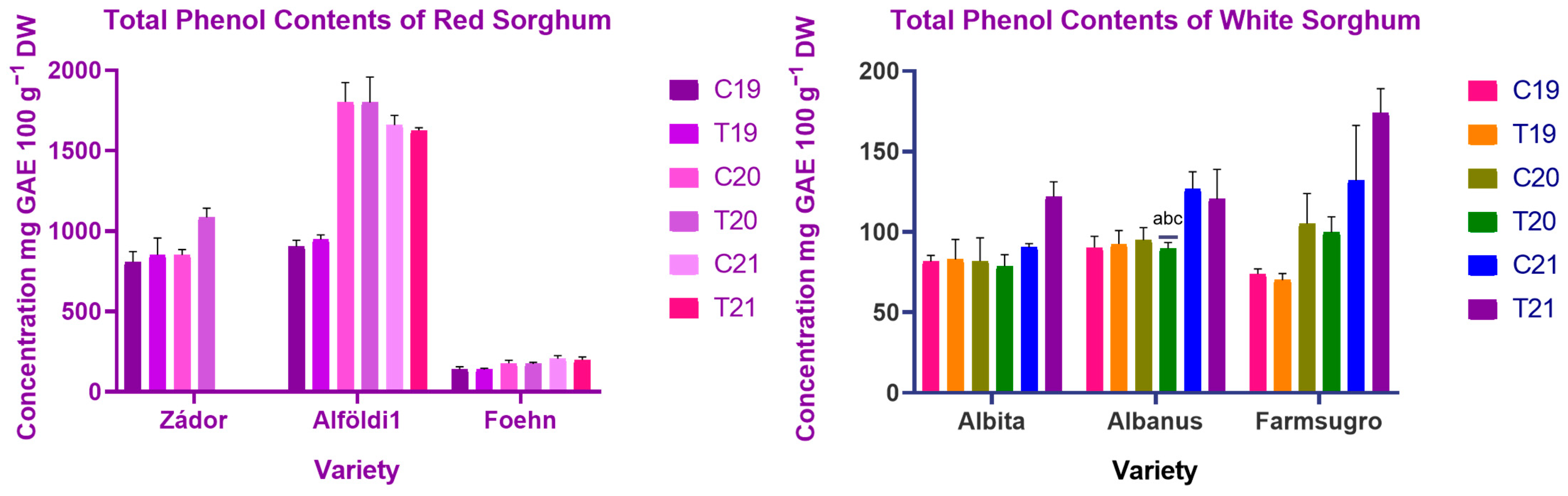
3.2. Evaluation of Polyphenol Content of Sorghum Brans
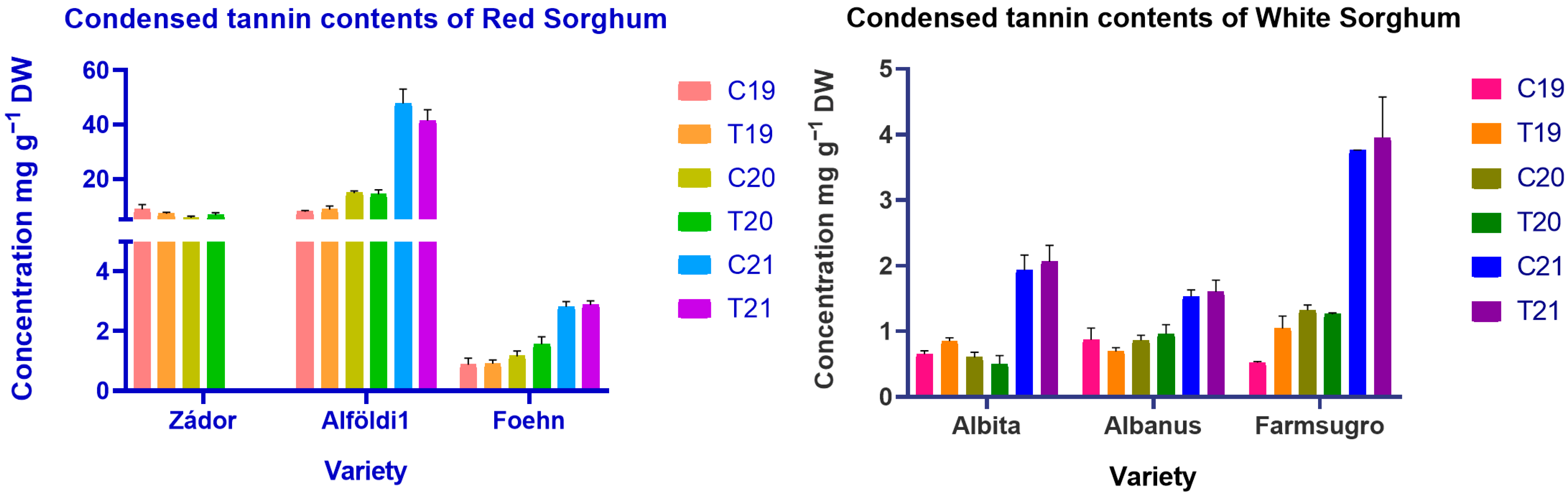
3.3. Estimation of Condensed Tannin Content and Differences of Brans
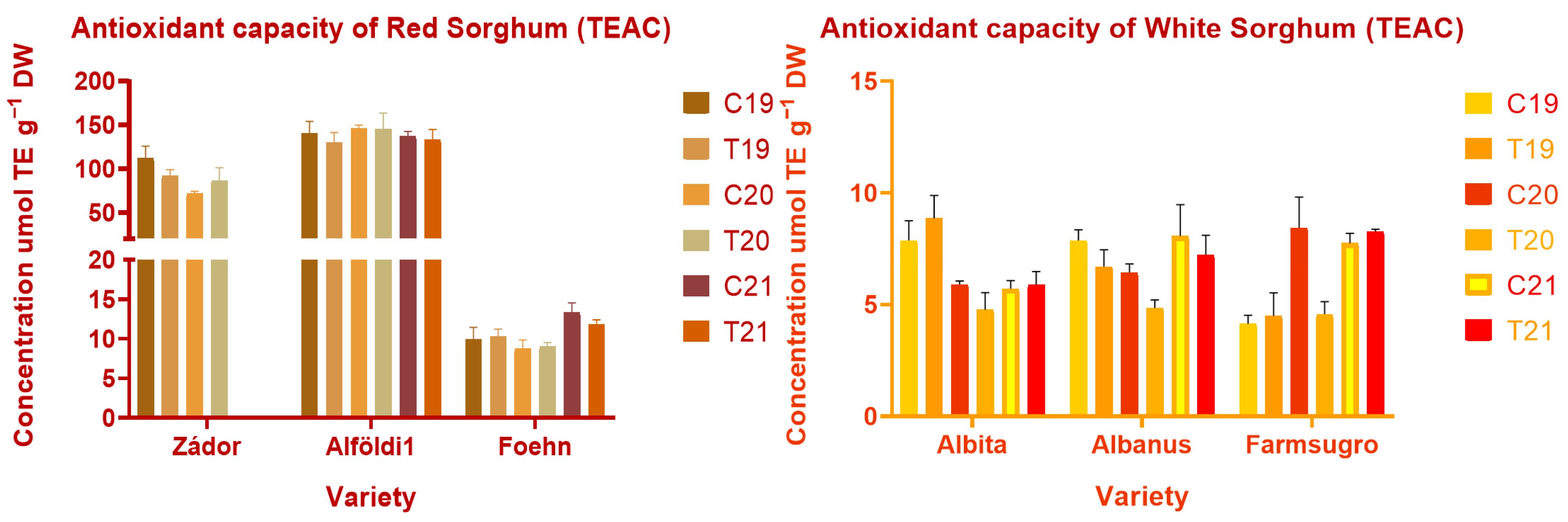
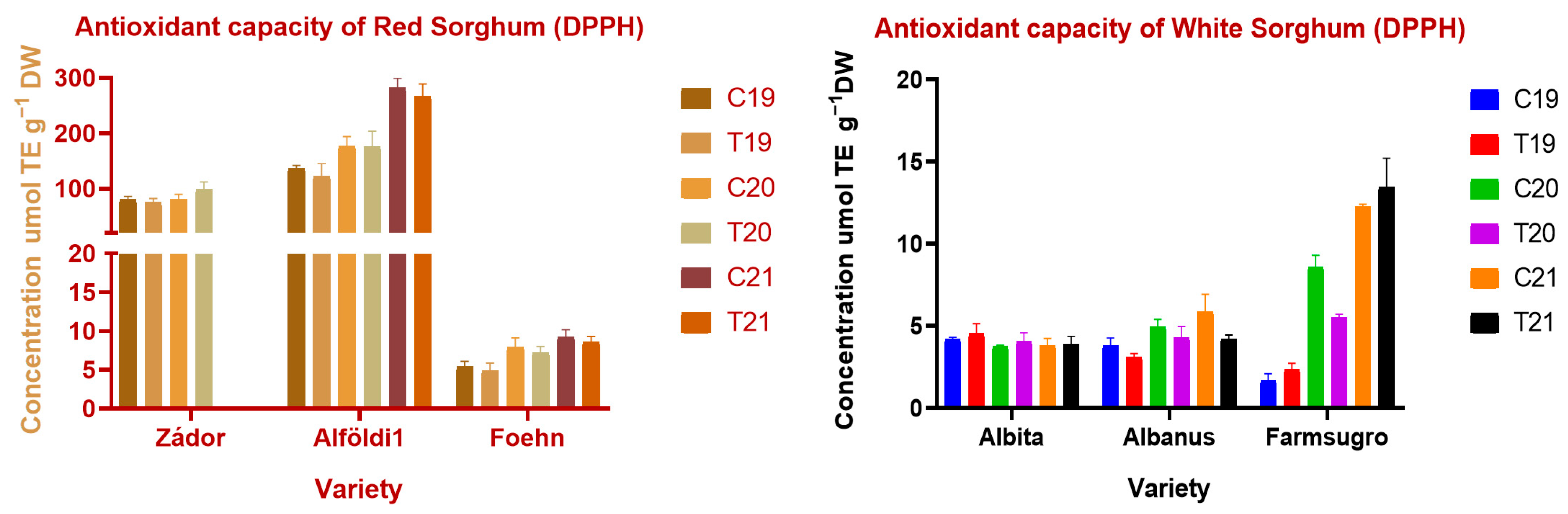
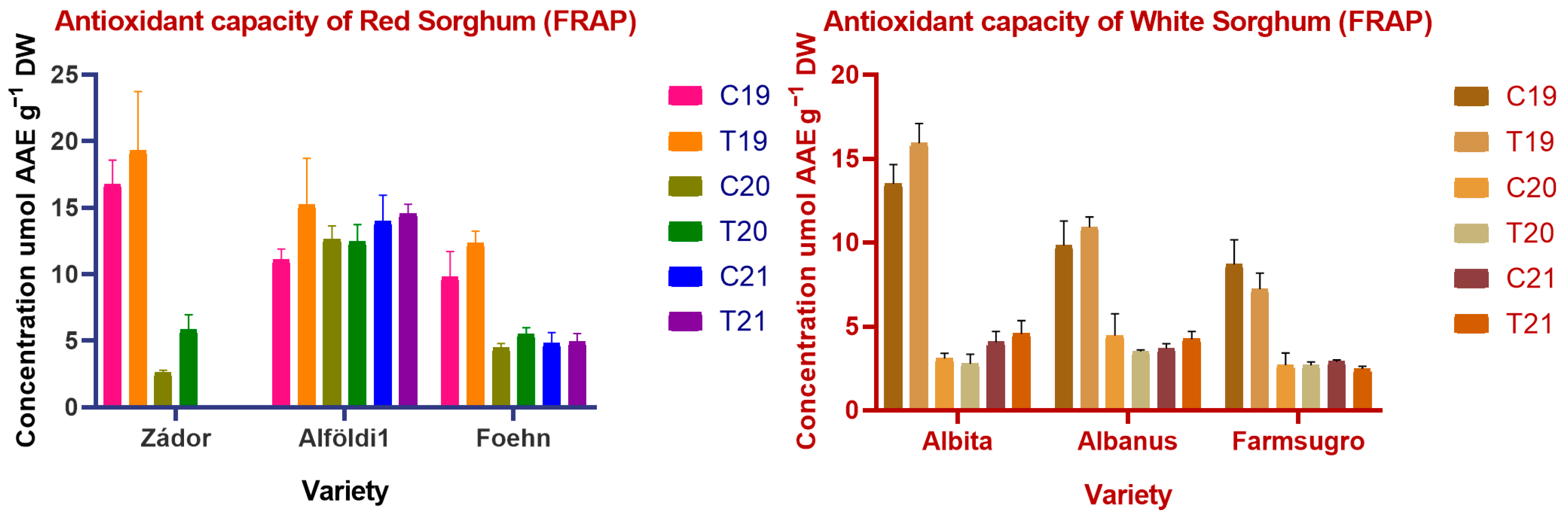
3.4. Antioxidant Properties of Sorghum Brans
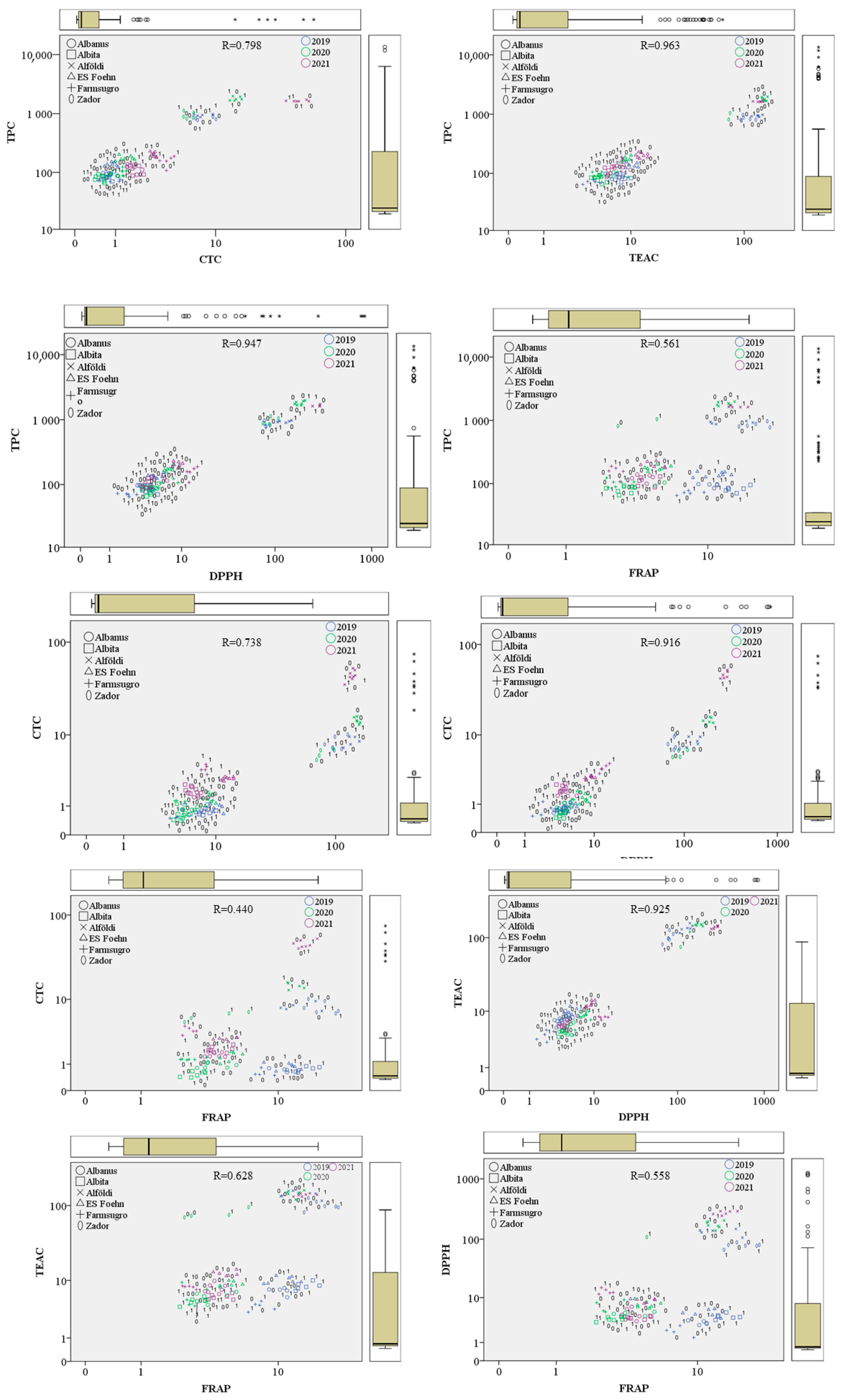
3.5. Connections between the Antioxidant Properties, Tannin, and Polyphenol Content of Sorghum Grains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Villanueva-Cañongo, C.; Hernández-Carlos, B. Antioxidant Compounds and Their Antioxidant Mechanism. In Antioxidants; Shalaby, E., Brozozowsky, T., Eds.; IntechOpen: London, UK, 2019; ISBN 978-1-78923-920-1. [Google Scholar]
- Cetin Cakmak, K.; Gülçin, İ. Anticholinergic and Antioxidant Activities of Usnic Acid-an Activity-Structure Insight. Toxico. Rep. 2019, 6, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.C.; Lee, J.Y.; Jang, M.G.; Song, H.; Kim, S.-J. Seasonal Variations in the Phenolic Compounds and Antioxidant Activity of Sasa Quelpaertensis. Ind. Crops Prod 2018, 122, 506–512. [Google Scholar] [CrossRef]
- Toydemir, G.; Gultekin Subasi, B.; Hall, R.D.; Beekwilder, J.; Boyacioglu, D.; Capanoglu, E. Effect of Food Processing on Antioxidants, Their Bioavailability and Potential Relevance to Human Health. Food. Chem. X 2022, 14, 100334. [Google Scholar] [CrossRef]
- Apak, R.; Calokerinos, A.; Gorinstein, S.; Segundo, M.A.; Hibbert, D.B.; Gülçin, İ.; Çekiç, S.D.; Güçlü, K.; Özyürek, M.; Çelik, S.E.; et al. Methods to Evaluate the Scavenging Activity of Antioxidants toward Reactive Oxygen and Nitrogen Species (IUPAC Technical Report). Pure Appl. Chem. 2022, 94, 87–144. [Google Scholar] [CrossRef]
- Al-juhaimi, F.; Ghafoor, K.; Özcan, M.M.; Jahurul, M.H.A.; Babiker, E.E.; Jinap, S.; Sahena, F.; Sharifudin, M.S.; Zaidul, I.S.M. Effect of Various Food Processing and Handling Methods on Preservation of Natural Antioxidants in Fruits and Vegetables. J. Food Sci. Technol. 2018, 55, 3872–3880. [Google Scholar] [CrossRef]
- Chen, P.; Bornhorst, J.; Diana Neely, M.; Avila, D.S. Mechanisms and Disease Pathogenesis Underlying Metal-Induced Oxidative Stress. Oxid. Med. Cell Longev. 2018, 2018, 7612172. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef]
- Awika, J.M.; Rose, D.J.; Simsek, S. Complementary Effects of Cereal and Pulse Polyphenols and Dietary Fiber on Chronic Inflammation and Gut Health. Food Funct. 2018, 9, 1389–1409. [Google Scholar] [CrossRef]
- Wu, G.; Bennett, S.J.; Bornman, J.F.; Clarke, M.W.; Fang, Z.; Johnson, S.K. Phenolic Profile and Content of Sorghum Grains under Different Irrigation Managements. Food Res. Int. 2017, 97, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Liu, C.; Jun, J.H. Metabolic Engineering of Anthocyanins and Condensed Tannins in Plants. Curr. Opin. Biotechnol. 2013, 24, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Giada, M.d.L.R. Food Phenolic Compounds: Main Classes, Sources and Their Antioxidant Power. In Oxidative Stress and Chronic Degenerative Diseases—A Role for Antioxidants; Morales-Gonzalez, J.A., Ed.; IntechOpen: London, UK, 2013; ISBN 978-953-51-1123-8. [Google Scholar]
- Awika, J.M. Chapter 3—Sorghum: Its Unique Nutritional and Health-Promoting Attributes. In Gluten-Free Ancient Grains; Taylor, J.R.N., Awika, J.M., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition: Sawston, UK, 2017; pp. 21–54. ISBN 978-0-08-100866-9. [Google Scholar]
- Borrell, A.; van Oosterom, E.; George-Jaeggli, B.; Rodriguez, D.; Eyre, J.; Jordan, D.J.; Mace, E.; Singh, V.; Vadez, V.; Bell, M.; et al. Chapter 5—Sorghum. In Crop Physiology Case Histories for Major Crops; Sadras, V.O., Calderini, D.F., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 196–221. ISBN 978-0-12-819194-1. [Google Scholar]
- Cornejo-Ramírez, Y.I.; Martínez-Cruz, O.; Toro-Sánchez, C.L.D.; Wong-Corral, F.J.; Borboa-Flores, J.; Cinco-Moroyoqui, F.J. The Structural Characteristics of Starches and Their Functional Properties. CyTA—J. Food 2018, 16, 1003–1017. [Google Scholar] [CrossRef]
- Rashwan, A.K.; Yones, H.A.; Karim, N.; Taha, E.M.; Chen, W. Potential Processing Technologies for Developing Sorghum-Based Food Products: An Update and Comprehensive Review. Trends Food Sci. Technol. 2021, 110, 168–182. [Google Scholar] [CrossRef]
- Scherf, K.A. Immunoreactive Cereal Proteins in Wheat Allergy, Non-Celiac Gluten/Wheat Sensitivity (NCGS) and Celiac Disease. Curr. Opin. Food Science 2019, 25, 35–41. [Google Scholar] [CrossRef]
- de Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, Bioactive Compounds, and Potential Impact on Human Health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef]
- Hossain, S.; Islam, N.; Rahman, M.; Mostofa, M.G.; Khan, A.R. Sorghum: A Prospective Crop for Climatic Vulnerability, Food and Nutritional Security. J. Agric. Food Res. 2022, 8, 100300. [Google Scholar] [CrossRef]
- Rumler, R.; Bender, D.; Schönlechner, R. Sorghum and Its Potential for the Western Diet. J. Cereal Sci. 2022, 104, 103425. [Google Scholar] [CrossRef]
- Przybylska-Balcerek, A.; Frankowski, J.; Stuper-Szablewska, K. Bioactive Compounds in Sorghum. Eur. Food Res. Technol. 2019, 245, 1075–1080. [Google Scholar] [CrossRef]
- Frankowski, J.; Przybylska-Balcerek, A.; Stuper-Szablewska, K. Concentration of Pro-Health Compound of Sorghum Grain-Based Foods. Foods 2022, 11, 216. [Google Scholar] [CrossRef]
- Bean, S.R.; Wilson, J.D.; Moreau, R.A.; Galant, A.; Awika, J.M.; Kaufman, R.C.; Adrianos, S.L.; Ioerger, B.P. Structure and Composition of the Sorghum Grain. In Sorghum; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2019; pp. 173–214. ISBN 978-0-89118-628-1. [Google Scholar]
- Li, Z.; Zhao, X.; Zhang, X.; Liu, H. Bioactive Compounds and Biological Activities of Sorghum Grains. Foods 2021, 10, 2868. [Google Scholar] [CrossRef]
- Nagy, R.; Szőllősi, E.; Molnár, P.B.; Murányi, E.; Czimbalmos, R.; Sipos, P. Condensed Tannin Content and Antioxidant Activity of Hungarian Sorghum Varieties Grown at Research Institute in Karcag. Acta Agrar. Debr. 2021, 11, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Ky, I.; Le Floch, A.; Zeng, L.; Pechamat, L.; Jourdes, M.; Teissedre, P.-L. Tannins. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 247–255. ISBN 978-0-12-384953-3. [Google Scholar]
- Jiang, Y.; Zhang, H.; Qi, X.; Wu, G. Structural Characterization and Antioxidant Activity of Condensed Tannins Fractionated from Sorghum Grain. J. Cereal Sci. 2020, 92, 102918. [Google Scholar] [CrossRef]
- Macáková, K.; Kolečkář, V.; Cahlíková, L.; Chlebek, J.; Hošťálková, A.; Kuča, K.; Jun, D.; Opletal, L. Chapter 6. Tannins and Their Influence on Health. In Recent Advances in Medicinal Chemistry; Bentham Science Publishers: Emirate of Sharjah, United Arab Emirates, 2014; Volume 1, p. 469. ISBN 978-1-60805-797-9. [Google Scholar]
- Yang, H.; Tuo, X.; Wang, L.; Tundis, R.; Portillo, M.P.; Simal-Gandara, J.; Yu, Y.; Zou, L.; Xiao, J.; Deng, J. Bioactive Procyanidins from Dietary Sources: The Relationship between Bioactivity and Polymerization Degree. Trends Food Sci. Technol. 2021, 111, 114–127. [Google Scholar] [CrossRef]
- Amoako, D.; Awika, J.M. Polyphenol Interaction with Food Carbohydrates and Consequences on Availability of Dietary Glucose. Curr. Opin. Food Sci. 2016, 8, 14–18. [Google Scholar] [CrossRef]
- Gu, L.; House, S.E.; Rooney, L.; Prior, R.L. Sorghum Bran in the Diet Dose Dependently Increased the Excretion of Catechins and Microbial-Derived Phenolic Acids in Female Rats. J. Agric. Food Chem. 2007, 55, 5326–5334. [Google Scholar] [CrossRef] [PubMed]
- Anunciação, P.C.; Cardoso, L.D.M.; Queiroz, V.A.V.; de Menezes, C.B.; de Carvalho, C.W.P.; Pinheiro-Sant’Ana, H.M.; Alfenas, R.D.C.G. Consumption of a Drink Containing Extruded Sorghum Reduces Glycaemic Response of the Subsequent Meal. Eur. J. Nutr. 2018, 57, 251–257. [Google Scholar] [CrossRef]
- Molino, S.; Lerma-Aguilera, A.; Jiménez-Hernández, N.; Gosalbes, M.J.; Rufián-Henares, J.Á.; Francino, M.P. Enrichment of Food With Tannin Extracts Promotes Healthy Changes in the Human Gut Microbiota. Front. Microbiol. 2021, 12, 625782. [Google Scholar] [CrossRef]
- Sallam, I.E.; Abdelwareth, A.; Attia, H.; Aziz, R.K.; Homsi, M.N.; von Bergen, M.; Farag, M.A. Effect of Gut Microbiota Biotransformation on Dietary Tannins and Human Health Implications. Microorganisms 2021, 9, 965. [Google Scholar] [CrossRef]
- Peng, K.; Zhao, H.; Wang, G.; Chen, B.; Mo, W.; Huang, Y. Effect of Condensed Tannins on Growth Performance, Intestinal Immune Capacity and Bacterial Microbiomes of Lateolabrax Japonicus. Aquac. Res. 2021, 52, 5321–5331. [Google Scholar] [CrossRef]
- Girard, A.L.; Awika, J.M. Sorghum Polyphenols and Other Bioactive Components as Functional and Health Promoting Food Ingredients. J. Cereal Sci. 2018, 84, 112–124. [Google Scholar] [CrossRef]
- Xu, J.; Wang, W.; Zhao, Y. Phenolic Compounds in Whole Grain Sorghum and Their Health Benefits. Foods 2021, 10, 1921. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.; Volino-Souza, M.; Conte-Júnior, C.A.; Alvares, T.S. Food-Derived Polyphenol Compounds and Cardiovascular Health: A Nano-Technological Perspective. Food Biosci. 2021, 41, 101033. [Google Scholar] [CrossRef]
- Mithul Aravind, S.; Wichienchot, S.; Tsao, R.; Ramakrishnan, S.; Chakkaravarthi, S. Role of Dietary Polyphenols on Gut Microbiota, Their Metabolites and Health Benefits. Food Res. Int. 2021, 142, 110189. [Google Scholar] [CrossRef]
- Awika, J.M.; Rooney, L.W. Sorghum Phytochemicals and Their Potential Impact on Human Health. Phytochemistry 2004, 65, 1199–1221. [Google Scholar] [CrossRef]
- Cisse, F.; Erickson, D.P.; Hayes, A.M.R.; Opekun, A.R.; Nichols, B.L.; Hamaker, B.R. Traditional Malian Solid Foods Made from Sorghum and Millet Have Markedly Slower Gastric Emptying than Rice, Potato, or Pasta. Nutrients 2018, 10, 124. [Google Scholar] [CrossRef]
- Agah, S.; Kim, H.; Mertens-Talcott, S.U.; Awika, J.M. Complementary Cereals and Legumes for Health: Synergistic Interaction of Sorghum Flavones and Cowpea Flavonols against LPS-Induced Inflammation in Colonic Myofibroblasts. Molr. Nutr. Food Res. 2017, 61, 1600625. [Google Scholar] [CrossRef]
- Taleon, V.; Dykes, L.; Rooney, W.L.; Rooney, L.W. Environmental Effect on Flavonoid Concentrations and Profiles of Red and Lemon-Yellow Sorghum Grains. J. Food Compos. Anal. 2014, 34, 178–185. [Google Scholar] [CrossRef]
- Przybylska-Balcerek, A.; Frankowski, J.; Stuper-Szablewska, K. The Influence of Weather Conditions on Bioactive Compound Content in Sorghum Grain. Eur. Food Res. Technol. 2020, 246, 13–22. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Z.; Li, Y.; Li, Z.; Liu, H.; Zhou, W. Metabolite Profiling of Sorghum Seeds of Different Colors from Different Sweet Sorghum Cultivars Using a Widely Targeted Metabolomics Approach. Int. J. Genomics 2020, 2020, 6247429. [Google Scholar] [CrossRef]
- Ma, D.; Li, Y.; Zhang, J.; Wang, C.; Qin, H.; Ding, H.; Xie, Y.; Guo, T. Accumulation of Phenolic Compounds and Expression Profiles of Phenolic Acid Biosynthesis-Related Genes in Developing Grains of White, Purple, and Red Wheat. Front. Plant Sci. 2016, 7, 528. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.; Behera, B.; Rahut, D.B. Climate Change and Indian Agriculture: A Systematic Review of Farmers’ Perception, Adaptation, and Transformation. Environ. Chall. 2022, 8, 100543. [Google Scholar] [CrossRef]
- Shaffril, H.A.M.; Krauss, S.E.; Samsuddin, S.F. A Systematic Review on Asian’s Farmers’ Adaptation Practices towards Climate Change. Sci. Total Environ. 2018, 644, 683–695. [Google Scholar] [CrossRef]
- Candau, F.; Regnacq, C.; Schlick, J. Climate Change, Comparative Advantage and the Water Capability to Produce Agricultural Goods. World Dev. 2022, 158, 105963. [Google Scholar] [CrossRef]
- Leghari, S.J.; Wahocho, N.; Laghari, G.; Laghari, A.; Bhabhan, G.; HussainTalpur, K.; Ahmed, T.; Wahocho, S.; Lashari, A. Role of Nitrogen for Plant Growth and Development: A Review. Adv. Environ. Biol. 2016, 10, 209–218. [Google Scholar]
- Xie, K.; Ren, Y.; Chen, A.; Yang, C.; Zheng, Q.; Chen, J.; Wang, D.; Li, Y.; Hu, S.; Xu, G. Plant Nitrogen Nutrition: The Roles of Arbuscular Mycorrhizal Fungi. J. Plant Physiol. 2022, 269, 153591. [Google Scholar] [CrossRef]
- Gu, J.; Li, Z.; Mao, Y.; Struik, P.C.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J. Roles of Nitrogen and Cytokinin Signals in Root and Shoot Communications in Maximizing of Plant Productivity and Their Agronomic Applications. Plant Sci. 2018, 274, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Wei, Y.; Yang, X.; Feng, Y.; Wu, C. Effects of Different Nitrogen Fertilizer Levels on Fe, Mn, Cu and Zn Concentrations in Shoot and Grain Quality in Rice (Oryza Sativa). Rice Sci. 2007, 14, 289–294. [Google Scholar] [CrossRef]
- Nguyen, P.M.; Niemeyer, E. Effects of Nitrogen Fertilization on the Phenolic Composition and Antioxidant Properties of Basil (Ocimum basilicum L.). J. Agric. Food Chem. 2008, 56, 8685–8691. [Google Scholar] [CrossRef]
- Lu, J.; Hu, T.; Zhang, B.; Wang, L.; Yang, S.; Fan, J.; Yan, S.; Zhang, F. Nitrogen Fertilizer Management Effects on Soil Nitrate Leaching, Grain Yield and Economic Benefit of Summer Maize in Northwest China. Agric. Water Manag. 2021, 247, 106739. [Google Scholar] [CrossRef]
- Maw, M.J.W.; Houx, J.H.; Fritschi, F.B. Nitrogen Fertilization of High Biomass Sorghum Affects Macro- and Micronutrient Accumulation and Tissue Concentrations. Ind. Crops Prod. 2020, 156, 112819. [Google Scholar] [CrossRef]
- Jevcsák, S.; Murányi, E.; Stündl, L.; Jóvér, J.; Sipos, P. Effect of Different Levels of Nitrogen on the Total Polyphenol and Total Flavonoid Content of Sorghum and Millet Flours. Acta Univ. Sapientiae, Aliment. 2017, 10, 107–115. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, L.; Zhang, G.; Xu, Y.; Zhang, L.; Gao, X.; Gao, J.; Jiang, N.; Shao, M. Optimal Fertilization for High Yield and Good Quality of Waxy Sorghum (Sorghum bicolor L. Moench). Field Crops Res. 2017, 203, 1–7. [Google Scholar] [CrossRef]
- Kovács, G.; Tuba, G.; Sinka, L.; Nagy, P.; Rivera–Garcia, A.; Zsembeli, J. A klímaváltozás lokális hatásainak értékelése a szélsőséges időjárási helyezetek tükrében. In 75 éves a Karcagi Kutatóintézet 1947–2022; Zsembeli, J., Ed.; Magyar Agrár- és Élettudományi Egyetem: Gödöllő, Hungary, 2022; pp. 27–35. ISBN 978-963-623-013-5. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Nemes, A.; Szőllősi, E.; Stündl, L.; Biró, A.; Homoki, J.R.; Szarvas, M.M.; Balogh, P.; Cziáky, Z.; Remenyik, J. Determination of Flavonoid and Proanthocyanidin Profile of Hungarian Sour Cherry. Molecules 2018, 23, 3278. [Google Scholar] [CrossRef] [PubMed]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A Critical Evaluation of the Vanillin Reaction as an Assay for Tannin in Sorghum Grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Zhu, F.; Cai, Y.Z.; Sun, M.; Ke, J.; Lu, D.; Corke, H. Comparison of major phenolic constituents and in vitro antioxidant activity of diverse Kudingcha genotypes from Ilex kudingcha, Ilex cornuta, and Ligustrum robustum. J. Agric. Food Chem. 2009, 57, 6082–6089. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Choi, S.C.; Kim, J.M.; Lee, Y.G.; Kim, C. Antioxidant Activity and Contents of Total Phenolic Compounds and Anthocyanins According to Grain Colour in Several Varieties of Sorghum bicolor (L.) Moench. Cereal Res. Commun. 2019, 47, 228–238. [Google Scholar] [CrossRef]
- Cheng, S.; Sun, Y.; Halgreen, L. The Relationships of Sorghum Kernel Pericarp and Testa Characteristics with Tannin Content. Asian J. Crop Sci. 2008, 1, 1–5. [Google Scholar] [CrossRef]
- Ayala-Soto, F.E.; Serna-Saldívar, S.O.; Welti-Chanes, J.; Gutierrez-Uribe, J.A. Phenolic Compounds, Antioxidant Capacity and Gelling Properties of Glucoarabinoxylans from Three Types of Sorghum Brans. J. Cereal Sci. 2015, 65, 277–284. [Google Scholar] [CrossRef]
- de Oliveira, L.D.L.; de Oliveira, G.T.; de Alencar, E.R.; Queiroz, V.A.V.; de Alencar Figueiredo, L.F. Physical, Chemical, and Antioxidant Analysis of Sorghum Grain and Flour from Five Hybrids to Determine the Drivers of Liking of Gluten-Free Sorghum Breads. LWT 2022, 153, 112407. [Google Scholar] [CrossRef]
- Ruiz-Hernández, A.A.; Cárdenas-López, J.L.; Cortez-Rocha, M.O.; González-Aguilar, G.A.; Robles-Sánchez, R.M. Optimization of Germination of White Sorghum by Response Surface Methodology for Preparing Porridges with Biological Potential. CyTA—J. Food 2021, 19, 49–55. [Google Scholar] [CrossRef]
- Awika, J.M.; Rooney, L.W.; Waniska, R.D. Properties of 3-Deoxyanthocyanins from Sorghum. J. Agric. Food Chem. 2004, 52, 4388–4394. [Google Scholar] [CrossRef] [PubMed]
- Brantsen, J.F.; Herrman, D.A.; Ravisankar, S.; Awika, J.M. Effect of Tannins on Microwave-Assisted Extractability and Color Properties of Sorghum 3-Deoxyanthocyanins. Food. Res. Intl. 2021, 148, 110612. [Google Scholar] [CrossRef]
- Shen, S.; Huang, R.; Li, C.; Wu, W.; Chen, H.; Shi, J.; Chen, S.; Ye, X. Phenolic Compositions and Antioxidant Activities Differ Significantly among Sorghum Grains with Different Applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef]
- Pinheiro, S.S.; Cardoso, L.D.M.; Anunciação, P.C.; de Menezes, C.B.; Queiroz, V.A.V.; Martino, H.S.D.; Della Lucia, C.M.; Pinheiro Sant’Ana, H.M. Water Stress Increased the Flavonoid Content in Tannin-Free Sorghum Grains. J. Food Compos. Anal. 2021, 100, 103892. [Google Scholar] [CrossRef]
- Verardo, V.; Cevoli, C.; Pasini, F.; Gómez-Caravaca, A.M.; Marconi, E.; Fabbri, A.; Caboni, M.F. Analysis of Oligomer Proanthocyanidins in Different Barley Genotypes Using High-Performance Liquid Chromatography–Fluorescence Detection–Mass Spectrometry and Near-Infrared Methodologies. J. Agric. Food Chem. 2015, 63, 4130–4137. [Google Scholar] [CrossRef]
- Ravisankar, S.; Abegaz, K.; Awika, J.M. Structural Profile of Soluble and Bound Phenolic Compounds in Teff (Eragrostis Tef) Reveals Abundance of Distinctly Different Flavones in White and Brown Varieties. Food Chem. 2018, 263, 265–274. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Satpal; Sangwan, S. Characterization of Phenolic Compounds and Antioxidant Activity in Sorghum [Sorghum bicolor (L.) Moench] Grains. Cereal Res. Commun. 2021, 49, 343–353. [Google Scholar] [CrossRef]
- Rao, S.; Santhakumar, A.B.; Chinkwo, K.A.; Wu, G.; Johnson, S.K.; Blanchard, C.L. Characterization of Phenolic Compounds and Antioxidant Activity in Sorghum Grains. J. Cereal Sci. 2018, 84, 103–111. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Pericarp Color | Ripening Time | Source |
|---|---|---|---|
| Zádor | Red/brown | early | Karcag, Hungary, DE-MATE |
| Alföldi1 | Red | semi-early | Karcag, Hungary, DE-MATE |
| ES Foehn | Red | semi-early | Karcag, Hungary, DE-MATE |
| Albita | White | semi-early | Karcag, Hungary, DE-MATE |
| Albanus | White | semi-late | Karcag, Hungary, DE-MATE |
| Farmsugro 180 | White | semi-late | Karcag, Hungary, DE-MATE |
| Year | pH (KCl) | KA | Water Soluble Total Salt Content (m/m%) | Carbonated Lime Content (m/m%) | Humus (m/m%) | (NO3 + NO2) N (mg kg−1) | AL-P2O5 (mg kg−1) | AL-K2O (mg kg−1) |
|---|---|---|---|---|---|---|---|---|
| 2019 | 5.1 | 46 | 0.02 | <0.05 | 3.4 | 6.4 | 136 | 486 |
| 2020 | 4.7 | 39 | <0.02 | 0.21 | 2.8 | 5.2 | 87 | 255 |
| 2021 | 4.9 | 44 | <0.02 | 0.21 | 3.3 | 9.2 | 175 | 462 |
| Source | df | TPC | CTC | TEAC | DPPH | FRAP |
|---|---|---|---|---|---|---|
| Variety | 5 | 6,051,167.304 *** | 1448.58 *** | 63,561.796115 *** | 112,949.518 *** | 183.634 *** |
| Year | 2 | 398,859.462 *** | 726.757 *** | 72.362315 | 8229.524 *** | 537.477 *** |
| Treatment | 1 | 14,645.948 ** | 3.925 | 77.76431 | 39.655 | 22.091 ** |
| Variety × Year | 9 | 230,327.66 *** | 437.516 *** | 223.486019 *** | 5689.3 *** | 48.15 *** |
| Variety × Treatment | 5 | 9275.932 ** | 3.733 | 20.806931 | 97.972 | 4.192 |
| Year × Treatment | 2 | 1128.094 | 4.959 | 97.713424 | 74.734 | 5.236 |
| Variety × Year × Treatment | 9 | 3547.406 | 5.31 ** | 78.991 * | 51.474 | 2.352 |
| Error | 79 | 1970.853 | 1.881 | 37.143 | 89.566 | 1.896 |
| Correlations | |||||
|---|---|---|---|---|---|
| TPC | CTC | TEAC | DPPH | FRAP | |
| TPC | 1 | 0.798 ** | 0.963 ** | 0.947 ** | 0.561 ** |
| CTC | 1 | 0.738 ** | 0.916 ** | 0.440 ** | |
| TEAC | 1 | 0.925 ** | 0.628 ** | ||
| DPPH | 1 | 0.558 ** | |||
| FRAP | 1 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagy, R.; Murányi, E.; Bíróné Molnár, P.; Szepesi, J.; Győri, Z.; Veres, S.; Remenyik, J.; Sipos, P. Assessment of Bioactive Profile of Sorghum Brans under the Effect of Growing Conditions and Nitrogen Fertilization. Agriculture 2023, 13, 760. https://doi.org/10.3390/agriculture13040760
Nagy R, Murányi E, Bíróné Molnár P, Szepesi J, Győri Z, Veres S, Remenyik J, Sipos P. Assessment of Bioactive Profile of Sorghum Brans under the Effect of Growing Conditions and Nitrogen Fertilization. Agriculture. 2023; 13(4):760. https://doi.org/10.3390/agriculture13040760
Chicago/Turabian StyleNagy, Róbert, Eszter Murányi, Piroska Bíróné Molnár, Judit Szepesi, Zoltán Győri, Szilvia Veres, Judit Remenyik, and Péter Sipos. 2023. "Assessment of Bioactive Profile of Sorghum Brans under the Effect of Growing Conditions and Nitrogen Fertilization" Agriculture 13, no. 4: 760. https://doi.org/10.3390/agriculture13040760
APA StyleNagy, R., Murányi, E., Bíróné Molnár, P., Szepesi, J., Győri, Z., Veres, S., Remenyik, J., & Sipos, P. (2023). Assessment of Bioactive Profile of Sorghum Brans under the Effect of Growing Conditions and Nitrogen Fertilization. Agriculture, 13(4), 760. https://doi.org/10.3390/agriculture13040760