
Current Status and Future Prospects of Head Rice Yield

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Factors Affecting Head Rice Yields
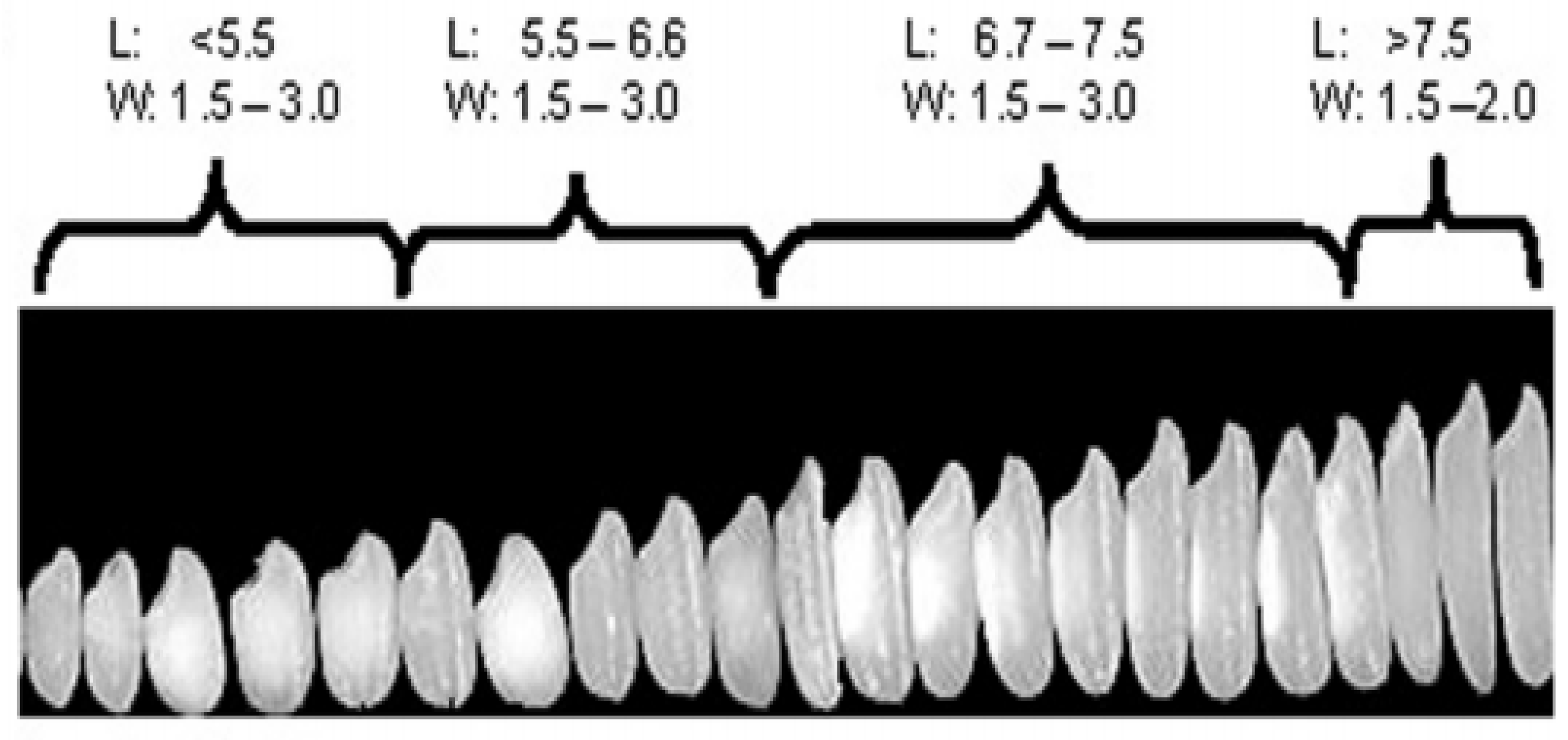
2.1. Rice Grain Size and Shape

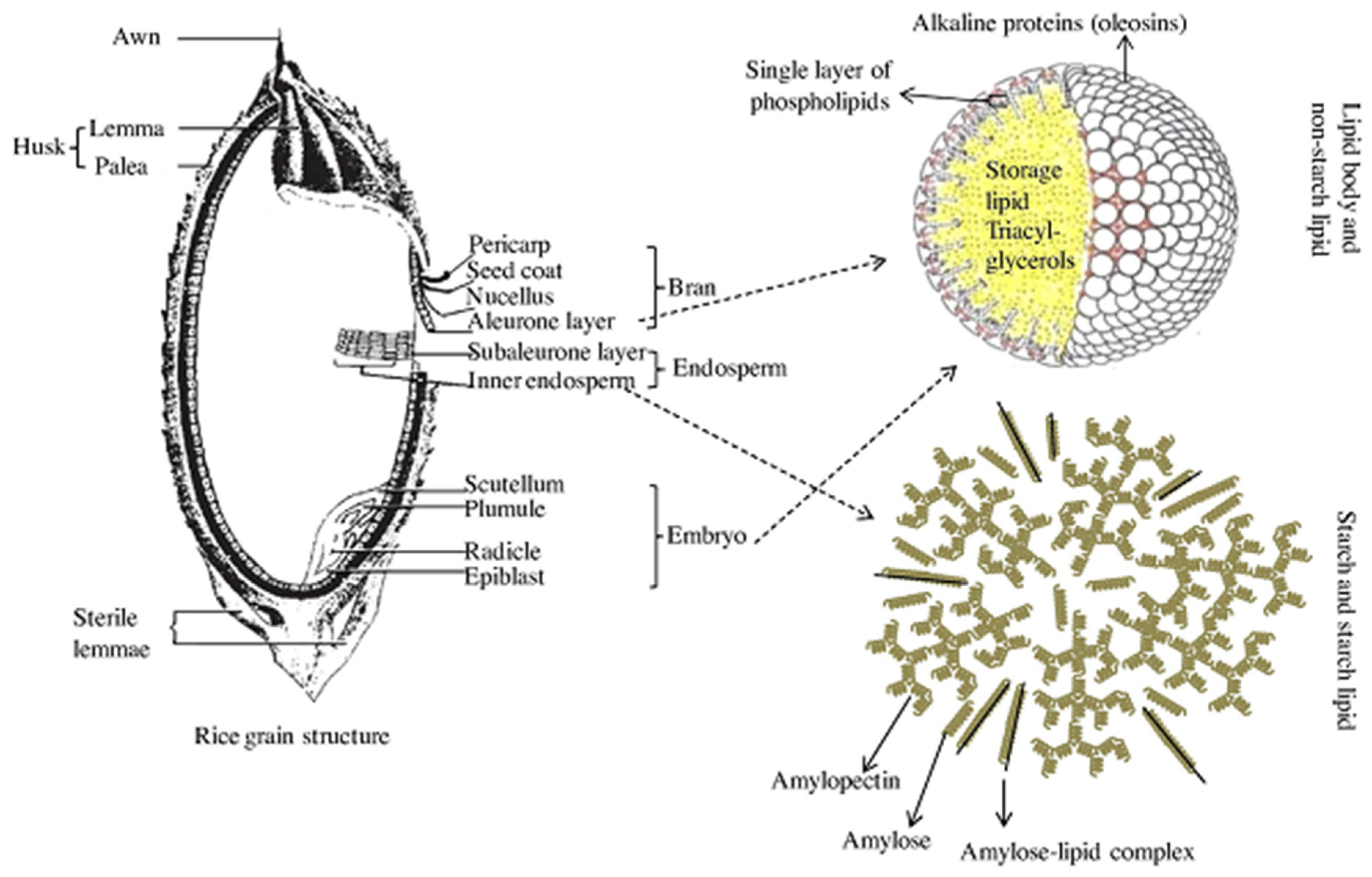
2.2. Rice Grain Molecular Composition
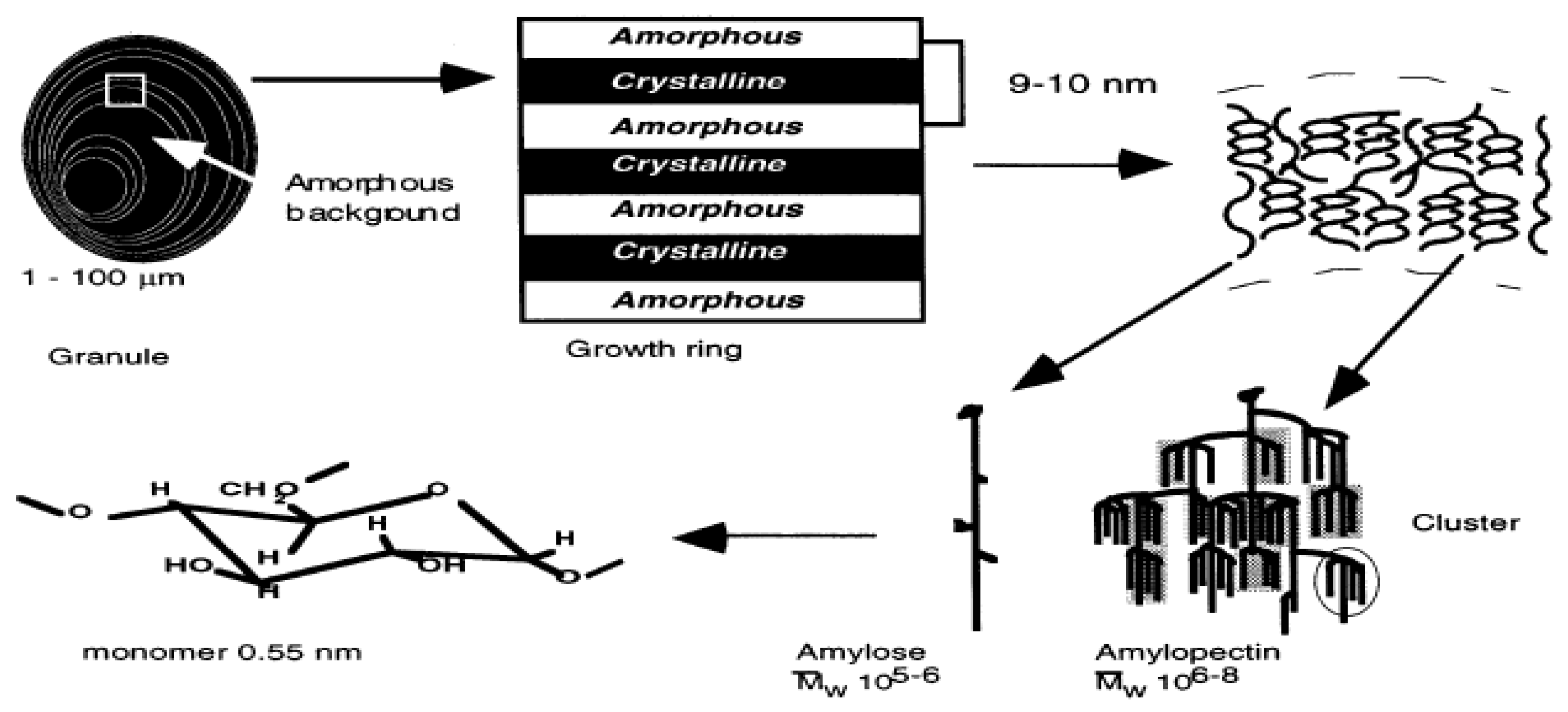
2.2.1. Starch
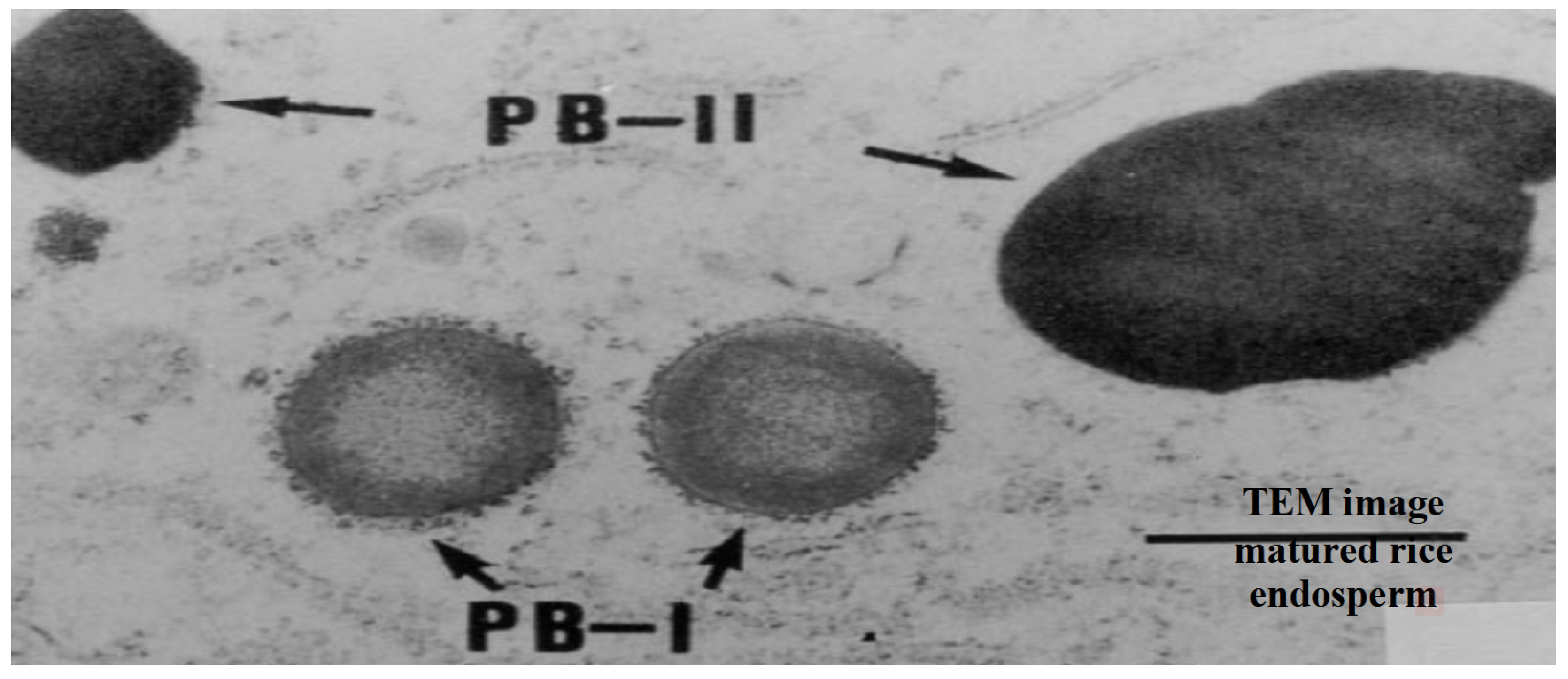
2.2.2. Seed Storage Proteins
2.2.3. Starch Lipids
2.2.4. Chalkiness
- (1)
- (2)
- (3)
- (4)
- (5)
- Environmental factors such as temperature, transpiration rate of canopy and evaporation rate contribute to developing chalkiness. Therefore, future studies are required to investigate which factors, environmental and genetic (in the context of Table 6), contribute to developing the chalky grains and finding the possible link between the rice grain molecular components and chalkiness.
2.3. Rice Plant Morphology and Grain Breakage
2.4. Pre-Harvest Factors That Affect Head Rice Yield
2.4.1. Temperature
2.4.2. Soil Moisture during Grain Filling
2.4.3. Nitrogen
2.5. Factors beyond Physiological Maturity Affecting Head Rice Yield
2.6. Genetics of Head Rice Yield
2.6.1. QTL Mapping for Head Rice Yield
2.6.2. GWAS (Genome-Wide Association Mapping)
2.6.3. Genomic Selection
3. Conclusions and Outlooks
Funding
Conflicts of Interest
References
- Juliano, B.O. The rice caryopsis and its composition. In Rice: Chemistry and Technology; Houston, D.F., Ed.; American Association of Cereal Chemists Inc.: St. Paul, MN, USA, 1985. [Google Scholar]
- Evers, T.; Millar, S. Cereal grain structure and development: Some implications for quality. J. Cereal Sci. 2002, 36, 261–284. [Google Scholar] [CrossRef]
- Bodie, A.R.; Micciche, A.C.; Atungulu, G.G.; Rothrock, M.J., Jr.; Ricke, S.C. Current Trends of Rice Milling Byproducts for Agricultural Applications and Alternative Food Production Systems. Front. Sustain. Food Syst. 2019, 3, 47. [Google Scholar] [CrossRef]
- Ohta, H.; Aibra, S.; Yamashita, H.; Sekiyama, F.; Morita, Y. Post-harvest drying of fresh rice grain and its effects on deterioration of lipids during storage. Agric. Biol. Chem. 1990, 54, 1157–1164. [Google Scholar]
- Urban-Alandete, L. Lipid Degradation during Grain Storage: Markers, Mechanisms and Shelf-Life Extension Treatments. Ph.D. Dissertation, School of Agriculture and Food Science, University of Queensland, Brisbane, Australia, 2018. [Google Scholar]
- Lyman, N.B.; Jagadish, K.S.V.; Nalley, L.L.; Dixon, B.L.; Siebenmorgen, T. Neglecting rice milling yield and quality underestimates economic losses from high-temperature stress. PLoS ONE 2013, 8, e72157. [Google Scholar] [CrossRef]
- Chandrajith, U.; Gunathilake, D.; Bandara, B.; Swarnasiri, D. Effects of Combine Harvesting on Head Rice Yield and Chaff Content of Long and Short Grain Paddy Harvest in Sri Lanka. Procedia Food Sci. 2016, 6, 242–245. [Google Scholar] [CrossRef]
- Esa, N.M.; Ling, T.B.; Peng, L.S. By-products of Rice Processing: An Overview of Health Benefits and Applications. Rice Res. Open Access 2013, 1, 1–11. [Google Scholar] [CrossRef]
- Ali, F.; Waters, D.; Ovenden, B.; Bundock, P.; Raymond, C.A.; Rose, T.J. Australian rice varieties vary in grain yield response to heat stress during reproductive and grain filling stages. J. Agron. Crop Sci. 2018, 205, 179–187. [Google Scholar] [CrossRef]
- Roy, P.; Ijiri, T.; Okadome, H.; Nei, D.; Orikasa, T.; Nakamura, N.; Shiina, T. Effect of processing conditions on overall energy consumption and quality of rice (Oryza sativa L.). J. Food Eng. 2008, 89, 343–348. [Google Scholar] [CrossRef]
- Sadeghi, M.; Hoseinian, S.H.; Hemmat, A. Influence of moisture content and whitening method on degree of milling and head rice yield of three Iranian rice varieties. Aust. J. Crop Sci. 2012, 6, 1481–1485. [Google Scholar]
- Sadeghi, M.; Nasrnia, E.; Masoumi, A.; Hemmat, A. Head rice yield response to low and high drying and tempering conditions. Int. Agrophysics 2013, 27, 219–223. [Google Scholar] [CrossRef]
- Statista. World Agriculture Production. 2022; p. 28. Available online: https://apps.fas.usda.gov/psdonline/circulars/production.pdf (accessed on 2 February 2023).
- Ali, F.; Waters, D.; Ovenden, B.; Bundock, P.; Raymond, C.; Rose, T. Heat stress during grain-filling reduces head rice yield through genotype dependant increased husk biomass and grain breakage. J. Cereal Sci. 2019, 90, 102820. [Google Scholar] [CrossRef]
- Abe, T.; Afzal, T.M. Tthin-layer infrared radiation drying of rough rice. J. Agric. Eng. Res. 1997, 67, 289–297. [Google Scholar] [CrossRef]
- Cnossen, A.G.; Siebenmorgen, T.J.; Yang, W. The glass transition temperature concept in rice drying and tempering: Effect on drying rate. Trans. ASAE 2002, 45, 759–766. [Google Scholar] [CrossRef]
- Dong, R.; Lu, Z.; Liu, Z.; Koide, S.; Cao, W. Effect of drying and tempering on rice fissuring analysed by integrating intra-kernel moisture distribution. J. Food Eng. 2010, 97, 161–167. [Google Scholar] [CrossRef]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef]
- Lanning, S.B.; Siebenmorgen, T.J.; Ambardekar, A.A.; Counce, P.A.; Bryant, R.J. Effects of Nighttime Air Temperature During Kernel Development of Field-Grown Rice on Physicochemical and Functional Properties. Cereal Chem. 2012, 89, 168–175. [Google Scholar] [CrossRef]
- Nalley, L.; Dixon, B.; Tack, J.; Barkley, A.; Jagadish, K. Optimal harvest moisture content for maximizing mid-south rice milling yields and returns. Agron. J. 2016, 108, 701–712. [Google Scholar] [CrossRef]
- Jongkaewwattana, S.; Geng, S.; Hill, J.E.; Miller, B.C. Within-Panicle Variability of grain filling in Rice Cultivars with Different Maturities. J. Agron. Crop Sci. 1993, 171, 236–242. [Google Scholar] [CrossRef]
- Monsoor, M.A.; Proctor, A.; Siebenmorgen, T.J. Surface lipid and free fatty acids (FFA) content of head and broken rice produced by milling after different drying treatments. Cereal Chem. 2004, 81, 705–709. [Google Scholar] [CrossRef]
- Li, X.; Wu, L.; Geng, X.; Xia, X.; Wang, X.; Xu, Z.; Xu, Q. Deciphering the Environmental Impacts on Rice Quality for Different Rice Cultivated Areas. Rice 2018, 11, 7. [Google Scholar] [CrossRef]
- Grigg, B.C.; Siebenmorgen, T.J.; Norman, R.J. Effects of Nitrogen Rate and Harvest Moisture Content on Physicochemical Properties and Milling Yields of Rice. Cereal Chem. 2015, 93, 172–181. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, C.; Li, Y.; Zhang, A.; Dong, G.; Xie, L.; Bin Zhang, B.; Ruan, B.; Hong, K.; Xue, D.; et al. QTL analysis for chalkiness of rice and fine mapping of a candidate gene for qACE9. Rice 2016, 9, 41. [Google Scholar] [CrossRef]
- Jennings, P.R.; Coffman, W.R.; Kauffman, H.E. Rice Improvement; IRRI: Los Banos, Laguna, Philippines, 1979; pp. 101–120. [Google Scholar]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Takano-Kai, N.; Jiang, H.; Kubo, T.; Sweeney, M.; Matsumoto, T.; Kanamori, H.; Padhukasahasram, B.; Bustamante, C.; Yoshimura, A.; Doi, K.; et al. Evolutionary History of GS3, a Gene Conferring Grain Length in Rice. Genetics 2009, 182, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Lin, Y.S.; Song, X.J.; Shen, J.B.; Huang, W.; Shan, J.X.; Zhu, M.Z.; Jiang, L.W.; Gao, J.P.; Lin, H.X. The novel quantitative trait locus GL3.1 control rice grain size and yield by regulating Cyclin-T1;3. Cell Res. 2012, 22, 1666–1680. [Google Scholar] [CrossRef] [PubMed]
- Song, X.J.; Huang, W.; Shi, M.; Zhu, M.Z.; Lin, H.X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet. 2007, 39, 623–630. [Google Scholar] [CrossRef]
- Calingacion, M.; Laborte, A.; Nelson, A.; Resurreccion, A.; Concepcion, J.C.; Daygon, V.D.; Mumm, R.; Reinke, R.; Dipti, S.; Bassinello, P.Z.; et al. Diversity of Global Rice Markets and the Science Required for Consumer-Targeted Rice Breeding. PLoS ONE 2014, 9, e85106. [Google Scholar] [CrossRef]
- Kubo, T.; Takano-Kai, N.; Yoshimura, A. RFLP mapping of genes for long kernel and awn on chromosome 3 in rice. Rice Genet. Newsl. 2001, 18, 26–28. [Google Scholar]
- Thomson, M.; Tai, T.H.; McClung, A.M.; Lai, X.-H.; Hinga, M.E.; Lobos, K.B.; Xu, Y.; Martinez, C.P.; McCouch, S.R. Mapping quantitative trait loci for yield, yield components and morphological traits in an advanced backcross population between Oryza rufipogon and the Oryza sativa cultivar Jefferson. Theor. Appl. Genet. 2003, 107, 479–493. [Google Scholar] [CrossRef]
- Li, J.; Thomson, M.; McCouch, S.R. Fine Mapping of a Grain-Weight Quantitative Trait Locus in the Pericentromeric Region of Rice Chromosome 3. Genetics 2004, 168, 2187–2195. [Google Scholar] [CrossRef]
- Zhou, L.Q.; Wang, Y.P.; Li, S.G. Genetic analysis and physical mapping of lk-4(t), a major gene controlling grain length in rice, with a BC2F2 population. Acta Genet. Sin. 2006, 33, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Gu, S.; Wan, X.; Gao, H.; Guo, T.; Su, N.; Lei, C.; Zhang, X.; Cheng, Z.; Guo, X.; et al. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 2008, 18, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Qian, Q.; Liu, Z.; Sun, H.; He, S.; Luo, D.; Xia, G.; Chu, C.; Li, J.; Fu, X. Natural variation at the DEP1 locus enhances grain yield in rice. Nat. Genet. 2009, 41, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Luo, L.; Yan, W.; Kovi, M.R.; Zhan, W.; Xing, Y. Genetic dissection of rice grain shape using a recombinant inbred line population derived from two contrasting parents and fine mapping a pleiotropic quantitative trait locus qGL7. BMC Genet. 2010, 11, 16. [Google Scholar] [CrossRef]
- Kato, T.; Segami, S.; Toriyama, M.; Kono, I.; Ando, T.; Yano, M.; Kitano, H.; Miura, K.; Iwasaki, Y. Detection of QTLs for grain length from large grain rice (Oryza sativa L.). Breed. Sci. 2011, 61, 269–274. [Google Scholar] [CrossRef]
- Qiu, X.; Gong, R.; Tan, Y.; Yu, S. Mapping and characterization of the major quantitative trait locus qSS7 associated with increased length and decreased width of rice seeds. Theor. Appl. Genet. 2012, 125, 1717–1726. [Google Scholar] [CrossRef]
- Ishimaru, K.; Hirotsu, N.; Madoka, Y.; Murakami, N.; Hara, N.; Onodera, H.; Kashiwagi, T.; Ujiie, K.; Shimizu, B.-I.; Onishi, A.; et al. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat. Genet. 2013, 45, 707–711. [Google Scholar] [CrossRef]
- Zhao, D.; Li, P.; Wang, L.; Sun, L.; Xia, D.; Luo, L.; Gao, G.; Zhang, Q.; He, Y. Genetic dissection of large grain shape in rice cultivar ‘Nanyangzhan’ and validation of a grain thickness QTL (qGT3.1) and a grain length QTL (qGL3.4). Mol. Breed. 2017, 37, 42. [Google Scholar] [CrossRef]
- Segami, S.; Takehara, K.; Yamamoto, T.; Kido, S.; Kondo, S.; Iwasaki, Y.; Miura, K. Overexpression of SRS5 improves grain size of brassinosteroid-related dwarf mutants in rice (Oryza sativa L.). Breed. Sci. 2017, 67, 393–397. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.; Fang, Y.; Zeng, L.; Xu, J.; Yu, H.; Shi, Z.; Pan, J.; Zhang, D.; Kang, S.; et al. A rare allele of gs2 enhances grain size and grain yield in rice. Mol. Plant 2015, 8, 1455–1465. [Google Scholar] [CrossRef]
- Xu, C.; Liu, Y.; Li, Y.; Xu, X.; Xu, C.; Li, X.; Xiao, J.; Zhang, Q. Differential expression of GS5 regulates grain size in rice. J. Exp. Bot. 2015, 66, 2611–2623. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Tong, H.; Xiao, Y.; Che, R.; Xu, F.; Hu, B.; Liang, C.; Chu, J.; Li, J.; Chu, C. Activation of Big Grain1 significantly improves grain size by regulating auxin transport in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11102–11107. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, G.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.; Zeng, L.; Xu, E.; Xu, J.; et al. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.; Wei, X.; Chen, M.; Tang, S.; Luo, J.; Jiao, G.; Xie, L.; Hu, P. Allelic variation for a candidate gene for GS7, responsible for grain shape in rice. Theor. Appl. Genet. 2012, 125, 1303–1312. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Huang, J.; Lan, H.; Wang, C.; Yin, C.; Wu, Y.; Tang, H.; Qian, Q.; Li, J.; et al. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice. Proc. Natl. Acad. Sci. USA 2012, 109, 21534–21539. [Google Scholar] [CrossRef]
- Segami, S.; Kono, I.; Ando, T.; Yano, M.; Kitano, H.; Miura, K.; Iwasaki, Y. Small and round seed 5 gene encodes alpha-tubulin regulating seed cell elongation in rice. Rice 2012, 5, 4. [Google Scholar] [CrossRef]
- Si, L.; Chen, J.; Huang, X.; Gong, H.; Luo, J.; Hou, Q.; Zhou, T.; Lu, T.; Zhu, J.; Shangguan, Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef]
- Zhou, Y.; Hou, J.; Li, P. Genetic dissection and validation of QTLs for grain shape and weight in rice and fine mapping of qGL1.3, a major QTL for grain length and weight. Mol. Breed. 2019, 39, 170. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, S.; Sun, S.; Zhang, Y.; Li, J.; You, J.; Su, T.; Chen, W.; Ling, Y.; He, G.; et al. Analysis of QTL for grain size in a rice chromosome segment substitution line Z1392 with long grains and fine mapping of qGL-6. Rice 2020, 13, 40. [Google Scholar] [CrossRef]
- Li, X.; Wei, Y.; Li, J.; Yang, F.; Chen, Y.; Chen, Y.; Guo, S.; Sha, A. Identification of QTL TGW12 responsible for grain weight in rice based on recombinant inbred line population crossed by wild rice (Oryza minuta) introgression line K1561 and indica rice G1025. BMC Genet. 2020, 21, 10. [Google Scholar] [CrossRef]
- Anilkumar, C.; Sah, R.P.; Azharudheen, T.P.M.; Behera, S.; Singh, N.; Prakash, N.R.; Sunitha, N.C.; Devanna, B.N.; Marndi, B.C.; Patra, B.C.; et al. Understanding complex genetic architecture of rice grain weight through QTL-meta analysis and candidate gene identification. Sci. Rep. 2022, 12, 13832. [Google Scholar] [CrossRef] [PubMed]
- Koutroubas, S.D.; Mazzini, F.; Pons, B.; Ntanos, D.A. Grain quality variation and relationships with morpho-physiological traits in rice (Oryza sativa L.) genetic resources in Europe. Field Crops Res. 2004, 86, 115–130. [Google Scholar] [CrossRef]
- Xie, L.; Tang, S.; Chen, N.; Luo, J.; Jiao, G.; Shao, G.; Wei, X.; Hu, P. Rice Grain Morphological Characteristics Correlate with Grain Weight and Milling Quality. Cereal Chem. 2013, 90, 587–593. [Google Scholar] [CrossRef]
- Jongkaewwattana, S.; Geng, S. Inter-relationships amongst grain characteristics, grain-filling parameters and rice (Oryza sativa L.) milling quality. J. Agron. Crop. Sci. 2001, 187, 223–229. [Google Scholar] [CrossRef]
- Jongkaewwattana, S.; Geng, S. Non-Uniformity of Grain Characteristics and Milling Quality of California Rice (Oryza sativa L.) of Different Maturities. J. Agron. Crop Sci. 2002, 188, 161–167. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef]
- Zhou, Z.K.; Robards, K.; Helliwell, S.; Blanchard, C. Composition and functional properties of rice. Int. J. Food Sci. Technol. 2002, 37, 849–868. [Google Scholar] [CrossRef]
- Wang, Y.J.; Wang, L.F.; Shephard, D.; Wang, F.D.; Patindol, J. Properties and structures of flours and starches from whole, broken, and yellowed rice kernels in a model study. Cereal Chem. 2002, 79, 383–386. [Google Scholar] [CrossRef]
- Buléon, A.; Colonna, P.; Planchot, V.; Ball, S. Starch granules: Structure and biosynthesis. Int. J. Biol. Macromol. 1998, 23, 85–112. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Complex Carbohydrates Are Formed by Linkage of Monosaccharides. Biochemistry, 5th ed.; W H Freeman: New York, NY, USA, 2002. [Google Scholar]
- Bemiller, J.; Whistler, R. Starch: Chemistry and Technology, 3rd ed.; Academy Press: Cambridge, UK, 2009; pp. 310–315. [Google Scholar]
- Sano, Y. Differential regulation of waxy gene expression in rice endosperm. Theor. Appl. Genet. 1984, 68, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Xu, M.; Liu, X.; Yan, C.; Korban, S.S.; Chen, X.; Gu, M. Genes coding for starch branching enzymes are major contributors to starch viscosity characteristics in waxy rice (Oryza sativa L.). Plant Sci. 2004, 166, 357–364. [Google Scholar] [CrossRef]
- Cuevas, R.P.; Daygon, V.D.; Morell, M.K.; Gilbert, R.G.; Fitzgerald, M.A. Using chain-length distributions to diagnose genetic diversity in starch biosynthesis. Carbohydr. Polym. 2010, 81, 120–127. [Google Scholar] [CrossRef]
- Counce, P.A.; Bryant, R.J.; Bergman, C.J.; Bautista, R.C.; Wang, Y.-J.; Siebenmorgen, T.J.; Moldenhauer, K.A.K.; Meullenet, J.-F.C. Rice milling quality, grain dimensions, and starch branching as affected by high night temperatures. Cereal Chem. 2005, 82, 645–648. [Google Scholar] [CrossRef]
- Kawasaki, T.; Mizuno, K.; Shimada, H.; Satoh, H.; Kishimoto, N.; Okumura, S.; Ichikawa, N.; Baba, T. Coordinated regulation of the genes participating in starch biosynthesis by the rice Floury-2 locus. Plant Physiol. 1996, 110, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Nishihara, M.; Mizuno, K.; Kawasaki, T.; Shimada, H.; Kobayashi, E.; Ohnishi, S.; Tanaka, K.; Arai, Y. Identification, cdna cloning, and gene-expression of soluble starch synthase in rice (Oryza sativa L) immature seeds. Plant Physiol. 1993, 103, 565–573. [Google Scholar] [CrossRef]
- She, K.-C.; Kusano, H.; Koizumi, K.; Yamakawa, H.; Hakata, M.; Imamura, T.; Fukuda, M.; Naito, N.; Tsurumaki, Y.; Yaeshima, M.; et al. A Novel factor Floury Endosperm2 is involved in regulation of rice grain size and starch quality. Plant Cell 2010, 22, 3280–3294. [Google Scholar] [CrossRef]
- Zhao, L.; Zhao, C.; Zhou, L.; Zhao, Q.; Zhu, Z.; Chen, T.; Yao, S.; Zhang, Y.; Wang, C. QTL mapping for starch paste viscosity of rice (Oryza sativa L.) using chromosome segment substitution lines derived from two sequenced cultivars with the same Wx allele. BMC Genom. 2021, 22, 596. [Google Scholar] [CrossRef]
- Yamagata, H.; Tanaka, K. The Site of Synthesis and Accumulation of Rice Storage Proteins. Plant Cell Physiol. 1986, 27, 135–145. [Google Scholar] [CrossRef]
- Miflin, B.J.; Field, J.M.; Shewry, P.R. Cereal storage proteins and their effects on technological properties. In Seed Proteins; Daussant, J., Mosse, J., Vaughan, J., Eds.; Academic Press: London, UK, 1983; pp. 255–319. [Google Scholar]
- Muench, D.G.; Ogawa, M.; Okita, T.W. The Prolamins of Rice. In Seed Proteins; Shewry, P.R., Casey, R., Eds.; Springer: Dordrecht, The Netherlands, 1999. [Google Scholar]
- Xu, J.H.; Messing, J. Amplification of prolamin storage protein genes in different subfamilies of the Poaceae. Theor. Appl. Genet. 2009, 119, 1397–1412. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Yamamoto, M.P.; Hirose, S.; Yano, M.; Takaiwa, F. Characterization of a new rice glutelin gene GluD-1 expressed in the starchy endosperm. J. Exp. Bot. 2008, 59, 4233–4245. [Google Scholar] [CrossRef] [PubMed]
- Kawakatsu, T.; Hirose, S.; Yasuda, H.; Takaiwa, F. Reducing Rice Seed Storage Protein Accumulation Leads to Changes in Nutrient Quality and Storage Organelle Formation. Plant Physiol. 2010, 154, 1842–1854. [Google Scholar] [CrossRef] [PubMed]
- Pedrazzini, E.; Mainieri, D.; Marrano, C.A.; Vitale, A. Where do protein bodies of cereal seeds come from? Front. Plant Sci. 2016, 7, 1139. [Google Scholar] [CrossRef] [PubMed]
- Nakase, M.; Hotta, H.; Adachi, T.; Aoki, N.; Nakamura, R.; Masumura, T.; Tanaka, K.; Matsuda, T. Cloning of the rice seed α-globulin-encoding gene: Sequence similarity of the 5′-flanking region to those of the genes encoding wheat high-molecular-weight glutenin and barley D hordein. Gene 1996, 170, 223–226, ISSN 0378-1119. [Google Scholar] [CrossRef]
- Tan, F.Y.; Sun, M.; Xing, Z.Y.; Hua, P.J.; Sun, L.X.; Zhang, F.Q.; Corke, H. Mapping quantitative trait loci for milling quality, protein content and colour characteristics of rice using a recombinant inbred line population derived from an elite rice hybrid. Theor. Appl. Genet. 2001, 103, 1037–1045. [Google Scholar] [CrossRef]
- Aluko, G.; Martinez, C.; Tohme, J.; Castano, C.; Bergman, C.; Oard, J.H. QTL mapping of grain quality traits from the interspecific cross Oryza sativa × O. glaberrima. Theor. Appl. Genet. 2004, 109, 630–639. [Google Scholar] [CrossRef]
- Hu, Z.-L.; Li, P.; Zhou, M.-Q.; Zhang, Z.-H.; Wang, L.-X.; Zhu, L.-H.; Zhu, Y.-G. Mapping of quantitative trait loci (QTLs) for rice protein and fat content using doubled haploid lines. Euphytica 2004, 135, 47–54. [Google Scholar] [CrossRef]
- Peng, B.; Kong, H.; Li, Y.; Wang, L.; Zhong, M.; Sun, L.; Gao, G.; Zhang, Q.; Luo, L.; Wang, G.; et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 2014, 5, 4847. [Google Scholar] [CrossRef]
- Chattopadhyay, K.; Behera, L.; Bagchi, T.B.; Sardar, S.S.; Moharana, N.; Patra, N.R.; Chakraborti, M.; Das, A.; Marndi, B.C.; Sarkar, A.; et al. Detection of stable QTLs for grain protein content in rice (Oryza sativa L.) employing high throughput phenotyping and genotyping platforms. Sci. Rep. 2019, 9, 3196. [Google Scholar] [CrossRef]
- Lee, S.; Park, J.; Lee, J.; Shin, D.; Marmagne, A.; Lim, P.O.; Masclaux-Daubresse, C.; An, G.; Gil Nam, H. OsASN1 Overexpression in Rice Increases Grain Protein Content and Yield under Nitrogen-Limiting Conditions. Plant Cell Physiol. 2020, 61, 1309–1320. [Google Scholar] [CrossRef]
- Fiaz, S.; Sheng, Z.; Zeb, A.; Barman, H.N.; Shar, T.; Ali, U.; Tang, S. Analysis of genomic regions for crude protein and fractions of protein using a recombinant inbred population in Rice (Oryza sativa L.). J. Taibah Univ. Sci. 2021, 15, 579–588. [Google Scholar] [CrossRef]
- Choudhury, N.H.; Juliano, B.O. Lipids in developing and mature rice grain. Phytochemistry 1980, 19, 1063–1069. [Google Scholar] [CrossRef]
- Taira, H.; Itani, T. Lipid content and fatty acid composition of brown rice of cultivars of the United States. J. Agric. Food Chem. 1988, 36, 460–462. [Google Scholar] [CrossRef]
- Mano, Y.; Kawaminami, K.; Kojima, M.; Ohnishi, M.; Ito, S. Comparative Composition of Brown Rice Lipids (Lipid Fractions) of Indica and Japonica Rices. Biosci. Biotechnol. Biochem. 1999, 63, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Whattam, J.; Cornell, H.J. Distribution and Composition of the Lipids in Starch Fractions from Wheat Flour. Starch Stärke 1991, 43, 152–156. [Google Scholar] [CrossRef]
- Liu, L.; Waters, D.L.; Rose, T.J.; Bao, J.; King, G.J. Phospholipids in rice: Significance in grain quality and health benefits: A review. Food Chem. 2013, 139, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Fujino, Y. Rice lipids. Cereal Chem. 1978, 55, 559–571. [Google Scholar]
- Lam, H.S.; Proctor, A. Lipid hydrolysis and oxidation on the surface of milled rice. J. Am. Oil Chem. Soc. 2003, 80, 563–567. [Google Scholar] [CrossRef]
- Finnie, S.; Jeannotte, R.; Morris, C.; Giroux, M.; Faubion, J. Variation in polar lipids located on the surface of wheat starch. J. Cereal Sci. 2010, 51, 73–80. [Google Scholar] [CrossRef]
- Pasha, I.; Anjum, F.; Morris, C. Grain Hardness: A major determinant of wheat quality. Food Sci. Technol. Int. 2010, 16, 511–522. [Google Scholar] [CrossRef]
- Pauly, A.; Pareyt, B.; Fierens, E.; Delcour, J.A. Wheat (Triticum aestivum L. and T. turgidum L. ssp. durum) kernel hardness: Current View on the Role of Puroindolines and Polar Lipids. Compr. Rev. Food Sci. Food Saf. 2013, 12, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Zhang, W.W.; Liu, L.L.; Shen, Y.Y.; Wang, J.K.; Jiang, L.; Zhai, H.Q.; Wan, J.M. Dynamic QTL Analysis on Rice Fat Content and Fat Index Using Recombinant Inbred Lines. Cereal Chem. 2008, 85, 769–775. [Google Scholar] [CrossRef]
- Liu, W.; Zeng, J.; Jiang, G.; He, Y. QTLs identification of crude fat content in brown rice and its genetic basis analysis using DH and two backcross populations. Euphytica 2009, 169, 197–205. [Google Scholar] [CrossRef]
- Yu, Y.-H.; Li, G.; Fan, Y.-Y.; Zhang, K.-Q.; Min, J.; Zhu, Z.-W.; Zhuang, J.-Y. Genetic relationship between grain yield and the contents of protein and fat in a recombinant inbred population of rice. J. Cereal Sci. 2009, 50, 121–125. [Google Scholar] [CrossRef]
- Qin, Y.; Kim, S.-M.; Zhao, X.; Lee, H.-S.; Jia, B.; Kim, K.-M.; Eun, M.-Y.; Sohn, J.-K. QTL detection and MAS selection efficiency for lipid content in brown rice (Oryza sativa L.). Genes Genom. 2010, 32, 506–512. [Google Scholar] [CrossRef]
- Ying, J.-Z.; Shan, J.-X.; Gao, J.-P.; Zhu, M.-Z.; Shi, M.; Lin, H.-X. Identification of Quantitative Trait Loci for Lipid Metabolism in Rice Seeds. Mol. Plant 2012, 5, 865–875. [Google Scholar] [CrossRef]
- Lee, G.-H.; Yun, B.-W.; Kim, K.-M. Analysis of QTLs Associated with the Rice Quality Related Gene by Double Haploid Populations. Int. J. Genom. 2014, 2014, 781832. [Google Scholar] [CrossRef]
- Zhou, H.; Xia, D.; Li, P.; Ao, Y.; Xu, X.; Wan, S.; Li, Y.; Wu, B.; Shi, H.; Wang, K.; et al. Genetic architecture and key genes controlling the diversity of oil composition in rice grains. Mol. Plant 2020, 14, 456–469, ISSN 0378-1119. [Google Scholar] [CrossRef]
- Li, Q.; Zhai, W.; Wei, J.; Jia, Y. Rice lipid transfer protein, OsLTPL23, controls seed germination by regulating starch-sugar conversion and ABA homeostasis. Front. Genet. 2023, 14, 1111318. [Google Scholar] [CrossRef]
- Lisle, A.J.; Martin, M.; Fitzgerald, M.A. Chalky and Translucent Rice Grains Differ in Starch Composition and Structure and Cooking Properties. Cereal Chem. 2000, 77, 627–632. [Google Scholar] [CrossRef]
- Yamakawa, H.; Ebitani, T.; Terao, T. Comparison between locations of QTLs for grain chalkiness and genes responsive to high temperature during grain filling on the rice chromosome map. Breed. Sci. 2008, 58, 337–343. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, L.; Jiang, L.; Zhang, W.; Liu, L.; Liu, X.; Zhao, Z.; Liu, S.; Zhang, L.; Wang, J.; et al. Fine mapping of the grain chalkiness QTL qPGWC-7 in rice (Oryza sativa L.). Theor. Appl. Genet. 2009, 118, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, C.; Xing, Y.; Yun, P.; Luo, L.; Yan, B.; Peng, B.; Xie, W.; Wang, G.; Li, X.; et al. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice. Nat. Genet. 2014, 46, 657. [Google Scholar] [CrossRef]
- Xi, M.; Lin, Z.; Zhang, X.; Liu, Z.; Li, G.; Wang, Q.; Wang, S.; Ding, Y. Endosperm Structure of White-Belly and White-Core Rice Grains Shown by Scanning Electron Microscopy. Plant Prod. Sci. 2014, 17, 285–290. [Google Scholar] [CrossRef]
- Zhou, L.; Liang, S.; Ponce, K.; Marundon, S.; Ye, G.; Zhao, X. Factors affecting head rice yield and chalkiness in indica rice. Field Crop. Res. 2015, 172, 1–10. [Google Scholar] [CrossRef]
- Lin, Z.; Zheng, D.; Zhang, X.; Wang, Z.; Lei, J.; Liu, Z.; Li, G.; Wang, S.; Ding, Y. Chalky part differs in chemical composition from translucent part of japonica rice grains as revealed by a notched-belly mutant with white-belly. J. Sci. Food Agric. 2016, 96, 3937–3943. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Daygon, V.D.; McNally, K.L.; Hamilton, R.S.; Xie, F.; Reinke, R.F.; Fitzgerald, M.A. Identification of stable QTLs causing chalk in rice grains in nine environments. Theor. Appl. Genet. 2016, 129, 141–153. [Google Scholar] [CrossRef]
- Jin, S.-K.; Xu, L.-N.; Yang, Q.-Q.; Zhang, M.-Q.; Wang, S.-L.; Wang, R.-A.; Tao, T.; Hong, L.-M.; Guo, Q.-Q.; Jia, S.-W.; et al. High-resolution quantitative trait locus mapping for rice grain quality traits using genotyping by sequencing. Front. Plant Sci. 2023, 13, 1050882. [Google Scholar] [CrossRef]
- Lin, C.-J.; Li, C.-Y.; Lin, S.-K.; Yang, F.-H.; Huang, J.-J.; Liu, Y.-H.; Lur, H.-S. Influence of High Temperature during Grain Filling on the Accumulation of Storage Proteins and Grain Quality in Rice (Oryza sativa L.). J. Agric. Food Chem. 2010, 58, 10545–10552. [Google Scholar] [CrossRef]
- Zhao, X.; Fitzgerald, M. Climate Change: Implications for the Yield of Edible Rice. PLoS ONE 2013, 8, e66218. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, W.; Xu, Z. Relationship between grain yield and quality in rice germplasms grown across different growing areas. Breed. Sci. 2015, 65, 226–232. [Google Scholar] [CrossRef]
- Fu, F.F.; Xue, H.W. Coexpression Analysis Identifies Rice Starch Regulator1, a Rice AP2/EREBP Family Transcription Factor, as a Novel Rice Starch Biosynthesis Regulator. Plant Physiol. 2010, 154, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-C.; Xu, H.; Zhu, Y.; Liu, Q.-Q.; Cai, X.-L. OsbZIP58, a basic leucine zipper transcription factor, regulates starch biosynthesis in rice endosperm. J. Exp. Bot. 2013, 64, 3453–3466. [Google Scholar] [CrossRef]
- Zhou, P.; Tan, Y.; He, Y.; Xu, C.; Zhang, Q. Simultaneous improvement for four quality traits of Zhenshan 97, an elite parent of hybrid rice, by molecular marker-assisted selection. Theor. Appl. Genet. 2003, 106, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhang, X.; Wang, Z.; Jiang, Y.; Liu, Z.; Alexander, D.; Li, G.; Wang, S.; Ding, Y. Metabolomic analysis of pathways related to rice grain chalkiness by a notched-belly mutant with high occurrence of white-belly grains. BMC Plant Biol. 2017, 17, 39. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Li, G.S.; Qian, Q.; Ma, Q.Y.; Li, Z.J.; Wang, M.W.; Chen, Y.; Zhu, H.L. Genetic analysis of rice grain quality. Theor. Appl. Genet. 1999, 98, 502–508. [Google Scholar] [CrossRef]
- Tan, Y.F.; Xing, Y.Z.; Li, J.X.; Yu, S.B.; Xu, C.G.; Zhang, Q. Genetic bases of appearance quality of rice grains in Shanyou 63, an elite rice hybrid. Theor. Appl. Genet. 2000, 101, 823–829. [Google Scholar] [CrossRef]
- Wan, X.Y.; Wan, J.M.; Weng, J.F.; Jiang, L.; Bi, J.C.; Wang, C.M.; Zhai, H.Q. Stability of QTLs for rice grain dimension and endosperm chalkiness characteristics across eight environments. Theor. Appl. Genet. 2005, 110, 1334–1346. [Google Scholar] [CrossRef]
- Yang, W.; Hao, Q.; Liang, J.; Tan, Q.; Luan, X.; Lin, S.; Zhu, H.; Bu, S.; Liu, Z.; Liu, G.; et al. Fine Mapping of Two Major Quantitative Trait Loci for Rice Chalkiness With High Temperature-Enhanced Additive Effects. Front. Plant Sci. 2022, 13, 957863. [Google Scholar] [CrossRef]
- Shi, H.; Zhu, Y.; Yun, P.; Lou, G.; Wang, L.; Wang, Y.; Gao, G.; Zhang, Q.; Li, X.; He, Y. Fine Mapping of qWCR4, a Rice Chalkiness QTL Affecting Yield and Quality. Agronomy 2022, 12, 706. [Google Scholar] [CrossRef]
- Kumari, A.; Sharma, D.; Sharma, P.; Wang, C.; Verma, V.; Patil, A.; Imran, M.; Singh, M.P.; Kumar, K.; Paritosh, K.; et al. Meta-QTL and haplo-pheno analysis reveal superior haplotype combinations associated with low grain chalkiness under high temperature in rice. Front. Plant Sci. 2023, 14, 1133115. [Google Scholar] [CrossRef]
- Wang, E.; Wang, J.; Zhu, X.; Hao, W.; Wang, L.; Li, Q.; Zhang, L.; He, W.; Lu, B.; Lin, H.; et al. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat. Genet. 2008, 40, 1370–1374. [Google Scholar] [CrossRef] [PubMed]
- Raju, G.N.; Chand, N.; Bhashyam, M.K.; Srinivas, T. Predictive model for grain cracking in terms of rice plant and panicle morphology derived from multivariate analysis. J. Sci. Food Agric. 1995, 68, 141–152. [Google Scholar] [CrossRef]
- Cho, J. Double fertilization in Oryza sativa L. and development of the endosperm with special reference to the aleurone layer. Bull. Natl. Inst. Agric. Sci. 1956, 6, 61–101. [Google Scholar]
- Ekanayake, I.J.; Datta, S.; Steponkus, P.L. Spikelet Sterility and Flowering Response of Rice to Water Stress at Anthesis. Ann. Bot. 1989, 63, 257–264. [Google Scholar] [CrossRef]
- Jagadish, S.; Craufurd, P.; Wheeler, T. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 1627–1635. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Craufurd, P.Q.; Wheeler, T.R. Phenotyping Parents of Mapping Populations of Rice for Heat Tolerance during Anthesis. Crop Sci. 2008, 48, 1140. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Murty, M.V.R.; Quick, W.P. Rice responses to rising temperatures–challenges, perspectives and future directions. Plant Cell Environ. 2015, 38, 1686–1698. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive Expression Profiling of Rice Grain Filling-Related Genes under High Temperature Using DNA Microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef]
- Shi, P.; Tang, L.; Wang, L.; Sun, T.; Liu, L.; Cao, W.; Zhu, Y. Post-Heading Heat Stress in Rice of South China during 1981–2010. PLoS ONE 2015, 10, e0130642. [Google Scholar] [CrossRef]
- Shi, W.; Ishimaru, T.; Gannaban, R.B.; Oane, W.; Jagadish, S.V.K. Popular Rice (Oryza sativa L.) Cultivars Show Contrasting Responses to Heat Stress at Gametogenesis and Anthesis. Crop Sci. 2015, 55, 589–596. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Butardo, V.M.; Misra, G.; Cuevas, R.P.; Anacleto, R.; Kishor, P.B.K. Designing climate-resilient rice with ideal grain quality suited for high-temperature stress. J. Exp. Bot. 2015, 66, 1737–1748. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.A.; Resurreccion, A.P. Maintaining the yield of edible rice in a warming world. Funct. Plant Biol. 2009, 36, 1037–1045. [Google Scholar] [CrossRef]
- Ambardekar, A.A.; Siebenmorgen, T.J.; Counce, P.A.; Lanning, S.B.; Mauromoustakos, A. Impact of field-scale night-time air temperatures during kernel development on rice milling quality. Field Crops Res. 2011, 122, 179–185. [Google Scholar] [CrossRef]
- Wu, Y.C.; Chang, S.J.; Lur, H.S. Effects of field high temperature on grain yield and quality of a subtropical type japonica rice—Pon-Lai rice. Plant Prod. Sci. 2016, 19, 145–153. [Google Scholar] [CrossRef]
- Abayawickrama, A.; Reinke, R.; Fitzgerald, M.; Harper, J.; Burrows, G. Influence of high daytime temperature during the grain filling stage on fissure formation in rice. J. Cereal Sci. 2017, 74, 256–262. [Google Scholar] [CrossRef]
- Miah, M.N.H.; Yoshida, T.; Yamamoto, Y.; Nitta, Y. Characteristics of Dry Matter Production and Partitioning of Dry Matter to Panicles in High Yielding Semidwarf Indica and Japonica-Indica Hybrid Rice Varieties. Jpn. J. Crop Sci. 1996, 65, 672–685. [Google Scholar] [CrossRef][Green Version]
- Yang, J.; Peng, S.; Visperas, R.M.; Sanico, A.L.; Zhu, Q.; Gu, S. Grain filling pattern and cytokinin content in the grains and roots of rice plants. Plant Growth Regul. 2000, 30, 261–270. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Huang, Z.; Zhu, Q.; Wang, L. Remobilization of Carbon Reserves Is Improved by Controlled Soil-Drying during Grain Filling of Wheat. Crop Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Wang, W. Hormonal Changes in the Grains of Rice Subjected to Water Stress during Grain Filling. Plant Physiol. 2001, 127, 315–323. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Huang, Z.; Wang, Z.; Zhu, Q.; Liu, L. Correlation of Cytokinin Levels in the Endosperms and Roots with Cell Number and Cell Division Activity during Endosperm Development in Rice. Ann. Bot. 2002, 90, 369–377. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q. Hormones in the grains in relation to sink strength and postanthesis development of spikelets in rice. Plant Growth Regul. 2003, 41, 185–195. [Google Scholar] [CrossRef]
- Yang, W.; Jia, C.-C.; Howell, T. Relationship of Moisture Content Gradients and Glass Transition Temperatures to Head Rice Yield during Cross-Flow Drying. Biosyst. Eng. 2003, 86, 199–206. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J. Grain filling of cereals under soil drying. New Phytol. 2006, 169, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y. Chapter 13-Impact of climate change on rice grain quality. In Rice, 4th ed.; Bao, J., Ed.; AACC International Press: Washington, DC, USA, 2019; pp. 427–441. [Google Scholar]
- Zhang, H.; Chen, T.; Wang, Z.; Yang, J.; Zhang, J. Involvement of cytokinins in the grain filling of rice under alternate wetting and drying irrigation. J. Exp. Bot. 2010, 61, 3719–3733. [Google Scholar] [CrossRef]
- Nangju, D.; De Datta, S.K. Effect of Time of Harvest and Nitrogen Level on Yield and Grain Breakage in Transplanted Rice 1. Agron. J. 1970, 62, 468–474. [Google Scholar] [CrossRef]
- Seetanun, W.; De Datta, S.K. Grain Yield, Milling Quality, and Seed Viability of Rice as Influenced by Time of Nitrogen Application and Time of Harvest. Agron. J. 1973, 65, 390–394. [Google Scholar] [CrossRef]
- Atilio, J.B.; Causin, H.F. The central role of amino acids on nitrogen utilization and plant growth. J. Plant Physiol. 1996, 149, 358–362. [Google Scholar] [CrossRef]
- Perez, C.M.; Juliano, B.O.; Liboon, S.P.; Alcantara, J.M.; Cassman, K.G. Effects of late nitrogen fertilizer application on head rice yield, protein content, and grain quality of rice. Cereal Chem. 1996, 73, 556–560. [Google Scholar]
- Leesawatwong, M.; Jamjod, S.; Kuo, J.; Dell, B.; Rerkasem, B. Nitrogen Fertilizer Increases Seed Protein and Milling Quality of Rice. Cereal Chem. 2005, 82, 588–593. [Google Scholar] [CrossRef]
- Yang, Y.H.; Dai, L.; Xia, H.C.; Zhu, K.M.; Liu, H.J.; Chen, K.P. Protein profile of rice (Oryza sativa L.) seeds. Genet. Mol. Biol. 2013, 36, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.; Sajitz-Hermstein, M.; Nikoloski, Z. Effects of Varying Nitrogen Sources on Amino Acid Synthesis Costs in Arabidopsis thaliana under Different Light and Carbon-Source Conditions. PLoS ONE 2015, 10, e0116536. [Google Scholar] [CrossRef] [PubMed]
- Bryant, R.J.; Yeater, K.M.; McClung, A.M. Effect of Nitrogen Rate and the Environment on Physicochemical Properties of Selected High-Amylose Rice Cultivars. Cereal Chem. 2015, 92, 604–610. [Google Scholar] [CrossRef]
- Beeny, J.M.; Chin, S.N. Multipass Drying of Paddy (Rice) in the Humid Tropics. J. Agric. Eng. Res. 1970, 15, 364–374. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, H. Effect of quality characteristics on brown rice produced from paddy rice with different moisture contents. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 289–293. [Google Scholar] [CrossRef]
- Siebenmorgen, T.J.; Nehus, Z.T.; Archer, T.R. Milled Rice Breakage Due to Environmental Conditions. Cereal Chem. 1998, 75, 149–152. [Google Scholar] [CrossRef]
- Siebenmorgen, T.J.; Yang, W.; Sun, Z. Glass transition temperature of rice kernels determined by dynamic mechanical thermal analysis. Trans. Am. Soc. Agric. Biol. Eng. 2004, 47, 835–839. [Google Scholar] [CrossRef]
- Srikaeo, K.; Boonrod, C.; Rahman, M.S. Effect of storage temperatures on the head rice yield in relation to glass transition temperatures and un-freezable water. J. Cereal Sci. 2016, 70, 164–169. [Google Scholar] [CrossRef]
- Truong, T.; Truong, V.; Fukai, S.; Bhandari, B. Changes in Cracking Behavior and Milling Quality of Selected Australian Rice Varieties Due to Postdrying Annealing and Subsequent Storage. Dry. Technol. 2012, 30, 1831–1843. [Google Scholar] [CrossRef]
- Perdon, A.A.; Siebenmorgen, T.J.; Mauromoustakos, A.; Griffin, V.K.; Johnson, E.R. Degree of Milling Effects on Rice Pasting Properties. Cereal Chem. 2001, 78, 205–209. [Google Scholar] [CrossRef]
- Mei, H.-W.; Luo, L.-J.; Guo, L.-B.; Wang, Y.-P.; Yu, X.-Q.; Ying, C.-S.; Li, Z.-K. Molecular mapping of QTLs for rice milling yield traits. Acta Genet. Sin. 2002, 29, 791–797. [Google Scholar] [PubMed]
- Septiningsih, E.M.; Trijatmiko, K.R.; Moeljopawiro, S.; McCouch, S.R. Identification of quantitative trait loci for grain quality in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O. rufipogon. Theor. Appl. Genet. 2003, 107, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Tsuzuki, E.; Lin, D.; Kamiunten, H.; Terao, H.; Matsuo, M.; Cheng, S. Molecular genetic mapping of quantitative trait loci for milling quality in rice (Oryza sativa L.). J. Cereal Sci. 2004, 40, 109–114, ISSN 0733-5210. [Google Scholar] [CrossRef]
- Zheng, T.Q.; Xu, J.L.; Li, Z.K.; Zhai, H.Q.; Wan, J.M. Genomic regions associated with milling quality and grain shape identified in a set of random introgression lines of rice (Oryza sativa L.). Plant Breed. 2007, 126, 158–163. [Google Scholar] [CrossRef]
- Kepiro, J.; McClung, A.; Chen, M.; Yeater, K.; Fjellstrom, R. Mapping QTLs for milling yield and grain characteristics in a tropical japonica long grain cross. J. Cereal Sci. 2008, 48, 477–485. [Google Scholar] [CrossRef]
- Jiang, G.-H.; Hong, X.-Y.; Xu, C.-G.; Li, X.-H.; He, Y.-Q. Identification of Quantitative Trait Loci for Grain Appearance and Milling Quality Using a Doubled-Haploid Rice Population. J. Integr. Plant Biol. 2005, 47, 1391–1403. [Google Scholar] [CrossRef]
- Hao, W.; Zhu, M.-Z.; Gao, J.-P.; Sun, S.-Y.; Lin, H.-X. Identification of Quantitative Trait Loci for Rice Quality in a Population of Chromosome Segment Substitution Lines. J. Integr. Plant Biol. 2009, 51, 500–512. [Google Scholar] [CrossRef]
- Nelson, J.C.; McClung, A.M.; Fjellstrom, R.G.; Moldenhauer, K.A.K.; Boza, E.; Jodari, F.; Oard, J.H.; Linscombe, S.; Scheffler, B.E.; Yeater, K.M. Mapping QTL main and interaction influences on milling quality in elite US rice germplasm. Theor. Appl. Genet. 2011, 122, 291–309. [Google Scholar] [CrossRef]
- Nelson, J.C.; Jodari, F.; Roughton, A.I.; McKenzie, K.M.; McClung, A.M.; Fjellstrom, R.G.; Scheffler, B.E. QTL mapping for milling quality in elite western U.S. rice germplasm. Crop Sci. 2012, 52, 242–252. [Google Scholar] [CrossRef]
- Wang, X.; Pang, Y.; Wang, C.; Chen, K.; Zhu, Y.; Shen, C.; Ali, J.; Xu, J.; Li, Z. New Candidate Genes Affecting Rice Grain Appearance and Milling Quality Detected by Genome-Wide and Gene-Based Association Analyses. Front. Plant Sci. 2017, 7, 1998. [Google Scholar] [CrossRef]
- Qiu, X.; Pang, Y.; Yuan, Z.; Xing, D.; Xu, J.; Dingkuhn, M.; Li, Z.; Ye, G. Genome-Wide Association Study of Grain Appearance and Milling Quality in a Worldwide Collection of Indica Rice Germplasm. PLoS ONE 2016, 10, e0145577. [Google Scholar] [CrossRef] [PubMed]
- Quero, G.; Gutiérrez, L.; Monteverde, E.; Blanco, P.; de Vida, F.P.; Rosas, J.; Fernández, S.; Garaycochea, S.; McCouch, S.; Berberian, N.; et al. Genome-Wide Association Study Using Historical Breeding Populations Discovers Genomic Regions Involved in High-Quality Rice. Plant Genome 2018, 11, 170076. [Google Scholar] [CrossRef]
- Misra, G.; Anacleto, R.; Badoni, S.; Butardo, V.; Molina, L.; Graner, A.; Demont, M.; Morell, M.K.; Sreenivasulu, N. Dissecting the genome-wide genetic variants of milling and appearance quality traits in rice. J. Exp. Bot. 2019, 70, 5115–5130. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhan, J.; Li, J.; Lu, X.; Liu, J.; Wang, Y.; Zhao, Q.; Ye, G. Genome-Wide Association Study (GWAS) for Mesocotyl Elongation in Rice (Oryza sativa L.) under Multiple Culture Conditions. Genes 2019, 11, 49. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Liu, Y.; Gong, C.; Chen, B.; Wang, T. Waxy is an important factor for grain fissure resistance and head rice yield as revealed by a genome-wide association study. J. Exp. Bot. 2022, 73, 6942–6954. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.U.; Ye, J.H.; Yang, Y.Y.; Li, R.S.; Zhen, L.I.; Shan, W.A.N.G.; Yang, Y.L. Genetic diversity analysis and GWAS reveal the adaptive loci of milling and appearance quality of japonica rice (Oryza sativa L.) in Northeast China. J. Integr. Agric. 2022, 21, 1539–1550. [Google Scholar] [CrossRef]
- Yu, P.; Ye, C.; Li, L.; Yin, H.; Zhao, J.; Wang, Y.; Zhang, Z.; Li, W.; Long, Y.; Hu, X.; et al. Genome-wide association study and genomic prediction for yield and grain quality traits of hybrid rice. Mol. Breed. 2022, 42, 16. [Google Scholar] [CrossRef]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef]
- Zhou, X.; Stephens, M. Efficient multivariate linear mixed model algorithms for genome-wide association studies. Nat. Methods 2014, 11, 407–409. [Google Scholar] [CrossRef]
- Singh, R.; Saripalli, G.; Gautam, T.; Kumar, A.; Jan, I.; Batra, R.; Kumar, J.; Kumar, R.; Balyan, H.S.; Sharma, S.; et al. Meta-QTLs, ortho-MetaQTLs and candidate genes for grain Fe and Zn contents in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2022, 28, 637–650. [Google Scholar] [CrossRef]
- Joukhadar, R.; Thistlethwaite, R.; Trethowan, R.; Keeble-Gagnère, G.; Hayden, M.J.; Ullah, S.; Daetwyler, H.D. Meta-analysis of genome-wide association studies reveal common loci controlling agronomic and quality traits in a wide range of normal and heat stressed environments. Theor. Appl. Genet. 2021, 134, 2113–2127. [Google Scholar] [CrossRef]
- Joukhadar, R.; Daetwyler, H.D.; Bansal, U.K.; Gendall, A.R.; Hayden, M.J. Genetic Diversity, Population Structure and Ancestral Origin of Australian Wheat. Front. Plant Sci. 2017, 8, 2115. [Google Scholar] [CrossRef] [PubMed]
- Jighly, A.; Benhajali, H.; Liu, Z.; Goddard, M.E. MetaGS: An accurate method to impute and combine SNP effects across populations using summary statistics. Genet. Sel. Evol. 2022, 54, 37. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Monteverde, E.; Rosas, J.E.; Blanco, P.; de Vida, F.P.; Bonnecarrère, V.; Quero, G.; Gutierrez, L.; McCouch, S. Multienvironment Models Increase Prediction Accuracy of Complex Traits in Advanced Breeding Lines of Rice. Crop Sci. 2018, 58, 1519–1530. [Google Scholar] [CrossRef]
- Monteverde, E.; Gutierrez, L.; Blanco, P.; Pérez de Vida, F.; Rosas, J.E.; Bonnecarrère, V.; McCouch, S. Integrating molecular markers and environmental covariates to interpret genotype by environment interaction in rice (Oryza sativa L.) grown in subtropical areas. G3 Genes Genomes Genet. 2019, 9, 1519–1531. [Google Scholar]
- Jighly, A.; Thayalakumaran, T.; O’Leary, G.J.; Kant, S.; Panozzo, J.; Aggarwal, R.; Hessel, D.; Forrest, K.L.; Technow, F.; Tibbits, J.F.G.; et al. Using genomic prediction with crop growth models enables the prediction of associated traits in wheat. J. Exp. Bot. 2022, 74, 1389–1402. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Countries | Head Rice Yields (Million Metric Tons) |
|---|---|
| China | 148.30 |
| India | 122.27 |
| Indonesia | 35.30 |
| Bangladesh | 34.60 |
| Vietnam | 27.38 |
| Thailand | 18.86 |
| Burma | 12.60 |
| Philippines | 12.42 |
| Pakistan | 8.42 |
| Brazil | 8 |
| Plant Material | QTL | Traits | Cloning | Chromosome Number | References |
|---|---|---|---|---|---|
| Rice | lk3 | Grain length | 3 | [32] | |
| Rice | gw3.1, gw3.2, gw9.1 | Grain weight | 3 | ||
| Rice | gw3.1 | Grain weight | Fine mapped | 3 | [33] |
| Rice | GS3 | grain length and weight | cloned | 3 | |
| Rice | GW2 | Control grain width and weight | cloned | 2 | [34] |
| Rice | Lk-4 | Grain length | Fine mapped | 4 | [35] |
| Rice | GW5 | Grain width and weight | Fine mapped | 5 | [36] |
| Rice | DEP1 | Reduce length of panicle for better grain-filling | 1 | [37] | |
| Rice | qGL7 | Grain length and size | 7 | [38] | |
| Rice | GL3.1 | Grain length and weight | cloned | 3 | [39] |
| Rice | qGL4b | Grain length and weight | 4 | [40] | |
| Rice | qSS7 | Length, width and weight | 7 | [41] | |
| Rice | GW1, GS3, GS7 | Grain weight and size | Fine mapped | 1, 3, 7 | [42] |
| Rice | GW7 | Grain width | OsSNB | 7 | [43] |
| Rice | GW2, Gl12 | Grain length and width | Fine mapped | 2, 12 | [44] |
| Rice | TGW6 | Thousand grain weight | cloned | 6 | [45] |
| Rice | qgl3 | Grain length, filling and weight | cloned | 3 | [46] |
| Rice | * Srs5 | Cell elongation | 5 | [47] | |
| Rice | GS2 | Grain weight | cloned | 2 | [48] |
| Rice | GS5-1, GS5-2 | Grain width, filling and weight | 5 | [49] | |
| Rice | * BG1 | Seed weight | cloned | 1 | [50] |
| Rice | * BG1 | Source-size sink and heading date | |||
| Rice | GLW7 | Grain length | Cloned OsSPL13 | 7 | [51] |
| Rice | GL1.3 | Grain length and weight | Fine mapped | 1 | [52] |
| Rice | GL-6 | Grain length | Fine mapped | 6 | [53] |
| Rice | TGW12 | Grain weight | Fine mapped | 12 | [54] |
| Rice | MQTL3.1 | Grain weight | Meta QTL | 3 | [55] |
| Plant Material | QTL | Traits | Chromosome Number | References |
|---|---|---|---|---|
| Seven indica (T65,N8,108,C8669,C8005,221,T65wx), three glaberrima (W025,GM1,GM2) | Wx gene (Wx a, Wx b) | Amylose | 6 | [67] |
| Kinmaze and its mutant lines | Flo-2 | Regulates RBE1 gene for starch synthesis | 6 | |
| Japonica | Soluble starch synthase | Starch | [71] | |
| 132 DH population via anther culture of F1 hybrid between ZYQ8 (indica) and JX17 (japonica) | qAC-5, Wx | Amylose content | 5, 6 | |
| Nipponbare, Koshiibuki, Tentakaku, Sasanishiki and Hatsuboshi | GBSSI, GBSSII, SSI, SSIIa, SSIIb, SSIIc, SSIIIa, SSIIIb, SSICa, SSIVb | Starch synthase under temperature stress | 6, 7, 6, 6, 2, 10, 8, 4, 1, 5 | [72] |
| BEI, BEIIb, | Branching enzyme | 6, 2 | ||
| ISA1, ISA3, PUL | Debranching enzyme | 8, 9, 4 | ||
| Amy1A, Amy2A, Amy3A, Amy3C, Amy3D, Amy3E, Amy4A | Alpha-Amylase | 2, 6, 9, 9, 8, 8, 1 | ||
| Ramy1 | Transcription factor for amylase gene | 1 | ||
| Nipponbare | Flo-2 | Starch quality and grain size | 6 | [73] |
| chromosome segment substitution lines of a cross between 9311 and Nipponbare | starch paste viscosity characteristics | Rapid viscos analyzer profile of rice genotypes | 1, 2, 4, 5, 6, 8, 10 | [74] |
| Plant Material | QTL | Traits | Chromosome Number | References |
|---|---|---|---|---|
| 210 F10 RIL population derived from Zhenshan 97 (maternal) and Minghui 63 (paternal) | Markers (C922, C1016, R1629 RM228, R19, C909B, TEL3, R496) | Protein % | 1, 4, 8, 10, 3, 11, 12 | [82] |
| 312 Double haploids from BC3F1 i.e., derived from Caiapo (Brazillian, long grain) × Oryza glaberrima, IRGC 103544 | Pro1, pro2, pro6, pro11 | Protein % | 1, 2, 6, 11 | |
| 81 DH lines from the F1 hybrid of a cross between Gui 630 (indica) × 02428 (japonica) | qRPC-1, qRPC-4, qRPC6, qRPC-7, qRPC-10 | Protein content | 1, 4, 6, 7, 10 | [83] |
| 71 RILs, derived from a cross between Asominori × IR24 | qGLB-1, qGLB-2.1, qGLB-2.2, qGLB-5 | Globulin | 1, 2, 2, 5 | |
| qPLA-1, qPLA-3, qPLA-10 | Prolamin | 1, 3, 10 | [84] | |
| qGLT-2, qGLT-10, qGLT-11, qGLT-12 | Glutelin | 2, 10, 11, 12 | ||
| Two BC3F2 populations, NILs and 197 accessions (indica and japonica) | * qPC1 | Glutelins, prolamins, globulins, albumins and starch. encodes a putative amino acid transporter OsAAP6 | 1 | [85] |
| 25 BC1F1 lines were developed by crossing “ARC 10075” × Naveen | qGPC1.1, qSGPC2.1, qSGPC7.1 | Grain protein content | 1, 2, 7 | [86] |
| Mutants rice lines | rice asparagine synthetase 1 | Grain protein content and grain yield | 1 | [87] |
| 193 recombinant inbred lines were developed by crossing Japonica rice Nipponbare with Indica super rice YK17 | qPC6, qPC7 and qGLU6 | Crude protein and protein contents | 6, 7 | [88] |
| Plant Material | QTL | Traits | Chromosome Number | References |
|---|---|---|---|---|
| 81 DH lines from the F1 hybrid of a cross between Gui 630 (indica) × 02428 (japonica) | qRFC-1, qRFC-2, qRFC-5 | Fat content | 1, 2, 5 | [98] |
| 71 RIL cross of Asominori (japonica) × IR24 (indica) | qFC-1-2, qFI-1-2, qFC-2-2, qFI-2, qFC-4, qFI-4, qFC-9, qFI-9 | Fat contents and fat index | 1, 1, 2, 2, 4, 4, 9, 9 | [99] |
| 190 DH lines, derived From anther culture of an F1 hybrid between Wuyujing2 (japonica) × Zhanshen 97B (indica). | qCFC1a, qCFC1c, qCFC3a, qCFC3c, qCFC5abc, qCFC6a, qCFC7ac, qCFC7b, qCFC8b, qCFC9a | Fat content in brown rice | 1, 1, 3, 3, 5, 6, 7, 7, 8, 9 | [100] |
| RIL population derived from Xieqingzao B × Milyang 46 | qFC-3, qFC-5, qFC-6, qFC-8 | Crude fat content | 3, 5, 6, 8 | [101] |
| 190 DH lines derived from F1 hybrid Wuyujing2 (japonica) × Zhanshen 97B (indica). BCF1 developed by crossing DH with both parents: 149 from Zhanshen 97B and 143 from Wuyujing2 | qCFC5 detected in both populations: DH and BCF1 | Crude Fat content | 5 | |
| 120 DH lines developed by anther culture, F1 cross Samgang × Nagdong | qLC6.1, qLC7.1, qLC9.1 | Lipid content brown rice | 6, 7, 9 | [102] |
| 182 backcross inbred lines (BILs), backcross (“Koshihikari” × “Kasalath”) and 39 chromosome segment substitution lines (CSSL) | qFC7.1 and qFC7.2 | Crude fat content brown rice | 7 | [103] |
| pal4, ara5, oil3 encodes enzymes: Ketoacyl–ACP Synthase I(KAS I), Ketoacyl–CoA Synthase (FAE1) | Fatty acid composition | 4, 5, 3 | [104] | |
| pal6, ste4, ole1, oil1 encodes enzymes: Acyl–ACP Thioesterase (FatB), Acyl–CoA Thioesterase, ER 2-Lysophosphatidate Acyltransferase (LPAAT) | 6, 4, 1 | |||
| myr2-2, lin6-1, ole6, lin6-2 encode enzyme: Acyl–CoA:Diacylglycerol Acyltransferase (DGAT) | 2, 6, 6 | |||
| 133 DH lines cross Cheongcheong × Nagdong | qLip-3, qLip-6 | Lipid contents brown rice | 3, 6 | [105] |
| 533 diverse O. sativa accessions | PAL6, LIN6, MYR2, and ARA6 | Oil biosynthesis in rice | 2, 6 | [106] |
| Rice mutant population | OsLTPL23 | Lipid transfer protein | 1, 2 | [107] |
| Plant Material | QTL | Traits | Chromosome Number | References |
|---|---|---|---|---|
| 132 DH population via anther culture of F1 hybrid between ZYQ8 (indica) and JX17 (japonica) | Ratio of white-core kernels to total number of observed kernels | qPGWC-8, qPGWC-12 | 8, 12 | [124] |
| Ratio square of the white core to square of white-core kernel. | qSWC-3 | 3 | ||
| 241 F2:3 segregating population | C161-R753, RG360-C734a, RG528-C1447, R1952-C226, R2625-C223 | Chalkiness in milled rice | 3 | |
| 238 F10 (Recombinant Inbred lines) | RG360-C734a, R1245-R1789, RG360-C734a, Wx-R1952 | Brown rice white belly (First two marker intervals), brown rice white core (Second two marker intervals) | 3 | |
| 66 CSSL lines (BC3F1), derived from the cross Asominori IR24 (Asominori recurrent parent) | qPGWC-8, qPGWC-9, qPGWC-1, qPGWC-7 | Percentage of grains with chalkiness (PGWC) | 1, 5, 5, 6, 10 | [125] |
| qACE-8, qACE-2, qACE-9 | Area of chalky endosperm (ACE) | 8, 9 | ||
| qDEC-8, qDEC-9, qDEC-1a, qDEC-1b, qDEC-2 | Degree of endosperm chalkiness (DEC) | 5, 7, 5, 6 | ||
| NIL (near-isogenic lines), Transgenic | Chalk 5 | Increased chalkiness | 8, 9, 1 | [126] |
| two single-segment substitution lines | qPGC5, qPGC6 | Lower percentage of chalkiness | 5, 6 | [127] |
| 184 BC5F2 lines were developed by crossing indica cultivar J23B (the recurrent parent) and a japonica cultivar BL130 | qWCR4 | White core rate | 4 | [128] |
| Meta QTL analysis | 64 MetaQTL | Grain chalkiness | 1, 2, 3, 4, 5, 6, 7, 9, 10, 11, 12 | [129] |
| Plant Material | QTLs | Chromosome Number | References | Climate Zone |
|---|---|---|---|---|
| 212 RILs developed by crossing Lemont (japonica) × Teqing (Indica) | QHr6, QHr7 | 6, 7 | [170] | Temperate zone (China) |
| 312 double haploids of an interspecific cross of O. sativa × O. glaberrima. | hr1,hr3,hr6,hr8, hr11 | 1, 3, 6, 8 | [171] | Tropical Zone (Columbia; Tropic of Cancer) |
| 165 F6 RILs of a cross between Asominori (japonica) × IR24 (indica). | qMHP-1,MHP-3, qMHP-5 | 1, 3, 5 | [172] | Temperate zone (Japan) |
| 190 DH lines from the cross of Zhenshan 97 (indica) × WYJ-2 (japonica) | RM570-RM85 RM38-RM25 | 3, 8 | [83] | Temperate zone (China) |
| 254 BC4F1 of a cross between Teqing (Indica) × Lemont (Japonica) | QHr1, QHr5, QHr6 | 1, 5, 6 | [173] | Temperate zone (China) |
| 137 F11 RILs developed from a cross of two long-grain rice Cypress (japonica) × Panda (japonica) | hr6 | 6 | [174] | Temperate zone (USA) |
| 286 F8 RILs developed from a cross between Chuan7 (Indica) and Nanyangzhan (japonica) | qHRR-3 | 3 | [175] | Temperate zone (China) |
| RM6283 and RM16 * | 3 | |||
| 154 BC3F2 derived from cross between Koshihikari (japonica) and Nona Bokra (Indica) | qHRP-1, qHRP-6, qHRP-8 | 1, 6, 8 | [176] | Temperate zone (China) |
| MY1: 129 RILs derived from (Cypress × RT0034) grown in 2005 on four locations), and MY2: 298 RILs (Cypress × LaGrue) grown in 2006, 2007 on four locations) | qhr_6.1, qhr_9.1 (MY1) | 6, 9 | [177] | Temperate zone (USA) |
| qhr_1.1, qhr_5.1, qhr_9.1, qhr_10.1 (MY2) | 1, 5, 9, 10 | |||
| 231 RILs derived from L-204 (japonica) × 01Y110 (japonica) | qhr_6.1, qhr_6.2, qhr_8.1, qhr_9.1, qhr_9.2, qhr_10.1, qhr_11.1 | 6, 8, 9, 10, 11 | [178] | Temperate zone (USA) |
| 258 rice accessions (Indica, japonica, aus, aromatic) | qHMRR3, qHMRR9 | 3, 9 | [179] | Temperate zone (China) |
| 529 plants F2 population were developed by crossing Koshihikari (japonica) with Nona Bokra, (indica) | HRY10 | 10 | [116] | Temperate zone (China) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, F.; Jighly, A.; Joukhadar, R.; Niazi, N.K.; Al-Misned, F. Current Status and Future Prospects of Head Rice Yield. Agriculture 2023, 13, 705. https://doi.org/10.3390/agriculture13030705
Ali F, Jighly A, Joukhadar R, Niazi NK, Al-Misned F. Current Status and Future Prospects of Head Rice Yield. Agriculture. 2023; 13(3):705. https://doi.org/10.3390/agriculture13030705
Chicago/Turabian StyleAli, Fawad, Abdulqader Jighly, Reem Joukhadar, Nabeel Khan Niazi, and Fahad Al-Misned. 2023. "Current Status and Future Prospects of Head Rice Yield" Agriculture 13, no. 3: 705. https://doi.org/10.3390/agriculture13030705
APA StyleAli, F., Jighly, A., Joukhadar, R., Niazi, N. K., & Al-Misned, F. (2023). Current Status and Future Prospects of Head Rice Yield. Agriculture, 13(3), 705. https://doi.org/10.3390/agriculture13030705