
QTL Mining and Validation of Grain Nutritional Quality Characters in Rice (Oryza sativa L.) Using Two Introgression Line Populations



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Field Experiments
2.2. Quantitative Evaluation of Albumin, Globulin, Prolamin, Glutelin, and Grain Protein Contents
2.3. QTL Fingerprinting, Genetic Linkage Map Assembly, and Validation
3. Results
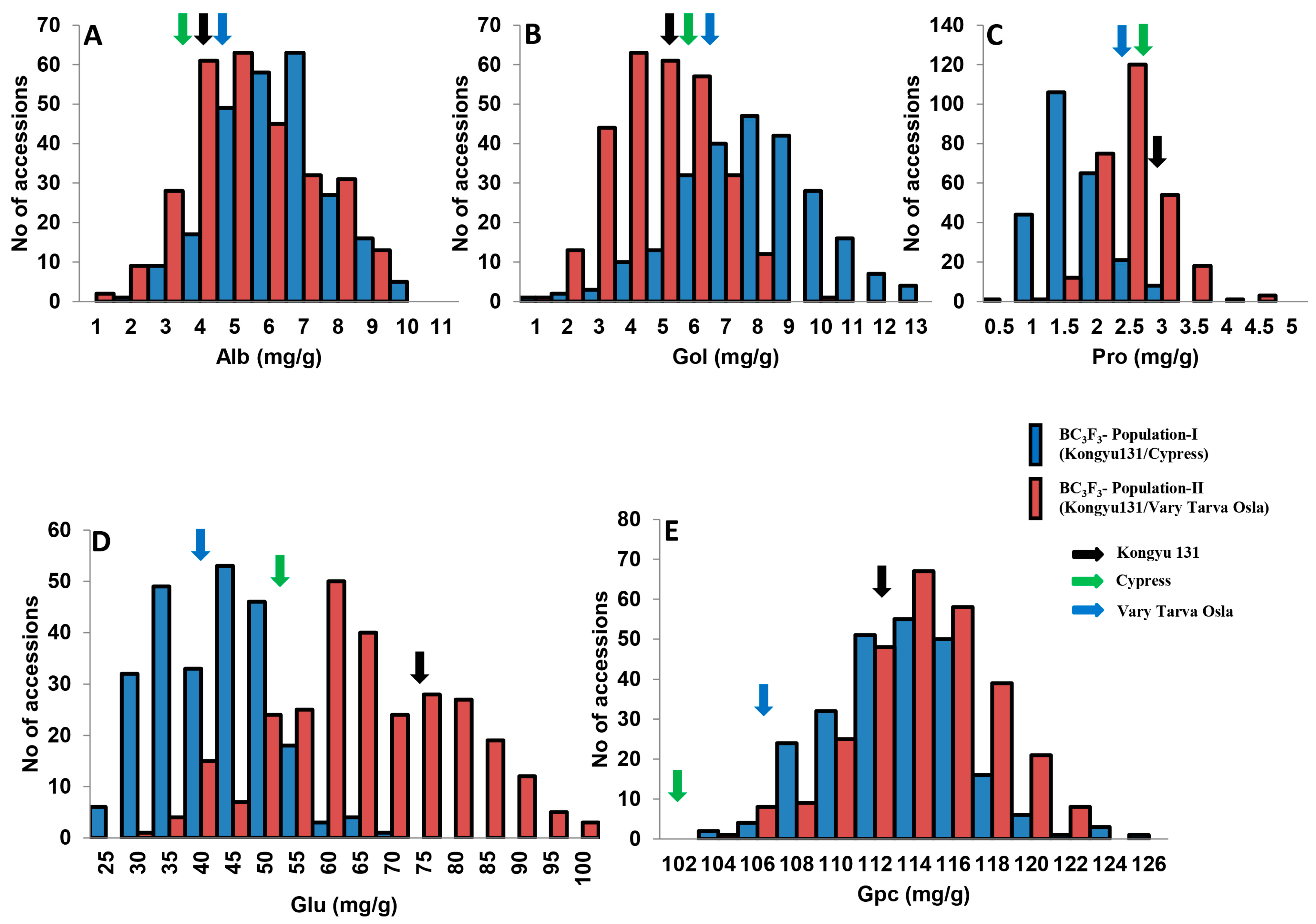
3.1. Phenotypic Variations in Nutritional Quality Traits in the Two ILs Populations
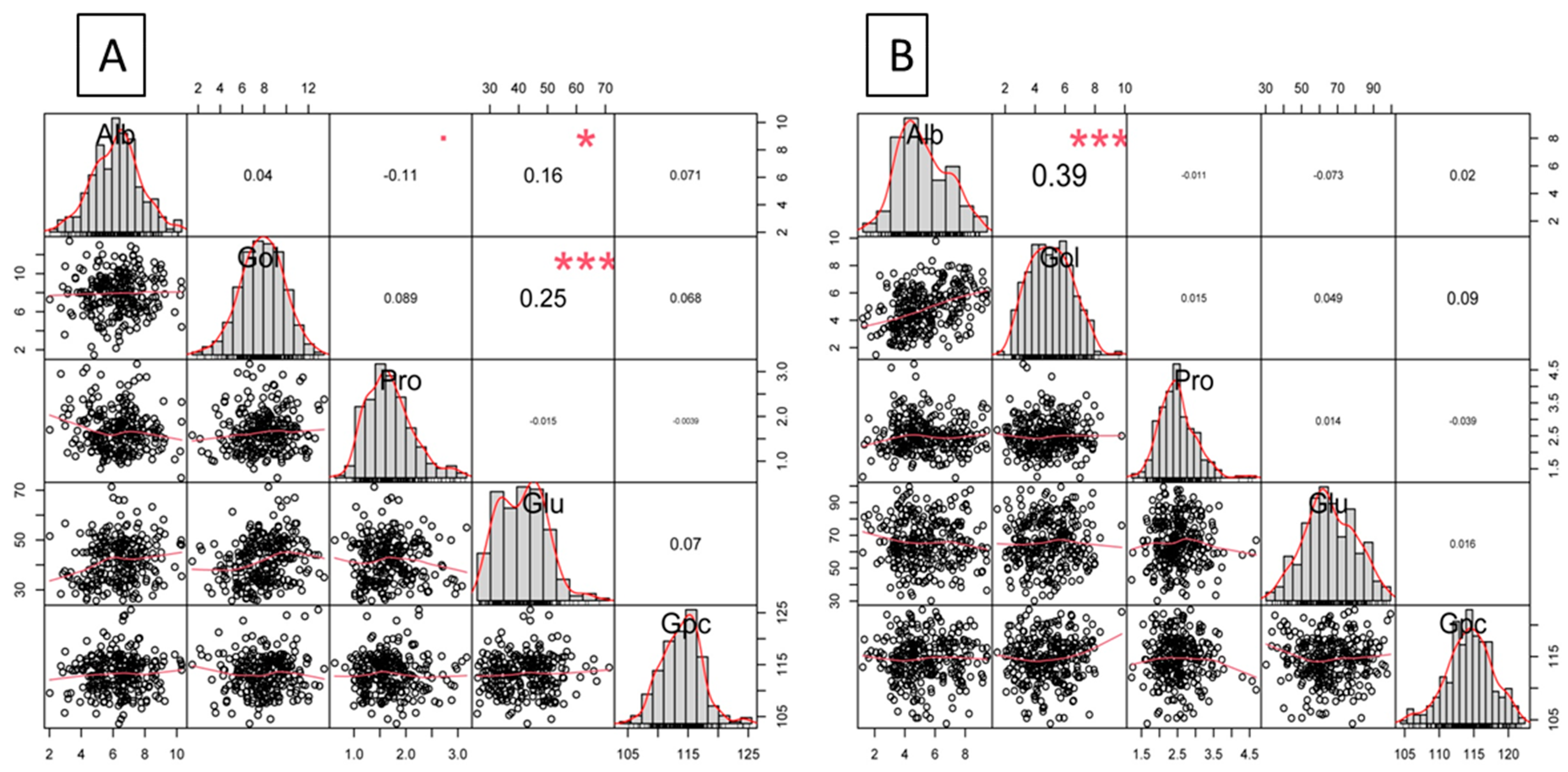
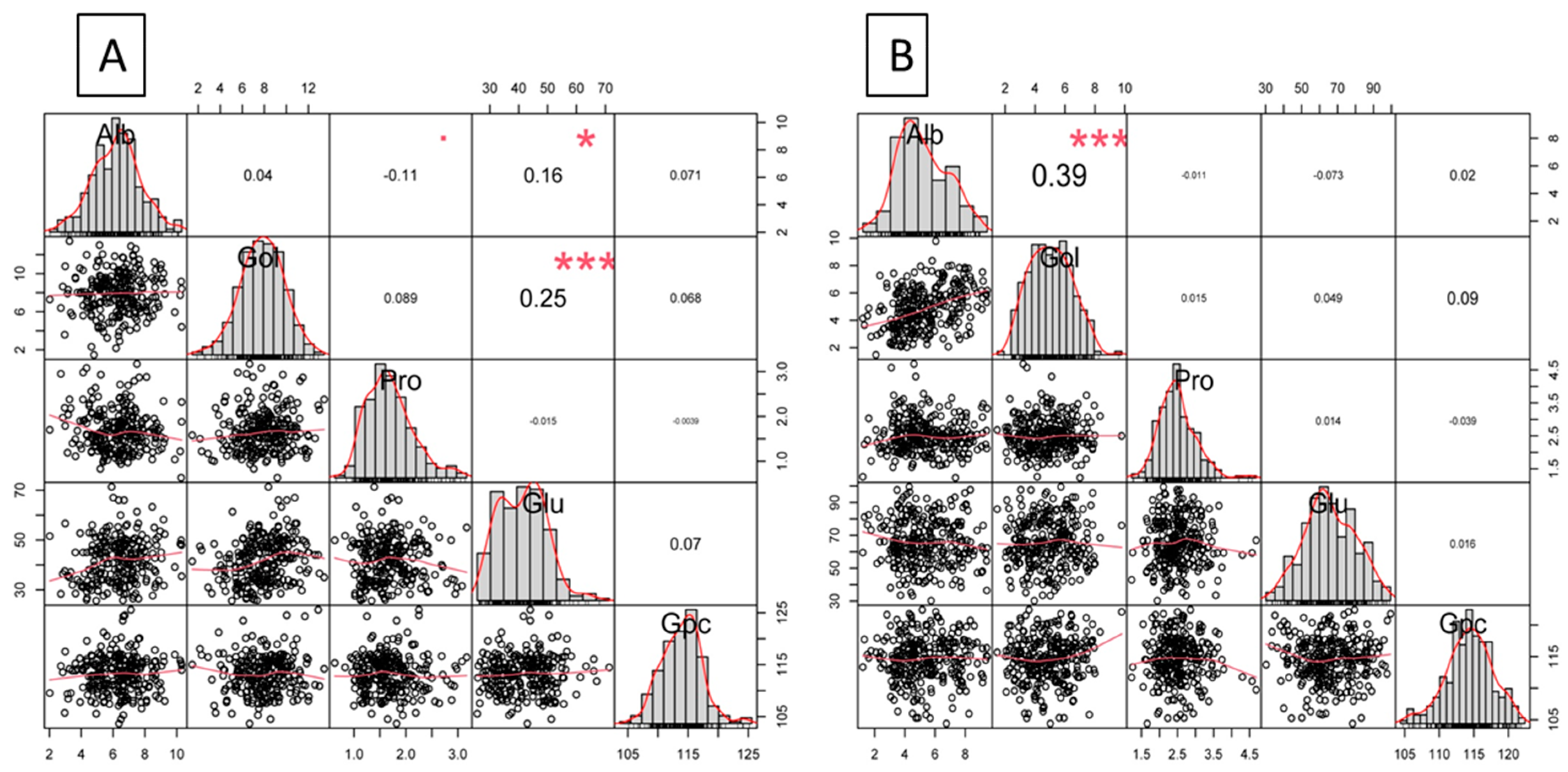
3.2. Correlation Delineation
3.3. QTL Fingerprinting in Albumin, Globulin, Prolamin, Glutelin, and Grain Protein Content
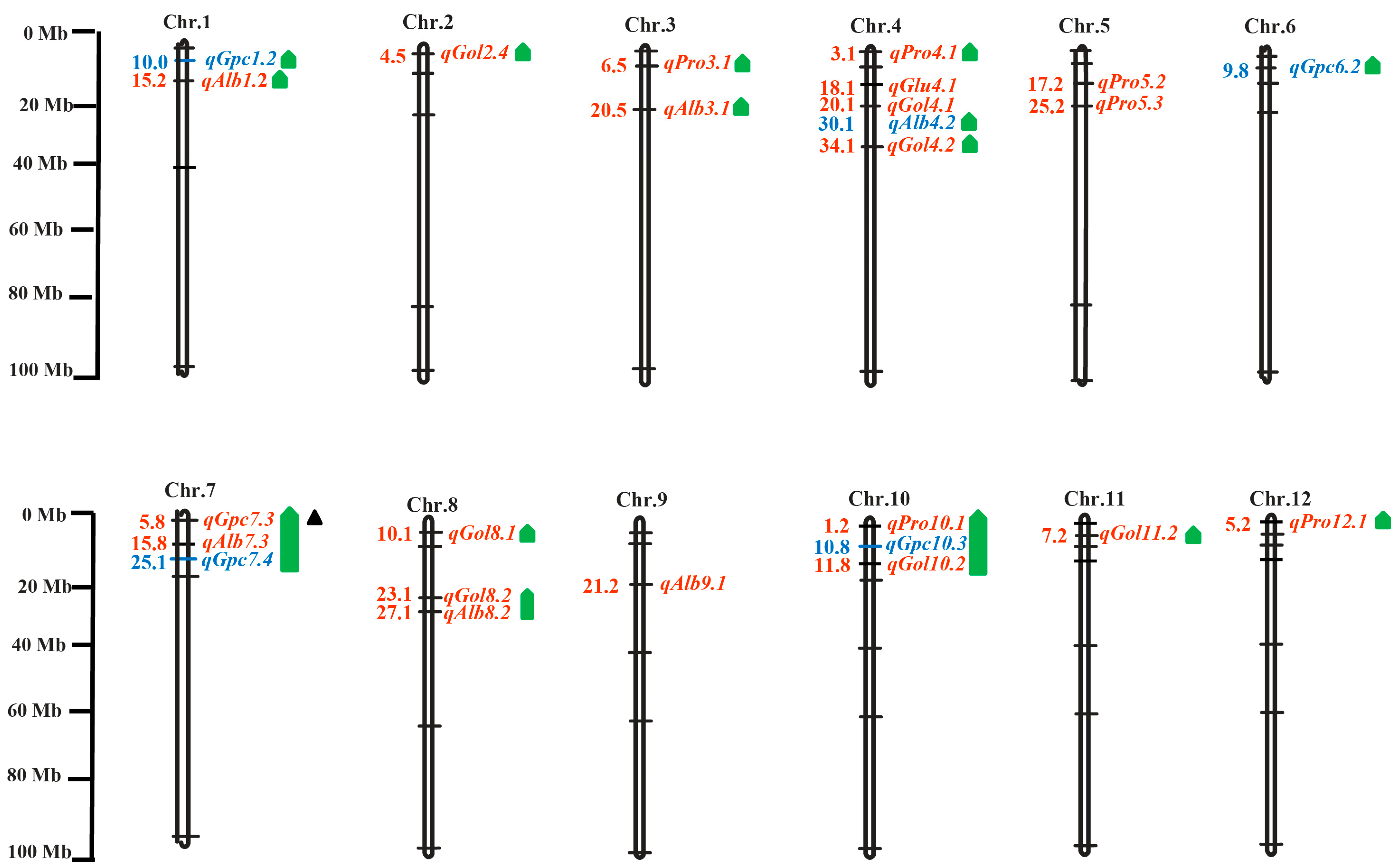
3.4. Co-Localization of Linked Positions with Previously Reported QTLs/Genes in Multiple Nutritional Quality Traits
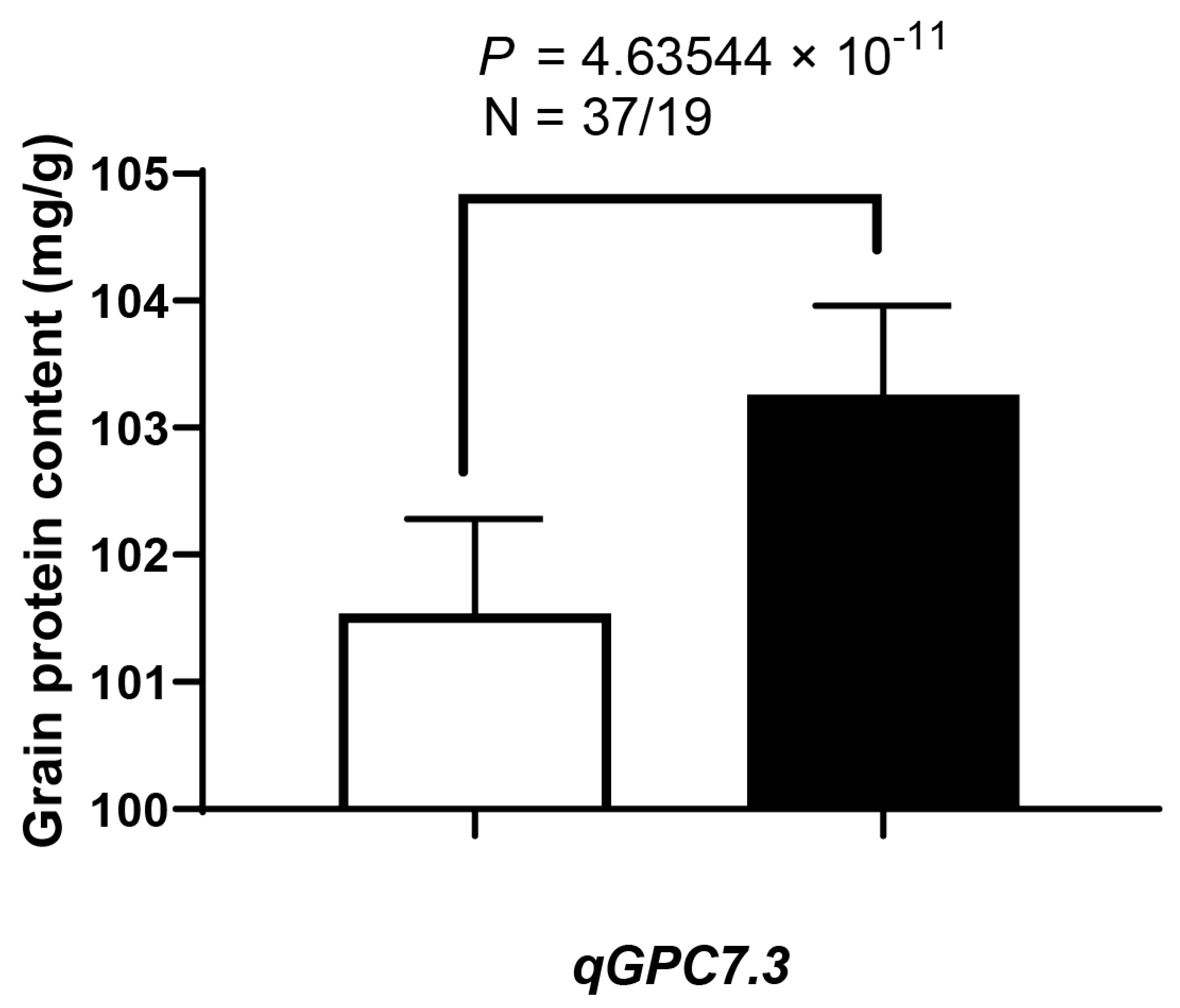
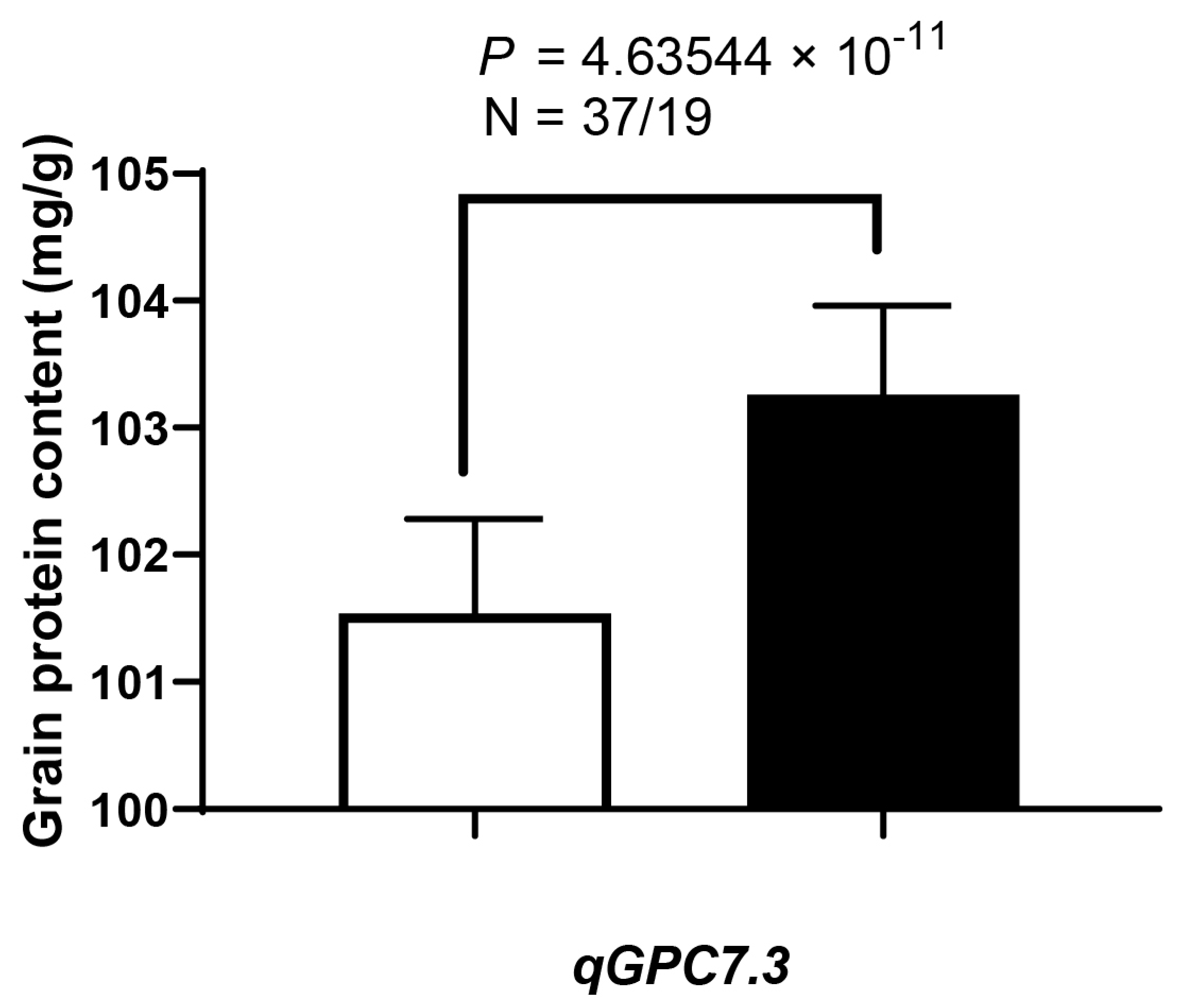
3.5. QTL Validation (qGpc7.3) and Co-Localization with Previously Reported QTLs
4. Discussion
4.1. Phenotypic Variations in Nutritional Quality Traits
4.2. QTL Localization and Pleiotropic Associations of Alb, Gol, Pro, Glu, and Gpc Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for global nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Butardo, V.M.; Sreenivasulu, N.; Juliano, B.O. Improving rice grain quality: State-of-the-art and future prospects. In Rice Grain Quality: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2019; pp. 19–55. [Google Scholar]
- Birla, D.S.; Malik, K.; Sainger, M.; Chaudhary, D.; Jaiwal, R.; Jaiwal, P.K. Nutrition. Progress and challenges in improving the nutritional quality of rice (Oryza sativa L.). Crit. Rev. Food Sci. Nutr. 2017, 57, 2455–2481. [Google Scholar] [CrossRef]
- Peng, B.; Kong, H.; Li, Y.; Wang, L.; Zhong, M.; Sun, L.; Gao, G.; Zhang, Q.; Luo, L.; Wang, G. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 2014, 5, 4847. [Google Scholar] [CrossRef]
- Lin, R.; Luo, Y.; Liu, D.; Huang, C. Determination and analysis on principal qualitative characters of rice germplasm. In Rice Germplasm Resources in China; Agricultural Science and Technology Publisher of China: Beijing, China, 1993; pp. 83–93. [Google Scholar]
- Zhou, L.; Liu, Q.; Zhang, C.; Xu, Y.; Tang, S.; Gu, M. Variation and distribution of seed storage protein content and composition among different rice varieties. Acta Agron. Sin. 2009, 35, 884–891. [Google Scholar] [CrossRef]
- Chen, P.; Lou, G.; Wang, Y.; Chen, J.; Chen, W.; Fan, Z.; Liu, Q.; Sun, B.; Mao, X.; Yu, H.; et al. The genetic basis of grain protein content in rice by genome-wide association analysis. Mol. Breed. 2022, 43, 1. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. Structure, chemistry, and function of the rice grain and its fractions. Cereal Foods World 1992, 37, 772–779. [Google Scholar]
- Jayaprakash, G.; Bains, A.; Chawla, P.; Fogarasi, M.; Fogarasi, S. A Narrative Review on Rice Proteins: Current Scenario and Food Industrial Application. Polymers 2022, 14, 3003. [Google Scholar] [CrossRef]
- He, W.; Wang, L.; Lin, Q.; Yu, F. Rice seed storage proteins: Biosynthetic pathways and the effects of environmental factors. J. Integr. Plant Biol. 2021, 63, 1999–2019. [Google Scholar] [CrossRef]
- Long, X.; Guan, C.; Wang, L.; Jia, L.; Fu, X.; Lin, Q.; Huang, Z.; Liu, C. Rice Storage Proteins: Focus on Composition, Distribution, Genetic Improvement and Effects on Rice Quality. Rice Sci. 2023, 30, 207–221. [Google Scholar]
- Chen, P.; Shen, Z.; Ming, L.; Li, Y.; Dan, W.; Lou, G.; Peng, B.; Wu, B.; Li, Y.; Zhao, D. Genetic basis of variation in rice seed storage protein (Albumin, Globulin, Prolamin, and Glutelin) content revealed by genome-wide association analysis. Front. Plant Sci. 2018, 9, 612. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, Y.; Liu, F.; Zhou, K.; Ding, Y.; Zhou, F.; Wang, Y.; Liu, K.; Gan, L.; Ma, W. GLUTELIN PRECURSOR ACCUMULATION3 encodes a regulator of post-Golgi vesicular traffic essential for vacuolar protein sorting in rice endosperm. Plant Cell 2014, 26, 410–425. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, S.; Liu, S.; Jiang, L.; Chen, L.; Ren, Y.; Han, X.; Liu, F.; Ji, S.; Liu, X. The vacuolar processing enzyme OsVPE1 is required for efficient glutelin processing in rice. Plant J. 2009, 58, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Kawakatsu, T.; Yamamoto, M.P.; Hirose, S.; Yano, M.; Takaiwa, F. Characterization of a new rice glutelin gene GluD-1 expressed in the starchy endosperm. J. Exp. Bot. 2008, 59, 4233–4245. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R. Improving the protein content and composition of cereal grain. J. Cereal Sci. 2007, 46, 239–250. [Google Scholar] [CrossRef]
- Zhang, W.; Bi, J.; Chen, L.; Zheng, L.; Ji, S.; Xia, Y.; Xie, K.; Zhao, Z.; Wang, Y.; Liu, L. QTL mapping for crude protein and protein fraction contents in rice (Oryza sativa L.). J. Cereal Sci. 2008, 48, 539–547. [Google Scholar] [CrossRef]
- Bhullar, N.K.; Gruissem, W. Nutritional enhancement of rice for human health: The contribution of biotechnology. Biotechnol. Adv. 2013, 31, 50–57. [Google Scholar] [CrossRef]
- Adachi, T.; Izumi, H.; Yamada, T.; Tanaka, K.; Takeuchi, S.; Nakamura, R.; Matsuda, T. Gene structure and expression of rice seed allergenic proteins belonging to the α-amylase/trypsin inhibitor family. Plant Mol. Biol. 1993, 21, 239–248. [Google Scholar] [CrossRef]
- Swamy, B.; Rahman, M.A.; Inabangan-Asilo, M.A.; Amparado, A.; Manito, C.; Chadha-Mohanty, P.; Reinke, R.; Slamet-Loedin, I.H. Advances in breeding for high grain zinc in rice. Rice 2016, 9, 49. [Google Scholar] [CrossRef]
- Li, H.; Yang, J.; Yan, S.; Lei, N.; Wang, J.; Sun, B. Molecular causes for the increased stickiness of cooked non-glutinous rice by enzymatic hydrolysis of the grain surface protein. Carbohydr. Polym. 2019, 216, 197–203. [Google Scholar] [CrossRef]
- Zhang, H.; Jang, S.-G.; Lar, S.M.; Lee, A.-R.; Cao, F.-Y.; Seo, J.; Kwon, S.-W. Genome-wide identification and genetic variations of the starch synthase gene family in rice. Plants 2021, 10, 1154. [Google Scholar] [CrossRef]
- Xiong, Q.; Sun, C.; Wang, R.; Wang, R.; Wang, X.; Zhang, Y.; Zhu, J. The Key Metabolites in Rice Quality Formation of Conventional japonica Varieties. Curr. Issues Mol. Biol. 2023, 45, 990–1001. [Google Scholar] [CrossRef]
- Chattopadhyay, K.; Behera, L.; Bagchi, T.B.; Sardar, S.S.; Moharana, N.; Patra, N.R.; Chakraborti, M.; Das, A.; Marndi, B.C.; Sarkar, A. Detection of stable QTLs for grain protein content in rice (Oryza sativa L.) employing high throughput phenotyping and genotyping platforms. Sci. Rep. 2019, 9, 3196. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, M.; Li, R.; Shen, L.; Wang, W.; Liu, M.; Zhu, Q.; Hu, Z.; He, Q.; Xue, Y. Identification of quantitative trait loci responsible for rice grain protein content using chromosome segment substitution lines and fine mapping of qPC-1 in rice (Oryza sativa L.). Mol. Breed. 2015, 35, 130. [Google Scholar] [CrossRef]
- Tan, Y.; Sun, M.; Xing, Y.; Hua, J.; Sun, X.; Zhang, Q.; Corke, H. Mapping quantitative trait loci for milling quality, protein content and color characteristics of rice using a recombinant inbred line population derived from an elite rice hybrid. Theor. Appl. Genet. 2001, 103, 1037–1045. [Google Scholar] [CrossRef]
- Wang, L.; Zhong, M.; Li, X.; Yuan, D.; Xu, Y.; Liu, H.; He, Y.; Luo, L.; Zhang, Q. The QTL controlling amino acid content in grains of rice (Oryza sativa) are co-localized with the regions involved in the amino acid metabolism pathway. Mol. Breed. 2008, 21, 127–137. [Google Scholar] [CrossRef]
- Lou, J.; Chen, L.; Yue, G.; Lou, Q.; Mei, H.; Xiong, L.; Luo, L. QTL mapping of grain quality traits in rice. J. Cereal Sci. 2009, 50, 145–151. [Google Scholar] [CrossRef]
- Ye, G.; Liang, S.; Wan, J. QTL mapping of protein content in rice using single chromosome segment substitution lines. Theor. Appl. Genet. 2010, 121, 741–750. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, W.; Chen, X.; Ma, J.; Chen, W.; Zhao, Z.; Zhai, H.; Wan, J. Dynamic QTL analysis of rice protein content and protein index using recombinant inbred lines. J. Plant Biol. 2011, 54, 321–328. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, W.; Liu, S.; Chen, L.; Liu, X.; Chen, X.; Ma, J.; Chen, W.; Zhao, Z.; Jiang, L.; et al. Genetic relationship between grain chalkiness, protein content, and paste viscosity properties in a backcross inbred population of rice. J. Cereal Sci. 2012, 56, 153–160. [Google Scholar] [CrossRef]
- Cheng, L.; Xu, Q.; Zheng, T.; Ye, G.; Luo, C.; Xu, J.; Li, Z. Identification of stably expressed quantitative trait loci for grain yield and protein content using recombinant inbred line and reciprocal introgression line populations in rice. Crop Sci. 2013, 53, 1437–1446. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Munakata, J. Identification and characteristics of quantitative trait locus for grain protein content, TGP12, in rice (Oryza sativa L.). Euphytica 2018, 214, 165. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, M.; Sun, S.; Zou, Y.; Yin, S.; Liu, Y.; Tang, S.; Gu, M.; Yang, Z.; Yan, C. Natural variation of OsGluA2 is involved in grain protein content regulation in rice. Nat. Commun. 2019, 10, 1949. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Griffin, V.K. Effect of disulfide bond-containing protein on rice starch gelatinization and pasting. Cereal Chem. 1993, 70, 377–380. [Google Scholar]
- Martin, M.; Fitzgerald, M. Proteins in rice grains influence cooking properties! J. Cereal Sci. 2002, 36, 285–294. [Google Scholar] [CrossRef]
- Kumamaru, T.; Satoh, H.; Iwata, N.; Omura, T.; Ogawa, M.; Tanaka, K.J.T.; Genetics, A. Mutants for rice storage proteins: 1. Screening of mutants for rice storage proteins of protein bodies in the starchy endosperm. Theoret. Appl. Genet. 1988, 76, 11–16. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Brubaker, C.L.; Wendel, J.F. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Rep. 1993, 11, 122–127. [Google Scholar] [CrossRef]
- Wang, S. Windows QTL Cartographer 2.5. 2006. Available online: http://statgen.ncsu.edu/qtlcart/WQTLCart.Htm (accessed on 1 November 2018).
- Lincoln, S. Mapping genes controlling quantitative traits with MAPMAKER/QTL 1.1. In Whitehead Institute for Biomedical Research Technical Report; Whitehead Institute: Cambridge, MA, USA, 1992. [Google Scholar]
- Kosambi, D.D. The estimation of map distances from recombination values. In D.D. Kosambi: Selected Works in Mathematics and Statistics; Springer: New Delhi, India, 2016; pp. 125–130. [Google Scholar]
- Kepiro, J.; McClung, A.; Chen, M.-H.; Yeater, K.; Fjellstrom, R. Mapping QTLs for milling yield and grain characteristics in a tropical japonica long grain cross. J. Cereal Sci. 2008, 48, 477–485. [Google Scholar] [CrossRef]
- Liu, X.; Wan, X.; Ma, X.; Wan, J. Dissecting the genetic basis for the effect of rice chalkiness, amylose content, protein content, and rapid viscosity analyzer profile characteristics on the eating quality of cooked rice using the chromosome segment substitution line population across eight environments. Genome 2011, 54, 64–80. [Google Scholar]
- Zhong, M.; Wang, L.-Q.; Yuan, D.-J.; Luo, L.-J.; Xu, C.-G.; He, Y.-Q. Identification of QTL affecting protein and amino acid contents in rice. Rice Sci. 2011, 18, 187–195. [Google Scholar] [CrossRef]
- Kinoshita, N.; Kato, M.; Koyasaki, K.; Kawashima, T.; Nishimura, T.; Hirayama, Y.; Takamure, I.; Sato, T.; Kato, K. Identification of quantitative trait loci for rice grain quality and yield-related traits in two closely related Oryza sativa L. subsp. japonica cultivars grown near the northernmost limit for rice paddy cultivation. Breed. Sci. 2017, 67, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhao, C.-F.; Zhou, L.-H.; Yao, S.; Zhao, Q.-Y.; Chen, T.; Zhu, Z.; Zhang, Y.-D.; Wang, C.-L. Mapping QTLs for rice (Oryza sativa L.) grain protein content via chromosome segment substitution lines. Cereal Res. Commun. 2022, 50, 699–708. [Google Scholar] [CrossRef]
- Jang, S.; Han, J.-H.; Lee, Y.K.; Shin, N.-H.; Kang, Y.J.; Kim, C.-K.; Chin, J.H. Mapping and validation of QTLs for the amino acid and total protein content in brown rice. Front. Genet. 2020, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-H.; Li, G.; Fan, Y.-Y.; Zhang, K.-Q.; Min, J.; Zhu, Z.-W.; Zhuang, J.-Y. Genetic relationship between grain yield and the contents of protein and fat in a recombinant inbred population of rice. J. Cereal Sci. 2009, 50, 121–125. [Google Scholar] [CrossRef]
- Hu, Z.-L.; Li, P.; Zhou, M.-Q.; Zhang, Z.-H.; Wang, L.-X.; Zhu, L.-H.; Zhu, Y.-G. Mapping of quantitative trait loci (QTLs) for rice protein and fat content using doubled haploid lines. Euphytica 2004, 135, 47–54. [Google Scholar] [CrossRef]
- Wang, X.; Pang, Y.; Zhang, J.; Wu, Z.; Chen, K.; Ali, J.; Ye, G.; Xu, J.; Li, Z. Genome-wide and gene-based association mapping for rice eating and cooking characteristics and protein content. Sci. Rep. 2017, 7, 17203. [Google Scholar] [CrossRef]
- Bruno, E.; Choi, Y.-S.; Chung, I.K.; Kim, K.M. QTLs and analysis of the candidate gene for amylose, protein, and moisture content in rice (Oryza sativa L.). 3 Biotech 2017, 7, 40. [Google Scholar] [CrossRef]
- Aluko, G.; Martinez, C.; Tohme, J.; Castano, C.; Bergman, C.; Oard, J. QTL mapping of grain quality traits from the interspecific cross Oryza sativa× O. glaberrima. Theor. Appl. Genet. 2004, 109, 630–639. [Google Scholar] [CrossRef]
- Nan, J.; Feng, X.; Wang, C.; Zhang, X.; Wang, R.; Liu, J.; Yuan, Q.; Jiang, G.; Lin, S. Improving rice grain length through updating the GS3 locus of an elite variety Kongyu 131. Rice 2018, 11, 21. [Google Scholar] [CrossRef]
- Harushima, Y.; Nakagahra, M.; Yano, M.; Sasaki, T.; Kurata, N. A genome-wide survey of reproductive barriers in an intraspecific hybrid. Genetics 2001, 159, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Chin, J.H.; Chu, S.-H.; Jiang, W.; Cho, Y.-I.; Basyirin, R.; Brar, D.S.; Koh, H.-J. Identification of QTLs for hybrid fertility in inter-subspecific crosses of rice (Oryza sativa L.). Genes Genom. 2011, 33, 39–48. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Baksh, S.Y.; Mukherjee, A.K.; Mohanty, S.P. Development of flash-flood tolerant and durable bacterial blight resistant versions of mega rice variety ‘Swarna’through marker-assisted backcross breeding. Sci. Rep. 2019, 9, 12810. [Google Scholar] [CrossRef] [PubMed]
- Hillerislambers, D.; Rutger, J.; Qualset, C.; Wiser, W. Genetic and environmental variation in protein content of rice (Oryza sativa L.). Euphytica 1973, 22, 264–273. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Hirose, S.; Yasuda, H.; Takaiwa, F. Reducing rice seed storage protein accumulation leads to changes in nutrient quality and storage organelle formation. Plant Physiol. 2010, 154, 1842–1854. [Google Scholar] [CrossRef]
- Ina, S.; Ninomiya, K.; Mogi, T.; Hase, A.; Ando, T.; Matsukaze, N.; Ogihara, J.; Akao, M.; Kumagai, H.; Kumagai, H. Rice (Oryza sativa japonica) albumin suppresses the elevation of blood glucose and plasma insulin levels after oral glucose loading. J. Agric. Food Chem. 2016, 64, 4882–4890. [Google Scholar] [CrossRef]
- Yamagata, H.; Sugimoto, T.; Tanaka, K.; Kasai, Z. Biosynthesis of storage proteins in developing rice seeds. Plant Physiol. 1982, 70, 1094–1100. [Google Scholar] [CrossRef]
- Park, S.-G.; Park, H.-S.; Baek, M.-K.; Jeong, J.-M.; Cho, Y.-C.; Lee, G.-M.; Lee, C.-M.; Suh, J.-P.; Kim, C.-S.; Kim, S.-M. Improving the glossiness of cooked rice, an important component of visual rice grain quality. Rice 2019, 12, 87. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Takaiwa, F. Cereal seed storage protein synthesis: Fundamental processes for recombinant protein production in cereal grains. Plant Biotechnol. J. 2010, 8, 939–953. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Traits | BC3F3 Introgression Line Populations | |||
|---|---|---|---|---|---|
| Mean | SD | Minimum | Maximum | ||
| Population I (Kongyu 131/Cypress) | Alb (mg/g) | 6.31 C | 1.53 | 2.02 | 10.31 |
| Gol (mg/g) | 7.91 C | 2.14 | 1.46 | 13.43 | |
| Pro (mg/g) | 1.68 D | 0.46 | 0.63 | 3.17 | |
| Glu (mg/g) | 42.08 B | 8.85 | 25.64 | 71.33 | |
| Gpc (mg/g) | 113.3 A | 3.59 | 103.7 | 125.6 | |
| Population-II (Kongyu 131/Vary Tarva Osla) | Alb (mg/g) | 5.33 C | 1.79 | 1.21 | 9.46 |
| Gol (mg/g) | 4.86 C | 1.52 | 1.47 | 9.79 | |
| Pro (mg/g) | 2.52 D | 0.51 | 1.23 | 4.68 | |
| Glu (mg/g) | 65.79 B | 14.32 | 30.26 | 99.47 | |
| Gpc (mg/g) | 114.5 A | 3.52 | 104.4 | 122.4 | |
| Trait | Pop | Chr. | QTL | Marker | Physical Interval (Mb) | LOD | Additive Effect | R2 (%) | Co-Localization/ Previous/Chr. Interval/QTLs/Genes | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Alb | P-I | 1 | qAlb1.2 | RM140-C1.23.3 | 12.4–23.3 | 3.92 | 4.55 | 48.53 | qPR1, qALB-1, qPC-1a, qPC-1b, qALB-1, Cpb1, Cph1 | [17,43,44,45] |
| 3 | qAlb3.1 | T3-4-C3.26.6 | 19.1–26.67 | 2.58 | −2.77 | 42.58 | qPC3, qPC3-1 | [46,47] | ||
| 7 | qAlb7.3 | RM501-T7-3 | 8.0–18.1 | 3.5 | 4.5 | 22.09 | qPC7, qPC7-1, qPC7-2, qPC-7, qAAC7.3 | ([12,28,45,47,48] | ||
| 8 | qAlb8.2 | Z8.25.3-RM264 | 25.3–28.0 | 2.96 | 4.46 | 1.6 | qPC-8b, qAAC8.2 | [12,48,49] | ||
| 9 | qAlb9.1 | RM160-Z9-22.4 | 20.4–22.3 | 2.62 | −2.63 | 40.16 | ||||
| P-II | 4 | qAlb4.2 | RM470-RM567 | 28.6–35.1 | 4.49 | −2.52 | 7.23 | qPC4, qRPC-4 | [47,50] | |
| Gol | P-I | 2 | qGol2.4 | T2-1-T2-2 | 4.5–6.9 | 5.15 | 9.4 | 2.67 | qPC-2,qGLB2.1, qSGpc2.1, Cph2 | [17,24,43,46] |
| 4 | qGol4.1 | Z4-17.7-T4-6 | 17.7–24.6 | 6.01 | 9.4 | 33.57 | ||||
| 4 | qGol4.2 | W4.30.6-Z435.1 | 30.66–35.1 | 5.96 | 5.49 | 13.68 | qPC4, qRPC-4 | [47,50] | ||
| 8 | qGol8.1 | RM404-RM515 | 15.5–20.3 | 5.33 | 9.42 | 33.26 | qPC8, qPC-8b | [44,47] | ||
| 8 | qGol8.2 | RM556--Z8-25.3 | 22.4–25.3 | 6.73 | 9.37 | 46.73 | qPC-8b, qAAC8.1, qAAC8.2 | [44,48] | ||
| 10 | qGol10.2 | A10.2.84-Z10-16.6 | 2.8–16.6 | 3.88 | −3.94 | 15.5 | qPC10, qPC10-2, qPC10-3, qPC10.1 | [47,49,51] | ||
| 11 | qGol11.2 | W11-6.6-RM536 | 6.6–9.0 | 6.79 | 9.37 | 55.78 | ||||
| Pro | P-I | 3 | qPro3.1 | T3-1-RM218 | 0.7–8.4 | 2.82 | −1.79 | 6.9 | qPC3, qPLA3, qPC-3.1, qPC-3.2, qPC-3.3, qAAC3.1, qAAC3.2 | [12,17,30,48,49] |
| 4 | qPro4.1 | RM518-Z4-4.0 | 2.0–4.0 | 2.98 | −2.11 | 34.28 | qPC-4, Cpb4, Cph4 | [30,43] | ||
| 5 | qPro5.2 | RM509-RM164 | 16.3–19.2 | 4.74 | −1.69 | 51.7 | ||||
| 5 | qPro5.3 | RM-164-RM480 | 19.2–27.5 | 7.53 | −1.95 | 47.17 | ||||
| 10 | qPro10.1 | W10-0.1-A10.2.84 | 0.1–2.8 | 6.83 | −2.09 | 61.85 | qPC10-1, qPC-10 | [30,47] | ||
| 12 | qPro12.1 | Z12-0.94-Z12-9.4 | 0.9–9.4 | 5.89 | −2.13 | 45.53 | qSGpc12.1, qPC-12 | [24,30] | ||
| Glu | P-I | 4 | qGlu4.1 | Z4-17.7-Z4-26.5 | 17.7–26.5 | 2.5 | −13.32 | 7.52 | ||
| Gpc | P-I | 7 | qGpc7.3 | RM481-RM501 | 2.9–8.0 | 3.71 | −1.67 | 3.14 | qPC7.1, qPC7,qPC-7, qRPC-7, qPC7, qAAC7.1, qAAC7.2, qAAC7.1 | [26,28,48,50,52] |
| P-II | 1 | qGpc1.2 | RM292-T1-3 | 9.5–12.5 | 3.54 | 2.5 | 4.1 | qSGpc1.3, qPC-1a, qPC-1b, qALB-1, Cpb1, Cph1 | [17,24,43,44] | |
| 6 | qGpc6.2 | T6-7-T6-4 | 8.5–18 | 5.26 | 2.18 | 6.27 | qPC-6, qPC-6, qPC-6 | [28,28,44] | ||
| 7 | qGpc7.4 | T7-1-T7-3 | 24.8–27.8 | 18.4 | 4.82 | 23.63 | qPC1 | [26] | ||
| 10 | qGpc10.3 | RM239-D10-7F | 9.8–11.8 | 3.31 | −4 | 4.47 | qPC10, qPC10.1 | [12,49,51] |
| QTLs | N | GN | (Mean ± SD) | p-Value | % |
|---|---|---|---|---|---|
| qGpc7.3 | 37 | KY131 | 101.54 ± 0.74 | 4.63544 × 10−11 | 1.69 |
| 19 | Cypress | 103.26 ± 0.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, M.; Tan, X.; Zhang, H.; Lou, G.; Yang, H.; Zhou, Y.; Hussain, A.; Bhantana, P.; Jiang, G.; He, Y. QTL Mining and Validation of Grain Nutritional Quality Characters in Rice (Oryza sativa L.) Using Two Introgression Line Populations. Agriculture 2023, 13, 1725. https://doi.org/10.3390/agriculture13091725
Alam M, Tan X, Zhang H, Lou G, Yang H, Zhou Y, Hussain A, Bhantana P, Jiang G, He Y. QTL Mining and Validation of Grain Nutritional Quality Characters in Rice (Oryza sativa L.) Using Two Introgression Line Populations. Agriculture. 2023; 13(9):1725. https://doi.org/10.3390/agriculture13091725
Chicago/Turabian StyleAlam, Mufid, Xuan Tan, Hao Zhang, Guangming Lou, Hanyuan Yang, Yin Zhou, Amjad Hussain, Parashuram Bhantana, Gonghao Jiang, and Yuqing He. 2023. "QTL Mining and Validation of Grain Nutritional Quality Characters in Rice (Oryza sativa L.) Using Two Introgression Line Populations" Agriculture 13, no. 9: 1725. https://doi.org/10.3390/agriculture13091725
APA StyleAlam, M., Tan, X., Zhang, H., Lou, G., Yang, H., Zhou, Y., Hussain, A., Bhantana, P., Jiang, G., & He, Y. (2023). QTL Mining and Validation of Grain Nutritional Quality Characters in Rice (Oryza sativa L.) Using Two Introgression Line Populations. Agriculture, 13(9), 1725. https://doi.org/10.3390/agriculture13091725