
DIO1 Gene Polymorphism Is Associated with Thyroid Profiles and Reproductive Performance in Dairy Cows

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Blood Sampling and Reproductive Traits Assessment
2.3. Hormone Determination
2.4. Genomic DNA Extraction
2.5. Primer Design
- 13,141 gagtgaaac/gg gcggctctag caatgtgtga gatccatcct aaggctctcc
- 16,741 gctgcttaac agcttacaac acacaaa/caca gcccccacaa caagaattac/t
- 16,921 catcaagaat caccggaatc tccaggaccg cctgcgggca gcccacctgc/a.
2.6. Real-Time PCR Analysis
2.7. Statistical Analysis
3. Results
3.1. Polymorphic Variants of the DIO1 Gene
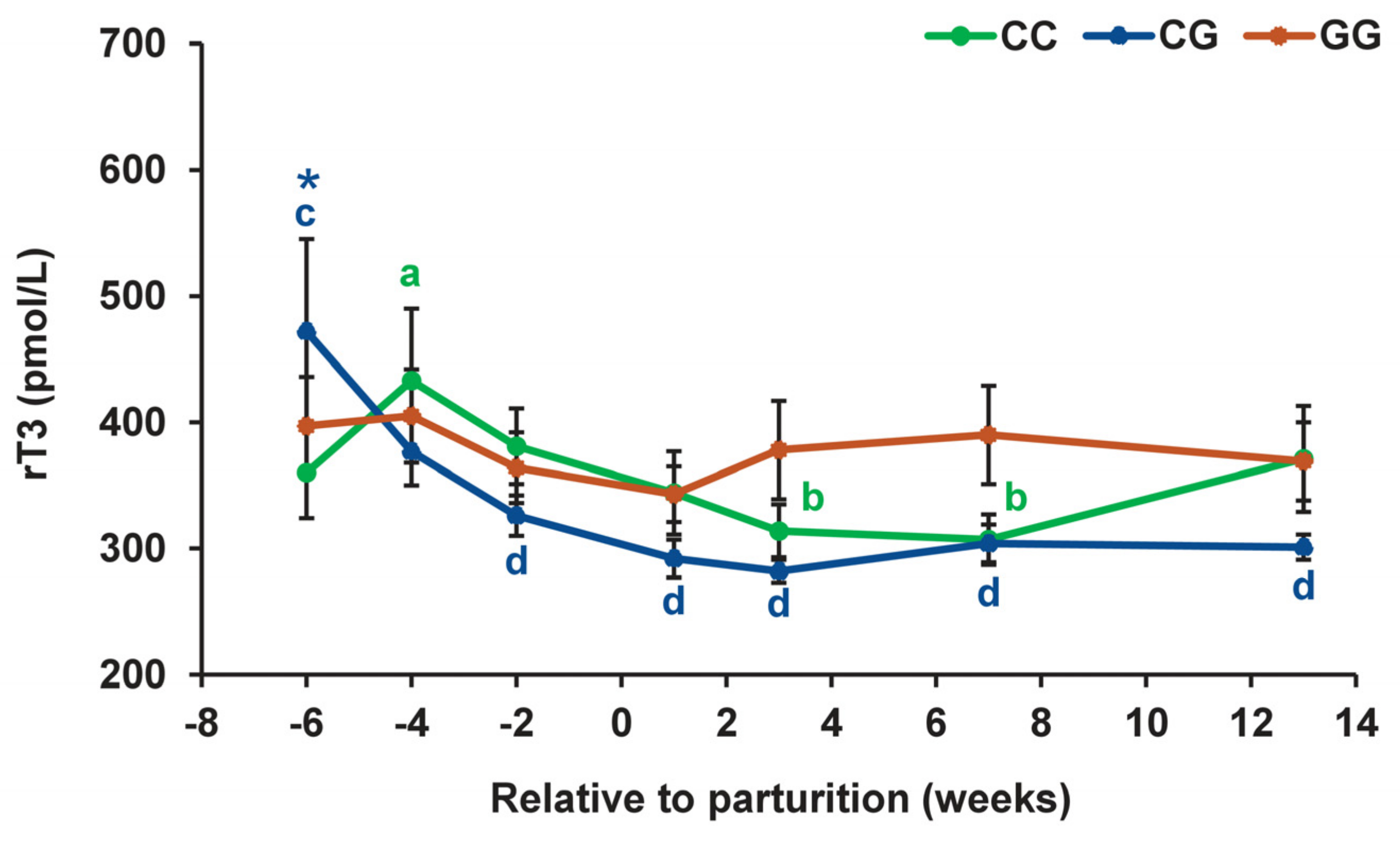
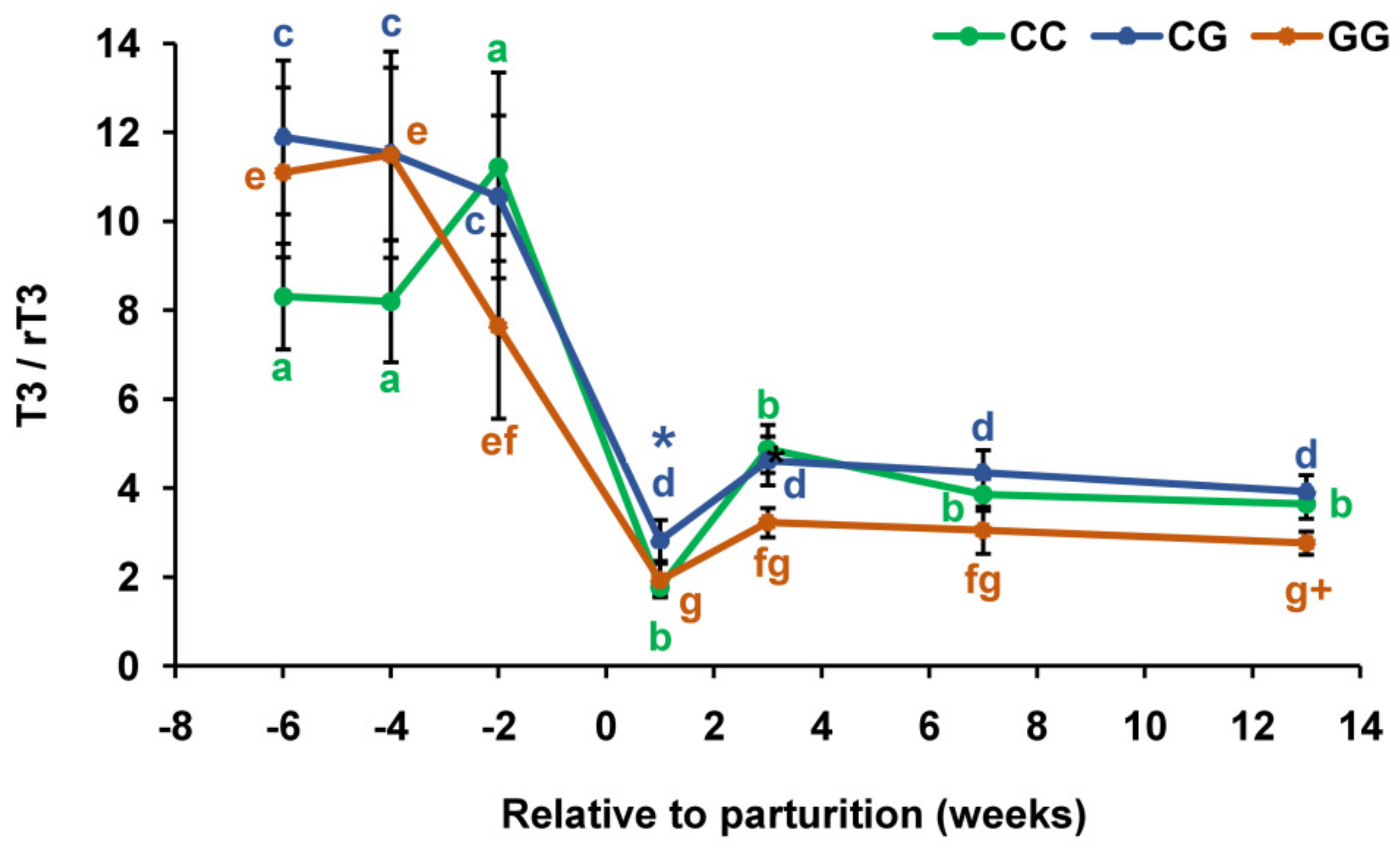
3.2. Thyroid Profiles in Cows with Polymorphic Variants of the DIO1 Gene
3.3. Reproductive Traits, Their Association with Genotypes and Thyroid Profiles, and BCS in Cows with Polymorphic Variants of the DIO1 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Roche, J.R.; Burke, C.R.; Crookenden, M.A.; Heiser, A.; Loor, J.; Meier, S.; Mitchell, M.; Phyn, C.V.C.; Turner, S.-A. Fertility and the transition dairy cow. Reprod. Fertil. Dev. 2018, 30, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Clempson, A.M.; Pollott, G.E. Associations between lipid metabolism and fertility in the dairy cow. Reprod. Fertil. Dev. 2012, 25, 48–61. [Google Scholar] [CrossRef] [PubMed]
- González, F.D.; Muiño, R.; Pereira, V.; Campos, R.; Benedito, J.L. Relationship among blood indicators of lipomobilization and hepatic function during early lactation in high-yielding dairy cows. J. Veter-Sci. 2011, 12, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.-K.; Jeong, J.-K.; Choi, I.-S.; Kang, H.-G.; Hur, T.-Y.; Jung, Y.-H.; Kim, I.-H. Relationships among ketosis, serum metabolites, body condition, and reproductive outcomes in dairy cows. Theriogenology 2015, 84, 252–260. [Google Scholar] [CrossRef]
- Wathes, D.C. Mechanisms Linking Metabolic Status and Disease with Reproductive Outcome in the Dairy Cow. Reprod. Domest. Anim. 2012, 47, 304–312. [Google Scholar] [CrossRef]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid Hormone Regulation of Metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef]
- Capuco, A.; Wood, D.; Elsasser, T.; Kahl, S.; Erdman, R.; Van Tassell, C.; Lefcourt, A.; Piperova, L. Effect of Somatotropin on Thyroid Hormones and Cytokines in Lactating Dairy Cows During Ad Libitum and Restricted Feed Intake. J. Dairy Sci. 2001, 84, 2430–2439. [Google Scholar] [CrossRef]
- Mohebbi-Fani, M.; Nazifi, S.; Rowghani, E.; Bahrami, S.; Jamshidi, O. Thyroid hormones and their correlations with serum glucose, beta hydroxybutyrate, nonesterified fatty acids, cholesterol, and lipoproteins of high-yielding dairy cows at different stages of lactation cycle. Comp. Clin. Path. 2009, 18, 211–216. [Google Scholar] [CrossRef]
- Fiore, E.; Piccione, G.; Gianesella, M.; Praticò, V.; Vazzana, I.; Dara, S.; Morgante, M. Serum thyroid hormone evaluation during transition periods in dairy cows. Arch. Anim. Breed. 2015, 58, 403–406. [Google Scholar] [CrossRef]
- Mityashova, O.S.; Solomakhin, A.A.; Bogolyubova, N.V.; Montvila, E.K.; Lebedeva, I.Y. Lipid metabolism and thyroid status of heifers with different functional conditions of the ovaries. Dostizheniya Nauk. I Tekhniki APK 2020, 34, 69–74. (In Russian) [Google Scholar] [CrossRef]
- Vernon, R.G.; Denis, R.G.P.; Sorensen, A.; Williams, G. Leptin and the Adaptations of Lactation in Rodents and Ruminants. Horm. Metab. Res. 2002, 34, 678–685. [Google Scholar] [CrossRef]
- Šamanc, H.; Stojić, V.; Kirovski, D.; Jovanović, M.; Cernescu, H.; Vujanac, I. Thyroid Hormones Concentrations during the Mid-Dry Period: An Early Indicator of Fatty Liver in Holstein-Friesian Dairy Cows. J. Thyroid. Res. 2010, 2010, 897602. [Google Scholar] [CrossRef][Green Version]
- Piechotta, M.; Holzhausen, L.; Araujo, G.M.; Heppelmann, M.; Sipka, A.; Pfarrer, C.; Schuberth, H.-J.; Bollwein, H. Antepartal insulin-like growth factor concentrations indicating differences in the metabolic adaptive capacity of dairy cows. J. Veter-Sci. 2014, 15, 343–352. [Google Scholar] [CrossRef]
- Kafi, M.; Tamadon, A.; Saeb, M.; Mirzaei, A.; Ansari-Lari, M. Relationships between thyroid hormones and serum energy metabolites with different patterns of postpartum luteal activity in high-producing dairy cows. Animal 2012, 6, 1253–1260. [Google Scholar] [CrossRef]
- Lebedeva, I.Y.; Mityashova, O.S. Dependence of reproductive ability of first-calf heifers on the dynamics of the content of thyroid hormones in the blood in the first trimester of lactation. Dostizheniya Nauk. I Tekhniki APK 2020, 34, 91–96. (In Russian) [Google Scholar] [CrossRef]
- Ashkar, F.A.; Bartlewski, P.M.; Singh, J.; Malhi, P.S.; Yates, K.M.; Singh, T.; King, W.A. Thyroid hormone concentrations in systemic circulation and ovarian follicular fluid of cows. Exp. Biol. Med. 2010, 235, 215–221. [Google Scholar] [CrossRef]
- Costa, N.; Cordeiro, M.; Silva, T.; Sastre, D.; Santana, P.; Sá, A.; Sampaio, R.; Santos, S.; Adona, P.; Miranda, M.; et al. Effect of triiodothyronine on developmental competence of bovine oocytes. Theriogenology 2013, 80, 295–301. [Google Scholar] [CrossRef]
- Spicer, L.; Alonso, J.; Chamberlain, C. Effects of Thyroid Hormones on Bovine Granulosa and Thecal Cell Function In Vitro: Dependence on Insulin and Gonadotropins. J. Dairy Sci. 2001, 84, 1069–1076. [Google Scholar] [CrossRef]
- Ortiga-Carvalho, T.M.; Chiamolera, M.I.; Pazos-Moura, C.C.; Wondisford, F.E. Hypothalamus-Pituitary-Thyroid Axis. Compr. Physiol. 2016, 6, 1387–1428. [Google Scholar] [CrossRef]
- Sabatino, L.; Vassalle, C.; Del Seppia, C.; Iervasi, G. Deiodinases and the Three Types of Thyroid Hormone Deiodination Reactions. Endocrinol. Metab. 2021, 36, 952–964. [Google Scholar] [CrossRef]
- Bos, M.M.; Smit, R.A.J.; Trompet, S.; Van Heemst, D.; Noordam, R. Thyroid Signaling, Insulin Resistance, and 2 Diabetes Mellitus: A Mendelian Randomization Study. J. Clin. Endocrinol. Metab. 2017, 102, 1960–1970. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.C.; Kim, B.S. Pathophysiological relevance of deiodinase polymorphism. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.E.; Sonstegard, T.S.; Kahl, S.; Bennett, G.L.; Snelling, W.M. The bovine type I iodothyronine deiodinase (DIO1) gene maps to chromosome 3. Anim. Genet. 2003, 34, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Li, D.; Wang, G.; Wu, X.; Zhang, M.; Zhang, C.; Cui, Y.; Li, S. Expression and imprinting of DIO3 and DIO3OS genes in Holstein cattle. J. Genet. 2017, 96, 333–339. [Google Scholar] [CrossRef]
- Coster, A.; Madsen, O.; Heuven, H.C.M.; Dibbits, B.; Groenen, M.A.M.; van Arendonk, J.A.M.; Bovenhuis, H. The Imprinted Gene DIO3 Is a Candidate Gene for Litter Size in Pigs. PLoS ONE 2012, 7, e31825. [Google Scholar] [CrossRef]
- Oczkowicz, M.; Dunkowska, A.; Piórkowska, K.; Mucha, A.; Tyra, M.; Ropka-Molik, K. 7. Associations between Polymorphisms in the DIO3 Gene and Reproductive Traits and Carcass Performance in Pigs. Ann. Anim. Sci. 2016, 16, 399–413. [Google Scholar] [CrossRef][Green Version]
- Wildman, E.E.; Jones, G.M.; Wagner, P.E.; Boman, R.L.; Troutt, H.F., Jr.; Lesch, T.N. A Dairy Cow Body Condition Scoring System and Its Relationship to Selected Production Characteristics. J. Dairy Sci. 1982, 65, 495–501. [Google Scholar] [CrossRef]
- Wiltbank, M.C.; Gümen, A.; Sartori, R. Physiological classification of anovulatory conditions in cattle. Theriogenology 2002, 57, 21–52. [Google Scholar] [CrossRef]
- Shrestha, H.K.; Nakao, T.; Suzuki, T.; Higaki, T.; Akita, M. Effects of abnormal ovarian cycles during pre-service period postpartum on subsequent reproductive performance of high-producing Holstein cows. Theriogenology 2004, 61, 1559–1571. [Google Scholar] [CrossRef]
- Motil, K.J.; Thotathuchery, M.; Montandon, C.M.; Hachey, D.L.; Boutton, T.W.; Klein, P.D.; Garza, C. Insulin, Cortisol and Thyroid Hormones Modulate Maternal Protein Status and Milk Production and Composition in Humans. J. Nutr. 1994, 124, 1248–1257. [Google Scholar] [CrossRef]
- Panicker, V.; Cluett, C.; Shields, B.; Murray, A.; Parnell, K.S.; Perry, J.R.B.; Weedon, M.N.; Singleton, A.; Hernandez, D.; Evans, J.; et al. A Common Variation in Deiodinase 1 Gene DIO1 Is Associated with the Relative Levels of Free Thyroxine and Triiodothyronine. J. Clin. Endocrinol. Metab. 2008, 93, 3075–3081. [Google Scholar] [CrossRef]
- Liang, S.-R.; Cai, J.; Yang, Y.; Zhang, L.; Taylor, P.; Ming, J.; Yu, X.-W.; Hu, R.-F.; Zhou, J.; Da-Yan, C.M.; et al. Relationship between thyroid hormones and metabolic syndrome in a normal thyroid function population in Western China: A cross-sectional study based on both epidemiological and genetic analysis. Chin. Med. J. 2021, 135, 350–352. [Google Scholar] [CrossRef]
- de Jong, F.J.; Peeters, R.P.; Heijer, T.D.; van der Deure, W.M.; Hofman, A.; Uitterlinden, A.G.; Visser, T.J.; Breteler, M.M.B. The Association of Polymorphisms in the Type 1 and 2 Deiodinase Genes with Circulating Thyroid Hormone Parameters and Atrophy of the Medial Temporal Lobe. J. Clin. Endocrinol. Metab. 2006, 92, 636–640. [Google Scholar] [CrossRef]
- Wolff, T.M.; Dietrich, J.W.; Müller, M.A. Optimal Hormone Replacement Therapy in Hypothyroidism—A Model Predictive Control Approach. Front. Endocrinol. 2022, 13, 884018. [Google Scholar] [CrossRef]
- Pascottini, O.B.; Leroy, J.L.M.R.; Opsomer, G. Maladaptation to the transition period and consequences on fertility of dairy cows. Reprod. Domest. Anim. 2022, 57 (Suppl. 4), 21–32. [Google Scholar] [CrossRef]
- De Koster, J.D.; Opsomer, G. Insulin Resistance in Dairy Cows. Veter-Clin. N. Am. Food Anim. Pract. 2013, 29, 299–322. [Google Scholar] [CrossRef]







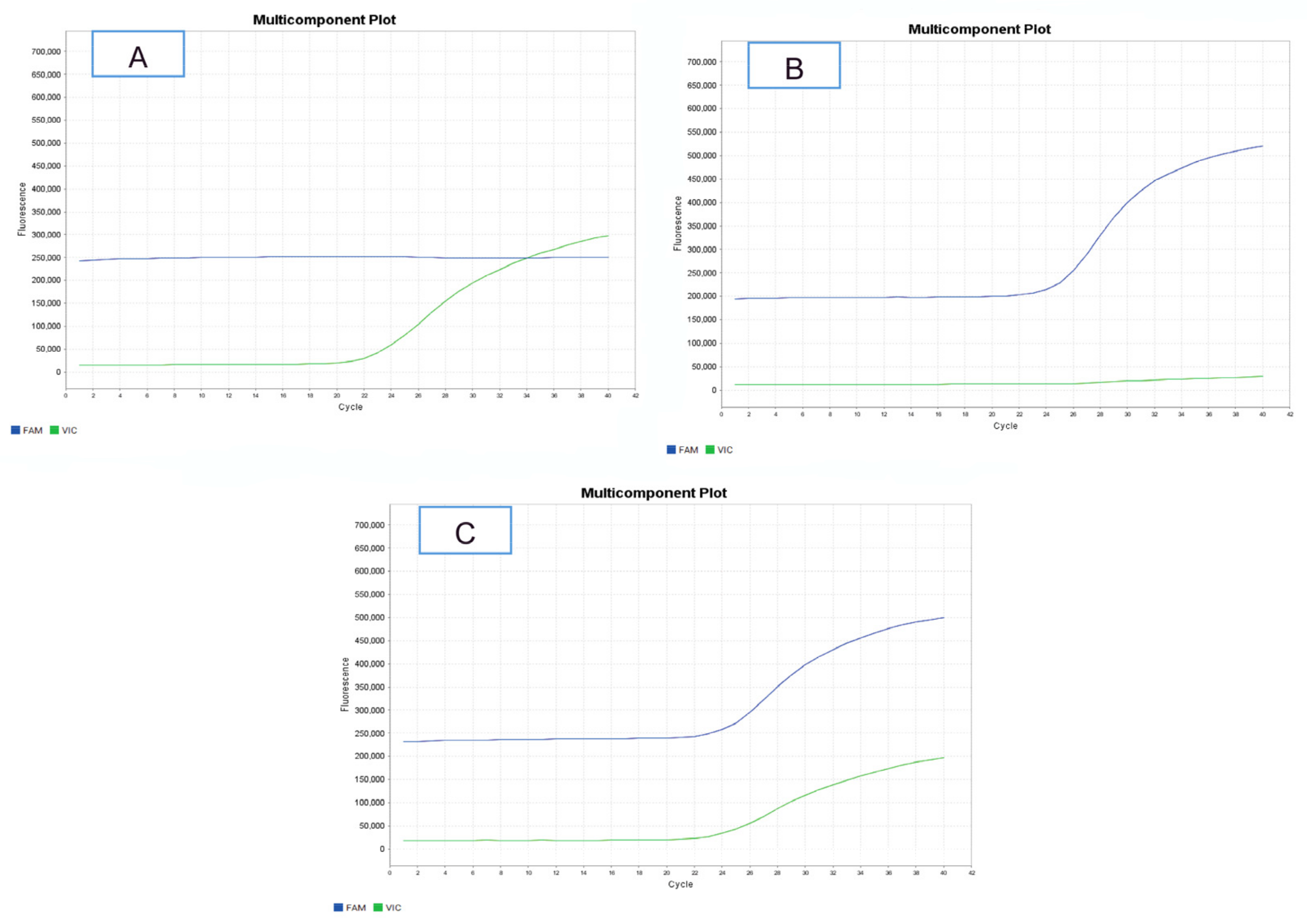{kind=link}
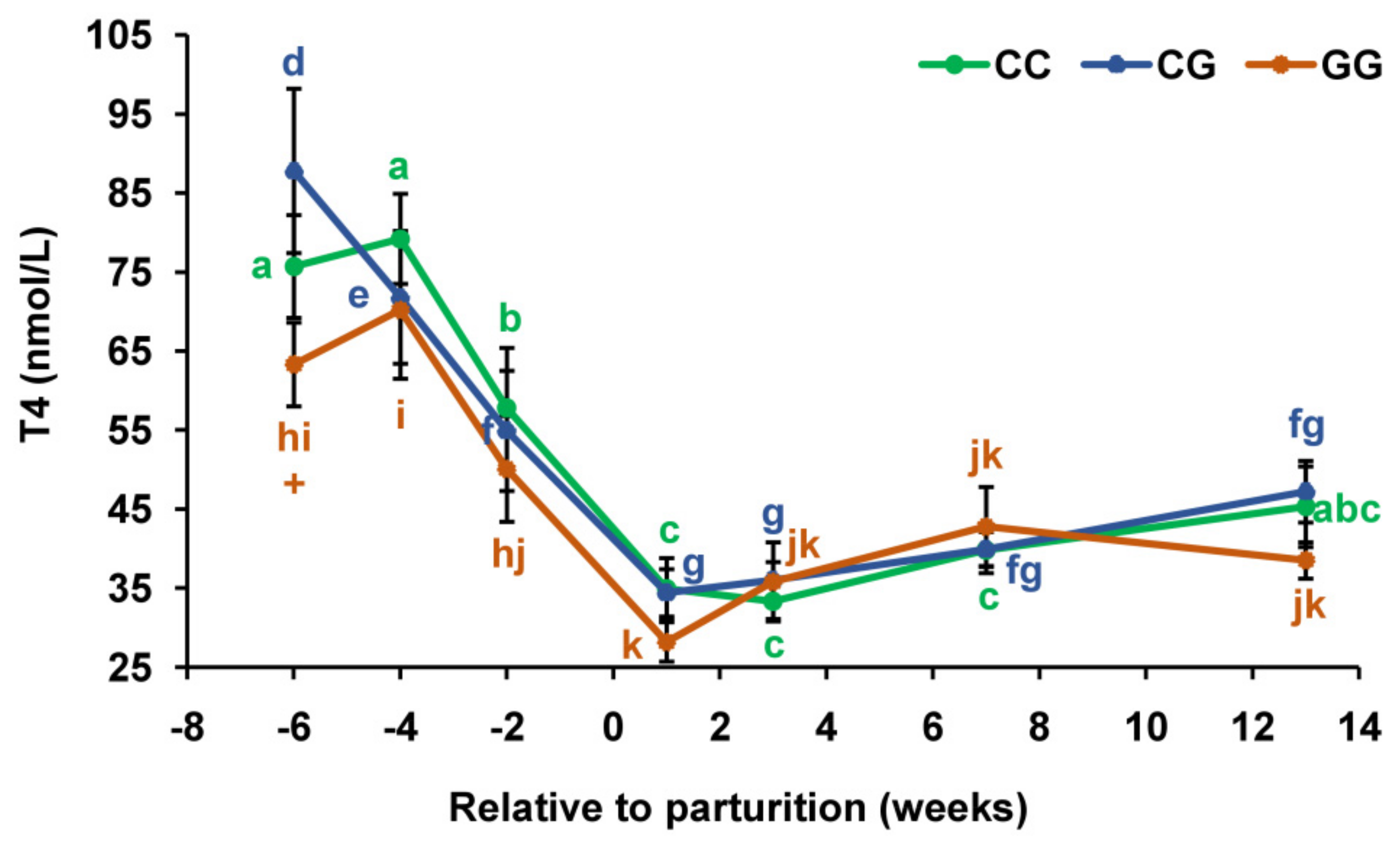{kind=link}
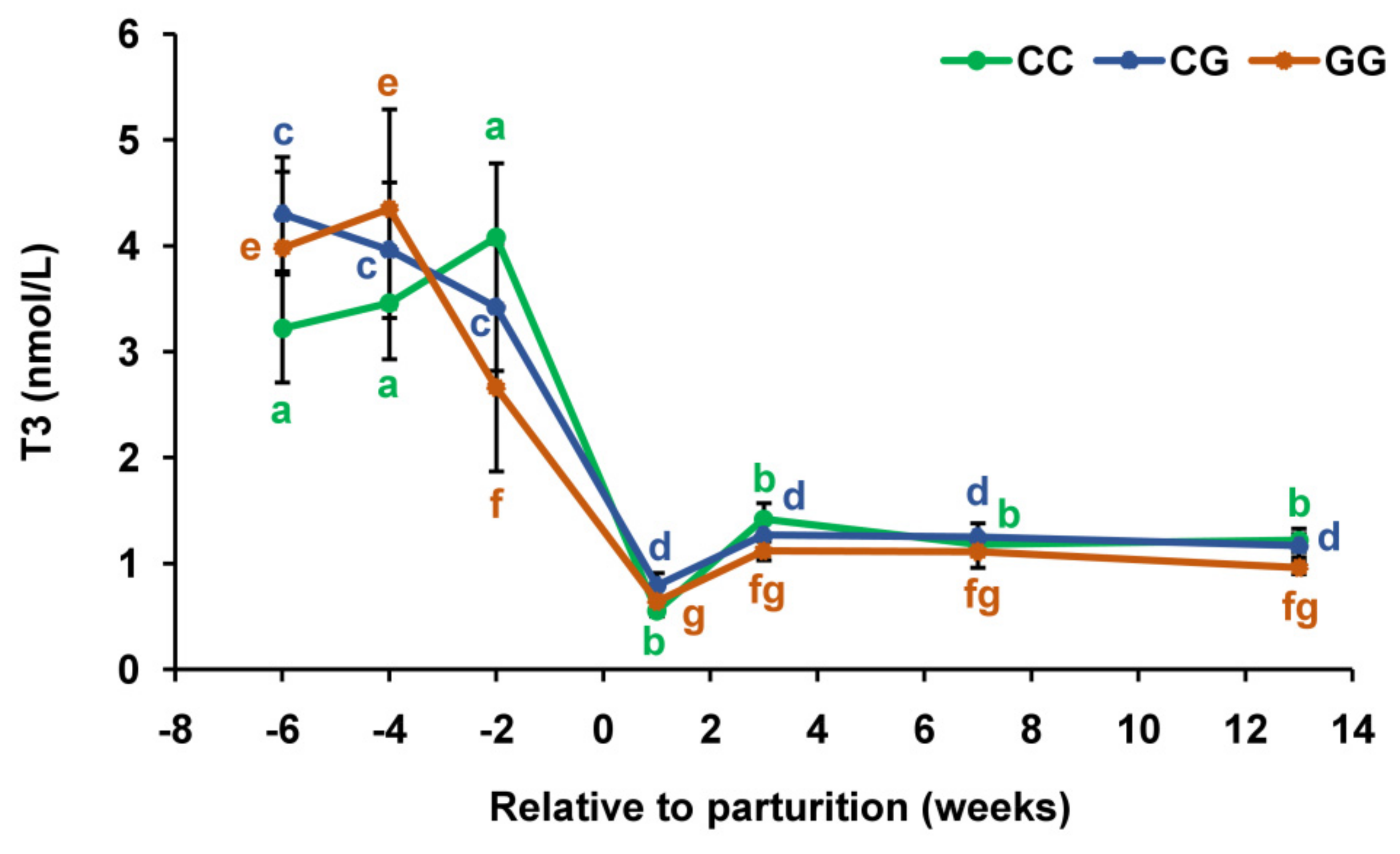{kind=link}
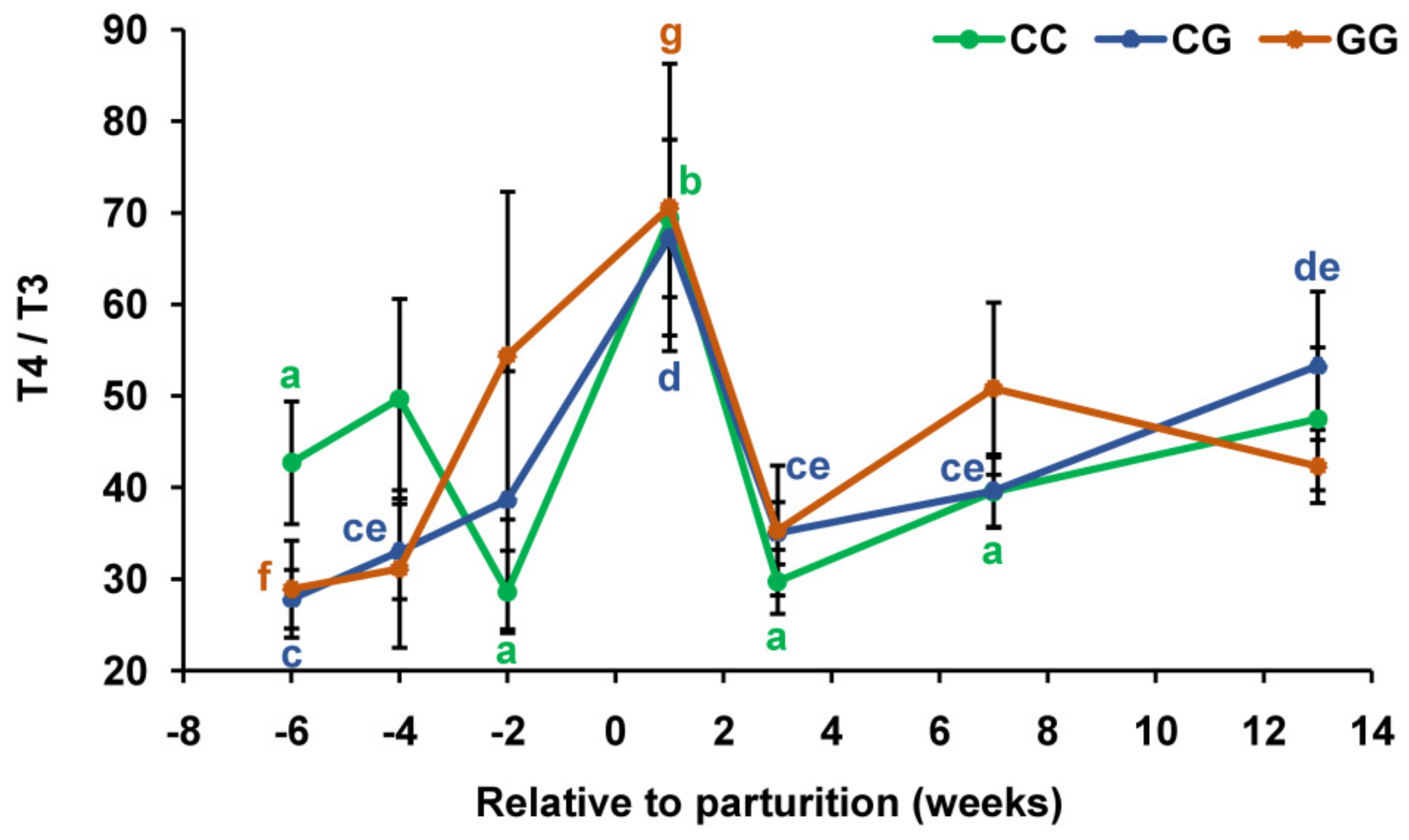{kind=link}
{kind=link}
{kind=link}
| SNP Position | Sequence of Primers and Probes | T (°C) |
|---|---|---|
| 16,767–16,790 | F: CTTTGGCACTACTTACAGTTTGG R: AAGACGCTGCTTAACAGCTTACA TAMRA-CACACAACACAGCCCCCA-BHQ-2 ROX-CACACAAAACAGCCCCCA-BHQ-2 FAM-CTAGATAATTCTTGTTGTGGG-BHQ-1 R6G-CTAGGTAATTCTTGTTGTGGG-BHQ-1 | 60 |
| 13,149 | F: TTAGACCTCATGAAAGGTCACGA R: GAGCCTTAGGATGGATCTCACA R6G-TGAAAGGGCGGCTCTAGCA-BHQ-1 FAM-TGAAACGGCGGCTCTAGCA-BHQ-1 | 60 |
| 16,970 | F: GGAATCTCCAGGACCGCCT R: CTGGTTCGTCATTGTGTCTACCA FAM-CACCTGATGCTGGACAGG-BHQ-1 R6G-CACCTGCTGCTGGACAGG-BHQ-1 | 60 |
| Reproductive Traits | Genotype | ||
|---|---|---|---|
| CC | CG | GG | |
| The period of recovery of the ovarian luteal activity: | n = 30 | n = 25 | n = 13 |
| <7 weeks after calving (I) | 50.0% | 44.0% | 53.8% |
| 7–13 weeks after calving (II) | 23.3% | 24.0% | 23.1% |
| >13 weeks after calving (III) | 26.7% | 32.0% | 23.1% |
| The open days period: | n = 29 | n = 21 | n = 11 |
| <120 days (I) | 48.3% a | 19.05% b | 45.45% |
| 120–240 days (II) | 34.5% c | 61.9% d | 9.10% e |
| 240–370 days (III) | 17.2% | 19.05% | 45.45% |
| The first service conception rate | n = 13 | n = 10 | n = 7 |
| 84.6% f | 40.0% g | 42.9% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostyunina, O.V.; Mityashova, O.S.; Bardukov, N.V.; Aleynikova, O.V.; Lebedeva, I.Y. DIO1 Gene Polymorphism Is Associated with Thyroid Profiles and Reproductive Performance in Dairy Cows. Agriculture 2023, 13, 398. https://doi.org/10.3390/agriculture13020398
Kostyunina OV, Mityashova OS, Bardukov NV, Aleynikova OV, Lebedeva IY. DIO1 Gene Polymorphism Is Associated with Thyroid Profiles and Reproductive Performance in Dairy Cows. Agriculture. 2023; 13(2):398. https://doi.org/10.3390/agriculture13020398
Chicago/Turabian StyleKostyunina, Olga V., Olga S. Mityashova, Nikolay V. Bardukov, Olga V. Aleynikova, and Irina Y. Lebedeva. 2023. "DIO1 Gene Polymorphism Is Associated with Thyroid Profiles and Reproductive Performance in Dairy Cows" Agriculture 13, no. 2: 398. https://doi.org/10.3390/agriculture13020398
APA StyleKostyunina, O. V., Mityashova, O. S., Bardukov, N. V., Aleynikova, O. V., & Lebedeva, I. Y. (2023). DIO1 Gene Polymorphism Is Associated with Thyroid Profiles and Reproductive Performance in Dairy Cows. Agriculture, 13(2), 398. https://doi.org/10.3390/agriculture13020398