
Amelioration of Hypothermia-Induced Damage on Peanut by Exogenous Application of Chitooligosaccharide


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Plant Materials
2.1.2. Experimental Agent
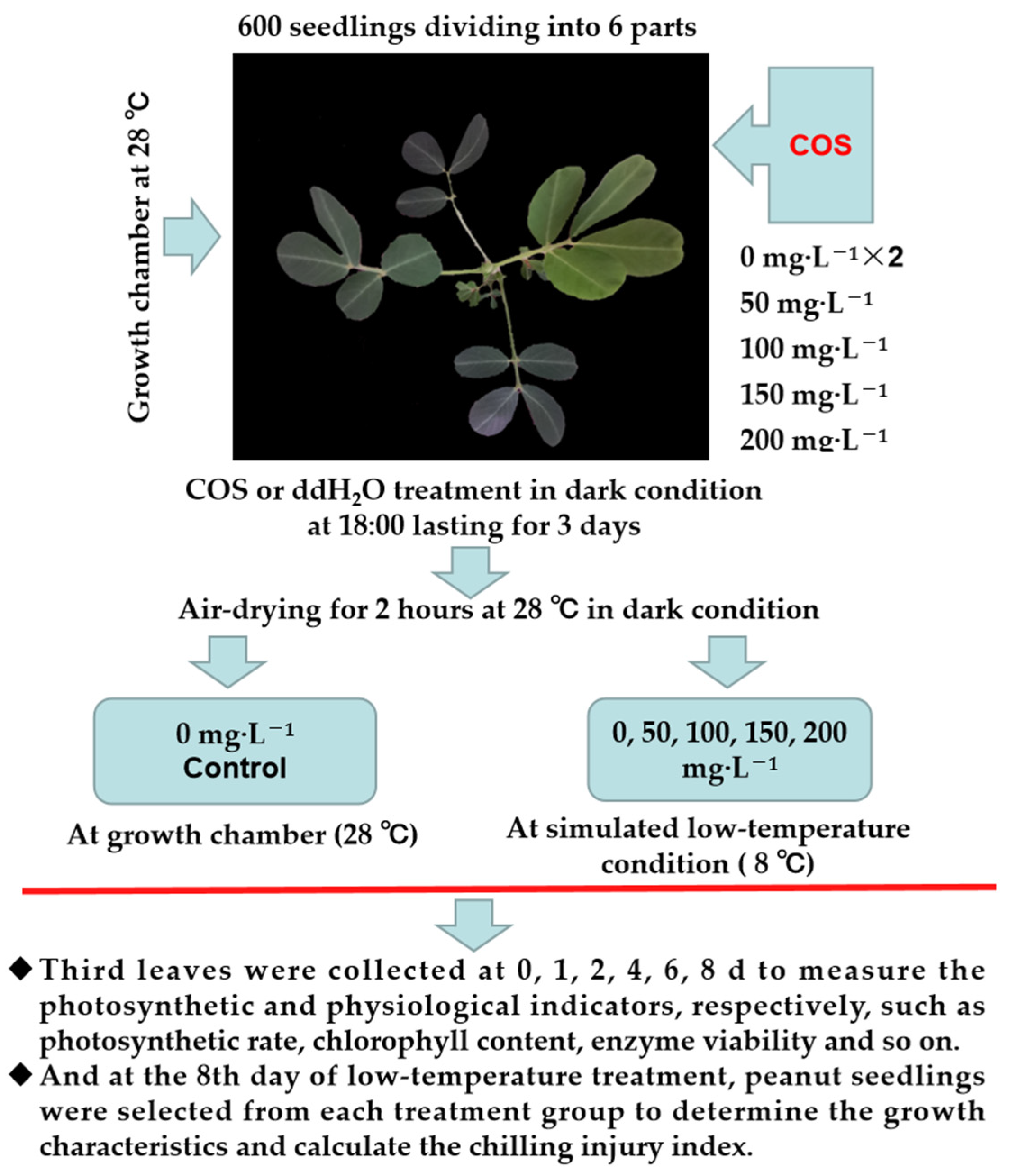
2.2. Experimental Design
2.2.1. Simulated Low-Temperature Growth Conditions
2.2.2. Growth Characteristics
2.2.3. Determination of the MDA and ROS Levels
2.2.4. Determination of Net Photosynthetic Rate and Chlorophyll Levels
2.2.5. Determination of Antioxidant Enzymatic Activities
2.2.6. Determination of Osmotic Substance Levels
2.3. Statistical Analysis
3. Results
3.1. Changes in Phenotypes of Peanut Affected by Chitooligosaccharide under Simulated Low-Temperature Conditions
3.2. Effects of Chitooligosaccharide on Lipid Peroxidation Contents of Peanut Seedlings under Simulated Low-Temperature Conditions
3.3. Effects of Chitooligosaccharides on the Photosynthetic efficiency of Peanut Seedlings under Simulated Low-Temperature Treatment
3.4. Effects of Chitooligosaccharide on Antioxidant-Related Enzyme Activities of Peanut Seedlings under Simulated Low-Temperature Conditions
3.5. Effects of Chitooligosaccharide on Osmotic Substance Contents of Peanut Seedlings under Simulated Low-Temperature Conditions
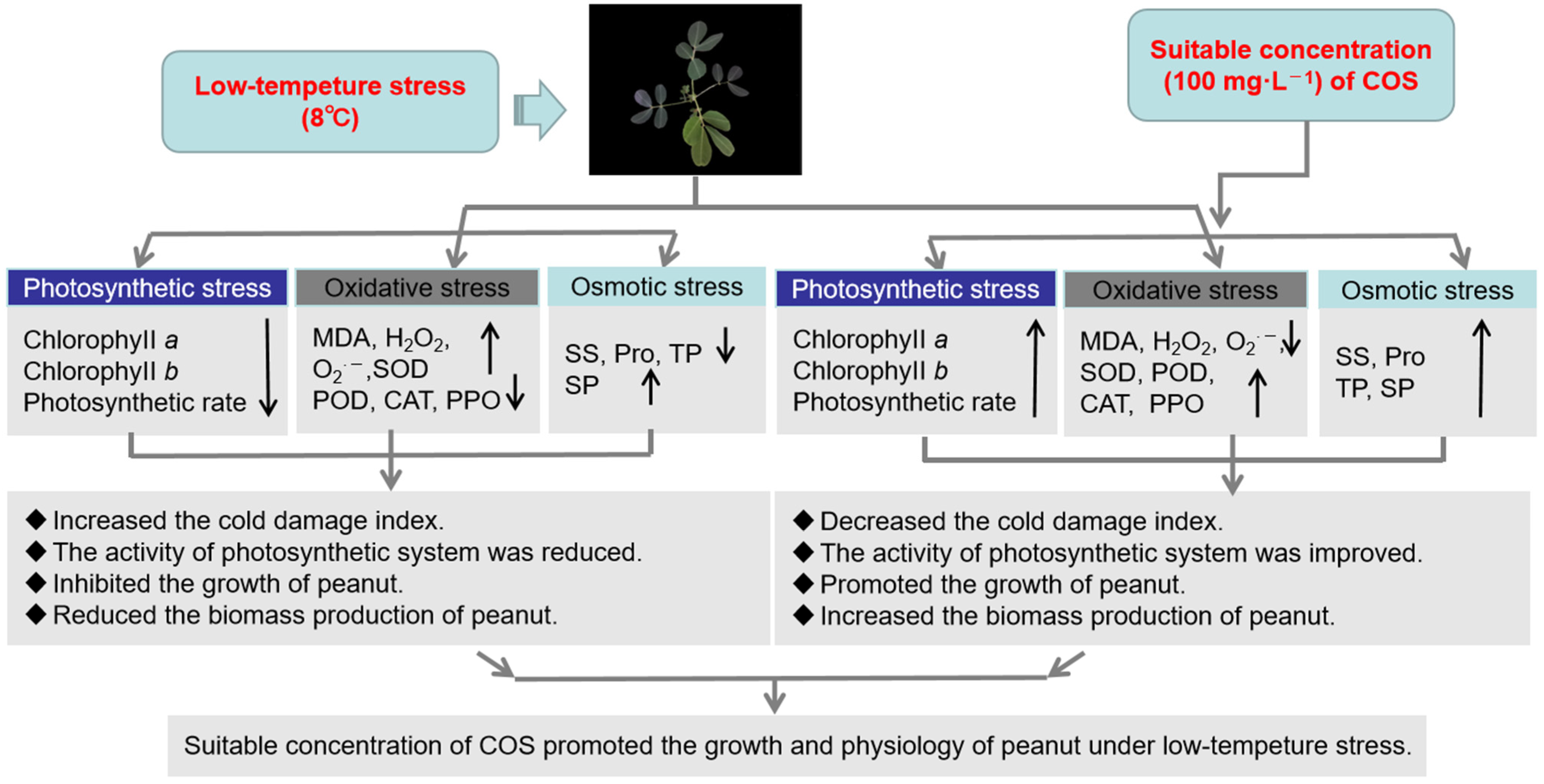
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, C.; Sun, Y.; Yang, W.; Zhang, S. Effect of spring low temperature and frost damage to the growth and yield of maize. J. Agric. Catastrophol. 2013, 3, 53–56. [Google Scholar]
- Ma, Z.; Hao, W.; Zheng, X.; Wang, L.; Xi, Y.; Hou, J. Effect of low temperature and freezing injury on whole growth period of sweet waxy corn in Wuzhai County of Shanxi Province and defensive measures. Agric. Eng. 2022, 12, 139–143. [Google Scholar]
- Xu, Y. Effect of low temperature and late freezing disaster on walnut growth in Chengxian county. J. Agric. Catastrophol. 2012, 2, 44–46. [Google Scholar]
- Mei, Y.Q.; Song, S.Q. Response to temperature stress of reactive oxygen species scavenging enzymes in the crosstolerance of barley seed germination. Zhejiang Univ. Sci. Biomed. 2010, 11, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jian, C.; Zhang, T.; Ma, C.; Shan, S. Effect of temperature on seed vigor of different peanut varieties. J. Peanut 2012, 41, 21–25. [Google Scholar]
- Pearce, R.S. Plant freezing and damage. Ann. Bot. 2001, 87, 417–424. [Google Scholar] [CrossRef]
- Wang, C.; Cheng, B.; Zheng, Y.; Sha, J.; Li, A.; Sun, X. Effects of temperature on peanut emergence, seedling growth and flowering. J. Peanut 2003, 32, 7–11. [Google Scholar]
- Tang, Y.; Wang, C.; Gao, H.; Feng, T.; Zhang, J.; Yu, S. Low temperature resistance and its correlation with quality traits during peanut seed imbibition. J. Nucl. Agric. 2011, 25, 436–442. [Google Scholar]
- Pan, R. Plant Physiology; Higher Education Press: Beijing, China, 2008. [Google Scholar]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Yan, W.; Li, J.; Wang, J.; Li, Z. Exogenous H2O2 improves the chilling tolerance of manilagrass and mascarenegrass by activating the antioxidative system. Plant Growth Regul. 2010, 61, 195–204. [Google Scholar]
- Aghdam, M.S.; Moradi, M.; Razavi, F.; Rabiei, V. Exogenous phenylalanine application promotes chilling tolerance in tomato fruits during cold storage by ensuring supply of NADPH for activation of ROS scavenging systems. Sci. Hortic. 2019, 246, 818–825. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Jannatizadeh, A.; Luo, Z.; Paliyath, G. Ensuring sufficient intracellular ATP supplying and friendly extracellular ATP signaling attenuates stresses, delays senescence and maintains quality in horticultural crops during postharvest life. Trends Food Sci. Technol. 2018, 76, 67–81. [Google Scholar] [CrossRef]
- Sharafi, Y.; Aghdam, M.S.; Luo, Z.; Jannatizadeh, A.; Razavi, F.; Fard, J.R.; Farmani, B. Melatonin treatment promotes endogenous melatonin accumulation and triggers gaba shunt pathway activity in tomato fruits during cold storage. Sci. Hortic. 2019, 254, 222–227. [Google Scholar] [CrossRef]
- Li, Q.; Li, L.L.; Hou, J.; Luo, R.R.; Wang, R.D.; Hu, J.B.; Huang, S. Advances on mechanism of cucurbit crops in response to lowtemperature stress. Acta Hortic. Sin. 2022, 49, 1382–1394. [Google Scholar]
- Mohammadrezakhani, S.; Rezanejad, F.; Hajilou, J. Effect of putrescine and proline on proflies of GABA, antioxidant activities in leaves of three Citrus species in response to low temperature stress. J. Plant Biochem. Biotechnol. 2021, 30, 545–553. [Google Scholar] [CrossRef]
- Pan, M.; Yin, Y.; Shen, F.; Luo, B.; Tang, X.; He, H.; Chen, D. Effect of calcalate ate on physiological indicators of tobacco seedlings under cold stress. J. Southwest Agric. 2016, 29, 288–293. [Google Scholar]
- Li, L. Mitigation effect of exogenous glutathione on maize seedlings under cold stress. Master’s Thesis, Heilongjiang University, Harbin, China, 2022. [Google Scholar]
- Xu, M. Effect of DA-6 on seed germination and photosynthetic characteristics of cotton seedlings. Master’s Thesis, Shihezi University, Shihezi, China, 2021. [Google Scholar]
- Cao, S.; Song, C.; Shao, J.; Bian, K.; Chen, W.; Yang, Z. Exogenous melatonin treatment increases chilling tolerance andinduces defense response in harvested peach fruit during cold storage. J. Agric. Food Chem. 2016, 64, 5215–5222. [Google Scholar] [CrossRef]
- He, Y.Q.; Bose, S.K.; Wang, M.Y.; Liu, T.; Wang, W.; Lu, H.; Yin, H. Effects of chitosan oligosaccharides postharvest treatment on the quality and ripening related gene expression of cultivated strawberry fruits. J. Berry Res. 2019, 9, 11–25. [Google Scholar] [CrossRef]
- Shi, X.; Yang, Y.; Hou, X.; Xue, X.; Li, M.; Duan, M.; Huo, J.; Zen, F. Effect of exogenous chitooligosaccharides on drought resistance of Tanggut white thorn. Jiangsu Agric. Sci. 2020, 48, 172–177. [Google Scholar]
- Gu, L. Effect of chitooligosaccharides on tomato seed germination. J. Southwest Agric. 2014, 27, 1233–1236. [Google Scholar]
- Lu, J.; Sun, D.; Zhang, C.; Qi, S.; Li, H. Effect of chitooligosaccharides on pepper seed germination and seedling antioxidant enzyme activity. Wild Plant Resour. China 2012, 31, 12–16. [Google Scholar]
- Jiang, R.; Li, L.; Li, Y.; Wang, S.; Liu, C.; Huang, X.; He, J. Effect of IAA, GA3 and chitooligosaccharide species on germination of Yunnan seeds. West. For. Sci. 2021, 50, 110–116+123. [Google Scholar]
- Ali, A.; Zhang, J.J.; Zhou, M.M.; Chen, T.; Liaqat, S.; ur Rehman, S.; Hayat, S.; Shi, J.; Chen, J. Chitosan oligosaccharides stimulate the efficacy of somatic embryogenesis in different genotypes of the Liriodendron hybrid. Forests 2021, 12, 557. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.; Ou, L.; Ji, D.; Liu, T.; Lan, R.; Li, X.; Jin, L. Response to the cold stress signaling of the tea plant (Camellia sinensis) elicited by chitosan oligosaccharide. Agronomy 2020, 10, 915. [Google Scholar] [CrossRef]
- Jia, X.C.; Qin, H.Q.; Bose, S.K.; Liu, T.; He, J.; Xie, S.; Ye, M.; Yin, H. Proteomics analysis reveals the defense priming effect of chitosan oligosaccharides in Arabidopsis-Pst DC3000 interaction. Plant Physiol. Biochem. 2020, 149, 301–312. [Google Scholar] [CrossRef]
- Liu, J.; Ou, X.; Wang, J. Effect of exogenous H2O2 on leaf physiological characteristics of naked oat seedlings under drought stres. Agric. Res. Arid Areas 2019, 37, 146–153. [Google Scholar]
- Cheplick, S.; Sarkar, D.; Bhowmik, P.C.; Shetty, K. Improved resilience and metabolic response of transplanted blackberry plugs using chitosan oligosaccharide elicitor treatment. Can. J. Plant Sci. 2018, 98, 717–731. [Google Scholar] [CrossRef]
- Song, L. Jiangxi Oil Crop Industry Development Report; Jiangxi Science and Technology Publishing House: Nanchang, China, 2011; pp. 63–64. [Google Scholar]
- Wu, X.; Yang, X.; Cha, D.; Zhu, Z.; Xu, S. Physiological mechanism of low-temperature tolerance in eggplant seedlings. Shanghai Agric. J. 2013, 29, 45–49. [Google Scholar]
- Zou, Q. Experimental Guidance of Plant Physiology; China Agriculture Press: Beijing, China, 2003. [Google Scholar]
- Wang, N.; Chu, Y. Effect of low temperature on the activity of osmoregulators and protective enzymes in peanut seedlings. Anhui Agric. Sci. 2007, 35, 9154–9156. [Google Scholar]
- Peng, W.; Guo, X. Effect of cold injury stress on the physiology and structure of watermelon seedlings. Chin. Melon Veg. 2017, 30, 8–12. [Google Scholar]
- Zhao, X.; Liang, T.; Zhang, H. Effects of chitooligosaccharide on seed germination, seedling growth and osmotic substances in wheat under PEG stress. Seeds 2020, 39, 91–95. [Google Scholar]
- Guo, W.H.; Zhao, X.M.; Du, Y.G. Effects of oligochitosan on the growth and photosynthesis and physiological index related to photosynthesis of tobacco seedlings. Plant Physiol. Commun. 2008, 1155–1157. [Google Scholar]
- Li, Y.; Zhao, X.M.; Xia, X.Y. Effects of oligochitosan on photosynthetic parameter of Brassica napus seedlings under drought stress. Acta Agron. Sin. 2008, 34, 326–329. [Google Scholar] [CrossRef]
- Prasad, P.; Boote, K.; Thomas, J.; Allen, L.H., Jr.; Gorbet, D.W. Influence of soil temperature on seedling emergence and early growth of peanut cultivars in field conditions. J. Agron. Crop Sci. 2006, 192, 168–177. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, F.; Hussain, H.A.; Nie, L. Physiological and biochemical mechanisms of seed priming-induced chilling tolerance in rice cultivars. Front. Plant Sci. 2016, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Reiter, R.J.; Chan, Z. Phytomelatonin: A universal abiotic stress regulator. J. Exp. Bot. 2018, 69, 963–974. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Zheng, J.; Dong, Y.; Liu, Q.; Yang, X.; Wei, C.H.; Zhang, Y.; Ma, F.W.; Zhang, X. Local melatonin application induces cold tolerance in distant organs of Citrullus lanatus L. Via long distance transport. Sci. Rep. 2017, 7, 40858. [Google Scholar]
- Du, B.; Luo, H.W.; He, L.X.; Zhang, L.; Liu, Y.; Mo, Z.; Pan, S.; Tian, H.; Duan, M.; Tang, X. Rice seed priming with sodium selenate: Effects on germination, seedling growth, and biochemical attributes. Sci. Rep. 2019, 9, 3631–3642. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Y.; Zhao, X.; Xu, T.; Jiang, Y.; Liu, T.; Zhou, K. H2O2 Regulation of peanut seed germination under low temperature stress. Chin. J. Chin. Oil Crops 2020, 42, 860–868. [Google Scholar]
- Chang, B.; Zhong, P.; Liu, J.; Tang, Z.; Gao, Y.; Yu, H.; Guo, W. Effect of cold stress and gibberellin on peanut seed germination and seedling physiological responses. J. Crop Sci. 2019, 45, 118–130. [Google Scholar]
- Dong, D.; Li, Y.; Jiang, L.; Liang, H.; Huang, J. Long-acting brassinosteroid TS303 and Acpropyl dihydrojasmonate enhances cold resistance in peanut. Guangxi Plants 2008, 28, 675–680. [Google Scholar]
- Li, C.; Ying, C.; Lin, Y.; Ye, H.; Zheng, X.; Xu, G. Effect of chilling stress the RPP, soluble protein and protective enzyme of Arachis. Duranensis leafs. China Grass Ind. Dev. Forum. 2008, 302–305. [Google Scholar]
- Liang, Y.; Chen, Q.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.H.; Wu, Y.; Zeng, J.Z.; Zeng, Q.M. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery. Photosynthetica 2010, 48, 537–544. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Name | Plant Height/cm | Main Root Length/cm | Fresh Weight/g | Dry Weight/g | Cold Damage Index | |
|---|---|---|---|---|---|---|
| Aboveground Part | Belowground Part | |||||
| Control | 15.63 ± 0.96 a | 13.58 ± 0.47 a | 10.17 ± 11.42 a | 1.22 ± 1.24 a | 0.27 ± 0.29 a | 0.00 ± 0.00 e |
| 0 mg·L−1 | 10.25 ± 0.70 d | 8.36 ± 0.35 e | 4.37 ± 4.84 d | 0.59 ± 0.73 e | 0.13 ± 0.16 e | 0.85 ± 0.00 a |
| 50 mg·L−1 | 11.87 ± 0.90 bc | 9.27 ± 0.29 d | 5.37 ± 9.67 c | 0.82 ± 0.85 d | 0.18 ± 0.22 d | 0.78 ± 0.02 b |
| 100 mg·L−1 | 12.62 ± 0.75 b | 12.64 ± 0.73 b | 9.68 ± 11.24 a | 1.17 ± 1.24 a | 0.24 ± 0.28 a | 0.63 ± 0.03 d |
| 150 mg·L−1 | 11.87 ± 0.28 bc | 11.21 ± 1.26 c | 8.49 ± 9.64 b | 1.04 ± 1.13 b | 0.22 ± 0.24 b | 0.73 ± 0.02 c |
| 200 mg·L−1 | 11.3 ± 0.72 c | 10.06 ± 0.25 d | 8.43 ± 8.71 b | 1.03 ± 1.08 c | 0.2 ± 0.22 c | 0.77 ± 0.01 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Sun, Z.; Xue, X.; Xu, H.; Wu, Y.; Zhang, Y.; Yang, Y.; Han, S.; Zhao, R.; Zhang, M.; et al. Amelioration of Hypothermia-Induced Damage on Peanut by Exogenous Application of Chitooligosaccharide. Agriculture 2023, 13, 217. https://doi.org/10.3390/agriculture13010217
Shi X, Sun Z, Xue X, Xu H, Wu Y, Zhang Y, Yang Y, Han S, Zhao R, Zhang M, et al. Amelioration of Hypothermia-Induced Damage on Peanut by Exogenous Application of Chitooligosaccharide. Agriculture. 2023; 13(1):217. https://doi.org/10.3390/agriculture13010217
Chicago/Turabian StyleShi, Xinlong, Ziqi Sun, Xian Xue, Huimin Xu, Yue Wu, Ya Zhang, Yueqin Yang, Suoyi Han, Ruifang Zhao, Mengyuan Zhang, and et al. 2023. "Amelioration of Hypothermia-Induced Damage on Peanut by Exogenous Application of Chitooligosaccharide" Agriculture 13, no. 1: 217. https://doi.org/10.3390/agriculture13010217
APA StyleShi, X., Sun, Z., Xue, X., Xu, H., Wu, Y., Zhang, Y., Yang, Y., Han, S., Zhao, R., Zhang, M., & Zhang, X. (2023). Amelioration of Hypothermia-Induced Damage on Peanut by Exogenous Application of Chitooligosaccharide. Agriculture, 13(1), 217. https://doi.org/10.3390/agriculture13010217