

Pseudomonas syringae Infection Modifies Chlorophyll Fluorescence in Nicotiana tabacum
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Inoculation of Tobacco Leaves

{kind=link}
{kind=link}
{kind=link}
| Chl-Fluorescence Parameters | Meaning |
| F0 | Minimum fluorescence when all RCs are open at the onset of fluorescence induction |
| Fm | Maximum fluorescence when all RCs are closed |
| Fv = Fm − F0 | Maximum variable fluorescence |
| φP0 = Fv/Fm | Maximum quantum efficiency of PSII |
| F0′ = F0/[(Fv/Fm) + (F0/Fm′)] | Minimum fluorescence for the light-adapted state |
| Fm′ | Maximum fluorescence for the light-adapted state |
| F′ | Actual (stationary) fluorescence during the slow fluorescence induction |
| Fv′ = Fm′ − F0′ | Variable fluorescence for a light-adapted state |
| ΔF = Fm′ − F′ | Difference in fluorescence between Fm′ and F′ |
| φPSII = Fv′/Fm′ | Maximum efficiency of PSII photochemistry for the light-adapted state, if all centers are open |
| qP = ΔF/Fv′ | Photochemical quenching coefficient |
| qL = qP/(F0′/F′) | Estimate of the fraction of open PSII RCs |
| NPQ = (Fm − Fm′)/Fm′ | Non-photochemical Chl fluorescence quenching |
| PQ = Fm/F′ − Fm/Fm′ | Photochemical Chl fluorescence quenching |
| Energy Partitioning | Meaning |
| φP = ΔF/Fm′ = φPSII × qP | Actual photochemical quantum yield of PSII photochemistry for the light-adapted state |
| φNPQ1 = F′/Fm′ − F′/Fm | Quantum yield of regulatory non-photochemical quenching after Genty et al., Cailly et al. and Hendrickson et al. [21,22,23] |
| φNPQ2 = (1 − (φPSII/0.8)) × (F′/F0′) | Quantum yield of regulatory non-photochemical quenching after Kornyeyev and Holaday [24] |
| φf,D1 = F′/Fm | Quantum yield of constitutive non-regulatory basal dissipation processes consisting of fluorescence emission and heat dissipation after Genty et al., Cailly et al. and Hendrickson et al. [21,22,23] |
| φf,D2 = (F′/Fm′) + (φPSII/φP0) × (F′/F0′) − (F′/F0′) | Quantum yield of constitutive non-regulatory basal dissipation processes consisting of fluorescence emission after Kornyeyev and Holaday [24] |
| φNF2 = (1 − (0.8/φP0)) × (φPSII/0.8) × (F′/F0′) | Quantum yield of heat dissipation by the photo-inactivated non-functional centers after Kornyeyev and Holaday [24] |
2.2. Chl a Fluorescence Procedures
2.3. Statistical Analyses
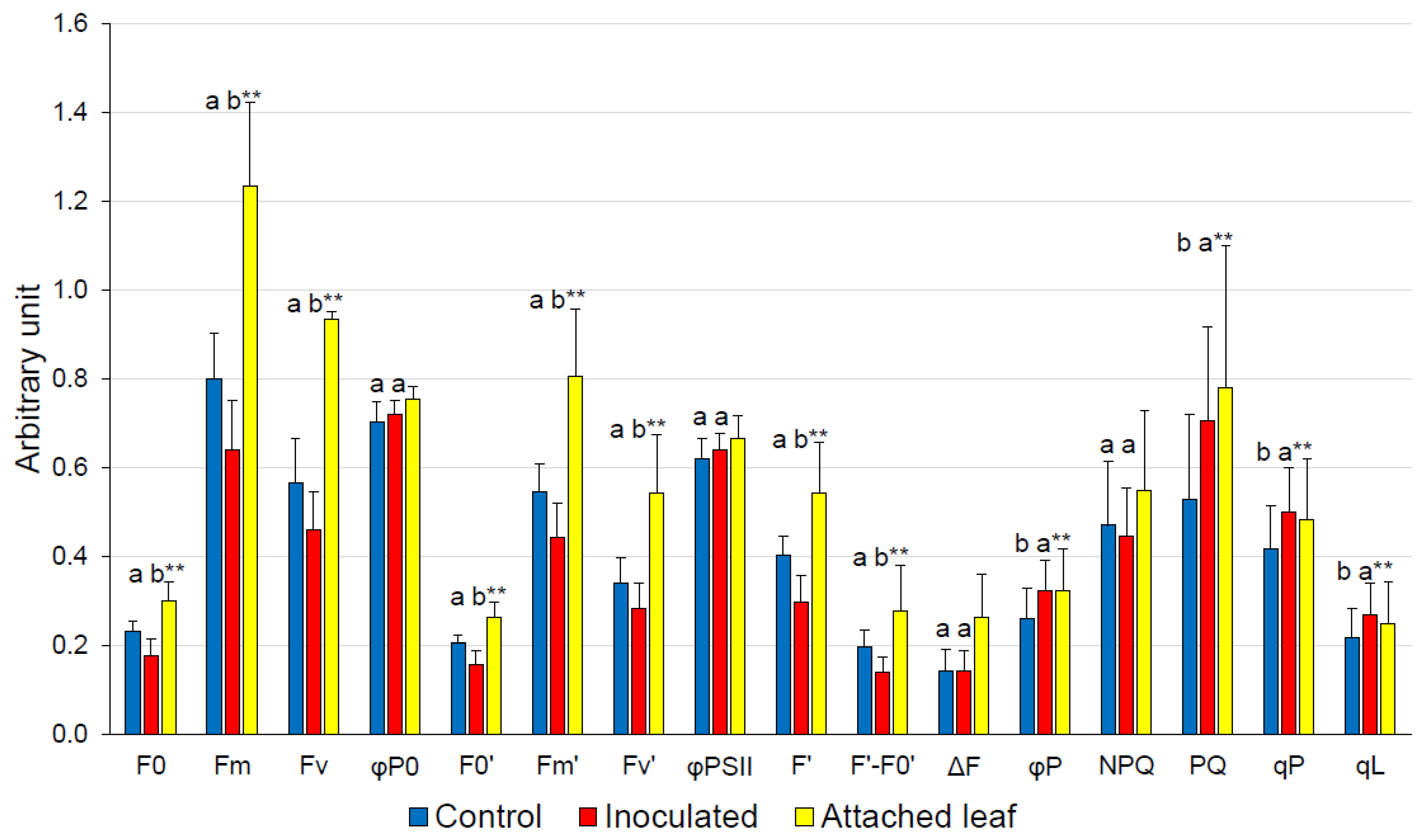
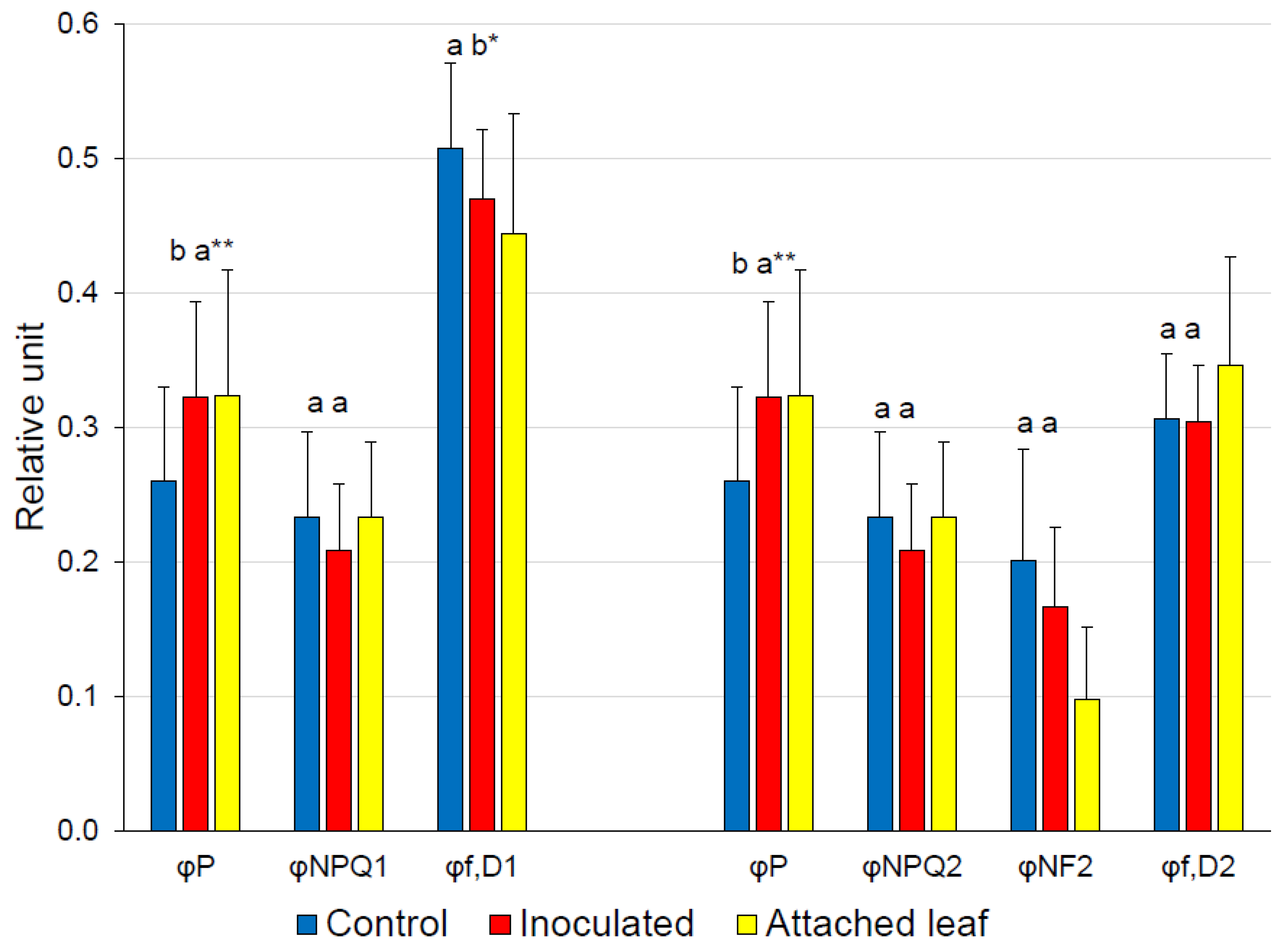
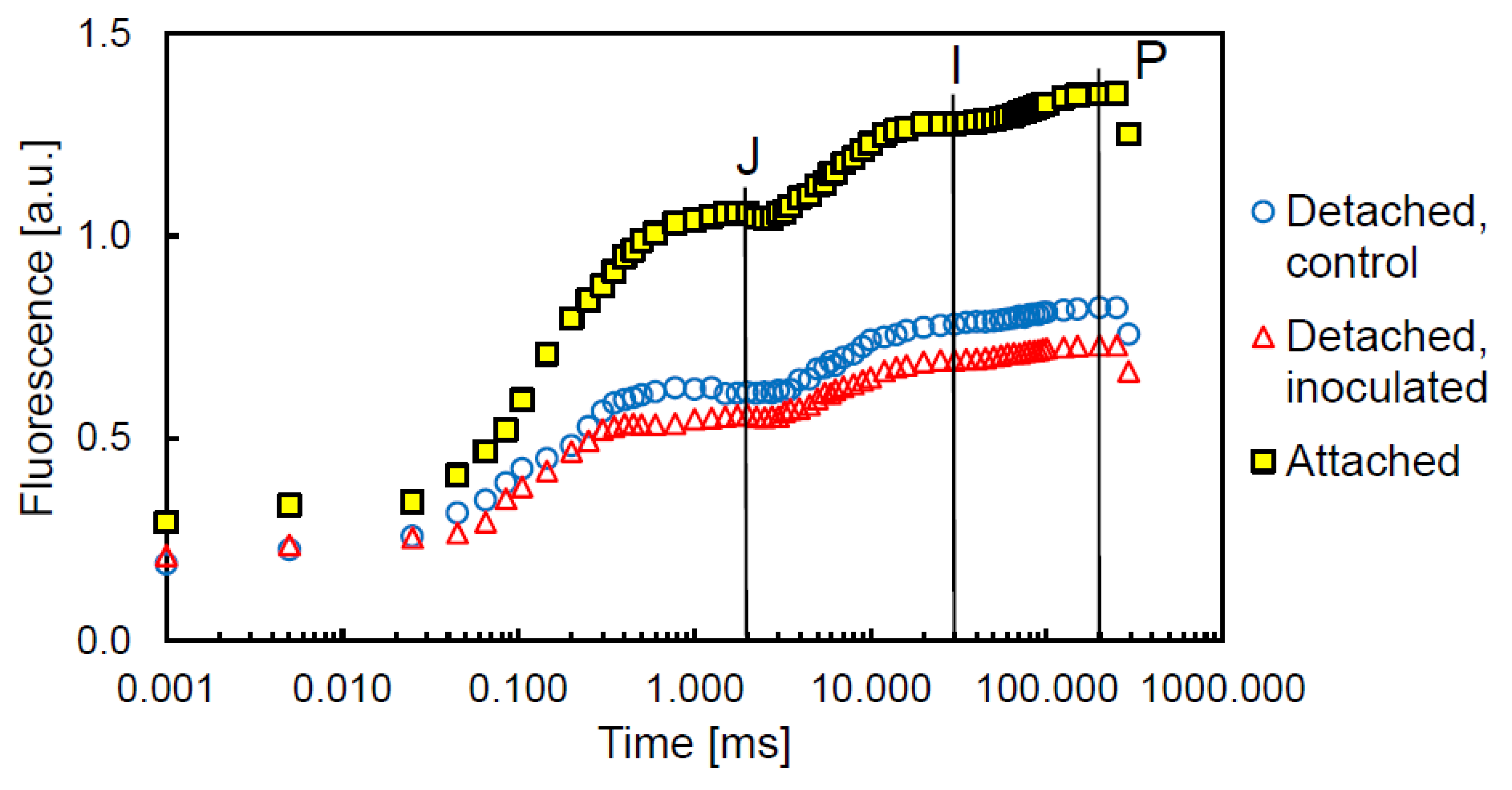
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, C.F.; Kvitko, B.H.; Shimizu, R.; Crabill, E.; Alfano, J.R.; Lin, N.C.; Martin, G.B.; Huang, H.C.; Collmer, A. A Pseudomonas syringae pv. tomato DC3000 mutant lacking the type III effector HopQ1-1 is able to cause disease in the model plant Nicotiana benthamiana. Plant J. 2007, 51, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Mysore, K.S.; Ryu, C.M. Nonhost resistance: How much do we know? Trends Plant Sci. 2004, 9, 97–104. [Google Scholar] [CrossRef]
- Cheng, D.D.; Zhang, Z.S.; Sun, X.B.; Zhao, M.; Sun, G.Y.; Chow, W.S. Photoinhibition and photoinhibition-like damage to the photosynthetic apparatus in tobacco leaves induced by Pseudomonas syringae pv. Tabaci under light and dark conditions. BMC Plant Biol. 2016, 16, 29. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Díaz-Casado, E.; Barón, M. Spatial and temporal dynamics of primary and secondary metabolism in Phaseolus vulgaris challenged by Pseudomonas syringae. Physiol. Plant. 2015, 153, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Bonfig, K.B.; Schreiber, U.; Gabler, A.; Roitsch, T.; Berger, S. Infection with virulent and avirulent P. syringae strains differentially affects photosynthesis and sink metabolism in Arabidopsis leaves. Planta 2006, 225, 1–12. [Google Scholar] [CrossRef]
- Rodríguez-Moreno, L.; Pineda, M.; Soukupová, J.; Macho, A.P.; Beuzón, C.R.; Barón, M.; Ramos, C. Early detection of bean infection by Pseudomonas syringae in asymptomatic leaf areas using chlorophyll fluorescence imaging. Photosynth. Res. 2008, 96, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Nguyen, D.; Rieu, I.; Mariani, C.; van Dam, N.M. How plants handle multiple stresses: Hormonal interactions underlying responses to abiotic stress and insect herbivory. Plant Mol. Biol. 2016, 91, 727–740. [Google Scholar] [CrossRef]
- Thevenet, D.; Pastor, V.; Baccelli, I.; Balmer, A.; Vallat, A.; Neier, R.; Glauser, G.; Mauch-Mani, B. The priming molecule β-aminobutyric acid is naturally present in plants and is induced by stress. New Phytol. 2017, 213, 552–559. [Google Scholar] [CrossRef]
- Pathak, R.; Singh, S.K.; Tak, A.; Gehlot, P. Impact of climate change on host, pathogen and plant disease adaptation regime: A review. Biosci. Biotechnol. Res. Asia 2018, 15, 529–540. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed]
- Boonekamp, P.M. Are plant diseases too much ignored in the climate change debate? Eur. J. Plant Pathol. 2012, 133, 291–294. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, S.; Kumar, R.; Kumar, R.; Imran, M. Impact of climate change on plant diseases and their management strategies. J. Pharmacogn. Phytochem. 2017, 6, 779–781. Available online: https://www.phytojournal.com/archives/2017/vol6issue6S/PartR/SP-6-6-176.pdf (accessed on 14 September 2022).
- Hanemian, M.; Zhou, B.; Deslandes, L.; Marco, Y.; Tremousaygue, D. Hrp mutant bacteria as biocontrol agents: Toward a sustainable approach in the fight against plant pathogenic bacteria. Plant Signal. Behav. 2013, 8, e25678. [Google Scholar] [CrossRef][Green Version]
- Velásquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant–pathogen warfare under changing climate conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef]
- CORESTA—Cooperation Centre for Scientific Research Relative to Tobacco. A Scale of Coding Growth in Tobacco Crops; CORESTA Guide No. 7; CORESTA: Paris, France, 2019; p. 15. Available online: https://www.coresta.org/scale-coding-growth-stages-tobacco-crops-29211.html (accessed on 14 September 2022).
- Oxborough, K.; Baker, N.R. Resolving chlorophyll a fluorescence images of photosynthetic efficiency into photochemical and non-photochemical components—Calculation of qP and Fv’/Fm’ without measuring Fo’. Photosynth. Res. 1997, 54, 135–142. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Lazár, D. Parameters of photosynthetic energy partitioning. J. Plant Physiol. 2015, 175, 131–147. [Google Scholar] [CrossRef]
- Genty, B.; Harbinson, J.; Cailly, A.L.; Rizza, F. Fate of excitation at PSII in leaves: The non-photochemical side. In Book of Abstract, Proceedings of the Third BBSRC Robert Hill Symposium on Photosynthesis, 31 March–3 April 1996, Abstract P 18; Department of Molecular Biology and Biotechnology, University of Sheffield: Sheffield, UK.
- Cailly, A.L.; Rizzal, F.; Genty, B.; Harbinson, J. Fate of excitation at PSII in leaves, the nonphotochemical side. Plant Physiol. Biochem. 1996, 86. [Google Scholar]
- Hendrickson, L.; Furbank, R.T.; Chow, W.S. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence. Photosynth. Res. 2004, 82, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Kornyeyev, D.; Holaday, A.S. Corrections to current approaches used to calculate energy partitioning in photosystem 2. Photosynthetica 2008, 46, 170–178. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Schansker, G.; Brestic, M.; Bussotti, F.; Calatayud, A.; Ferroni, L.; Goltsev, V.; Guidi, L.; Jajoo, A.; Li, P.; et al. Frequently asked questions about chlorophyll fluorescence, the sequel. Photosynth. Res. 2017, 132, 13–66, Erratum in Photosynth. Res. 2017, 132, 67–68. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Goltsev, V.N.; Żuk-Gołaszewska, K.; Zivcak, M.; Brestic, M. Chlorophyll Fluorescence: Understanding Crop Performance—Basics and Applications; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar] [CrossRef]
- Keutgen, N.; Tomaszewska-Sowa, M.; Keutgen, A.J. Chlorophyll fluorescence of Nicotiana tabacum expressing the green fluorescent protein. Photosynthetica 2020, 58, 275–282. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient: Chlorophyll a fluorescence. In Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. BBA-Bioenergetics 2010, 1797, 1313–1326. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Kromdijk, J.; Govindjee. Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Weng, J.H.; Chien, C.T.; Chen, C.W.; Lai, X.M. Effects of osmotic- and high-light stresses on PSII efficiency of attached and detached leaves of three tree species adapted to different water regimes. Photosynthetica 2011, 49, 555–563. [Google Scholar] [CrossRef]
- McAdam, S.; Brodribb, T. Mesophyll cells are the main site of abscisic acid biosynthesis in water-stressed leaves. Plant Physiol. 2018, 177, 911–917. [Google Scholar] [CrossRef]
- De Torres-Zabala, M.; Truman, W.; Bennett, M.H.; Lafforgue, G.; Mansfield, J.W.; Rodriguez Egea, P.; Bögre, L.; Grant, M. Pseudomonas syringae pv. tomato hijacks the Arabidopsis abscisic acid signalling pathway to cause disease. EMBO J. 2007, 26, 1434–1443. [Google Scholar] [CrossRef]
- Jia, H.; Lu, C. Effects of abscisic acid on photoinhibition in maize plants. Plant Sci. 2003, 165, 1403–1410. [Google Scholar] [CrossRef]
- Lu, S.; Su, W.; Li, H.; Guo, Z. Abscisic acid improves drought tolerance of triploid bermudagrass and involves H2O2- and NO-induced antioxidant enzyme activities. Plant Physiol. Biochem. 2009, 47, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Desotgiu, R.; Cascio, C.; Pollastrini, M.; Gerosa, G.; Marzuoli, R.; Bussotti, F. Short and long term photosynthetic adjustments in sun and shade leaves of Fagus sylvatica L., investigated by fluorescence transient (FT) analysis. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. Off. J. Soc. Bot. Ital. 2012, 146 (Suppl. S1), 206–216. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Balatova, Z.; Drevenakova, P.; Olsovska, K.; Kalaji, H.M.; Yang, X.; Allakhverdiev, S.I. Photosynthetic electron transport and specific photoprotective responses in wheat leaves under drought stress. Photosynth. Res. 2013, 117, 529–546. [Google Scholar] [CrossRef] [PubMed]
- Umar, M.; Uddin, Z.; Siddiqui, Z.S. Responses of photosynthetic apparatus in sunflower cultivars to combined drought and salt stress. Photosynthetica 2019, 57, 627–639. [Google Scholar] [CrossRef]
- Mur, L.A.J.; Kenton, P.; Lloyd, A.J.; Ougham, H.; Prats, E. The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot. 2008, 59, 501–520. [Google Scholar] [CrossRef]
- Vidal, G.; Ribas–Carbo, M.; Garmier, M.; Dubertret, G.; Rasmusson, A.G.; Mathieu, C.; Foyer, C.H.; De Paepe, R. Lack of respiratory chain complex I impairs alternative oxidase engagement and modulates redox signaling during elicitor-induced cell death in tobacco. Plant Cell 2007, 19, 640–655. [Google Scholar] [CrossRef]
- Hirano, S.S.; Upper, C.D. Bacteria in the leaf ecosystem with emphasis on Pseudomonas syringae—a pathogen, ice nucleus and epiphyte. Microbiol. Mol. Biol. Rev. 2000, 64, 624–653. [Google Scholar] [CrossRef]



| Chl-Fluorescence Parameters | Meaning |
| Vt = (Ft − F0)/Fv | Relative variable fluorescence, e.g., at the ‘J’ (t = 2 ms) and ‘I’ steps (t = 30 ms) |
| M0 = (ΔV/Δt)0 ≅ 4(F0.3ms − F0)/Fv | Initial slope (in ms−1) of the F0–J fluorescence rise |
| Efficiencies and Quantum Yields | Meaning |
| ψE0 = 1 − VJ | Efficiency with which a PSII-trapped electron is transferred from quinone A− (QA−) to plastoquinone |
| φE0 = φP0 × ψE0 | Quantum yield of electron transport from QA− to plastoquinone |
| ψR0 = 1 − VI | Efficiency with which a PSII-trapped electron is transferred to final PSI acceptors |
| φR0 = φP0 × ψR0 | Quantum yield of electron transport from QA− to final PSI acceptors |
| δR0 = ψR0/ψE0 | Efficiency with which an electron from plastoquinol is transferred to final PSI acceptors |
| Specific Energy Fluxes (per Active PSII) | Meaning |
| ABS/RC = (M0/VJ)/φP0 | Absorption flux of antenna Chls per RC; apparent antenna size of an active PSII |
| Performance Indexes | Meaning |
| PIABS = (RC/ABS) × [φP0/(1 − φP0)] × [ψE0/(1 − ψE0)] | Performance index for energy conservation of photons absorbed by PSII to the reduction in intersystem electron acceptors |
| PITotal = PIABS × [δR0/(1 − δR0)] | Performance index for energy conservation from photons absorbed by PSII to the reduction in PSI end acceptors |
| Transformation of RCs to So-Called Silent RCs | Meaning |
| RCsi/RCat = 100 × (1 − (ABS/RCat)/(ABS/RCde)) | Fraction of detached leaves that is transformed into silent RCs relative to an attached leaf in percent (si: silent; at: attached; de: detached) |
| Parameter | Control | Inoculated | PKS-test | PSW-test | PF-test | Pt-test | PMW-test |
|---|---|---|---|---|---|---|---|
| ABS/RC [ms−1] | 4.596 ± 0.496 | 4.503 ± 0.604 | 0.002 ** | 0.001 ** | 0.117 | ||
| φE0 [au] | 0.246 ± 0.046 | 0.254 ± 0.043 | 0.200 x | 0.799 | 0.611 | 0.369 | |
| δR0 [au] | 0.174 ± 0.037 | 0.196 ± 0.037 | 0.200 x | 0.769 | 0.931 | 0.009 ** | |
| φR0 [au] | 0.043 ± 0.013 | 0.050 ± 0.012 | 0.200 x | 0.749 | 0.507 | 0.019 * | |
| PIABS [au] | 0.325 ± 0.116 | 0.362 ± 0.139 | 0.200 x | 0.949 | 0.260 | 0.211 | |
| PITotal [au] | 0.072 ± 0.036 | 0.088 ± 0.040 | 0.200 x | 0.065 | 0.586 | 0.066 | |
| RCsi/RCat [%] | 10.7 ± 9.6 | 8.5 ± 10.8 | 0.200 x | 0.639 | 0.461 | 0.332 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomaszewska-Sowa, M.; Keutgen, N.; Lošák, T.; Figas, A.; Keutgen, A.J. Pseudomonas syringae Infection Modifies Chlorophyll Fluorescence in Nicotiana tabacum. Agriculture 2022, 12, 1504. https://doi.org/10.3390/agriculture12091504
Tomaszewska-Sowa M, Keutgen N, Lošák T, Figas A, Keutgen AJ. Pseudomonas syringae Infection Modifies Chlorophyll Fluorescence in Nicotiana tabacum. Agriculture. 2022; 12(9):1504. https://doi.org/10.3390/agriculture12091504
Chicago/Turabian StyleTomaszewska-Sowa, Magdalena, Norbert Keutgen, Tomáš Lošák, Anna Figas, and Anna J. Keutgen. 2022. "Pseudomonas syringae Infection Modifies Chlorophyll Fluorescence in Nicotiana tabacum" Agriculture 12, no. 9: 1504. https://doi.org/10.3390/agriculture12091504
APA StyleTomaszewska-Sowa, M., Keutgen, N., Lošák, T., Figas, A., & Keutgen, A. J. (2022). Pseudomonas syringae Infection Modifies Chlorophyll Fluorescence in Nicotiana tabacum. Agriculture, 12(9), 1504. https://doi.org/10.3390/agriculture12091504