

Identification and Expression Profiling of TGA Transcription Factor Genes in Sugarcane Reveals the Roles in Response to Sporisorium scitamineum Infection
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. S. scitamineum Inoculation and Applied Treatments
2.3. Sequence Retrieval
2.4. Screening and Identification of TGA Family Genes in Sugarcane
2.5. Analysis of Gene Structure and Conserved Motifs
2.6. Phylogenetic Analysis of ShTGA Proteins and Calculation of Ka/Ks
2.7. Cis-Regulatory Elements Prediction of ShTGA Gene Family
2.8. Experimental Validation of ShTGA Gene Expression Levels by qRT-PCR
2.9. Statistical Analysis
3. Results
3.1. Screening TGA Transcription Factors Family Genes for Basic Physic-Chemical Properties of ShTGA Coding Proteins
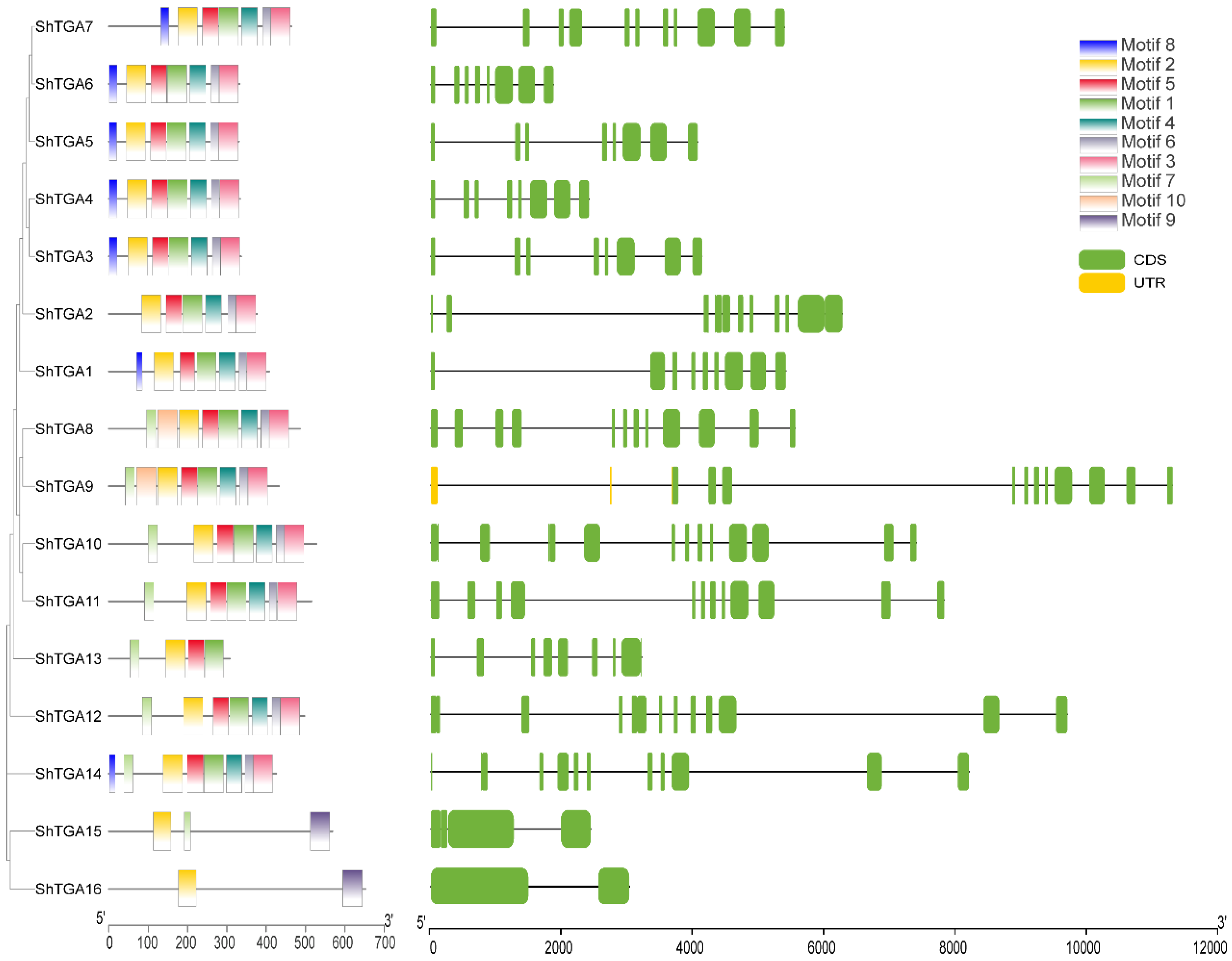
3.2. Gene Structures and Protein Domains Analysis of ShTGAs
3.3. Evolution Analysis of ShTGA Family Members
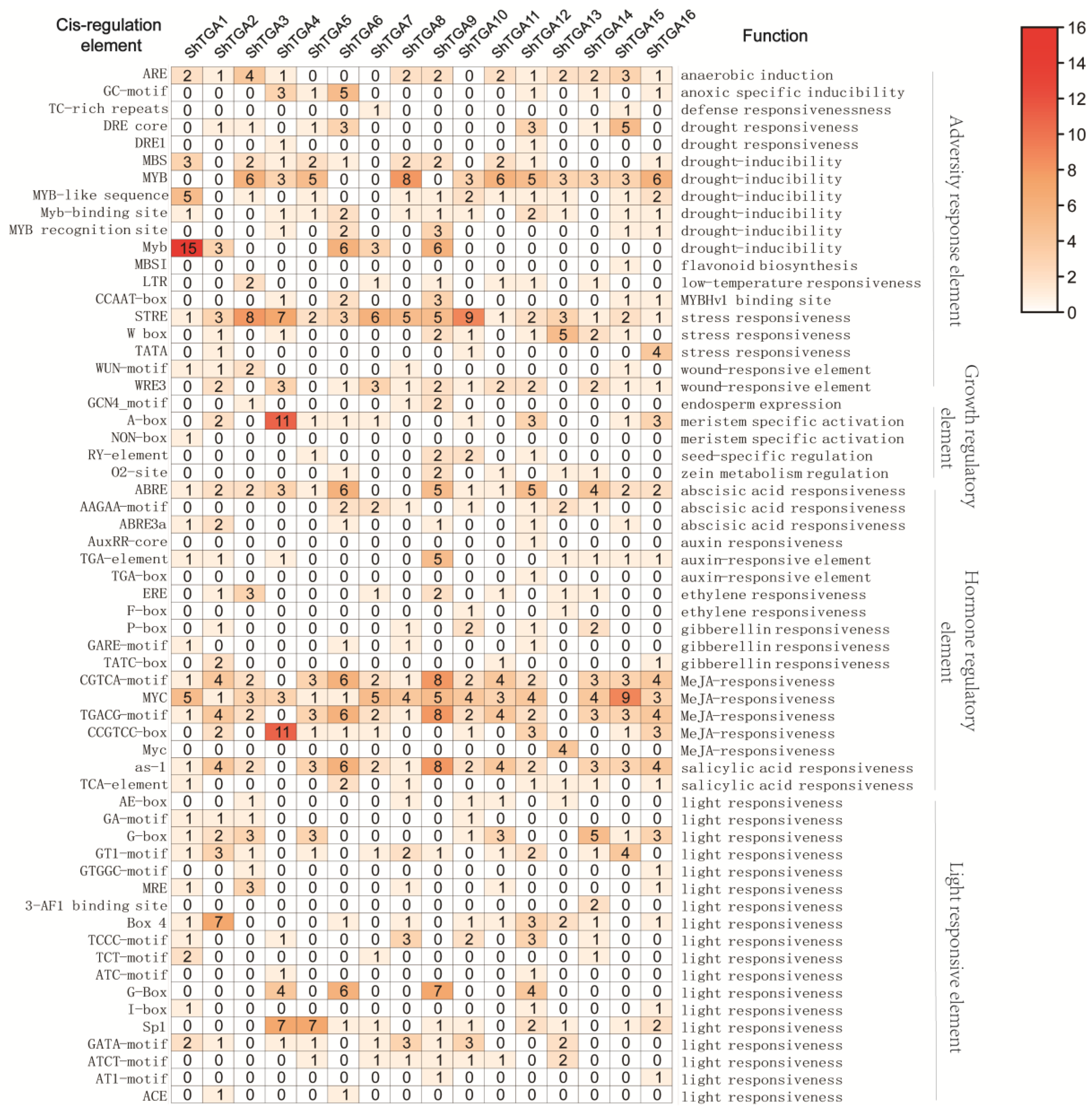
3.4. Cis-Elements Analysis in ShTGAs Promoter Regions
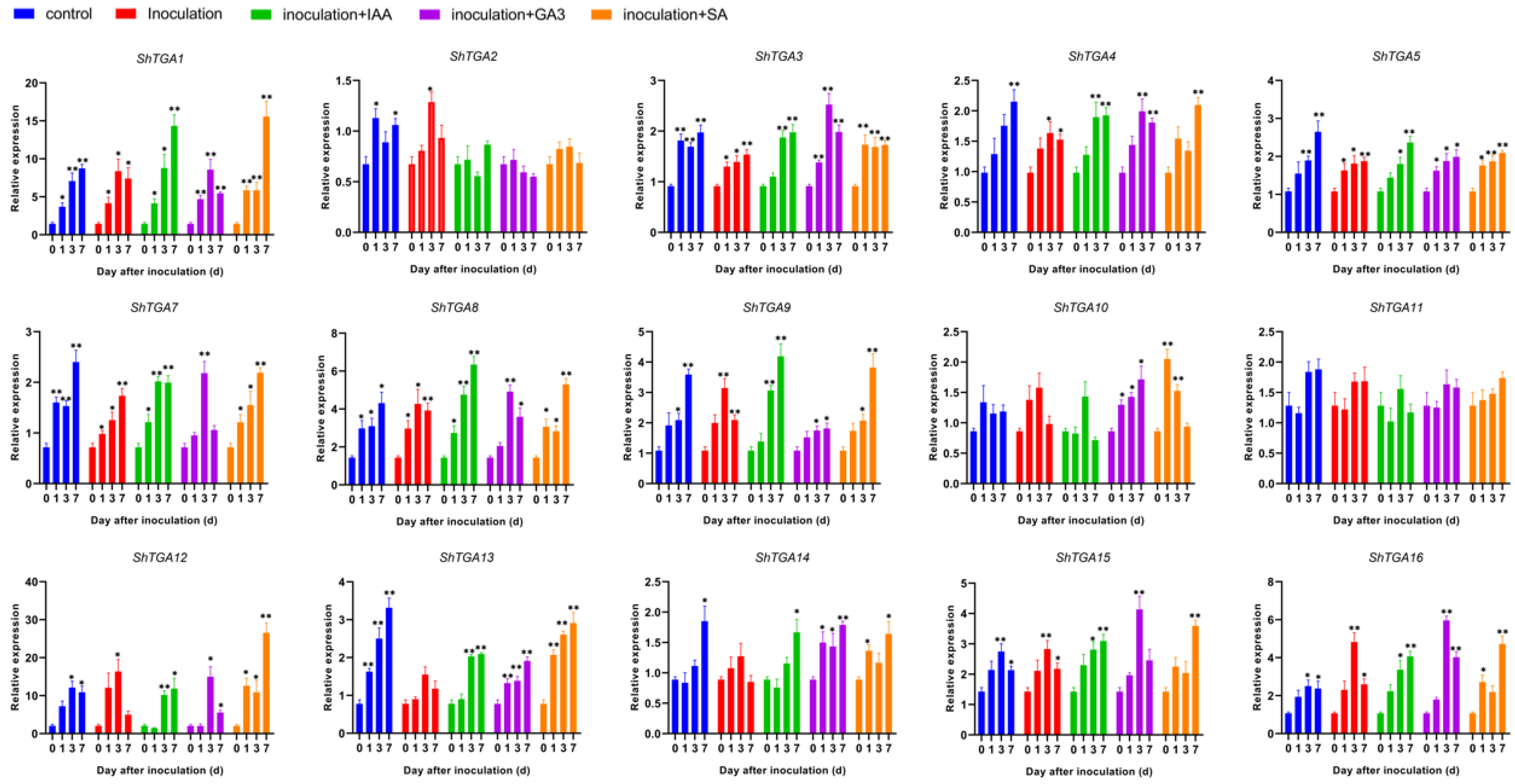
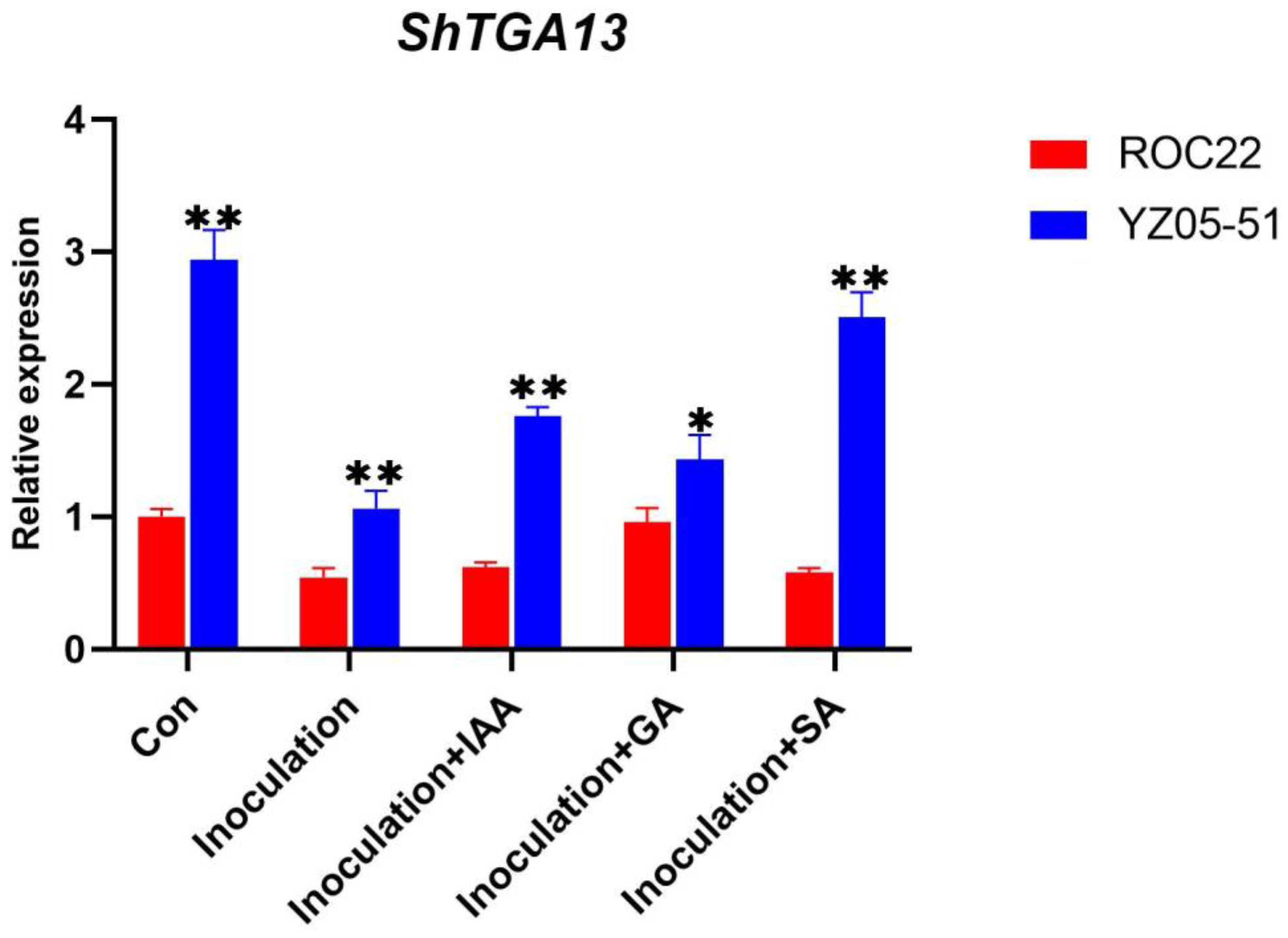
3.5. Expression Profile Analysis of ShTGA Genes in the Response to S. scitamineum Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izawa, T.; Foster, R.; Chua, N.H. Plant bZIP protein DNA binding specificity. J. Mol. Biol. 1993, 230, 1131–1144. [Google Scholar] [CrossRef]
- Schindler, U.; Beckmann, H.; Cashmore, A.R. TGA1 and G-box binding factors: Two distinct classes of Arabidopsis leucine zipper proteins compete for the G-box-like element TGACGTGG. Plant Cell 1992, 4, 1309–1319. [Google Scholar] [PubMed]
- Katagiri, F.; Lam, E.; Chua, N.H. Two tobacco DNA-binding proteins with homology to the nuclear factor CREB. Nature 1989, 340, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, B.; Droge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Li, C.H.; Wang, K.T.; Lei, C.Y.; Xu, F.; Ji, N.N.; Jiang, Y.B. Identification of TGA gene family in peach and analysis of expression mode involved in a BABA-induced disease resistance. Acta Hortic. Sin. 2022, 49, 265–280. [Google Scholar]
- Lin, P.; Wang, M.Y.; Li, Y.Q.; Liu, J.F.; Zhang, H.Y.; Lin, S.D. Identification of banana TGA transcription factor family and the expression analysis under Fusarium wilt infection. Chin. J. Trop. Crops 2021, 42, 2134–2142. [Google Scholar]
- Jin, X.F. Identification of Tga Transcription Factor and Its Functional Analysis in The Regulation of Taxol Synthesis in Taxus chinensis. Master’s Thesis, Huazhong University of Science and Technology, Wuhan, China, 2020. [Google Scholar]
- Tian, M. Genome-Wide Characterization of the TGA Gene Family in Melon and Their Roles in Disease Resistance. Master’s Thesis, Zhejiang University, Hangzhou, China, 2021. [Google Scholar]
- Ullah, I.; Magdy, M.; Wang, L.X.; Liu, M.Y.; Li, X. Genome-wide identification and evolutionary analysis of TGA transcription factors in soybean. Sci. Rep. 2019, 9, 11186. [Google Scholar] [CrossRef]
- Loake, G.; Grant, M. Salicylic acid in plant defence-the players and protagonists. Curr. Opin. Plant Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef]
- Kesarwani, M.; Yoo, J.; Dong, X.N. Genetic interactions of TGA transcription factors in the regulation of pathogenesis-related genes and disease resistance in Arabidopsis. Plant Physiol. 2007, 144, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Tessaro, M.J.; Lassner, M.; Li, X. Knockout analysis of Arabidopsis transcription factors TGA2, TGA5 and TGA6 reveals their redundant and essential roles in systemic acquired resistance. Plant Cell 2003, 15, 2647–2653. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.J.; Busta, L.; Zhang, Q.; Ding, P.T.; Jetter, R.; Zhang, Y.L. TGACG-BINDING FACTOR 1 (TGA1) and TGA4 regulate salicylic acid and pipecolic acid biosynthesis by modulating the expression of SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1 (SARD1) and CALMODULINBINDING PROTEIN 60g (CBP60g). New Phytol. 2018, 217, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Zander, M.; La Camera, S.; Lamotte, O.; Métraux, J.P.; Gatz, C. Arabidopsis thaliana class-II TGA transcription factors are essential activators of jasmonic acid/ethylene-induced defense responses. Plant J. 2010, 61, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Zander, M.; Thurow, C.; Gatz, C. TGA transcription factors activate the salicylic acid-suppressible branch of the ethylene-induced defense program by regulating ORA59 expression. Plant Physiol. 2014, 165, 1671. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.; Hilbert, B.; Dueckershoff, K.; Roitsch, T.; Krischke, M.; Mueller, M.J.; Berger, S. General detoxification and stress responses are mediated by oxidized lipids through TGA transcription factors in Arabidopsis. Plant Cell 2008, 20, 768–785. [Google Scholar] [CrossRef]
- Herrera-Vásquez, A.; Fonseca, A.; Ugalde, J.M.; Lamig, L.; Seguel, A.; Moyano, T.C.; Gutiérrez, R.A.; Salinas, P.; Vidal, E.A.; Holuigue, L. TGA class II transcription factors are essential to restrict oxidative stress in response to UV-B stress in Arabidopsis. J. Exp. Bot. 2021, 72, 1891–1905. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.H.; Liu, Z.Q.; Long, Y.P.; Liang, Y.L.; Jin, Z.P.; Zhang, L.P.; Liu, D.M.; Li, H.; Zhai, J.X.; Pei, Y.X. The Ca2+/calmodulin2-binding transcription factor TGA3 elevates LCD expression and H2S production to bolster Cr6+ tolerance in Arabidopsis. Plant J. 2017, 91, 1038–1050. [Google Scholar] [CrossRef]
- Zhong, L.; Chen, D.D.; Min, D.H.; Li, W.W.; Xu, Z.S.; Zhou, Y.B.; Li, L.C.; Chen, M.; Ma, Y.Z. AtTGA4, a bZIP transcription factor, confers drought resistance by enhancing nitrate transport and assimilation in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 457, 433–439. [Google Scholar] [CrossRef]
- Hou, J.Y.; Sun, Q.; Li, J.J.; Ahammed, G.J.; Yu, J.Q.; Fang, H.; Xia, X.J. Glutaredoxin S25 and its interacting TGACG motif-binding factor TGA2 mediate brassinosteroid-induced chlorothalonil metabolism in tomato plants. Environ. Pollut. 2019, 255, 113256. [Google Scholar] [CrossRef]
- Chen, L.J.; Zou, W.S.; Fei, C.Y.; Wu, G.; Li, X.Y.; Lin, H.H.; Xi, D.H. α-Expansin EXPA4 positively regulates abiotic stress tolerance but negatively regulates pathogen resistance in Nicotiana tabacum. Plant Cell Physiol. 2018, 59, 2317–2330. [Google Scholar] [CrossRef]
- Wang, Y.; Salasini, B.C.; Khan, M.; Devi, B.; Bush, M.; Subramaniam, R.; Hepworth, S.R. Clade I TGA bZIP transcription factors mediate BLADE-ON-PETIOLE-dependent regulation of development. Plant Physiol. 2019, 180, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.Y.; Wi, S.J.; Park, K.Y. Functional switching of NPR1 between chloroplast and nucleus for adaptive response to salt stress. Sci. Rep. 2020, 10, 4339. [Google Scholar] [CrossRef]
- Kumar, S.; Zavaliev, R.; Wu, Q.L.; Zhou, Y.; Cheng, J.; Dillard, L.; Powers, J.; Withers, J.; Zhao, J.S.; Guan, Z.Q.; et al. Structural basis of NPR1 in activating plant immunity. Nature 2022, 605, 561–566. [Google Scholar] [CrossRef]
- Chai, L.X.; Dong, K.; Liu, S.Y.; Zhang, Z.; Zhang, X.P.; Tong, X.; Zhu, F.F.; Zou, J.Z.; Wang, X.B. A putative nuclear copper chaperone promotes plant immunity in Arabidopsis. J. Exp. Bot. 2020, 71, 6684–6696. [Google Scholar] [CrossRef] [PubMed]
- Maier, A.T.; Stehling-Sun, S.; Offenburger, S.-L.; Lohmann, J.U. The bZIP transcription factor PERIANTHIA: A multifunctional hub for meristem control. Front. Plant Sci. 2011, 2, 79. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.R.; Xu, J.Y.; Yuan, C.; Hu, Y.K.; Liu, Q.G.; Chen, Q.Q.; Zhang, P.C.; Shi, N.N.; Qin, C. Characterization of genes associated with TGA7 during the floral transition. BMC Plant Biol. 2021, 21, 367. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, W.; Karapetyan, S.; Mwimba, M.; Marqués, J.; Buchler, N.E.; Dong, X.N. Redox rhythm reinforces the circadian clock to gate immune response. Nature 2015, 523, 472–476. [Google Scholar] [CrossRef]
- Hu, X.C.; Yang, L.Y.; Ren, M.F.; Liu, L.; Fu, J.; Cui, H.C. TGA factors promote plant root growth by modulating redox homeostasis or response. J. Integr. Plant Biol. 2022, 8, 1543–1559. [Google Scholar] [CrossRef]
- Bhuiyan, S.A.; Magarey, R.C.; McNeil, M.D.; Aitken, K.S. Sugarcane smut, caused by Sporisorium scitamineum, a major disease of sugarcane: A contemporary review. Phytopathology 2021, 111, 1905–1917. [Google Scholar] [CrossRef]
- Schaker, P.D.C.; Palhares, A.C.; Taniguti, L.M.; Peters, L.P.; Creste, S.; Aitken, K.S.; Van Sluys, M.A.; Kitajima, J.P.; Vieira, M.L.; Monteiro-Vitorello, C.B. RNA-Seq transcriptional profiling following whip development in sugarcane smut disease. PLoS ONE 2016, 11, e0162237. [Google Scholar] [CrossRef]
- Que, Y.X.; Guo, J.L.; Wu, Q.B.; Xu, L.P. A global view of transcriptome dynamics during Sporisorium scitamineum challenge in sugarcane by RNA-seq. PLoS ONE 2014, 9, e106476. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.C.; Xu, L.P.; Wang, Z.Q.; Peng, Q.; Yang, Y.T.; Chen, Y.; Que, Y.X. Comparative proteomics reveals that central metabolism changes are associated with resistance against Sporisorium scitamineum in sugarcane. BMC Genom. 2016, 17, 800. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.A.E.; Oirdi, M.E.; Gonzalez-Lamothe, R.; Bouarab, K. Necrotrophic pathogens use the salicylic acid signaling pathway to promote disease development in tomato. Mol. Plant-Microbe Interact. 2012, 25, 1584–1593. [Google Scholar] [CrossRef]
- Su, Y.C.; Xu, L.P.; Xue, B.T.; Wu, Q.B.; Guo, J.L.; Wu, L.J.; Que, Y.X. Molecular cloning and characterization of two pathogenesis-related β-1, 3-glucanase genes ScGluA1 and ScGluD1 from sugarcane infected by Sporisorium scitamineum. Plant Cell Rep. 2013, 32, 1503–1519. [Google Scholar] [CrossRef]
- Chen, J.W.; Kuang, J.F.; Peng, G.; Wan, S.B.; Liu, R.; Yang, Z.D.; Deng, H.H. Molecular cloning and expression analysis of a NPR1 gene from sugarcane. Pak. J. Bot. 2012, 44, 193–200. [Google Scholar]
- Huang, N.; Ling, H.; Su, Y.C.; Liu, F.; Xu, L.P.; Su, W.H.; Wu, Q.B.; Guo, J.L.; Gao, S.W.; Que, Y.X. Transcriptional analysis identifies major pathways as response components to Sporisorium scitamineum stress in sugarcane. Gene 2018, 678, 207–218. [Google Scholar] [CrossRef]
- Tang, N.; Ma, S.Q.; Zong, W.; Yang, N.; Lv, Y.; Yan, C.; Guo, Z.L.; Li, J.; Li, X.; Xiang, Y.; et al. MODD mediates deactivation and degradation of OsbZIP46 to negatively regulate ABA signaling and drought resistance in rice. Plant Cell 2016, 28, 2161–2177. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.N.; Xue, L.J.; Zou, M.J.; Liu, J.Y.; Chen, F.; Xue, H.W. Rice ABI5-Like1 regulates abscisic acid and auxin responses by affecting the expression of ABRE-containing genes. Plant Physiol. 2011, 156, 1397–1409. [Google Scholar] [CrossRef]
- Tomaž, Š.; Gruden, K.; Coll, A. TGA transcription factors—Structural characteristics as basis for functional variability. Front. Plant Sci. 2022, 13, 935819. [Google Scholar] [CrossRef]
- Gatz, C. From pioneers to team players: TGA transcription factors provide a molecular link between different stress pathways. Mol. Plant Microbe Interact. 2013, 26, 151–159. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Li, Q.; Li, Z.M.; Staswick, P.E.; Wang, M.Y.; Zhu, Y.; He, Z.H. Dual regulation role of GH3.5 in salicylic acid and auxin signaling during Arabidopsis-Pseudomonas syringae interaction. Plant Physiol. 2007, 145, 450–464. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.F.; Liu, J.Y.; Yang, K.; Xia, H.M.; Wu, C.W.; Chen, X.K.; Zhao, J.; Yang, H.C.; Li, J.; Zan, F.G. Registration of ‘YZ0551’ sugarcane. J. Plant Regist. 2015, 9, 172–178. [Google Scholar] [CrossRef]
- Almeida, C.M.A.; Donato, V.M.T.; Amaral, D.O.J.; Lima, G.S.A.; Brito, G.G.; Lima, M.M.A.; Correia, M.T.S.; Silva, M.V. Differential gene expression in sugarcane induced by salicylic acid and under water deficit conditions. Agric. Sci. Res. J. 2014, 3, 38–44. [Google Scholar]
- Szőke, L.; Moloi, M.J.; Kovács, G.E.; Biró, G.; Radócz, L.; Hájos, M.T.; Kovács, B.; Rácz, D.; Danter, M.; Tóth, B. The Application of Phytohormones as Biostimulants in Corn Smut Infected Hungarian Sweet and Fodder Corn Hybrids. Plants 2021, 10, 1822. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.Z.; Zhou, S.G.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, 5–9. [Google Scholar] [CrossRef]
- Lu, S.N.; Wang, J.Y.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, 265–268. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, 427–432. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 1987, 13, 1402–1406. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Schiermeyer, A.; Thurow, C.; Gatz, C. Tobacco bZIP factor TGA10 is a novel member of the TGA family of transcription factors. Plant Mol. Biol. 2003, 51, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Noshi, M.; Mori, D.; Tanabe, N.; Maruta, T.; Shigeoka, S. Arabidopsis clade IV TGA transcription factors, TGA10 and TGA9, are involved in ROS mediated responses to bacterial PAMP flg22. Plant Sci. 2016, 252, 12–21. [Google Scholar] [CrossRef]
- Venturuzzi, A.L.; Rodriguez, M.C.; Conti, G.; Leone, M.; Caro, M.D.P.; Montecchia, J.F.; Zavallo, D.; Asurmendi, S. Negative modulation of SA signaling components by the capsid protein of tobacco mosaic virus is required for viral long-distance movement. Plant J. 2021, 106, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.H.; Lin, X.H.; Zuo, Y.; Yu, Z.; Baerson, S.R.; Pan, Z.Q.; Zeng, R.S.; Song, Y.Y. Transcription factor OsbZIP49 controls tiller angle and plant architecture through the induction of indole-3-acetic acid-amido synthetases in rice. Plant J. 2021, 108, 1346–1364. [Google Scholar] [CrossRef] [PubMed]
- Glickmann, E.; Gardan, L.; Jacquet, S.; Hussain, S.; Elasri, M.; Petit, A.; Dessaux, Y. Auxin production is a common feature of most pathovars of Pseudomonas syringae. Mol. Plant-Microbe Interact. 1998, 11, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Jagadeeswaran, G.; Raina, S.; Acharya, B.R.; Maqbool, S.B.; Mosher, S.L.; Appel, H.M.; Schultz, J.C.; Klessig, D.F.; Raina, R. Arabidopsis GH3-LIKE DEFENSE GENE 1 is required for accumulation of salicylic acid, activation of defense responses and resistance to Pseudomonas syringae. Plant J. 2007, 51, 234–246. [Google Scholar] [CrossRef]
- Park, J.E.; Park, J.Y.; Kim, Y.S.; Staswick, P.E.; Jeon, J.; Yun, J.; Kim, S.Y.; Kim, J.; Lee, Y.H.; Park, C.M. GH3-mediated auxin homeostasis links growth regulation with stress adaptation response in Arabidopsis. J. Biol. Chem. 2007, 282, 10036–10046. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pajerowska-Mukhtar, K.; Culler, A.H.; Dong, X.N. Salicylic acid inhibits pathogen growth in plants through repression of the auxin signaling pathway. Curr. Biol. 2007, 17, 1784–1790. [Google Scholar] [CrossRef]
- Ding, X.H.; Cao, Y.L.; Huang, L.L.; Zhao, J.; Xu, C.G.; Li, X.H.; Wang, S.P. Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice. Plant Cell 2008, 20, 228–240. [Google Scholar] [CrossRef]
- Domingo, C.; Andrés, F.; Tharreau, D.; Iglesias, D.J.; Talón, M. Constitutive expression of OsGH3.1 reduces auxin content and enhances defense response and resistance to a fungal pathogen in rice. Mol. Plant-Microbe Interact. 2009, 22, 201–210. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Accession Number | Number of Amino Acid | MW | pI | BZIP Domain Location | DOG1 Domain Location | GRAVY | Instability Index | Aliphatic Index | Subcellular Location |
|---|---|---|---|---|---|---|---|---|---|---|
| ShTGA1 | OP205433 | 407 | 45,278.52 | 6.35 | 115–159 | 186–400 | −0.399 | 48.79 | 77.79 | Nuclear |
| ShTGA2 | OP205434 | 375 | 41,336.72 | 6.06 | 84–128 | 151–372 | −0.319 | 51.74 | 80.21 | Nuclear |
| ShTGA3 | OP205435 | 335 | 37,385.27 | 8.92 | 49–93 | 116–332 | −0.533 | 55.77 | 82.54 | Nuclear |
| ShTGA4 | OP205436 | 333 | 37,116.98 | 8.9 | 47–91 | 114–330 | −0.522 | 55.89 | 83.63 | Nuclear |
| ShTGA5 | OP205437 | 330 | 36,652.12 | 7.08 | 44–88 | 111–327 | −0.593 | 58.34 | 79.64 | Nuclear |
| ShTGA6 | OP205438 | 332 | 36,931.58 | 6.68 | 45–89 | 112–329 | −0.560 | 57.99 | 84.52 | Nuclear |
| ShTGA7 | OP205439 | 463 | 50,666.80 | 6.23 | 176–220 | 243–460 | −0.498 | 55.12 | 78.55 | Nuclear |
| ShTGA8 | OP205440 | 485 | 53,170.47 | 6.1 | 179–223 | 243–456 | −0.460 | 56.59 | 72 | Nuclear |
| ShTGA9 | OP205441 | 431 | 47,270.06 | 6.81 | 125–169 | 189–402 | −0.472 | 48.01 | 76.91 | Nuclear |
| ShTGA10 | OP205442 | 527 | 57,470.71 | 6.29 | 216–260 | 281–495 | −0.374 | 58.98 | 77.61 | Nuclear |
| ShTGA11 | OP205443 | 514 | 56,653.49 | 5.91 | 198–240 | 263–477 | −0.460 | 65.57 | 73.04 | Nuclear |
| ShTGA12 | OP205444 | 496 | 55,227.21 | 6.88 | 190–232 | 270–484 | −0.565 | 68.92 | 75.02 | Nuclear |
| ShTGA13 | OP205445 | 306 | 33,014.46 | 9.67 | 145–188 | 207–306 | −0.503 | 50.70 | 72.91 | Nuclear |
| ShTGA14 | OP205446 | 425 | 46,972.51 | 7.87 | 138–181 | 205–415 | −0.393 | 61.96 | 80.64 | Nuclear |
| ShTGA15 | OP205447 | 567 | 60,463.29 | 5.86 | 113–173 | / | −0.382 | 51.12 | 73.69 | Nuclear |
| ShTGA16 | OP205448 | 652 | 68,705.2 | 8.83 | 177–237 | / | −0.449 | 45.38 | 71.27 | Nuclear |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Z.; Hu, X.; Wu, Z.; Liu, X.; Wu, C.; Zeng, Q. Identification and Expression Profiling of TGA Transcription Factor Genes in Sugarcane Reveals the Roles in Response to Sporisorium scitamineum Infection. Agriculture 2022, 12, 1644. https://doi.org/10.3390/agriculture12101644
Luo Z, Hu X, Wu Z, Liu X, Wu C, Zeng Q. Identification and Expression Profiling of TGA Transcription Factor Genes in Sugarcane Reveals the Roles in Response to Sporisorium scitamineum Infection. Agriculture. 2022; 12(10):1644. https://doi.org/10.3390/agriculture12101644
Chicago/Turabian StyleLuo, Zhengying, Xin Hu, Zhuandi Wu, Xinlong Liu, Caiwen Wu, and Qianchun Zeng. 2022. "Identification and Expression Profiling of TGA Transcription Factor Genes in Sugarcane Reveals the Roles in Response to Sporisorium scitamineum Infection" Agriculture 12, no. 10: 1644. https://doi.org/10.3390/agriculture12101644
APA StyleLuo, Z., Hu, X., Wu, Z., Liu, X., Wu, C., & Zeng, Q. (2022). Identification and Expression Profiling of TGA Transcription Factor Genes in Sugarcane Reveals the Roles in Response to Sporisorium scitamineum Infection. Agriculture, 12(10), 1644. https://doi.org/10.3390/agriculture12101644