
Sex and Body Colour Affect the Variation in Internal Body Temperature of Oedaleus decorus asiaticus in Natural Habitats in Inner Mongolia, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Measurements
2.3. Statistical Analyses
3. Results
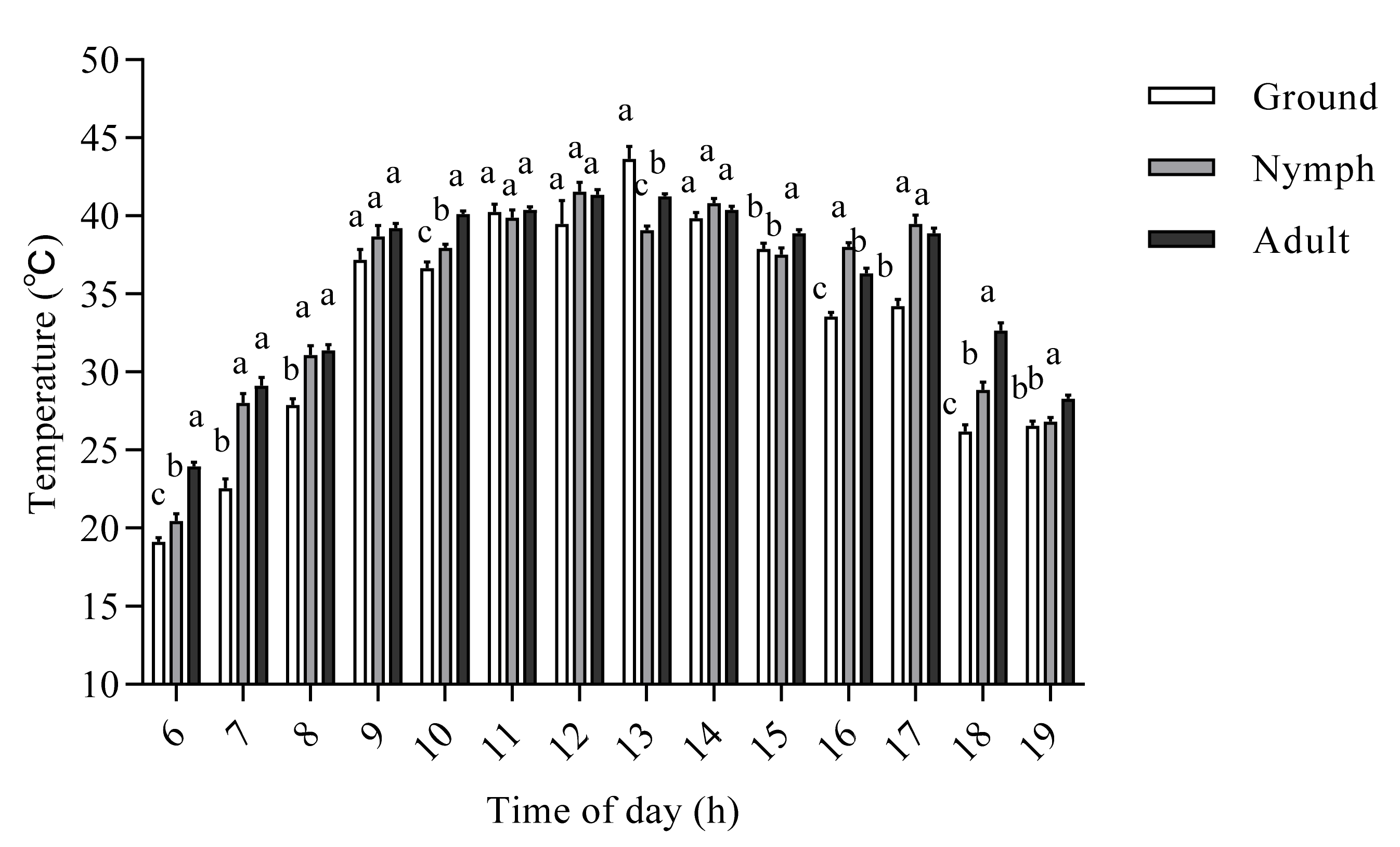
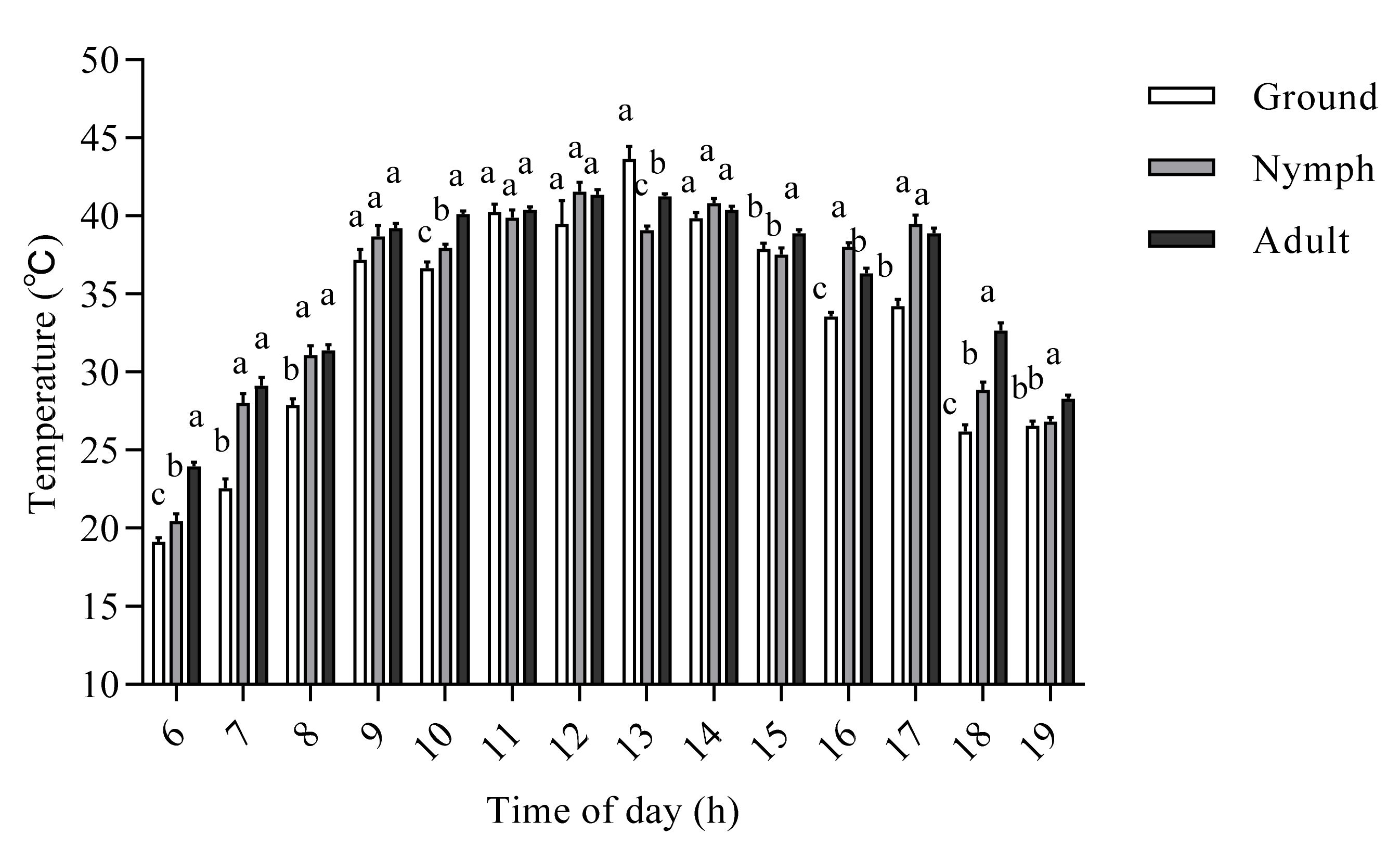
3.1. The Effect of Developmental Stage on the Internal Body Temperature of O. d. asiaticus
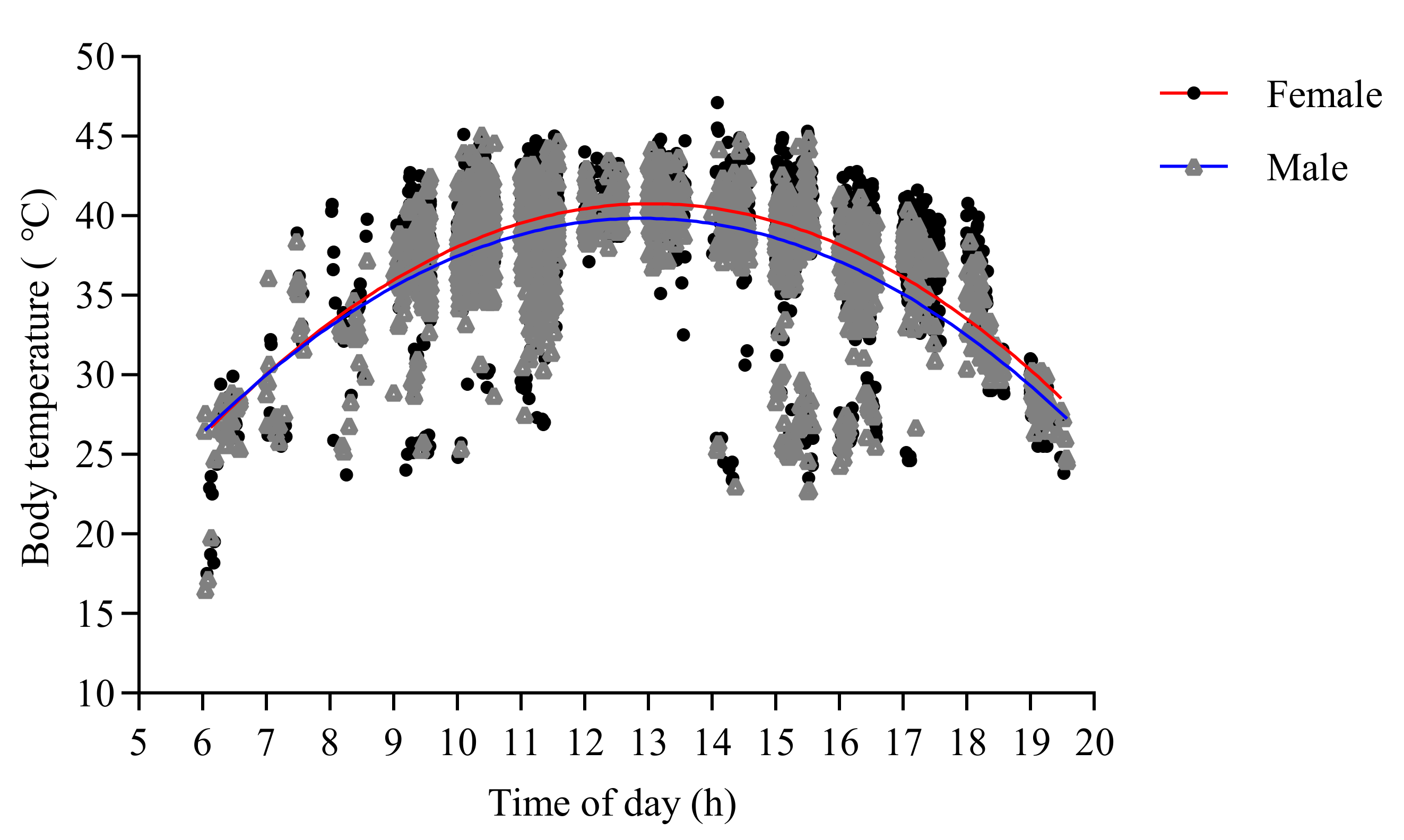
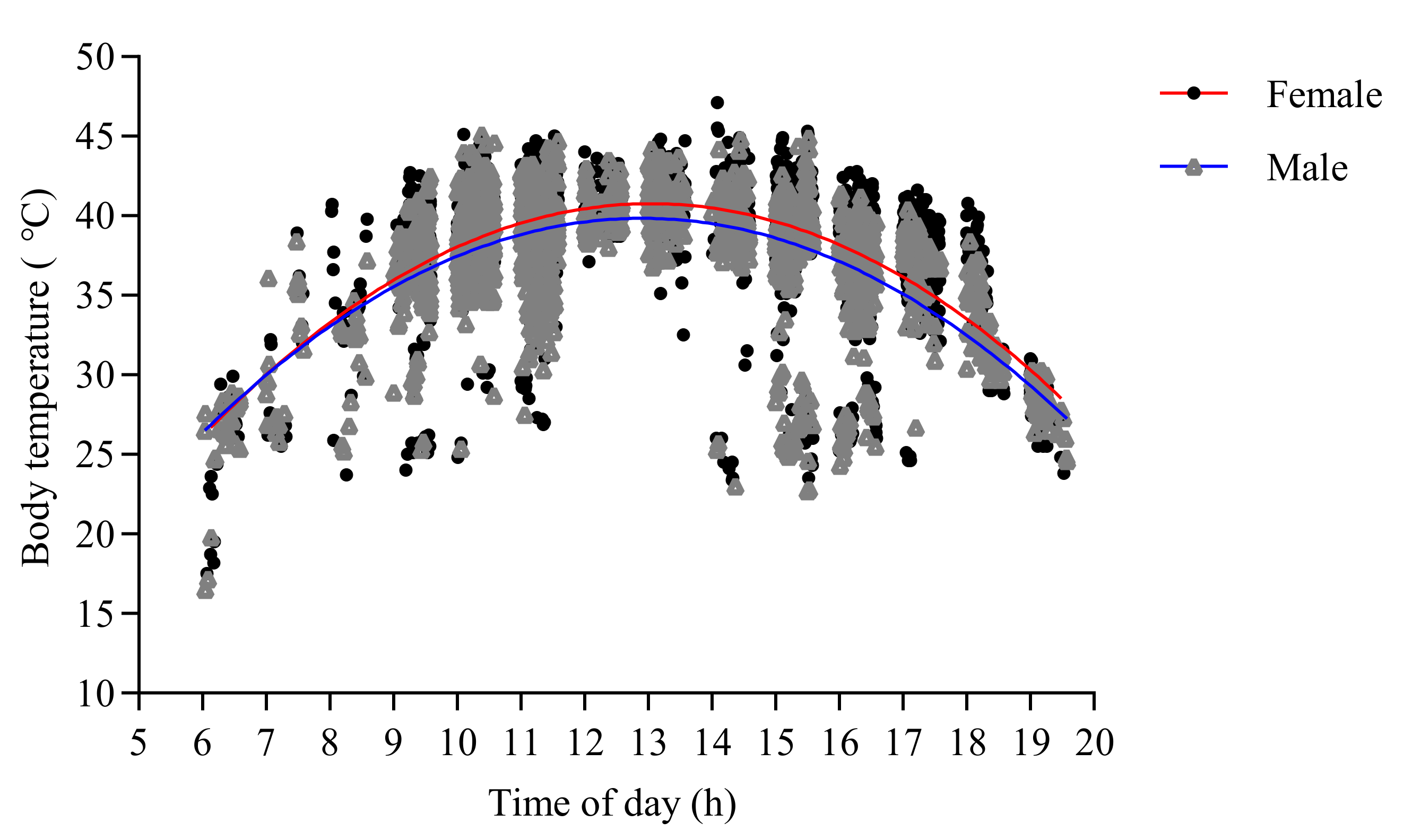
3.2. Factors Affecting the Internal Body Temperature of O. d. asiaticus Adults
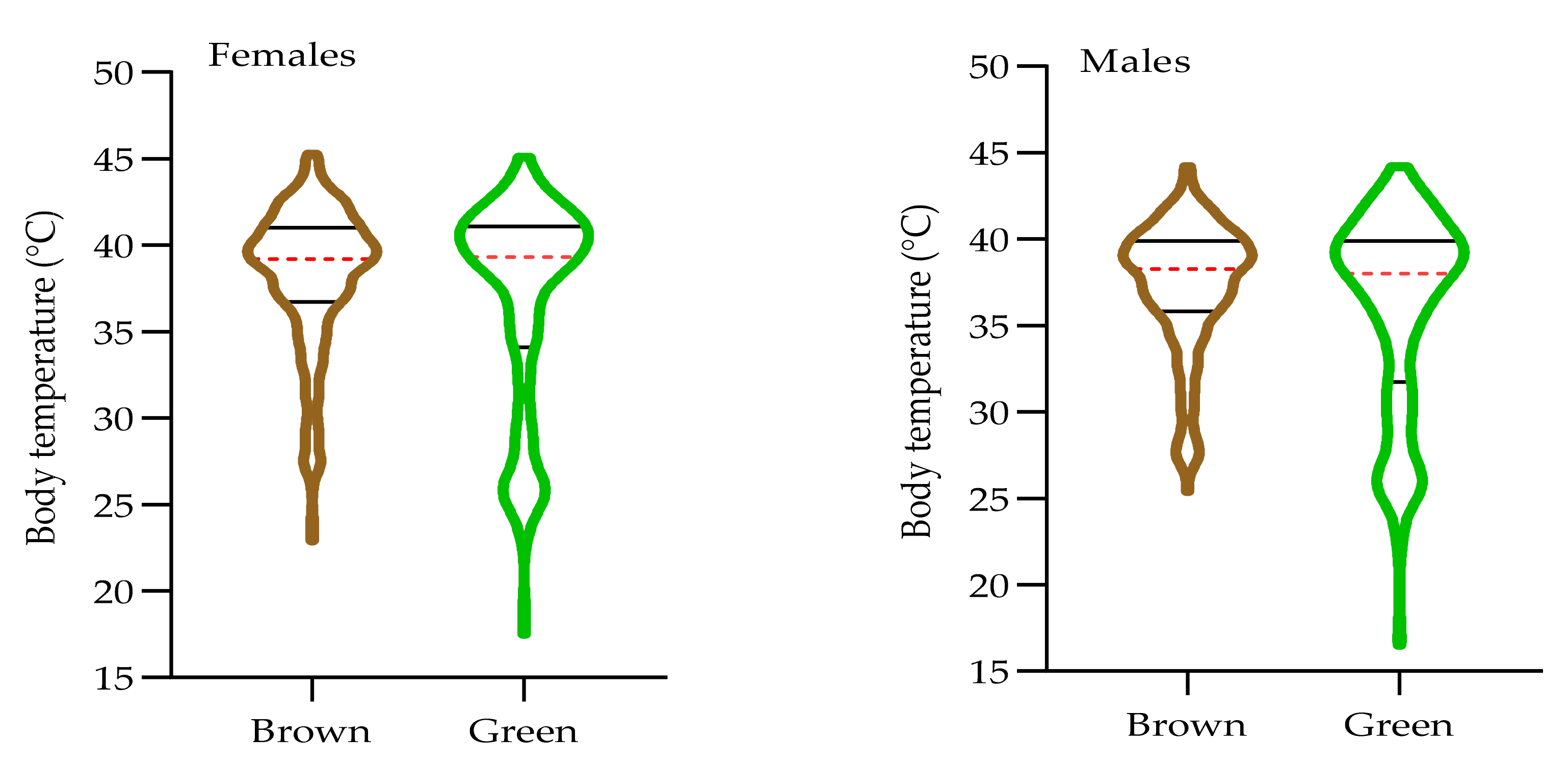
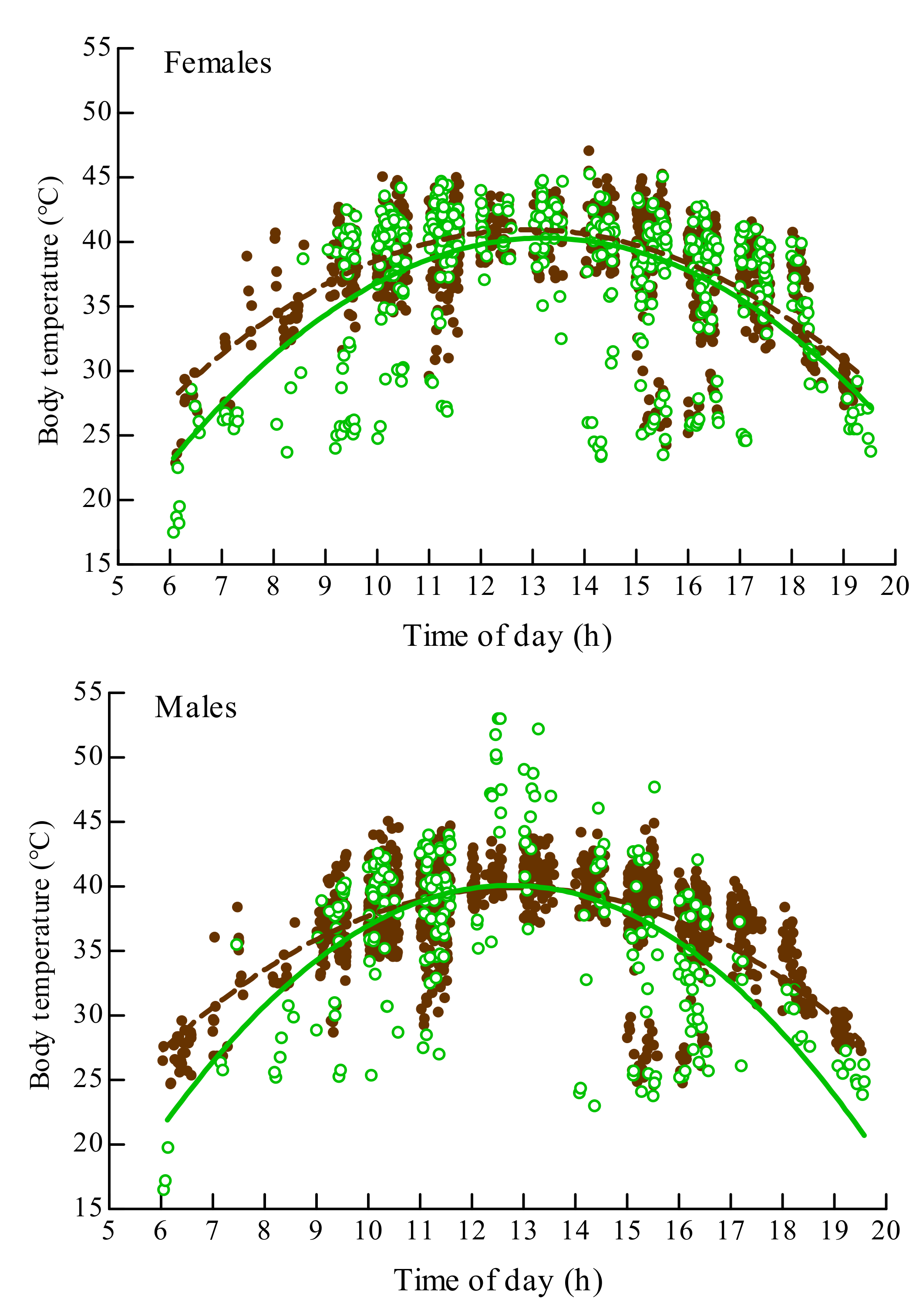
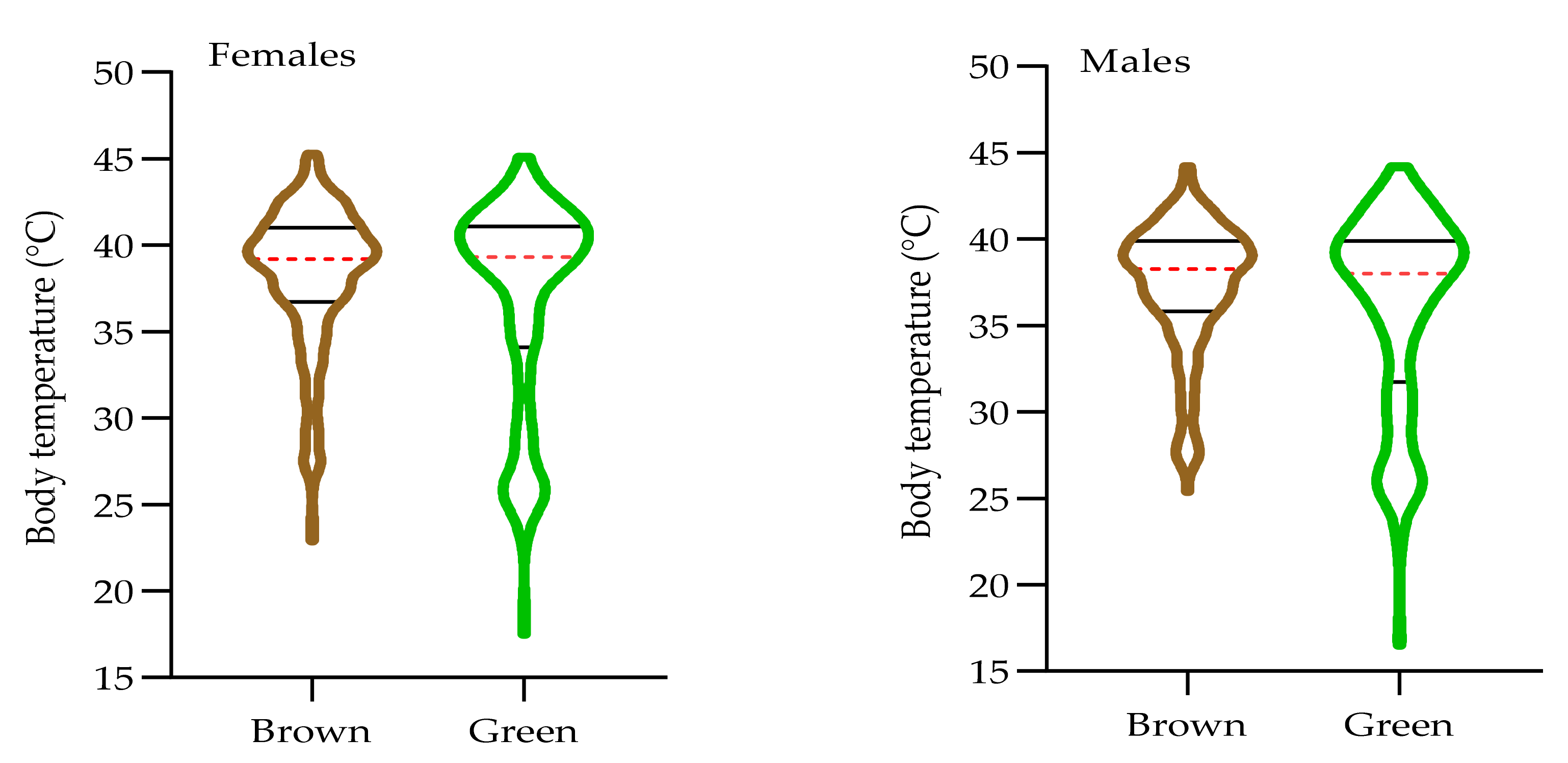
3.3. The Effect of Morph on Internal Body Temperature of O. d. asiaticus Female and Male Adults
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gall, M.L.; Overson, R.; Cease, A. A global review on locusts (Orthoptera: Acrididae) and their interactions with livestock grazing practices. Front. Ecol. Evol. 2019, 7, 263. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and grasshopper management. Annu. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Gao, S.; Zhang, G.Y.; Wang, Y.; Yang, B.D.; Zhang, Z.R.; Zheng, S.Y.; Wang, M.L. Using Metarhizium flavoviride oil spray to control grasshoppers in Inner Mongolia grassland. Chin. J. Biol. Control 2000, 16, 49–52. [Google Scholar]
- Hong, J.; Zhao, H.L.; Wang, G.J. The occurring and control situation of grasshopper in the grassland of China (review). Acta Agrestia Sin. 2014, 22, 929–934. [Google Scholar]
- Ritchie, J.M. A taxonomic revision of the genus Oedaleus Fieber (Orthoptera: Acrididae). Bull. Br. Mus. Entomol. 1981, 42, 83–183. [Google Scholar]
- Lomer, C.J.; Bateman, R.P.; Johnson, D.L.; Langewald, J.; Thomas, M. Biological control of locusts and grasshoppers. Annu. Rev. Entomol. 2001, 46, 667–702. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.Z.; Meng, Z.P.; Guo, Y.H.; Shen, J.P. Occurrences and control of Oedaleus decorus asiaticus in agro-pastoral intersecting area of Ulanqab. Inn. Mong. Agric. Sci. Technol. 2005, 26, 384–387. [Google Scholar]
- Gao, S.J.; Wei, Y.S.; Te, M.R.; Liu, A.P.; Xu, L.B.; Wang, N. The flight ability of Oedaleus asiaticus and its relationship to population density. Pratacult. Sci. 2012, 29, 1915–1919. [Google Scholar]
- Blanford, S.; Thomas, M.B. Host thermal biology: The key to understanding host-pathogen interactions and microbial pest control? Agr. For. Entomol. 1999, 1, 195–202. [Google Scholar] [CrossRef]
- Maeno, K.O.; Piou, C.; Kearney, M.R.; Ely, S.O.; Ebbe, M. A general model of the thermal constraints on the world’s most destructive locust, Schistocerca gregaria. Ecol. Appl. 2021, 31, e02310. [Google Scholar] [CrossRef]
- Hunt, V.L.; Charnley, A.K. The inhibitory effect of the fungal toxin, destruxin A, on behavioural fever in the desert locust. J. Insect Physiol. 2011, 57, 1341–1346. [Google Scholar] [CrossRef]
- Willott, S.J. The Thermal Ecology and population. Dynamics of Grasshoppers in Relation to Grazing on a Breckland Grass Heath. Ph.D. Thesis, University of East Anglia, Norwich, UK, 1992; pp. 333–337. [Google Scholar]
- Huang, G.H.; Long, Q.C. The change of body temperature of the oriental migratory locust (Locusta migratoria manilensis Meyen) during flight. Acta Entomol. Sin. 1962, 11, 419–421. [Google Scholar]
- Hu, L.J.; Du, W.G. Thermoregulation and thermal dependence of locomotor performance in the gecko Gekko japonicus. Acta Zool. Sin. 2007, 53, 227–232. [Google Scholar]
- Thomas, M.B.; Blanford, S. Thermal biology in insect-parasite interactions. Trends Ecol. Evol. 2003, 18, 344–350. [Google Scholar] [CrossRef]
- Digby, P.S.B. Factors affecting the temperature excess of insects in sunshine. J. Exp. Biol. 1955, 32, 279–298. [Google Scholar] [CrossRef]
- Liu, Y.M.; Yang, J.W.; Fan, C.B.; Shang, S.Q.; Bryony, T.; Li, H.M. Research on environmental factors regulating body temperature of oriental migratory locust Locusta migratoria manilensis. J. Plant Protec. 2018, 45, 1296–1301. [Google Scholar]
- Watt, W.B. Adaptive significance of pigment polymorphismsin Colias butterflies I. Variation of melanin pigmentin relation to thermoregulation. Evolution 1968, 22, 437–458. [Google Scholar]
- Jacobs, M.D.; Watt, W.B. Seasonal adaptation vsphysiological constraint: Photoperiod, thermoregulationand flight in Colias butterflies. Funct. Ecol. 1994, 8, 366–376. [Google Scholar] [CrossRef]
- Partridge, L.; French, V. Thermal Evolution of Ectotherm Body Size: Why Get Big in the Cold? Cambridge University Press: Cambridge, UK, 1996; pp. 265–292. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Scaling: Why Is Body Size So Important; Cambridge University Press: Cambridge, UK, 1994; pp. 56–73. [Google Scholar]
- Shepherd, B.L.; Prange, H.D.; Moczek, A.P. Some like it hot: Body and weapon size affect thermoregulation in horned beetles. J. Insect Physiol. 2008, 54, 604–611. [Google Scholar] [CrossRef]
- Chown, S.L.; Nicolson, S. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, UK, 2004; pp. 225–226. [Google Scholar]
- Nielsen, M.E.; Papaj, D.R. Effects of developmental change in body size on ectotherm body temperature and behavioural thermoregulation: Caterpillars in a heat-stressed environment. Oecologia 2015, 177, 171–179. [Google Scholar] [CrossRef]
- Woods, H.A. Ontogenetic changes in the body temperature of an insect herbivore. Funct. Ecol. 2013, 27, 1322–1331. [Google Scholar] [CrossRef] [Green Version]
- Michael, J.A.; Todd, D.S.; Michael, W.S. Temperature, growth rate, and body size in ectotherms: Fitting pieces of a life-history puzzle. Integr. Comp. Biol. 2004, 44, 498–509. [Google Scholar]
- Zuo, W.; Moses, M.E.; West, G.B.; Hou, C.; Brown, J.H. A general model for effects of temperature on ectotherm ontogenetic growth and development. Proc. Biol. Sci. 2012, 279, 1840–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, P.; Cramp, R.L.; Franklin, C.E. Physiological responses of ectotherms to daily temperature variation. J. Exp. Biol. 2015, 218, 3068–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, B.J.; Marshall, K.E.; Sewell, M.A.; Levesque, D.L.; Willett, C.S.; Slotsbo, S.; Dong, Y.; Harley, C.D.; Marshall, D.J.; Helmuth, B.S.; et al. Can we predict ectotherm responses to climate change using thermal performance curves and body temperatures? Ecol. Lett. 2016, 19, 1372–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubalcaba, J.G.; Gouveia, S.; Olalla-Tárraga, M.A. Upscaling microclimatic conditions into body temperature distributions of ectotherms. Am. Nat. 2019, 193, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Köhler, G.; Schielzeth, H. Green-brown polymorphism in alpine grasshoppers affects body temperature. Ecol. Evol. 2019, 10, 441–450. [Google Scholar] [CrossRef]
- Veron, J.E.N. The role of physiological colour change in the thermoregulation of Austrolestes annulosus (Selys) (Odonata). Aust. J. Zool. 1974, 22, 457–469. [Google Scholar] [CrossRef]
- Umbers, K.D.L. Turn the temperature to turquoise: Cues for colour change in the male chameleon grasshopper (Kosciuscola tristis) (Orthoptera: Acrididae). J. Insect Physiol. 2011, 57, 1198–1204. [Google Scholar] [CrossRef]
- Seebacher, F.; White, C.R.; Franklin, C.E. Physiological plasticity increases resilience of ectothermic animals to climate change. Nat. Clim. Change 2015, 5, 61–66. [Google Scholar] [CrossRef]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism: An update. Adv. Insect Physiol. 2009, 36, 1–286. [Google Scholar]
- Ayali, A. The puzzle of locust density-dependent phasepolyphenism. Curr. Opin. Insect Sci. 2019, 35, 41–47. [Google Scholar] [CrossRef]
- Blanford, S.; Thomas, M.B.; Langewald, J. Behavioural fever in the Senegalese grasshopper, Oedaleus senegalensis, and its implications for biological control using pathogens. Ecol. Entomol. 1998, 23, 9–14. [Google Scholar] [CrossRef]
- Schielzeth, H. Simple means to improve the interpretability of regression coefficients. Methods Ecol. Evol. 2010, 1, 103–113. [Google Scholar] [CrossRef]
- Rskov, C.K.; Tregenza, T.; Overgaard, J. Using radiotelemetry to study behavioural thermoregulation in insects under field conditions. Methods Ecol. Evol. 2019, 10, 1773–1782. [Google Scholar] [CrossRef]
- Clusella-Trullas, S.; Nielsen, M. The evolution of insect body coloration under changing climates. Curr. Opin. Insect Sci. 2020, 41, 25–32. [Google Scholar] [CrossRef]
- Bartholomew, G.A.; Heinrich, B. Endothermy in African dung beetles during flight, ball making, and ball rolling. J. Exp. Biol. 1978, 73, 65–83. [Google Scholar] [CrossRef]
- Zeuss, D.; Brandl, R.; Brändle, M.; Rahbek, C.; Brunzel, S. Global warming favours light-coloured insects in Europe. Nat. Commun. 2014, 5, 3874. [Google Scholar] [CrossRef]
- Franco, A.M.A.; Hill, J.K.; Kitschke, C.; Collingham, Y.C.; Roy, D.B.; Fox, R.; Huntley, B.; Thomas, C.D. Impacts of climate warming and habitat loss on extinctions at species’ low-latitude range boundaries. Glob. Change Biol. 2006, 12, 1545–1553. [Google Scholar] [CrossRef]
- Pateman, R.M.; Hill, J.K.; Roy, D.B.; Fox, R.; Thomas, C.D. Temperature-dependent alterations in host use drive rapid range expansion in a butterfly. Science 2012, 336, 1028–1030. [Google Scholar] [CrossRef]
- Buxton, P.A. Heat, moisture and animal life in deserts. Proc. R. Soc. B Biol. Sci. 1924, 96, 123–131. [Google Scholar]
- Schilthuizen, M.; Kellermann, V. Contemporary climate change and terrestrial invertebrates: Evolutionary versus plastic changes. Evol. Appl. 2014, 7, 56–67. [Google Scholar] [CrossRef]
- MacLean, H.J.; Nielsen, M.E.; Kingsolver, J.G.; Buckley, B. Using museum specimens to track morphological shifts through climate change. Philos. Trans. R. Soc. B Biol. Sci. 2018, 374, 20170404. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.K.; Thomas, C.D.; Fox, R.; Telfer, M.G.; Willis, S.G.; Asher, J.; Huntley, B. Responses of butterflies to twentieth century climate warming: Implications for future ranges. Proc. R. Soc. B Biol. Sci. 2002, 269, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography. Divers. Distrib. 2010, 16, 321–333. [Google Scholar] [CrossRef]
- Kerr, J.T.; Kharouba, H.M.; Currie, D.J. The macroecological contribution to global change solutions. Science 2007, 316, 1581–1584. [Google Scholar] [CrossRef] [Green Version]
- Dillon, M.E.; Wang, G.; Huey, R.B. Global metabolic impacts of recent climate warming. Nature 2010, 467, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Lisansky, S. Microbial biopesticides. In Microbial Insecticides: Novel or Necessity? Proceedings No. 68. British Crop Protection Council; Evans, H.F., Ed.; Springer: Farnham, UK, 1997; pp. 3–10. [Google Scholar]
- Blanford, S.; Thomas, M.B. Role of thermal biology in disease dynamics. Asp. Appl. Biol. 1999, 53, 73–82. [Google Scholar]
- Blanford, S.; Thomas, M.B. Thermal Behavior of Two Acridid Species: Effects of Habitat and Season on Body Temperature and the Potential Impact on Biocontrol with Pathogens. Environ. Entomol. 2000, 29, 1060–1069. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment Locations | Longitude (E) | Latitude (N) | Altitude (m) |
|---|---|---|---|
| Scientific Observation and Experimental Station of Pests in Xilingol Rangeland, MARA | 116.001 | 43.950 | 984 ± 3 |
| The serpentine bay tourist area | 116.114 | 43.821 | 1066 ± 3 |
| Abag banner (40–50 km) | 115.380 | 43.877 | 1159 ± 3 |
| Shagouzi | 116.334 | 43.827 | 1167 ± 3 |
| Xilingol League reservoir | 116.089 | 43.848 | 1026 ± 3 |
| West Ujimqin banner 1 | 116.808 | 44.482 | 1023 ± 3 |
| Xilingol city | 116.284 | 44.057 | 1092 ± 3 |
| West Ujimqin banner 2 | 116.483 | 44.285 | 1040 ± 3 |
| Sex | Morphs | Fit Curves | R2 | df | F | p |
|---|---|---|---|---|---|---|
| Female | Brown | Tb = –0.58 + 7.12 t − 0.28 t2 | 0.41 | 1122 | 361.93 | 0.00 |
| Green | Tb = –17.90 + 8.80 t − 0.33 t2 | 0.31 | 470 | 101.03 | 0.00 | |
| Male | Brown | Tb = –4.70 + 6.93 t − 0.27 t2 | 0.45 | 1281 | 545.00 | 0.00 |
| Green | Tb = –27.34 + 10.58 t − 0.42 t2 | 0.39 | 245 | 77.76 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.; Li, H.; Liu, L.; Wang, G.; Gu, H.; Luke, B. Sex and Body Colour Affect the Variation in Internal Body Temperature of Oedaleus decorus asiaticus in Natural Habitats in Inner Mongolia, China. Agriculture 2022, 12, 878. https://doi.org/10.3390/agriculture12060878
Cheng Y, Li H, Liu L, Wang G, Gu H, Luke B. Sex and Body Colour Affect the Variation in Internal Body Temperature of Oedaleus decorus asiaticus in Natural Habitats in Inner Mongolia, China. Agriculture. 2022; 12(6):878. https://doi.org/10.3390/agriculture12060878
Chicago/Turabian StyleCheng, Yumeng, Hongmei Li, Lulu Liu, Guangjun Wang, Haojing Gu, and Belinda Luke. 2022. "Sex and Body Colour Affect the Variation in Internal Body Temperature of Oedaleus decorus asiaticus in Natural Habitats in Inner Mongolia, China" Agriculture 12, no. 6: 878. https://doi.org/10.3390/agriculture12060878
APA StyleCheng, Y., Li, H., Liu, L., Wang, G., Gu, H., & Luke, B. (2022). Sex and Body Colour Affect the Variation in Internal Body Temperature of Oedaleus decorus asiaticus in Natural Habitats in Inner Mongolia, China. Agriculture, 12(6), 878. https://doi.org/10.3390/agriculture12060878