Rootstock Effects on Anthocyanin Accumulation and Associated Biosynthetic Gene Expression and Enzyme Activity during Fruit Development and Ripening of Blood Oranges


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sampling
2.2. Chemicals and Solvents
2.3. Fruit Weight, Shape, and Color Measurement
2.4. Determination of Soluble Solid Content (SSC), Total Soluble Sugar (TSS), Titratable Acidity (TA), Vitamin C (Vc), and Total Anthocyanin Concentration (TAC)
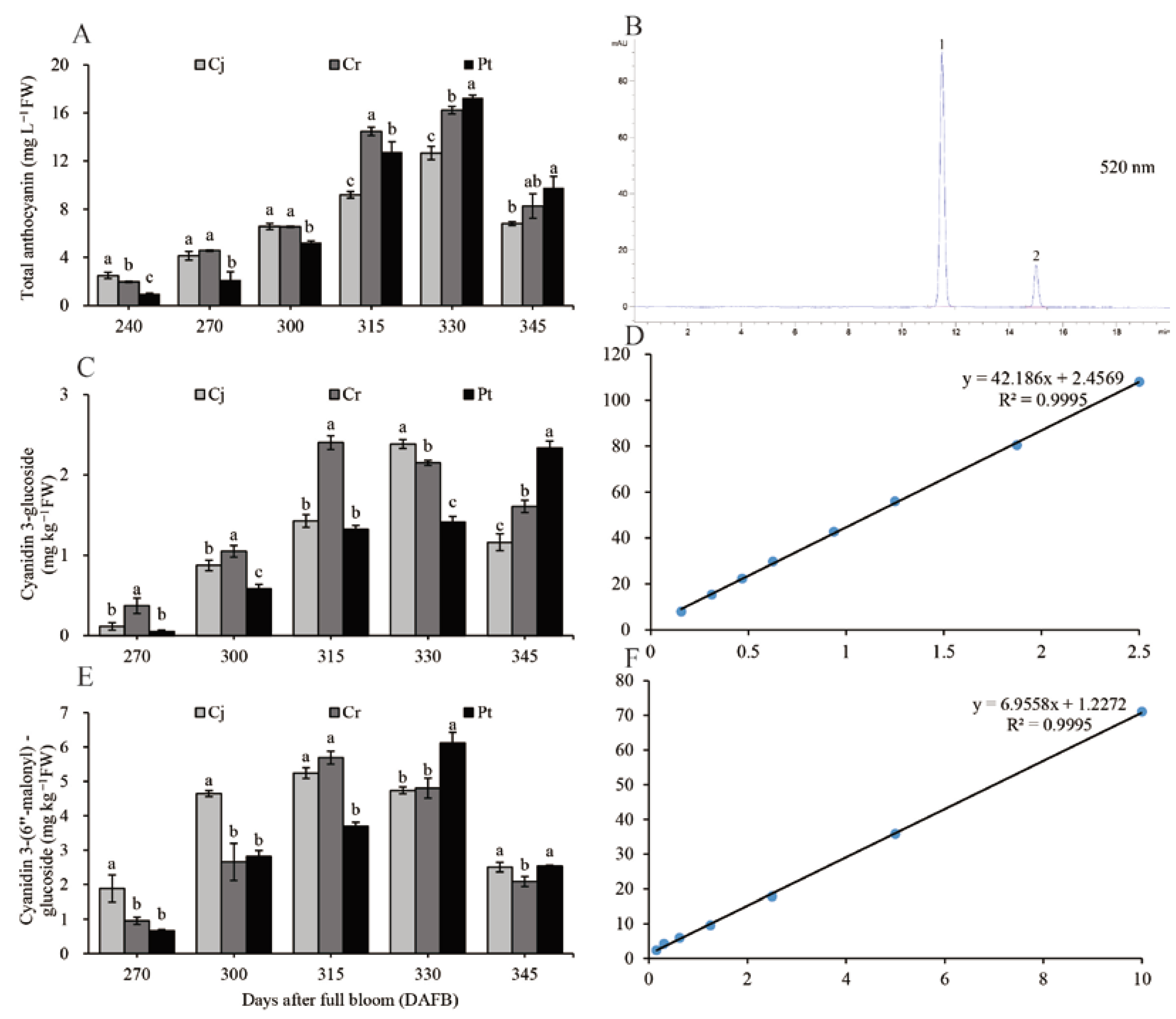
2.5. Extraction, Characterization, and Quantification of Anthocyanin Using High-Performance Liquid Chromatography (HPLC)
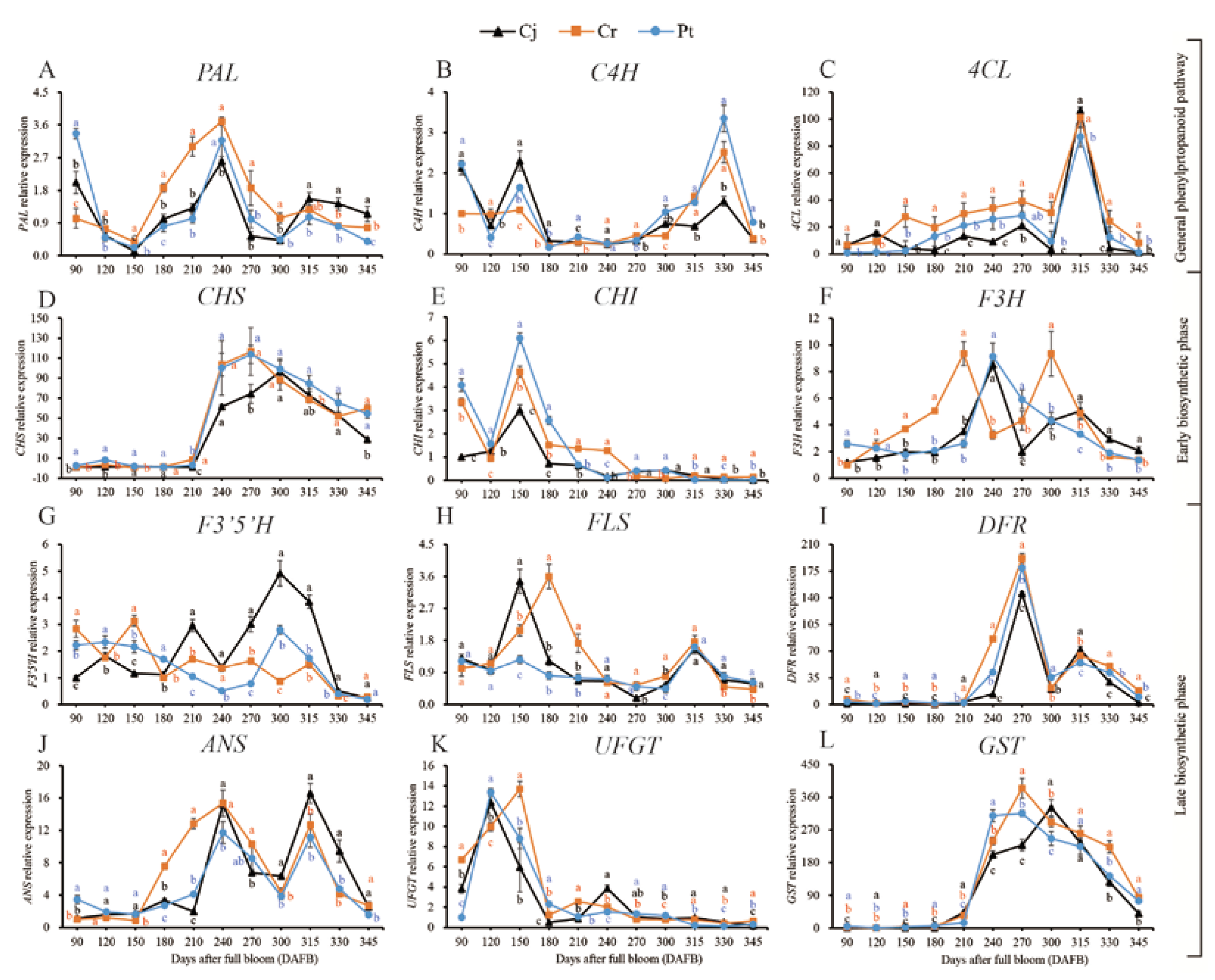
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
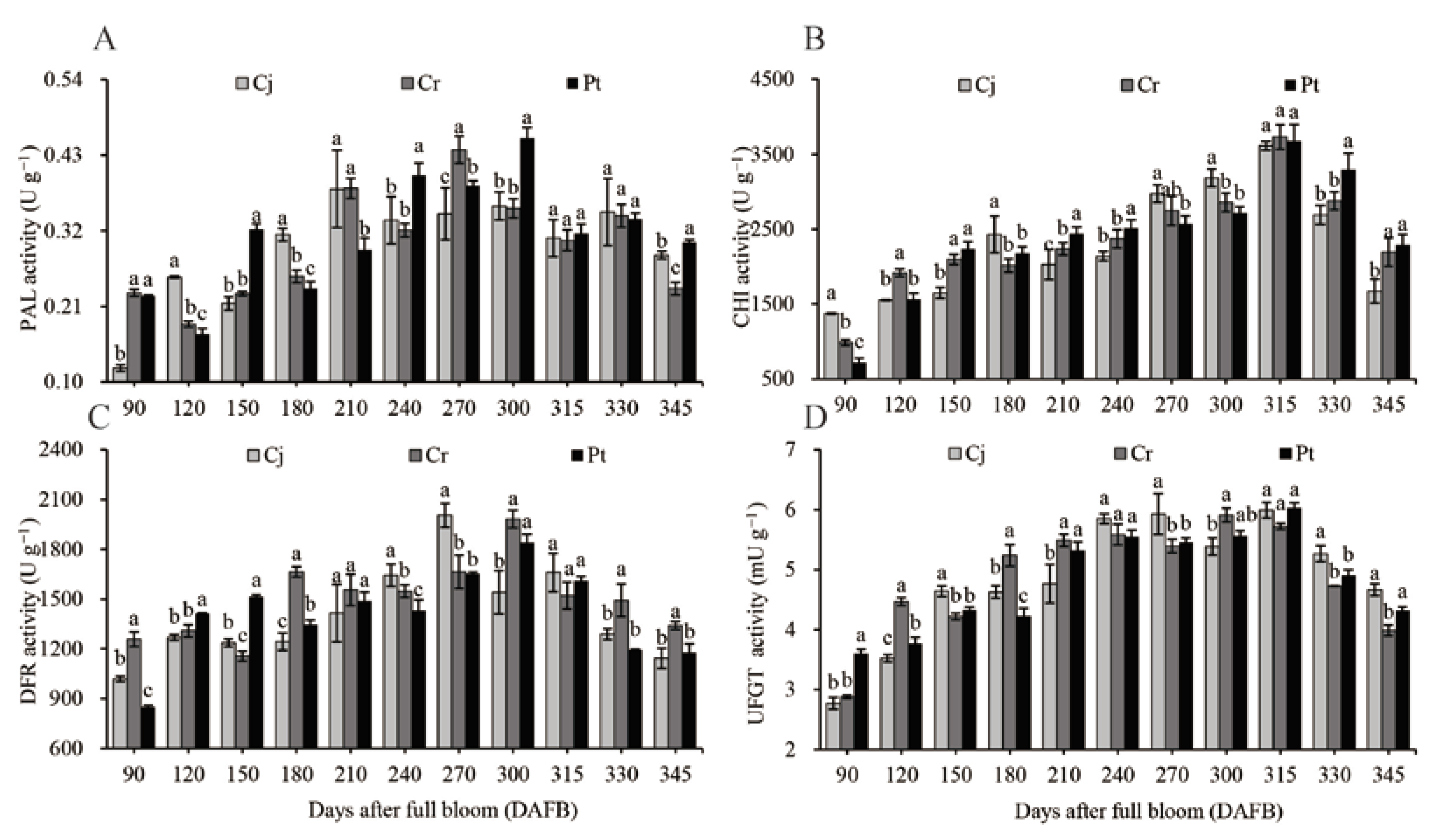
2.7. Assay Enzymes
2.8. Statistical Analyses
3. Results
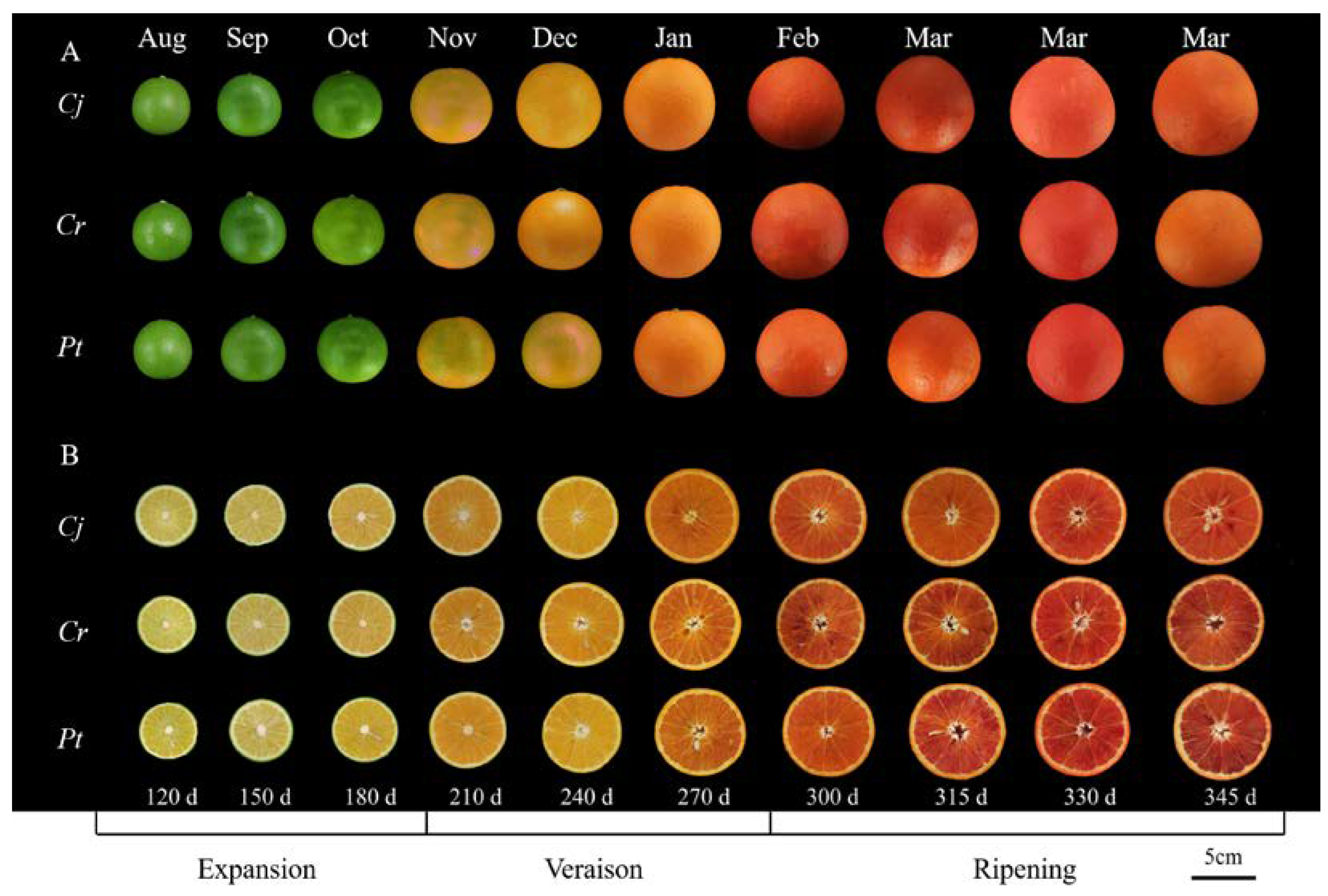
3.1. The Fruit Phenotypes
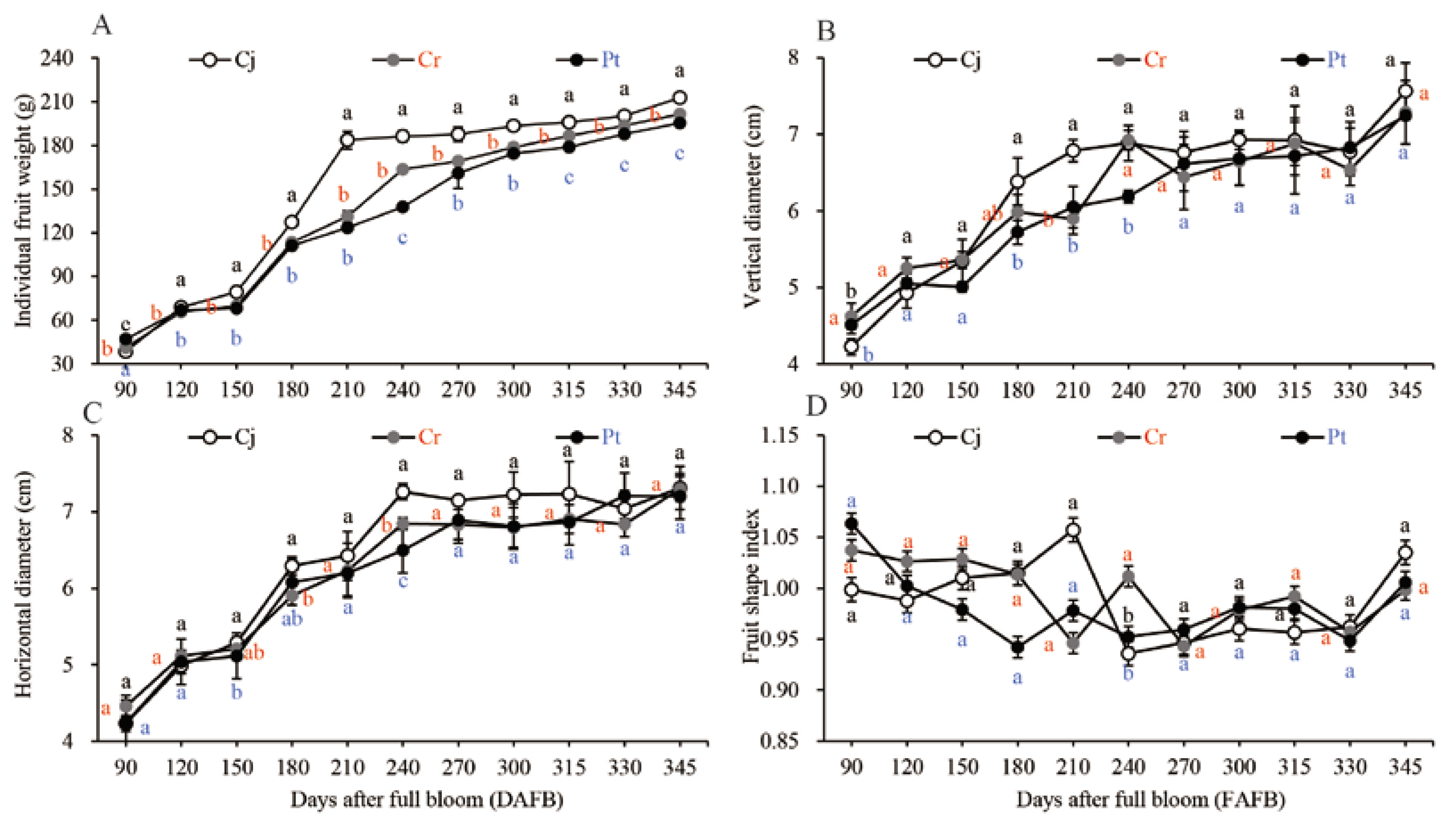
3.2. The General Features of the Fruits
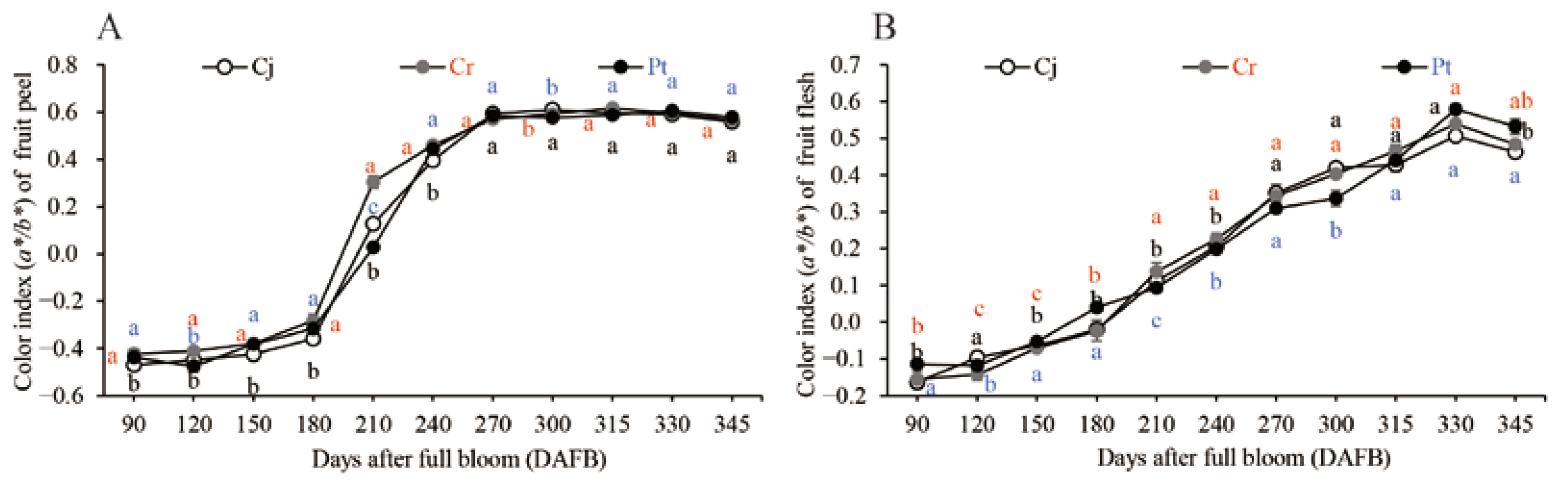
3.3. The Color Index of the Fruits
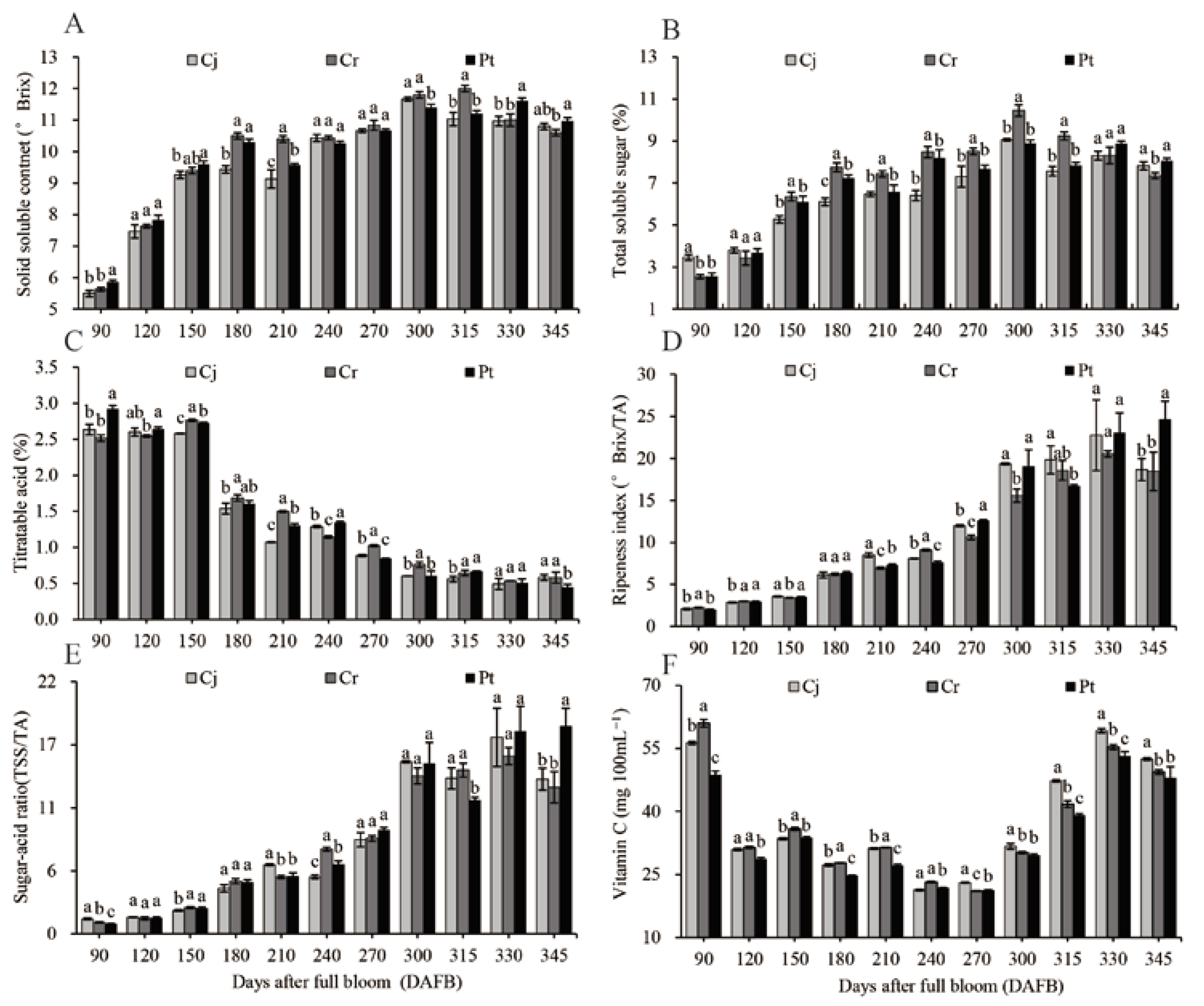
3.4. The SSC, TSS, TA, and Vc Analysis
3.5. The Anthocyanin Concentration
3.6. Activity of Enzyme for Anthocyanin Biosynthesis
3.7. Expression of Genes for Anthocyanin Biosynthetic
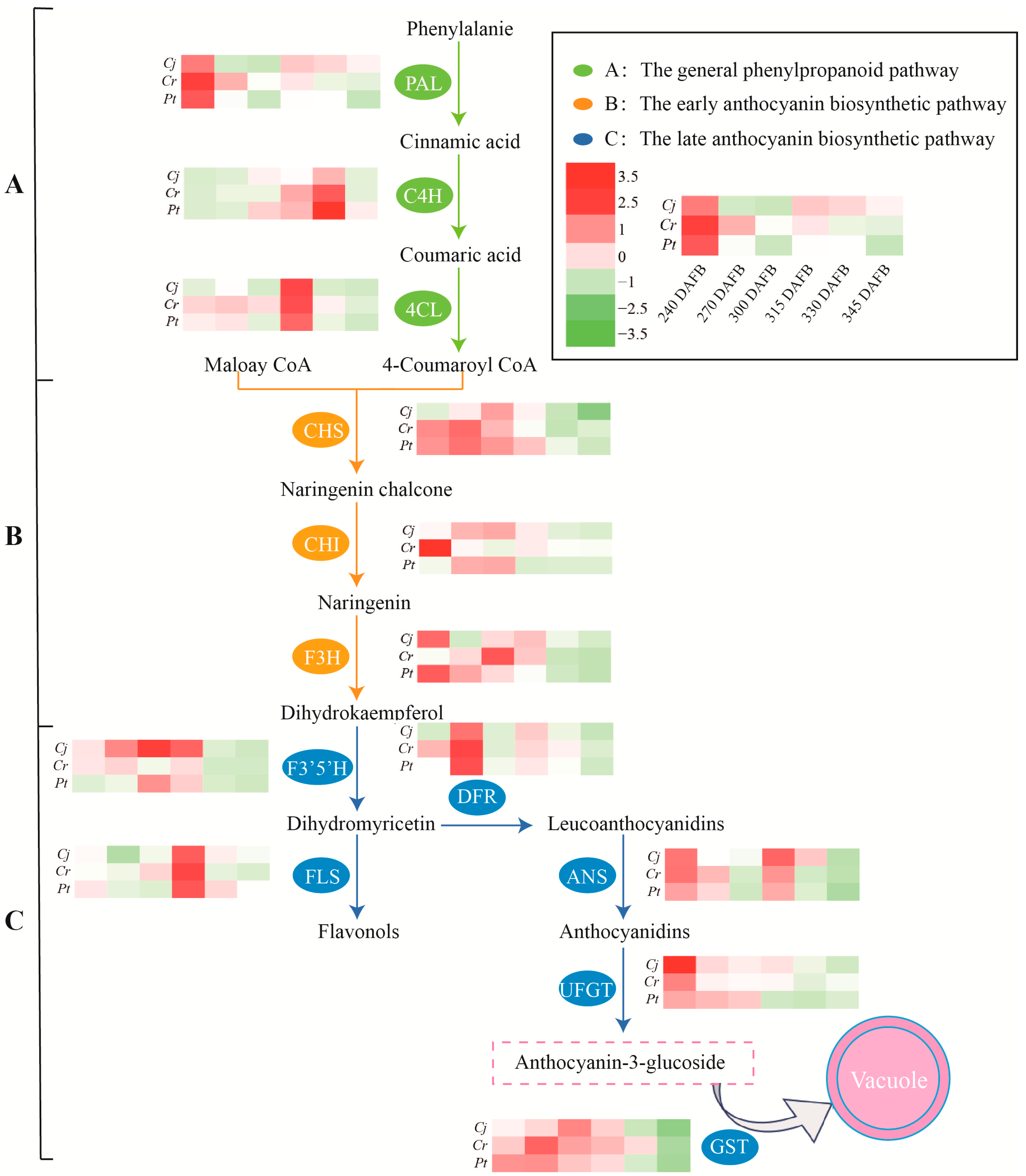
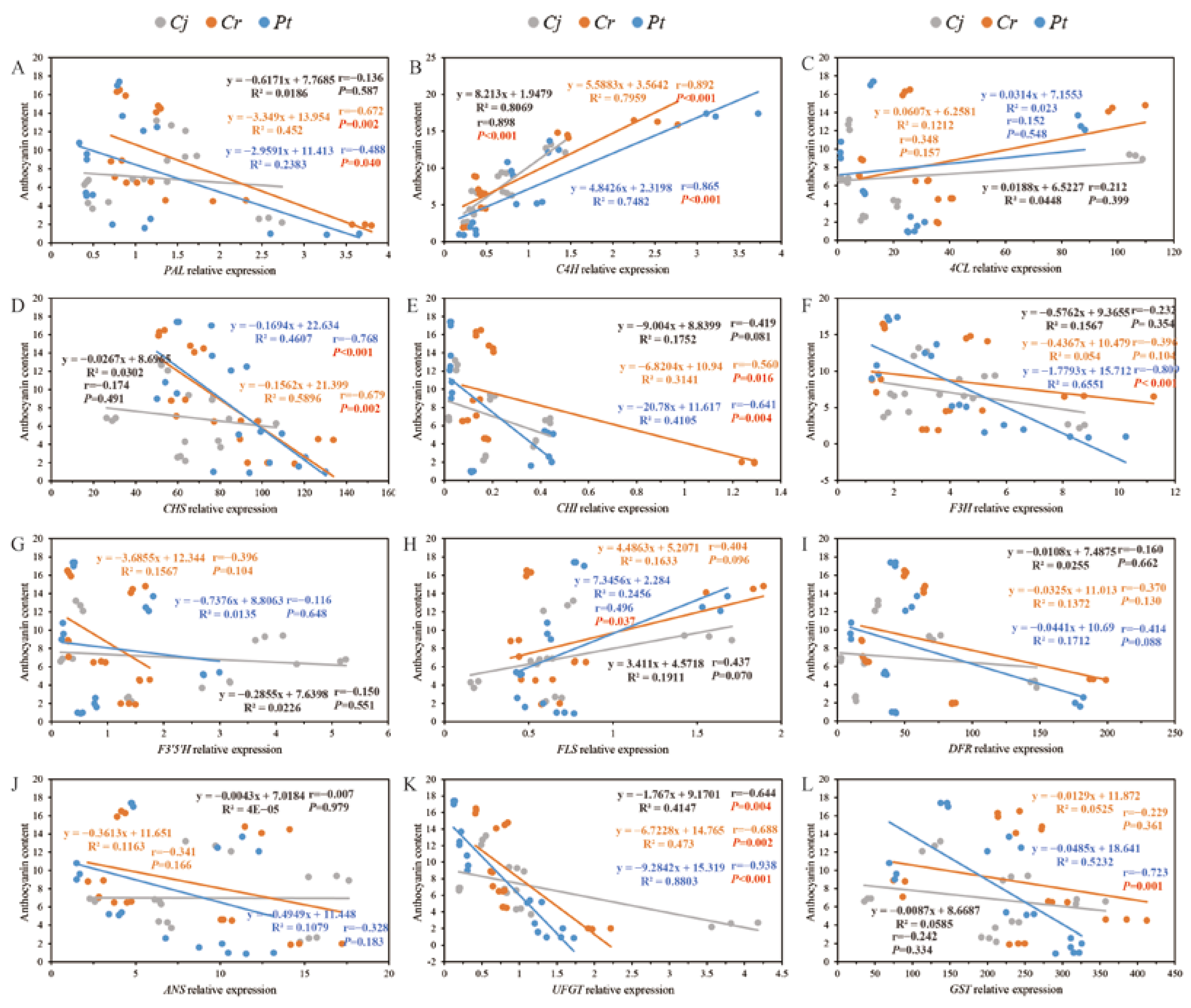
3.8. The Relationship between Anthocyanin Accumulation and Other Parameters (Enzyme Activity and Relative Gene Expression)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cebadera-Miranda, L.; Domínguez, L.; Dias, M.I.; Barros, L.; Ferreira, I.C.; Igual, M.; Martínez-Navarrete, N.; Fernández-Ruiz, V.; Morales, P.; Cámara, M. Sanguinello and Tarocco (Citrus sinensis [L.] Osbeck): Bioactive compounds and colour appearance of blood oranges. Food Chem. 2014, 270, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Grosso, G.; Galvano, F.; Mistretta, A.; Marventano, S.; Nolfo, F.; Calabrese, G.; Buscemi, S.; Drago, F.; Veronesi, U.; Scuderi, A. Red orange: Experimental models and epidemiological evidence of its benefits on human health. Oxidative Med. Cell. Longev. 2013, 2013, 157240. [Google Scholar] [CrossRef] [PubMed]
- Fallico, B.; Ballistreri, G.; Arena, E.; Brighina, S.; Rapisarda, P. Bioactive compounds in blood oranges (Citrus sinensis (L.) Osbeck): Level and intake. Food Chem. 2017, 215, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons control fruit-specific, cold-dependent accumulation of anthocyanins in blood oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef] [Green Version]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and human health—a focus on oxidative stress, inflammation and disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Ferlito, F.; Licciardello, C.; Allegra, M.; Strano, M.C.; Di Silvestro, S.; Russo, M.P.; Paolo, D.P.; Caruso, P.; Casas, G.L.; et al. Pomological diversity of the Italian blood orange germplasm. Sci. Hortic. 2016, 213, 331–339. [Google Scholar] [CrossRef]
- Bowman, K.D.; Joubert, J. Citrus Rootstocks. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F., Eds.; Woodhead Publishing: Duxford, UK; Elsevier: Duxford, UK, 2020; pp. 105–127. [Google Scholar] [CrossRef]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; Von Wettberg, E.J.B.; Miller, A.J. Rootstocks: Diversity, domestication, and impacts on shoot phenotypes. Trends Plant Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Liu, J.; Chen, K.; Hu, Q.; Yang, M.; Zhou, Q.; Li, H.; Jian, H.; Guan, B. Preliminary study on Ziyang xiangcheng (Citrus junos Sieb.ex Tanaka), a special local citrus germplasm. Southwest China J. Agric. Sci. 2008, 21, 1658–1660, (In Chinese with English Abstract). [Google Scholar]
- Usman, M.; Fatima, B. Mandarin (Citrus Reticulata Blanco) breeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J.M., Jain, S.M., Johnson, D., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 465–533. [Google Scholar] [CrossRef]
- Guo, W.; Deng, X. Wide somatic hybrids of Citrus with its related genera and their potential in genetic improvement. Euphytica 2001, 118, 175–183. [Google Scholar] [CrossRef]
- Swingle, W.T.; Reece, P.C. The Botany of Citrus and Its Wild Relatives. In The Citrus Industry; Reuther, W., Webber, H.J., Batchelor, L., Eds.; University of California Press: Berkeley, CA, USA, 1967; Volume 1, pp. 190–430. [Google Scholar]
- Parvaneh, T.; Abedi, B.; Davarynejad, G.H.; Ganji Moghadam, E. Enzyme activity, phenolic and flavonoid compounds in leaves of Iranian red flesh apple cultivars grown on different rootstocks. Sci. Hortic. 2019, 246, 862–870. [Google Scholar] [CrossRef]
- Font i Forcada, C.; Gogorcena, Y.; Moreno, M. Fruit sugar profile and antioxidants of peach and nectarine cultivars on almond×peach hybrid rootstocks. Sci. Hortic. 2013, 164, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Milošević, T.; Milošević, N.; Mladenović, J. Combining fruit quality and main antioxidant attributes in the sour cherry: The role of new clonal rootstock. Sci. Hortic. 2020, 265, 109236. [Google Scholar] [CrossRef]
- Ochmian, I.; Oszmiański, J.; Lachowicz, S.; Krupa-Małkiewicz, M. Rootstock effect on physico-chemical properties and content of bioactive compounds of four cultivars Cornelian cherry fruits. Sci. Hortic. 2019, 256, 108588. [Google Scholar] [CrossRef]
- Cheng, J.; Wei, L.; Mei, J.; Wu, J. Effect of rootstock on phenolic compounds and antioxidant properties in berries of grape (Vitis vinifera L.) cv. ‘Red Alexandria’. Sci. Hortic. 2017, 217, 137–144. [Google Scholar] [CrossRef]
- Incesu, M.; Çimen, B.; Yesiloglu, T.; Yilmaz, B. Rootstock effects on yield, fruit quality, rind and juice color of “Moro” blood orange. J. Food Agric. Environ. 2013, 11, 867–871. [Google Scholar]
- Lana, G.; Modica, G.; Casas, G.L.; Siracusa, L.; La Malfa, S.; Gentile, A.; Sicilia, A.; Distefano, G.; Continella, A. Molecular insights into the effects of rootstocks on maturation of blood oranges. Horticulturae 2021, 7, 468. [Google Scholar] [CrossRef]
- Continella, A.; Pannitteri, C.; La Malfa, S.; Legua, P.; Distefano, G.; Nicolosi, E.; Gentile, A. Influence of different rootstocks on yield precocity and fruit quality of ‘Tarocco Scirè’ pigmented sweet orange. Sci. Hortic. 2018, 230, 62–67. [Google Scholar] [CrossRef]
- Nakazawa, A.; Nozue, M.; Yasuda, H.; Takeba, G.; Kubo, H. Expression pattern and gene structure of phenylalanine ammonia-lyase in Pharbitis nil. J. Plant Res. 2001, 114, 323–328. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Yun, C.S.; Matsuda, F.; Sasaki, T.; Saito, K.; Tozawa, Y. Expression of bacterial tyrosine ammonia-lyase creates a novel p-coumaric acid pathway in the biosynthesis of phenylpropanoids in Arabidopsis. Planta 2010, 232, 209–218. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Sicilia, A.; Scialò, E.; Puglisi, I.; Lo Piero, A.R. Anthocyanin biosynthesis and DNA methylation dynamics in sweet orange fruit [Citrus sinensis L. (Osbeck)] under cold stress. J. Agric. Food Chem. 2020, 68, 7024–7031. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Alseekh, S.; Fernie, A.R. Conservation and diversification of flavonoid metabolism in the plant kingdom. Curr. Opin. Plant Biol. 2020, 55, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Lo Piero, A.R.; Puglisi, I.; Petrone, G. Gene isolation, analysis of expression, and in vitro synthesis of glutathione s-transferase from orange fruit [Citrus sinensis L. (Osbeck)]. J. Agric. Food Chem. 2006, 54, 9227–9233. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Lo Piero, A.R. The state of the art in biosynthesis of anthocyanins and its regulation in pigmented sweet oranges [(Citrus sinensis L. (Osbeck)]. J. Agric. Food Chem. 2015, 63, 4031–4041. [Google Scholar] [CrossRef]
- Carmona, L.; Zacarías, L.; Rodrigo, M.J. Stimulation of coloration and carotenoid biosynthesis during postharvest storage of “Navelina” orange fruit at 12 °C. Postharvest Biol. Technol. 2012, 74, 108–117. [Google Scholar] [CrossRef]
- Chen, M.; Xie, X.; Lin, Q.; Chen, J.; Grierson, D.; Yin, X.; Sun, C.; Chen, K. Differential expression of organic acid degradation-related genes during fruit development of Navel oranges (Citrus sinensis) in two habitats. Plant Mol. Biol. Report. 2013, 31, 1131–1140. [Google Scholar] [CrossRef]
- Rapisarda, P.; Fanella, F.; Maccarone, E. Reliability of analytical methods for determining anthocyanins in blood orange juices. J. Agric. Food Chem. 2000, 48, 2249–2252. [Google Scholar] [CrossRef]
- Liang, D.; Deng, H.; Deng, Q.; Lin, L.; Lv, X.; Wang, J.; Wang, Z.; Xiong, B.; Zhao, X.; Xia, H. Dynamic changes of phenolic compounds and their associated gene expression profiles occurring during fruit development and ripening of the Donghong kiwifruit. J. Agric. Food Chem. 2020, 68, 11421–11433. [Google Scholar] [CrossRef]
- Crifò, T.; Puglisi, I.; Petrone, G.; Recupero, G.R.; Piero, A.R.L. Expression analysis in response to low temperature stress in blood oranges: Implication of the flavonoid biosynthetic pathway. Gene 2011, 476, 1–9. [Google Scholar] [CrossRef]
- Liu, C.; Long, J.; Zhu, K.; Liu, L.; Chaoyang, L.; Zhang, H.; Linlin, L.; Xu, Q.; Deng, X. Characterization of a Citrus R2R3-MYB transcription factor that regulates the flavonol and hydroxycinnamic acid biosynthesis. Sci. Rep. 2016, 6, 25352. [Google Scholar] [CrossRef]
- Baldwin, E.A.; Bai, J.; Plotto, A.; Ritenour, M.A. Citrus fruit quality assessment; producer and consumer perspectives. Stewart Postharvest Rev. 2014, 10, 1–7. [Google Scholar]
- Legua, P.; Bellver, R.; Forner, J.B.; Forner-Giner, M.A. Trifoliata hybrids rootstocks for “Lane Late” navel orange in Spain. Sci. Agric. 2011, 68, 548–553. [Google Scholar] [CrossRef]
- Morales, J.; Tárrega, A.; Salvador, A.; Navarro, P.; Besada, C. Impact of ethylene degreening treatment on sensory properties and consumer response to citrus fruits. Food Res. Int. 2020, 127, 108641. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Díaz, D.; Hervalejo, A.; Pereira-Caro, G.; Muñoz-Redondo, J.M.; Romero-Rodriguez, E.; Arenas-Arenas, F.J.; Moreno-Rojas, J.M. Effect of rootstock and harvesting period on the bioactive compounds and antioxidant activity of two orange cultivars (‘Salustiana’ and ‘Sanguinelli’) widely used in juice industry. Processes 2020, 8, 1212. [Google Scholar] [CrossRef]
- Catalano, C.; Ciacciulli, A.; Salonia, F.; Russo, M.P.; Caruso, P.; Caruso, M.; Russo, G.; Distefano, G.; Licciardello, C. Target-genes reveal species and genotypic specificity of anthocyanin pigmentation in citrus and related genera. Genes 2020, 11, 807. [Google Scholar] [CrossRef]
- Wu, J.X.; Cao, J.Y.; Su, M.; Feng, G.Z.; Xu, Y.H.; Yi, H.L. Genome-wide comprehensive analysis of transcriptomes and small RNAs offers insights into the molecular mechanism of alkaline stress tolerance in a citrus rootstock. Hortic. Res. 2019, 6, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadernejad, N.; Ahmadimoghadam, A.; Hossyinifard, J.; Poorseyedi, S. Effect of different rootstocks on PAL activity and phenolic compounds in flowers, leaves, hulls and kernels of three pistachio (Pistacia vera L.) cultivars. Trees 2013, 27, 1681–1689. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Y.; Ren, L.; Shi, Q.; Zheng, B.; Miao, K.; Guo, X. Overexpression of Ps-CHI1, a homologue of the chalcone isomerase gene from tree peony (Paeonia suffruticosa), reduces the intensity of flower pigmentation in transgenic tobacco. Plant Cell Tissue Organ Cult. 2014, 116, 285–295. [Google Scholar] [CrossRef]
- Liu, Y.; Che, F.; Wang, L.; Meng, R.; Zhang, X.; Zhao, Z. Fruit coloration and anthocyanin biosynthesis after bag removal in non-red and red apples (Malus × domestica Borkh.). Molecules 2013, 18, 1549–1563. [Google Scholar] [CrossRef]
- Sparvoli, F.; Martin, C.; Scienza, A.; Gavazzi, G.; Tonelli, C. Cloning and molecular analysis of structural genes involved in flavonoid and stilbene biosynthesis in grape (Vitis vinifera L.). Plant Mol. Biol. 1994, 24, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.W.; Breen, P.J. Activity of phenylalanine ammonia-lyase (PAL) and concentrations of anthocyanins and phenolics in developing strawberry fruit. J. Am. Soc. Hortic. Sci. 1991, 116, 865–869. [Google Scholar] [CrossRef]
- Ju, Z. Fruit bagging, a useful method for studying anthocyanin synthesis and gene expression in apples. Sci. Hortic. 1998, 77, 155–164. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, J.R.; Hong, S.T.; Yoo, Y.K.; An, G.; Kim, S.R. Molecular cloning and analysis of anthocyanin biosynthesis genes preferentially expressed in apple skin. Plant Sci. 2003, 165, 403–413. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Genes Name | Froward 5′-3′ | Reverse 5′-3′ | Reference |
|---|---|---|---|---|
| XM_006481431.3 | PAL | GATTTGAGACATTTGGAGGA | ATGGATGAAGCTCTCCACTA | [33] |
| NM_001288840.1 | C4H | CGTGGGAAGGCGAAGCT | TCACAAGCCCCAAGATTGGT | [34] |
| NM_001288931.1 | CHI | AACAAAGTCGCATTTCCTCGGC | GCACGGCGTTCTCCTCCAAGTA | [34] |
| XM_006478851.2 | 4CL | CCGAAACGCACCTCATCTG | TGATGCCGAGATTCGACAAA | [34] |
| XM_006420545.2 | CHS | TCTATCGACGGGCATCTTC | TGCCTCGGTTAGGCTTTTC | [33] |
| NM_001288887.1 | F3H | CCCCTAATTTAAGGCTACCC | GACATCGTTCACGAAAACATAC | [29] |
| XM_006446247.2 | F3′5′H | GAATGGGGATTGTGCTTGTTG | CGAGGCGAGGCCGAAGC | [29] |
| NM_001288931.1 | DRF | GCTGTTCGTGCTACTGTTC | GGCTAAATCGGCTTTCCATA | [33] |
| XM_006466120.1 | FLS | GCCTCCGTCTTCTATCAACTACC | TACTCCTCATTCACCGCTCG | [33] |
| XM_025097974.1 | ANS | GGGTGACTAAATGTGTT | CAAGTCCCCTGTGAAGAATA | [33] |
| NM_001320060.1 | UFGT | TCTTCAGCACTCCGCAATC | TCCATCGGATACGTCGTAAG | [33] |
| NM_001288887.1 | GST | GCAGCAAAGTATGCAAACC | GAGCCATTGATCCACCAAAG | [33] |
| XM_006488084.3 | EF-1α | GGCTAGGTACGATGAAATTG | GTTGTCACCCTCGAAACC | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Deng, H.; Xiong, B.; Li, S.; Yang, L.; Yang, Y.; Huang, S.; Tan, L.; Sun, G.; Wang, Z. Rootstock Effects on Anthocyanin Accumulation and Associated Biosynthetic Gene Expression and Enzyme Activity during Fruit Development and Ripening of Blood Oranges. Agriculture 2022, 12, 342. https://doi.org/10.3390/agriculture12030342
Chen Z, Deng H, Xiong B, Li S, Yang L, Yang Y, Huang S, Tan L, Sun G, Wang Z. Rootstock Effects on Anthocyanin Accumulation and Associated Biosynthetic Gene Expression and Enzyme Activity during Fruit Development and Ripening of Blood Oranges. Agriculture. 2022; 12(3):342. https://doi.org/10.3390/agriculture12030342
Chicago/Turabian StyleChen, Zhaofang, Honghong Deng, Bo Xiong, Sichen Li, Liancong Yang, Youting Yang, Shengjia Huang, Liping Tan, Guochao Sun, and Zhihui Wang. 2022. "Rootstock Effects on Anthocyanin Accumulation and Associated Biosynthetic Gene Expression and Enzyme Activity during Fruit Development and Ripening of Blood Oranges" Agriculture 12, no. 3: 342. https://doi.org/10.3390/agriculture12030342
APA StyleChen, Z., Deng, H., Xiong, B., Li, S., Yang, L., Yang, Y., Huang, S., Tan, L., Sun, G., & Wang, Z. (2022). Rootstock Effects on Anthocyanin Accumulation and Associated Biosynthetic Gene Expression and Enzyme Activity during Fruit Development and Ripening of Blood Oranges. Agriculture, 12(3), 342. https://doi.org/10.3390/agriculture12030342