
Late to the Party—Transferred Mulch from Green Manures Delays Colorado Potato Beetle Infestation in Regenerative Potato Cropping Systems

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
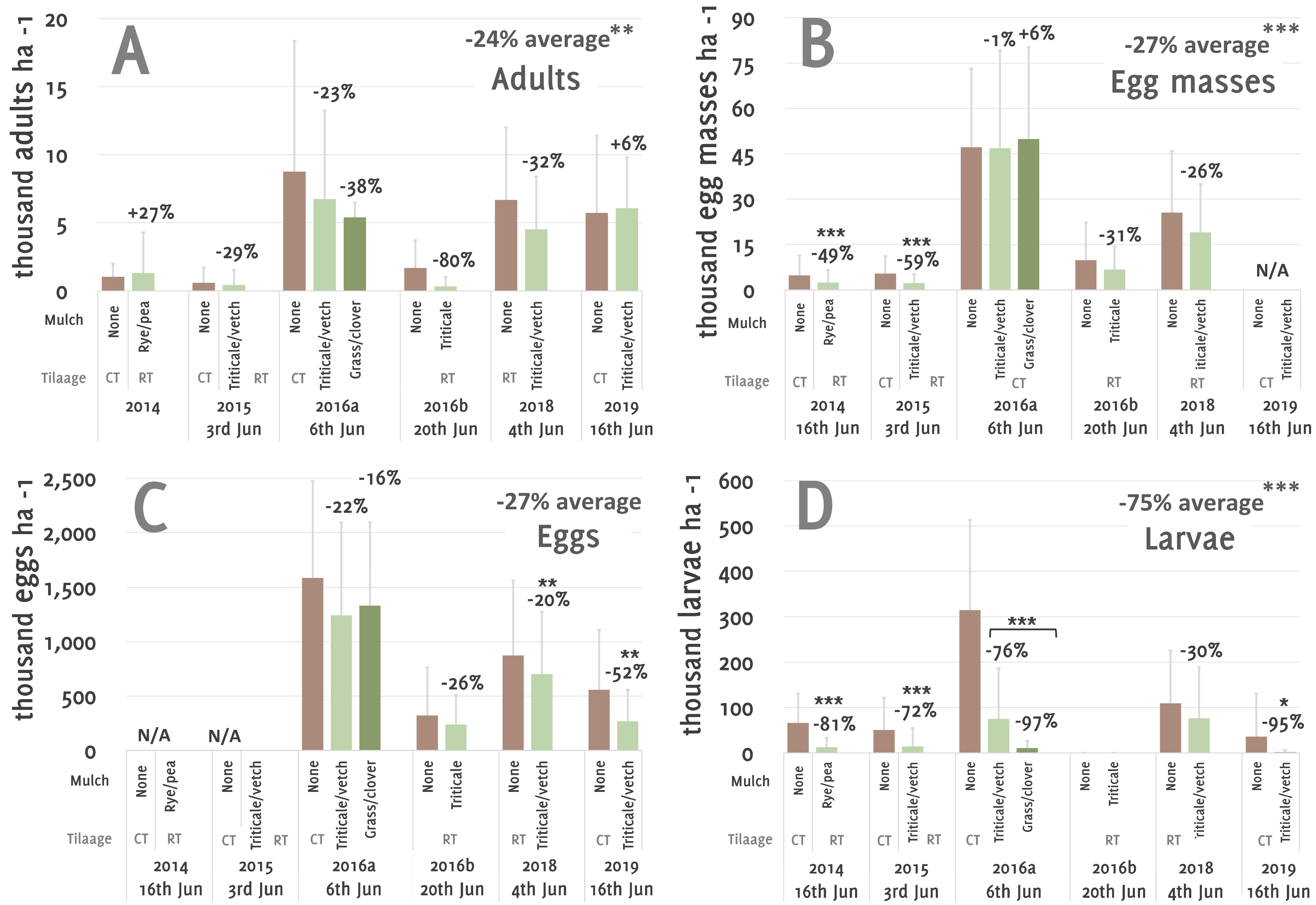
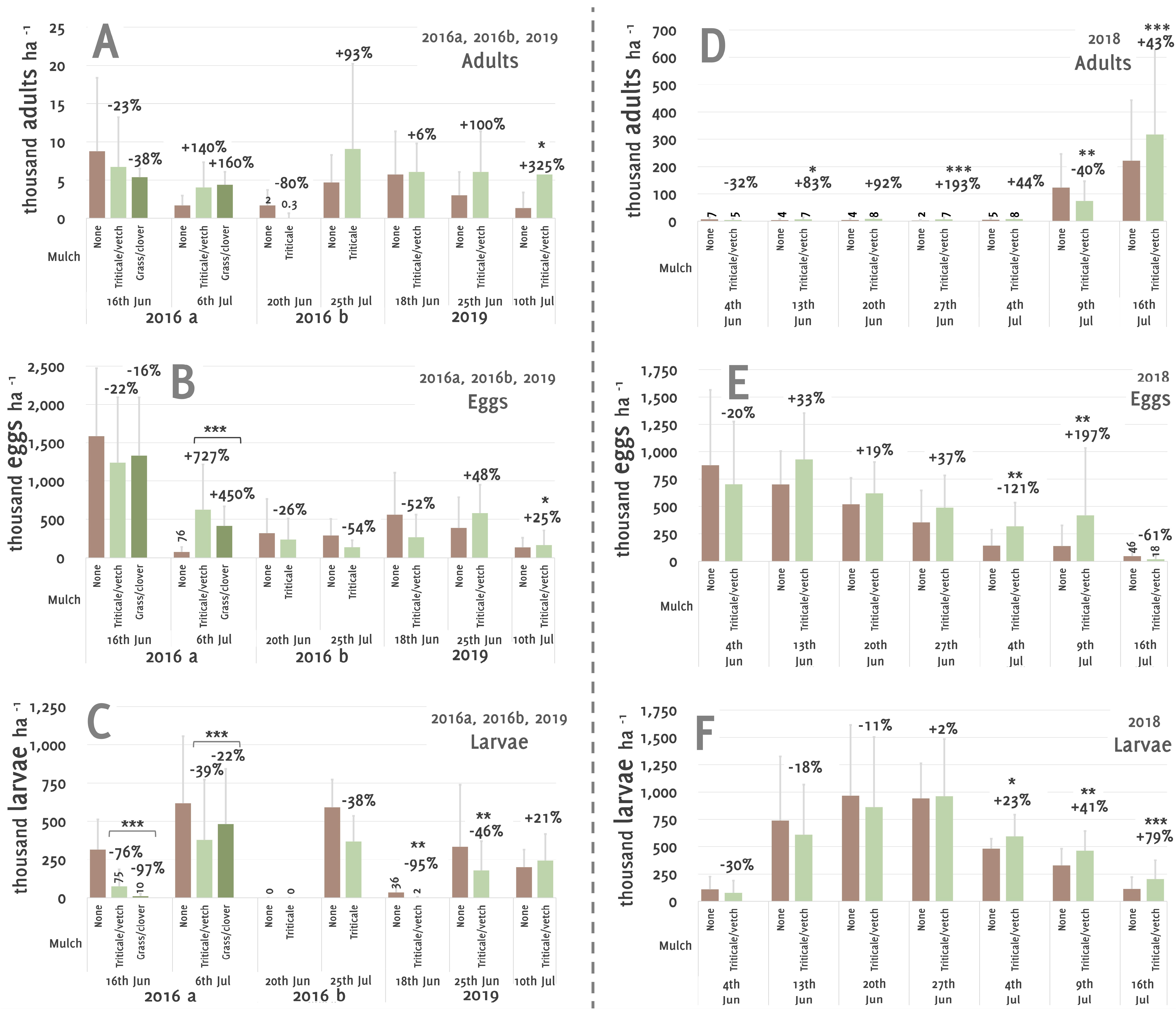
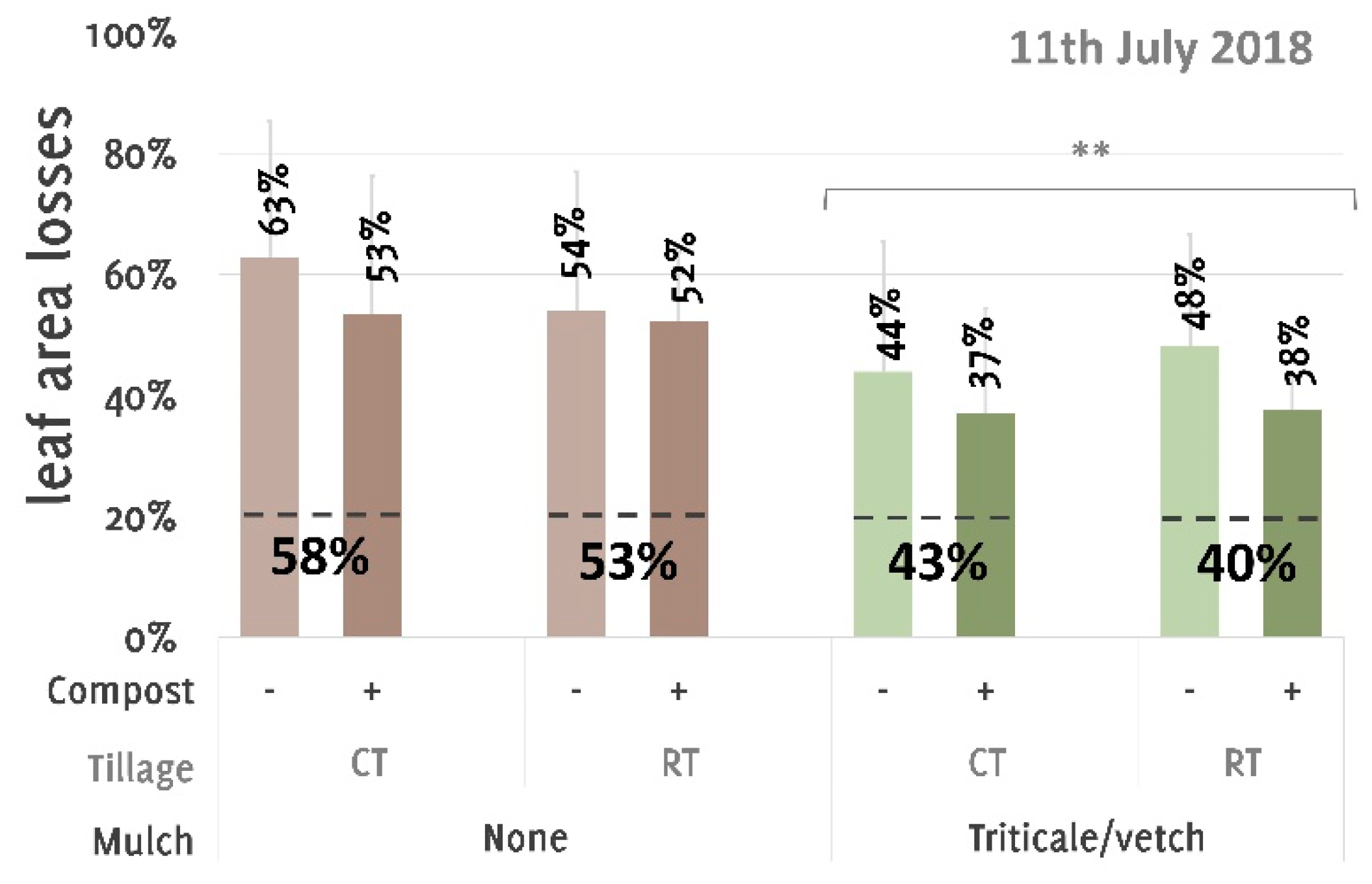
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Alyokhin, A.; Benkovskaya, G.; Udalov, M. Colorado Potato Beetle. In Insect Pests of Potato; Elsevier: Amsterdam, The Netherlands, 2022; pp. 29–43. ISBN 978-0-12-821237-0. [Google Scholar]
- Ferro, D.N.; Logan, J.A.; Voss, R.H.; Elkinton, J.S. Colorado Potato Beetle (Coleoptera: Chrysomelidae) Temperature-Dependent Growth and Feeding Rates. Environ. Entomol. 1985, 14, 343–348. [Google Scholar] [CrossRef]
- Auerswald, K.; Gerl, G.; Kainz, M. Influence of Cropping System on Harvest Erosion under Potato. Soil Tillage Res. 2006, 89, 22–34. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Biklé, A.; Archuleta, R.; Brown, P.; Jordan, J. Soil Health and Nutrient Density: Preliminary Comparison of Regenerative and Conventional Farming. PeerJ 2022, 10, e12848. [Google Scholar] [CrossRef] [PubMed]
- Schreefel, L.; Schulte, R.P.O.; de Boer, I.J.M.; Schrijver, A.P.; van Zanten, H.H.E. Regenerative Agriculture—The Soil Is the Base. Glob. Food Secur. 2020, 26, 100404. [Google Scholar] [CrossRef]
- Giller, K.E.; Hijbeek, R.; Andersson, J.A.; Sumberg, J. Regenerative Agriculture: An Agronomic Perspective. Outlook Agric. 2021, 50, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Cabral, L.; Sumberg, J. The Use of Epic Narratives in Promoting ‘Natural Agriculture’. Outlook Agric. 2022, 51, 129–136. [Google Scholar] [CrossRef]
- Newton, P.; Civita, N.; Frankel-Goldwater, L.; Bartel, K.; Johns, C. What Is Regenerative Agriculture? A Review of Scholar and Practitioner Definitions Based on Processes and Outcomes. Front. Sustain. Food Syst. 2020, 4, 577723. [Google Scholar] [CrossRef]
- Junge, S.M.; Storch, J.; Finckh, M.R.; Schmidt, J.H. Developing Organic Minimum Tillage Farming Systems for Central and Northern European Conditions. In No-till Farming Systems for Sustainable Agriculture; Dang, Y.P., Dalal, R.C., Menzies, N.W., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 173–192. [Google Scholar] [CrossRef]
- Finckh, M.R.; Junge, S.M.; Schmidt, J.H.; Weedon, O.D. Disease and Pest Management in Organic Farming: A Case for Applied Agroecology. In Burleigh Dodds Series in Agricultural Science; Köpke, U., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; Volume 33, pp. 271–301. [Google Scholar] [CrossRef]
- Alyokhin, A.; Nault, B.; Brown, B. Soil Conservation Practices for Insect Pest Management in Highly Disturbed Agroecosystems—A Review. Entomol. Exp. Appl. 2020, 168, 7–27. [Google Scholar] [CrossRef]
- Hunt, D. Reduced Tillage Practices for Managing the Colorado Potato Beetle in Processing Tomato Production. HortScience Publ. Am. Soc. Hortic. Sci. (USA) 1998, 33, 279–282. [Google Scholar]
- Zehnder, G.W.; Linduska, J.J. Influence of Conservation Tillage Practices on Populations of Colorado Potato Beetle (Coleoptera: Chrysomelidae) in Rotated and Nonrotated Tomato Fields. Environ. Entomol. 1987, 16, 135–139. [Google Scholar] [CrossRef]
- Hofmeester, Y. Effects of Fertilization on Pests and Diseses. Neth. J. Plant Pathol. 1992, 98, 257–264. [Google Scholar] [CrossRef]
- Schaerffenberg, B. Der Einfluß der Edelkompostdüngung auf das Auftreten des Kartoffelkäfers (Leptinotarsa decemlineata Say). Z. für Angew. Entomol. 1968, 62, 90–97. [Google Scholar] [CrossRef]
- Zehnder, G.W.; Hough-Goldstein, J. Colorado Potato Beetle (Coleoptera: Chrysomelidae) Population Development and Effects on Yield of Potatoes With and Without Straw Mulch. J. Econ. Entomol. 1990, 83, 1982–1987. [Google Scholar] [CrossRef]
- Stoner, K.A. Effects of Straw and Leaf Mulches and Trickle Irrigation on the Abundance of Colorado Potato Beetles (Coleoptera: Chrysomelidae) on Potato in Connecticut. J. Entomol. Sci. 1993, 28, 393–403. [Google Scholar] [CrossRef]
- Brust, G.E. Natural Enemies in Straw-Mulch Reduce Colorado Potato Beetle Populations and Damage in Potato. Biol. Control 1994, 4, 163–169. [Google Scholar] [CrossRef]
- Brust, G.E. Interaction of Mulch and Bacillus thuringiensis subsp. Tenebrionis on Colorado Potato Beetle (Coleoptera: Chrysomelidae) Populations and Damage in Potato. J. Econ. Entomol. 1996, 89, 467–474. [Google Scholar] [CrossRef]
- Hoy, C.W.; Wyman, J.A.; Vaughn, T.T.; East, D.A.; Kaufman, P. Food, Ground Cover, and Colorado Potato Beetle (Coleoptera: Chrysomelidae) Dispersal in Late Summer. J. Econ. Entomol. 1996, 89, 963–969. [Google Scholar] [CrossRef]
- Stoner, K.A.; Ferrandino, F.J.; Gent, M.P.N.; Elmer, W.H.; Lamondia, J.A. Effects of Straw Mulch, Spent Mushroom Compost, and Fumigation on the Density of Colorado Potato Beetles (Coleoptera: Chrysomelidae) in Potatoes. J. Econ. Entomol. 1996, 89, 1267–1280. [Google Scholar] [CrossRef]
- Johnson, J.M.; Hough-Goldstein, J.A.; Vangessel, M.J. Effects of Straw Mulch on Pest Insects, Predators, and Weeds in Watermelons and Potatoes. Environ. Entomol. 2004, 33, 1632–1643. [Google Scholar] [CrossRef]
- Döring, T.; Heimbach, U.; Thieme, T.; Saucke, H. Aspects of Straw Mulching in Organic Potatoes—II. Effects on Potato Virus Y, Leptinotarsa Decemlineata (Say) and Tuber Yield. Nachrichtenbl Deut Pflanzenschutzd. 2006, 58, 93–97. [Google Scholar]
- Szendrei, Z.; Kramer, M.; Weber, D.C. Habitat Manipulation in Potato Affects Colorado Potato Beetle Dispersal. J. Appl. Entomol. 2009, 133, 711–719. [Google Scholar] [CrossRef]
- Dvořák, P.; Tomášek, J.; Kuchtová, P.; Hamouz, K.; Hajšlová, J.; Schulzová, V. Effect of Mulching Materials on Potato Production in Different Soil-Climatic Conditions. Roman Agric. Res. 2012, 29, 201–209. [Google Scholar]
- Dvořák, P.; Kuchtová, P.; Tomásek, J. Response of Surface Mulching of Potato (Solanum Tuberosum) on SPAD Value, Colorado Potato Beetle and Tuber Yield. Int. J. Agric. Biol. 2013, 15, 798. [Google Scholar]
- Genger, R.K.; Rouse, D.I.; Charkowski, A.O. Straw Mulch Increases Potato Yield and Suppresses Weeds in an Organic Production System. Biol. Agric. Hortic. 2018, 34, 53–69. [Google Scholar] [CrossRef]
- Junge, S.M.; Wedemeyer, R.; Saucke, H.; Finckh, M.R. Einflüsse von Minimalbodenbearbeitung und Transfermulch auf den Initialbefall und die Populationsentwicklung des Kartoffelkäfers (Leptinotarsa decemlineata). In Proceedings of the Ökologischen Landbau weiterdenken—Verantwortung Übernehmen—Vertrauen Stärken. Beiträge der 14. Wissenschaftstagung Ökologischer Landbau, Freising, Germany, 7–10 March 2017; Dr. Köster: Berlin, Germany, 2017; Volume 14, p. 174. [Google Scholar] [CrossRef]
- Döring, T.F.; Brandt, M.; Heß, J.; Finckh, M.R.; Saucke, H. Effects of Straw Mulch on Soil Nitrate Dynamics, Weeds, Yield and Soil Erosion in Organically Grown Potatoes. Field Crops Res. 2005, 94, 238–249. [Google Scholar] [CrossRef]
- Dvorak, P.; Tomasek, J.; Hamouz, K. Ecological Growing of Potatoes with Using of Grass Mulch and Black Textile Mulch. In Proceedings of the 45th Croatian and 5th International Symposium on Agriculture, Opatija, Croatia, 15–19 February 2010; Poljoprivredni fakultet Sveucilista Jurja Strossmayera u Osijeku = Faculty of Agriculture University of Josip Juraj Strossmayer in Osijek: Osijek, Croatia, 2010; pp. 65–69. [Google Scholar]
- Schmidt, J.H.; Junge, S.; Finckh, M.R. Cover Crops and Compost Prevent Weed Seed Bank Buildup in Herbicide-Free Wheat-Potato Rotations under Conservation Tillage. Ecol. Evol. 2019, 9, 2715–2724. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Dixon, P. VEGAN, a Package of R Functions for Community Ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 1–15. ISBN 978-1-118-44511-2. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.; Singmann, H. Lme4: Linear Mixed-Effects Models Using “Eigen” and S4 Classes. R Package Version 2020, 1, 15. [Google Scholar]
- Hartig, F.; Lohse, L. DHARMa—Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version. 2022, Volume 320. Available online: https://cran.r-project.org/web/packages/DHARMa/index.html (accessed on 2 December 2022).
- Schmidt, J.H.; Hallmann, J.; Finckh, M.R. Bacterivorous Nematodes Correlate with Soil Fertility and Improved Crop Production in an Organic Minimum Tillage System. Sustainability 2020, 12, 6730. [Google Scholar] [CrossRef]
- Wilson, C.; Zebarth, B.J.; Burton, D.L.; Goyer, C. Short-Term Effects of Diverse Compost Products on Soil Quality in Potato Production. Soil Sci. Soc. Am. J. 2018, 82, 889–900. [Google Scholar] [CrossRef]
- Herencia, J.F.; Ruiz, J.C.; Morillo, E.; Melero, S.; Villaverde, J.; Maqueda, C. The Effect of Organic and Mineral Fertilization on Micronutirent Abailability in Soil. Soil Sci. 2008, 173, 69–80. [Google Scholar] [CrossRef]
- Wilson, C.; Zebarth, B.J.; Goyer, C.; Burton, D.L. Effect of Diverse Compost Products on Soilborne Diseases of Potato. Compost Sci. Util. 2018, 26, 156–164. [Google Scholar] [CrossRef]
- Nguyen, H.D.D.; Nansen, C. Edge-Biased Distributions of Insects. A Review. Agron. Sustain. Dev. 2018, 38, 11. [Google Scholar] [CrossRef]
- Junge, S.M.; Kaplan, F.; Wedemeyer, R.; Finckh, M.R. Transfermulch aus Gründüngern als agrarökologisches Werkzeug gegen Alternaria solani im Ökologischen Kartoffelanbau. In Proceedings of the Gesunde Pflanzen in Verantwortung für unsere Welt, Webinar, 21–23 September 2021. [Google Scholar] [CrossRef]
- Jermy, T.; Szentesi, Á.; Horváth, J. Host Plant Finding in Phytophagous Insects: The Case of the Colorado Potato Beetle. Entomol. Exp. Appl. 1988, 49, 83–98. [Google Scholar] [CrossRef]
- Alyokhin, A.; Porter, G.; Groden, E.; Drummond, F. Colorado Potato Beetle Response to Soil Amendments: A Case in Support of the Mineral Balance Hypothesis? Agric. Ecosyst. Environ. 2005, 109, 234–244. [Google Scholar] [CrossRef]
- Thiery, D.; Visser, J.H. Masking of Host Plant Odour in the Olfactory Orientation of the Colorado Potato Beetle. Entomol. Exp. Appl. 1986, 41, 165–172. [Google Scholar] [CrossRef]
- Ji, S.; Unger, P.W. Soil Water Accumulation under Different Precipitation, Potential Evaporation, and Straw Mulch Conditions. Soil Sci. Soc. Am. J. 2001, 65, 442. [Google Scholar] [CrossRef]
- Lashomb, J.H.; Ng, Y.-S. Colonization by Colorado Potato Beetles, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae), in Rotated and Nonrotated Potato Fields. Environ. Entomol. 1984, 13, 1352–1356. [Google Scholar] [CrossRef]
- Phelan, P.L. Soil-Management History and the Role of Plant Mineral Balance as a Determinant of Maize Susceptibility to the European Corn Borer. Biol. Agric. Hortic. 1997, 15, 25–34. [Google Scholar] [CrossRef]
- Wright, R.J. Evaluation of Crop Rotation for Control of Colorado Potato Beetles (Coleoptera: Chrysomelidae) in Commercial Potato Fields on Long Island. J. Econ. Entomol. 1984, 77, 1254–1259. [Google Scholar] [CrossRef]
- Boiteau, G.; Lynch, D.H.; Martin, R.C. Influence of Fertilization on the Colorado Potato Beetle, Leptinotarsa decemlineata, in Organic Potato Production. Environ. Entomol. 2008, 37, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Boiteau, G.; Picka, J.D.; Watmough, J. Potato Field Colonization by Low-Density Populations of Colorado Potato Beetle as a Function of Crop Rotation Distance. J. Econ. Entomol. 2008, 101, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, S.M.; Hiltunen, L.H.; Santala, J.; Döring, T.F.; Ketola, J.; Kankaala, A.; Virtanen, E.; Valkonen, J.P.T. Comparison of Straw Mulch, Insecticides, Mineral Oil, and Birch Extract for Control of Transmission of Potato Virus Y in Seed Potato Crops. Potato Res. 2014, 57, 59–75. [Google Scholar] [CrossRef]
- De Nazareno, N.R.X.; Finckh, M.R.; dos Santos Canalli, L.B.; Yada, I.F.U.; Kawakami, J. Potential Use of Fresh Mulch to Curb Potato Late Blight Epidemics in Brazil. Summa Phytopathol. 2020, 46, 191–197. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mulch (Quantity) | Mulch Effects | Mechanisms Suggested by Authors | Adults | Egg Masses | Larvae | Predators | Defoliation | Yield | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Wheat straw (6–10 cm) | ↓ Adults, egg masses ↑ Development time larvae ↑ Yield | Effects on plant volatile compounds ↓ Water stress Physical obstruction | x | x | x | x | [16] | ||
| Rye straw (10 cm) leaf Mulch (0–5 cm) | ↓ Defoliation by 50% ↑ Yield | ↑ Predators ↓ Ability of larva to climb back after falling mulch type or depth affects the effect | x | x | x | x | [17] | ||
| Wheat straw (8–10 cm) | ↓ Larvae ↑ 2nd Gen. Adults and Larvae ↑ Predators on Plant ↓ Defoliation | ↑ Predators | x | x | x | x | x | [18] | |
| Wheat straw (2.4 t ha) | ↓ Larvae (1st, 2nd and 4th instar) ↑ Soil predators ↓ Defoliation | ↑ Predators | x | x | x | x | x | x | [19] |
| Straw (15–30 cm) | ↓ Defoliation | Difficulty in finding plants due to mulch Potato variety and senesces process of the foliage | x | x | [20] | ||||
| Straw mulch 1 | ↓ Adults ↓ Larvae ↓ Defoliation | ↑ Predators ↓ Attractiveness of potato plants infected by Verticillium dahliae and Pratylenchus penetrans | x | x | x | x | x | [21] | |
| Wheat straw 1–1.3 t ha−1 | ↑ inititial Adults ↓ Adults in later Season ↑ Soil Predators | ↑ Soil Predators Temporal influences on CPB population | x | x | x | x | [22] | ||
| Wheat straw 1.5–5 t ha−1 | ↓ CPB infestation (ns.) Dose effects | ↑ Soil Predators | x | x | x | x | x | [23] | |
| Rye 5cm, vetch (Residues) 10 cm | Interfere field colonization | visual or chemical deterrence and/or physical barrier | x | [24] | |||||
| polypropylene-textile Mulch Chopped grass (2.5 cm) | ↑ eggs in textile mulch ↓ Larvae ↑ Yield | ↑ Soil Temperature | x | x | x | x | [25,26] | ||
| Winter wheat or rye straw 18–22 t ha−1 | ↓Larvae in 3/4 Experiments ↑ Predators ↑ Yield | ↑ Number of predators in pan and pit traps | x | x | x | x | [27] | ||
| Triticale mulch 133 t ha−1(fresh matter) | ↓ eggs ↓ Larvae ↓ Defoliation ↑ Yield | balanced nutrition by mulch makes potato plants less susceptible to potato beetle | x | x | x | x | x | [10,28] |
| Mulch 2 | Number of Assessed | Assessments | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | >Tillage 1 | >Other Factors | >Type | >t FM ha−1 | >t DM ha−1 | >C/N-ratio | >Plot Size (m) | >Plots | >Plants | >CPB-Assessment | >BBCH 3 | >Adults | >Egg Masses | >Eggs | >Larvae | >Defoliation 4 | Diseases 5 |
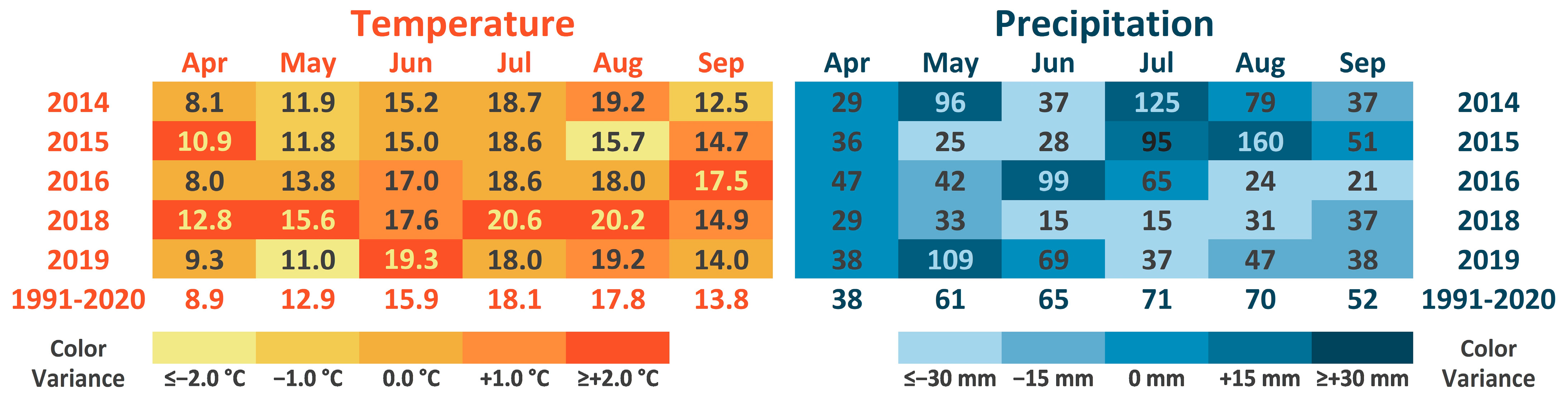
| 2014 (LTE) | CT, RT | 2 precrops, +/− compost | RP | 49, 6 | 25 | 28 | 6 × 15 | 64 | 50 | 17.6. | 51 | x | x | x | P. infestans | ||
| 2015 (LTE) | CT, RT | 2 precrops, +/− compost | TV | 88, 3 | 30 | 23 | 6 × 15 | 64 | 30 | 3.7. | 54 | x | x | x | P. infestans, Alternaria | ||
| 2016a | CT | GC, TV | 147, 129 | 35, 33 | 39, 41 | 4.5 × 17 | 12 | 30 | 16.6.; 6.7. | 50 | x | x | x | x | P. infestans | ||
| 2016b | CT | T | 133, 0 | 37 | 52 | 4.5 × 15 | 8 | 30 | 20.6.; 25.7. | 45 | x | x | x | x | P. infestans | ||
| 2018 (LTE) | CT, RT | +/− compost | TV | 63 | 33 | 19 | 6 × 15 | 16/64; 64/64 defoliation | 30 | 4., 13, 20, 27.6.; 4., 9., 16.7. | 31 | x | x | x | x | x 4 | - |
| 2019 6 (LTE) | CT, RT | TV | 47 | 23 | 24 | 6 × 15 | 16/64 | 15 | 18., 25.6., 10.7. | 52 | x | x | x | x | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Junge, S.M.; Leisch-Waskönig, S.; Winkler, J.; Kirchner, S.M.; Saucke, H.; Finckh, M.R. Late to the Party—Transferred Mulch from Green Manures Delays Colorado Potato Beetle Infestation in Regenerative Potato Cropping Systems. Agriculture 2022, 12, 2130. https://doi.org/10.3390/agriculture12122130
Junge SM, Leisch-Waskönig S, Winkler J, Kirchner SM, Saucke H, Finckh MR. Late to the Party—Transferred Mulch from Green Manures Delays Colorado Potato Beetle Infestation in Regenerative Potato Cropping Systems. Agriculture. 2022; 12(12):2130. https://doi.org/10.3390/agriculture12122130
Chicago/Turabian StyleJunge, Stephan Martin, Simeon Leisch-Waskönig, Julian Winkler, Sascha Michael Kirchner, Helmut Saucke, and Maria Renate Finckh. 2022. "Late to the Party—Transferred Mulch from Green Manures Delays Colorado Potato Beetle Infestation in Regenerative Potato Cropping Systems" Agriculture 12, no. 12: 2130. https://doi.org/10.3390/agriculture12122130
APA StyleJunge, S. M., Leisch-Waskönig, S., Winkler, J., Kirchner, S. M., Saucke, H., & Finckh, M. R. (2022). Late to the Party—Transferred Mulch from Green Manures Delays Colorado Potato Beetle Infestation in Regenerative Potato Cropping Systems. Agriculture, 12(12), 2130. https://doi.org/10.3390/agriculture12122130