
Nutrient Composition and Growth of Yellow Mealworm (Tenebrio molitor) at Different Ages and Stages of the Life Cycle

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample
2.2. Measurement Procedure
2.3. Statistical Analysis
3. Results
3.1. Growth of Yellow Mealworm
3.1.1. Overall Growth of Yellow Mealworm
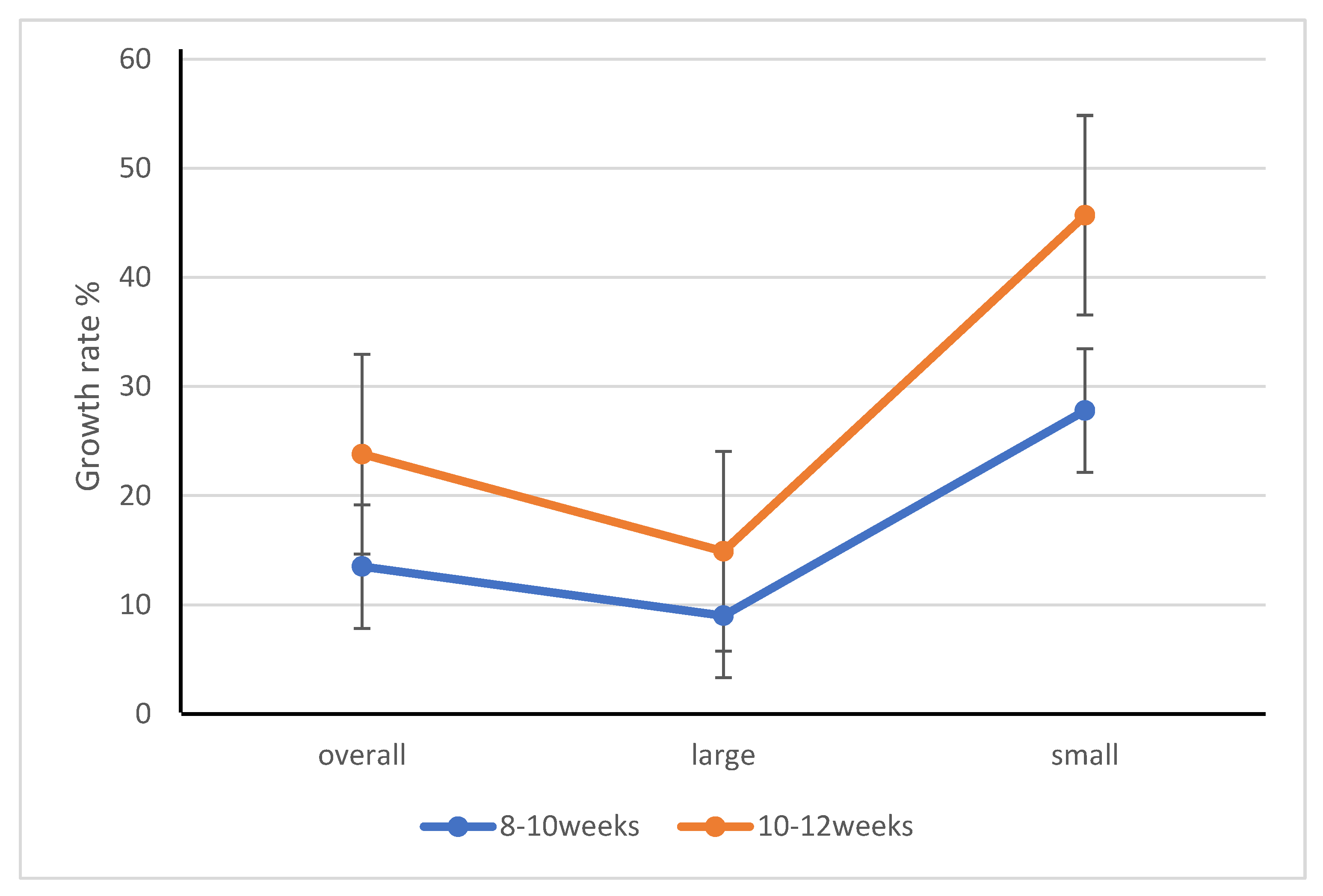
3.1.2. Growth of Different Age and Size Groups of Yellow Mealworm
3.2. Surface Area/Body Mass Ratio, Length and Width of Yellow Mealworm at Week 12
3.3. Nutrient Composition
3.3.1. Nutrient Composition at Different Ages
3.3.2. Nutrient Composition of Yellow Mealworm at Different Development Stages and Nutrient Composition of the Cuticle
4. Discussion
4.1. Growth
4.2. Nutrient Composition
4.2.1. Nutrient Composition Based on Age and Size
4.2.2. Nutrient Composition Based on the Developmental Stages and of the Cuticle
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| YM | Yellow mealworm |
| DM | Dry matter content |
| CP | Crude protein |
| CF | Crude fat |
| ADF | Acid detergent fibre |
| ADL | Acid detergent lignin |
| SA | Surface area |
| BM | Body mass |
| NFE | Nitrogen free extract |
References
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, J. Edible Insects of the World; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Jongema, Y. List of Edible Insect Species of the World. Wageningen, Laboratory of Entomology, Wageningen University. 2017. Available online: https://www.wur.nl/en/Research-Results/Chair-groups/Plant-Sciences/Laboratory-of-Entomology/Edible-insects/Worldwide-species-list.htm (accessed on 14 September 2021).
- Meyer-Rochow, V.B. Can insects help to ease the problem of world food shortage? Search 1975, 6, 261–262. [Google Scholar]
- Bodenheimer, F.S. Insects as Human Food. In Insects as Human Food: A Chapter of the Ecology of Man; Bodenheimer, F.S., Ed.; W. Junk: Hague, The Netherlands, 1951; pp. 7–38. [Google Scholar]
- Ghosh, S.; Lee, S.M.; Jung, C.; Meyer-Rochow, V.B. Nutritional composition of five commercial edible insects in South Korea. J. Asia-Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- Lange, K.W.; Nakamura, Y. Edible insects as future food: Chances and challenges. J. Future Foods 2021, 1, 38–46. [Google Scholar] [CrossRef]
- Lahteenmaki-Uutela, A.; Grmelová, N.; Hénault-Ethier, L.; Deschamps, M.H.; Vandenberg, G.W.; Zhao, A.; Zhang, Y.; Yang, B.; Nemane, V. Insects as food and feed: Laws of the European Union, United States, Canada, Mexico, Australia, and China. Eur. Food Feed L. Rev. 2017, 12, 22. [Google Scholar]
- Commission Implementing Regulation (EU) 2017/2469 of 20 December 2017 Laying Down Administrative and Scientific REQUIREMENTS for Applications Referred to in Article 10 of Regulation (EU) 2015/2283 of the European Parliament and of the Council on Novel Foods. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R2469 (accessed on 31 August 2021).
- Commission Implementing Regulation (EU) 2021/882 of 1 June 2021 Authorising the Placing on the Market of Dried Tenebrio molitor Larva as a Novel Food under Regulation (EU) 2015/2283 of the European Parliament and of the Council, and Amending Commission Implementing Regulation (EU) 2017/2470 (Text with EEA Relevance) (OJ L 194 01.06.2021), p. 16. Available online: http://data.europa.eu/eli/reg_impl/2021/882/oj (accessed on 31 August 2021).
- Araújo, R.R.S.; dos Santos Benfica, T.A.R.; Ferraz, V.P.; Santos, E.M. Nutritional composition of insects Gryllus assimilis and Zophobas morio: Potential foods harvested in Brazil. J. Food Compos. Anal. 2019, 76, 22–26. [Google Scholar] [CrossRef]
- Tubin, J.S.B.; Paiano, D.; de Oliveira Hashimoto, G.S.; Furtado, W.E.; Martins, M.L.; Durigon, E.; Emerenciano, M.G.C. Tenebrio molitor meal in diets for Nile tilapia juveniles reared in biofloc system. Aquaculture 2020, 519, 734763. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Józefiak, D.; Józefiak, A.; Kierończyk, B.; Rawski, M.; Świątkiewicz, S.; Długosz, J.; Engberg, R.M. Insects—A natural nutrient source for poultry—A review. Ann. Anim. Sci. 2016, 16, 297–313. [Google Scholar] [CrossRef]
- Motte, C.; Rios, A.; Lefebvre, T.; Do, H.; Henry, M.; Jintasataporn, O. Replacing fish meal with defatted insect meal (Yellow Mealworm Tenebrio molitor) improves the growth and immunity of pacific white shrimp (Litopenaeus vannamei). Animals 2019, 9, 258. [Google Scholar] [CrossRef]
- Caparros Megido, R.; Alabi, T.; Larreché, S.; Alexandra, L.; Haubruge, E.; Francis, F. Risks and valorization of insects in a food and feed context. Int. J. Entomol. 2015, 51, 215–258. [Google Scholar]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Czachorowski, S.; Peni, D. Will yellow mealworm become a source of safe proteins for Europe? Agriculture 2020, 10, 233. [Google Scholar] [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06343. [Google Scholar] [PubMed]
- Grau, T.; Vilcinskas, A.; Joop, G. Sustainable farming of the mealworm Tenebrio molitor for the production of food and feed. Z. Nat. C 2017, 72, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Sankian, Z.; Khosravi, S.; Kim, Y.; Lee, S. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters, and antioxidant enzyme activities of mandarin fish (Siniperca scherzeri) juveniles. Aquaculture 2018, 496, 79–87. [Google Scholar]
- Commission Regulation (EU) 2021/1372 of 17 August 2021 Amending Annex IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council as Regards the Prohibition to Feed Non-Ruminant Farmed Animals, Other than fur Animals, with Protein Derived from Animals. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=OJ:L:2021:295:TOC (accessed on 16 March 2022).
- Jeong, S.; Khosravi, S.; Yoon, K.; Kim, K.; Lee, B.; Hur, S.; Lee, S. Mealworm, Tenebrio molitor, as a feed ingredient for juvenile olive flounder, Paralichthys olivaceus. Aquac. Rep. 2021, 20, 100747. [Google Scholar] [CrossRef]
- Su, J.; Gong, Y.; Cao, S.; Lu, F.; Han, D.; Liu, H.; Jin, J.; Yang, Y.; Zhu, X.; Xie, S. Effects of dietary Tenebrio molitor meal on the growth performance, immune response, and disease resistance of yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish. Immunol. 2017, 69, 59–66. [Google Scholar] [CrossRef]
- Ido, A.; Hashizume, A.; Ohta, T.; Takahashi, T.; Miura, C.; Miura, T. Replacement of fish meal by defatted yellow mealworm (Tenebrio molitor) larvae in diet improves growth performance and disease resistance in red seabream (Pargus major). Animals 2019, 9, 100. [Google Scholar] [CrossRef]
- Govorushko, S. Global status of insects as food and feed source: A review. Trends Food Sci. Technol. 2019, 91, 436–445. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Gahukar, R.T.; Ghosh, S.; Jung, C. Chemical composition, nutrient quality, and acceptability of edible insects are affected by species, developmental stage, gender, diet, and processing method. Foods 2021, 10, 1036. [Google Scholar] [CrossRef]
- Gasco, L.; Józefiak, A.; Henry, M. Beyond the protein concept: Health aspects of using edible insects on animals. J. Insects Food Feed. 2020, 7, 715–741. [Google Scholar] [CrossRef]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary fatty acids influence the growth and fatty acid composition of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Jajić, I.; Krstović, S.; Petrović, M.; Urošević, M.; Glamočić, D.; Samardžić, M.; Popović, A.; Guljaš, D. Changes in the chemical composition of the yellow mealworm (Tenebrio molitor L.) reared on different feedstuffs. J. Anim. Feed. Sci. 2022, 31, 191–200. [Google Scholar] [CrossRef]
- Tabata, E.; Kashimura, A.; Wakita, S.; Ohno, M.; Sakaguchi, M.; Sugahara, Y.; Yoshihiro, K.; Vaclav, M.; Bauer, P.O.; Oyama, F. Gastric and intestinal proteases resistance of chicken acidic chitinase nominates chitin-containing organisms for alternative whole edible diets for poultry. Sci. Rep. 2017, 7, 6662. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Osafune, T.; Tamehira, S.; Yano, K. Piglets can secrete acidic mammalian chitinase from the pre weaning stage. Sci. Rep. 2021, 11, 1297. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, M.G.; Norberto, L.; Damini, R.; Musumeci, S. Human gastric juice contains chitinase that can degrade chitin. Ann. Nutr. Metab. 2007, 51, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Kipkoech, C.; Kinyuru, J.N.; Imathiu, S.; Meyer-Rochow, V.B.; Roos, N. In vitro study of cricket chitosan’s potential as a prebiotic and a promoter of probiotic microorganisms to control pathogenic bacteria in the human gut. Foods 2021, 10, 2310. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Methods 955.04, 922.06, 973.18, 930.15; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- ISO 6865:2000; Animal Feeding Stuffs—Determination of Crude Fibre Content—Method with Intermediate Filtration. International Organization for Standardization: Geneva, Switzerland, 2000.
- Hahn, T.; Roth, A.; Febel, E.; Fijalkowska, M.; Schmitt, E.; Arsiwalla, T.; Zibek, S. New methods for high-accuracy insect chitin measurement. J. Sci. Food Agric. 2018, 98, 5069–5073. [Google Scholar] [CrossRef]
- ISO 13906:2008; Animal Feeding Stuffs—Determination of Acid Detergent Fibre (ADF) and Acid Detergent Lignin (ADL) Contents. International Organization for Standardization: Geneva, Switzerland, 2008.
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Peni, D. Growth Potential of Yellow Mealworm Reared on Industrial Residues. Agriculture 2020, 10, 599. [Google Scholar] [CrossRef]
- Jones, L.; Cooper, R.; Harding, R. Composition of mealworm Tenebrio molitor larvae. J. Zoo Anim. Med. 1972, 3, 34–41. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.M.; Sagis, L.M.; Eisner-Schadler, V.; van Huis, A.; van Boekel, M.A. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.D. Complete nutrient composition of selected invertebrates commonly fed to insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.H.; Choi, W.H.; Hong, S.J.; Kim, N.J. Nutritional value of mealworm, Tenebrio molitor as food source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef]
- Zhao, X.; Vázquez-Gutiérrez, J.L.; Johansson, D.P.; Landberg, R.; Langton, M. Yellow mealworm protein for food purposes-extraction and functional properties. PLoS ONE 2016, 11, 0147791. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.D. Estimate of chitin in raw whole insects. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2007, 26, 105–115. [Google Scholar] [CrossRef]
- Subramanian, S.; Shankarganesh, K. Chapter 20—Insect Hormones (as Pesticides). In Ecofriendly Pest Management for Food Security; Academic Press: Cambridge, MA, USA, 2016; pp. 613–650. ISBN 9780128032657. [Google Scholar] [CrossRef]
- Cauchie, H.M. Chitin production by arthropods in the hydrosphere. Hydrobiologia 2002, 470, 63–95. [Google Scholar] [CrossRef]
- Klasing, K.C. Comparative Avian Nutrition; Cab International: Wallingford, UK, 1998. [Google Scholar]
- Tajiri, R. Cuticle itself as a central and dynamic player in shaping cuticle. Curr. Opin. Insect Sci. 2017, 19, 30–35. [Google Scholar] [CrossRef]
- Mondotte, J.A.; Saleh, M.C. Antiviral immune response and the route of infection in Drosophila melanogaster. Adv. Virus Res. 2018, 100, 247–278. [Google Scholar]


{kind=link}
| Egg | Larvae | Pupae | Bug | |
|---|---|---|---|---|
| Days | 1–21 | 22–105 | 106–114 | 115–198 |
| Survival Rate % | - | 98.9 | 98.7 | - |
| Larvae (Small) | Larvae (Large) | Moulted | Pre-Moult | Pupae | Cuticle | |
|---|---|---|---|---|---|---|
| Week 8 | ✓ | ✓ | x | x | x | x |
| Week 10 | ✓ | ✓ | x | x | x | x |
| Week 12 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| Week 8 | Week 10 | Week 12 | |
|---|---|---|---|
| Weight | 0.074 ± 0.006 a | 0.084 ± 0.005 a | 0.104 ± 0.007 b |
| Week 8 | Week 10 | Week 12 | |
|---|---|---|---|
| Large | 0.111 ± 0.007 a | 0.121 ± 0.009 ab | 0.139 ± 0.009 b |
| Small | 0.036 ± 0.008 c | 0.046 ± 0.005 c | 0.067 ± 0.015 d |
| Large | Small | |
|---|---|---|
| SA:SB | 1:55 ± 9 a | 1:62 ± 24 a |
| Length (cm) | 2.7 ± 0.20 a | 1.9 ± 0.10 b |
| Width (cm) | 0.4 ± 0.05 a | 0.3 ± 0.05 a |
| Week 8 | Week 10 | Week 12 | |
|---|---|---|---|
| DM | 37.42 ± 1.33 a | 37.27 ± 1.88 a | 36.57 ± 1.47 a |
| Crude protein | 43.35 ± 0.67 a | 44.72 ± 2.16 a | 44.93 ± 1.75 a |
| Crude Fat | 39.47 ± 0.61 a | 38.02 ± 1.68 a | 37.85 ± 1.78 a |
| Crude fibre | 11.30 ± 0.04 | 11.38 ± 0.05 | 11.52 ± 0.06 |
| Chitin | 22.73 ± 1.00 a | 22.25 ± 1.07 a | 21.68 ± 1.03 a |
| Crude ash | 3.64 ± 0.05 a | 3.66 ± 0.06 a | 3.68 ± 0.05 a |
| NFE | 2.29 ± 0.03 a | 2.26 ± 0.13 a | 2.10 ± 0.10 a |
| Week 8 | Week 10 | Week 12 | ||||
|---|---|---|---|---|---|---|
| Large | Small | Large | Small | Large | Small | |
| DM | 38.60 ± 0.30 a | 36.23 ± 0.40 b | 38.53 ± 1.60 a | 36.00 ± 1.22 b | 37.50 ± 1.13 a | 35.63 ± 1.25 b |
| Crude Protein | 43.70 ± 0.50 a | 43.00 ± 0.70 a | 45.23 ± 2.57 a | 44.20 ± 2.06 a | 45.13 ± 2.38 a | 44.73 ± 1.34 a |
| Crude fat | 39.60 ± 0.53 a | 39.33 ± 0.77 a | 38.10 ± 2.00 a | 37.93 ± 1.74 a | 37.80 ± 2.23 a | 37.90 ± 1.71 a |
| Fibre | 11.53 ± 0.61 a | 11.00 ± 0.10 b | 11.83 ± 0.15 a | 10.93 ± 0.25 b | 12.13 ± 0.25 a | 10.90 ± 0.26 b |
| Chitin | 21.93 ± 0.56 b | 23.53 ± 0.50 a | 21.40 ± 0.61 b | 23.10 ± 0.55 a | 20.87 ± 0.58 b | 22.50 ± 0.56 a |
| Crude ash | 3.65 ± 0.08 a | 3.62 ± 0.05 a | 3.64 ± 0.05 a | 3.62 ± 0.05 a | 3.64 ± 0.01 a | 3.60 ± 0.06 a |
| NFE | 1.52 ± 0.03 b | 3.05 ± 0.10 a | 1.2 ± 0.01 b | 3.32 ± 0.09 a | 1.3 ± 0.13 b | 2.87 ± 0.02 a |
| Pre-Moult | Moult | Pupae | Cuticle | |
|---|---|---|---|---|
| DM | 40.15 ± 0.14 b | 39.13 ± 0.98 b | 37.12 ± 0.11 c | 97.5 ± 1.32 a |
| Crude protein | 44.18 ± 0.03 b | 43.97 ± 0.02 b | 44.00 ± 0.08 b | 53.40 ± 0.17 a |
| Crude fat | 39.38 ± 0.03 a | 39.29 ± 0.01 a | 38.25 ± 0.04 b | 12.20 ± 0.13 c |
| Crude fibre | 10.53 ± 0.03 b | 10.82 ± 0.02 b | 11.9 ± 0.06 b | 19.8 ± 0.21 a |
| Chitin | 23.82 ± 0.01 b | 23.65 ± 0.03 b | 23.42 ± 0.04 c | 35.60 ± 0.26 a |
| Ash | 3.66 ± 0.04 b | 3.65 ± 0.02 b | 3.62 ± 0.05 b | 9.8 ± 0.19 a |
| NFE | 2.25 ± 0.03 b | 2.27 ± 0.02 b | 2.23 ± 0.03 b | 4.4 ± 0.20 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toviho, O.A.; Bársony, P. Nutrient Composition and Growth of Yellow Mealworm (Tenebrio molitor) at Different Ages and Stages of the Life Cycle. Agriculture 2022, 12, 1924. https://doi.org/10.3390/agriculture12111924
Toviho OA, Bársony P. Nutrient Composition and Growth of Yellow Mealworm (Tenebrio molitor) at Different Ages and Stages of the Life Cycle. Agriculture. 2022; 12(11):1924. https://doi.org/10.3390/agriculture12111924
Chicago/Turabian StyleToviho, Odunayo A., and Péter Bársony. 2022. "Nutrient Composition and Growth of Yellow Mealworm (Tenebrio molitor) at Different Ages and Stages of the Life Cycle" Agriculture 12, no. 11: 1924. https://doi.org/10.3390/agriculture12111924
APA StyleToviho, O. A., & Bársony, P. (2022). Nutrient Composition and Growth of Yellow Mealworm (Tenebrio molitor) at Different Ages and Stages of the Life Cycle. Agriculture, 12(11), 1924. https://doi.org/10.3390/agriculture12111924