
Assessment of Drought Tolerance of Miscanthus Genotypes through Dry-Down Treatment and Fixed-Soil-Moisture-Content Techniques

 and
and
Abstract
:1. Introduction
well-watered condition) × 100
2. Materials and Methods
2.1. Screening Experiment for Dry-Down-Imposed Drought Stress
2.1.1. Experiment in Hokkaido University, Japan
of drought)/(value of traits on day 0 as well-watered treatment) × 100
2.1.2. Experiment at Brigham Young University, USA
(value of traits of 0-day dry-down) × 100
2.2. Precise-Comparison Experiment with Automated Irrigation System at HU
(value of traits of 30% SMC) × 100
2.3. Post-Drought Recovery in the BYU Experiment
(value of traits of day 21 in BYU screening experiment) × 100
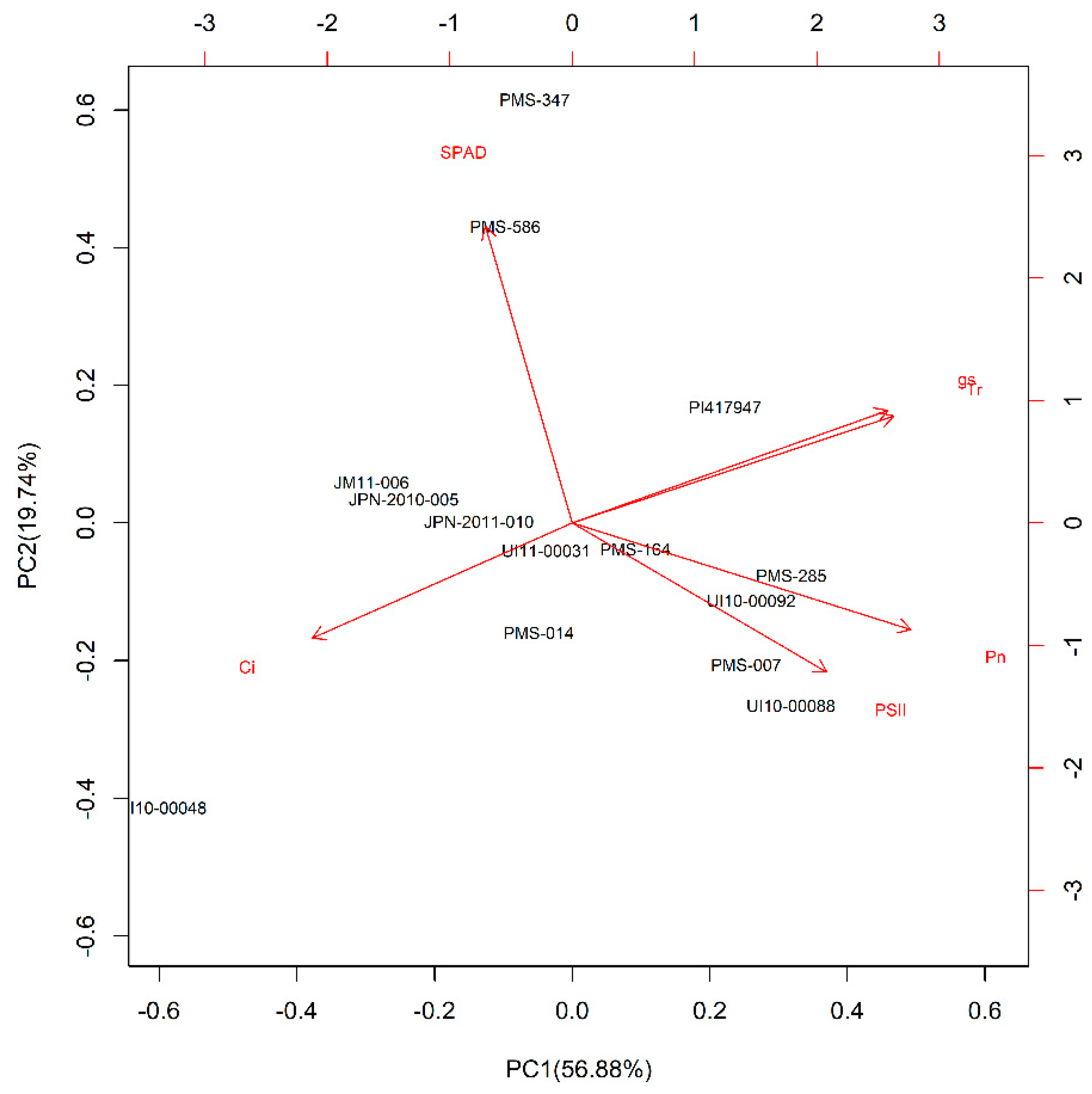
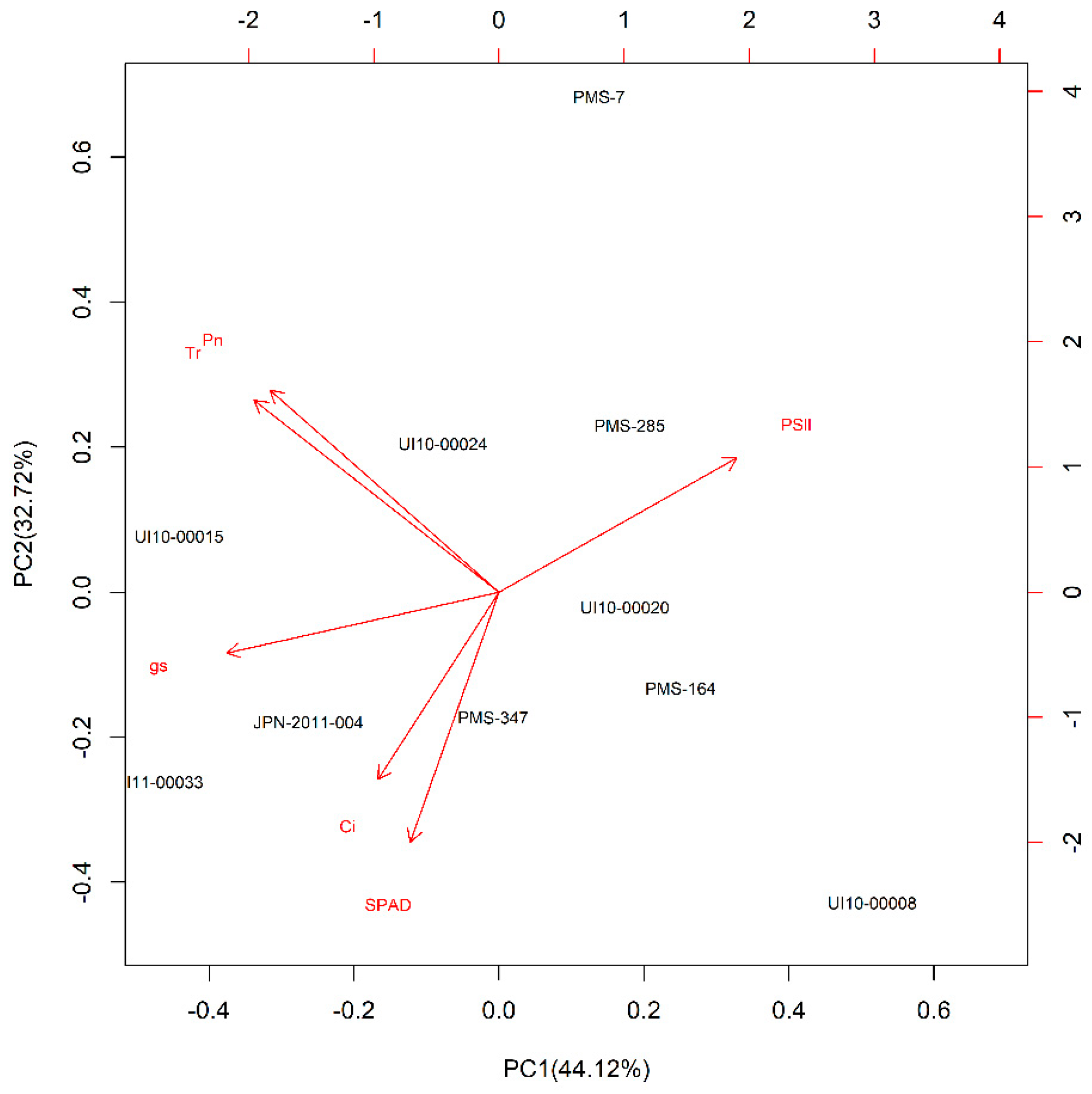
2.4. Drought Tolerance Evaluation and Statistical Analysis
PC1) + (contribution of the second principal components (PC2) (%) × PC2) +
(contribution of the third principal components (PC3) (%) × PC3)
3. Results
3.1. Comparison of Miscanthus Genotype Performance between HU and BYU Experiments
3.2. Changes in Soil Water Potential across Treatments in the BYU Experiment
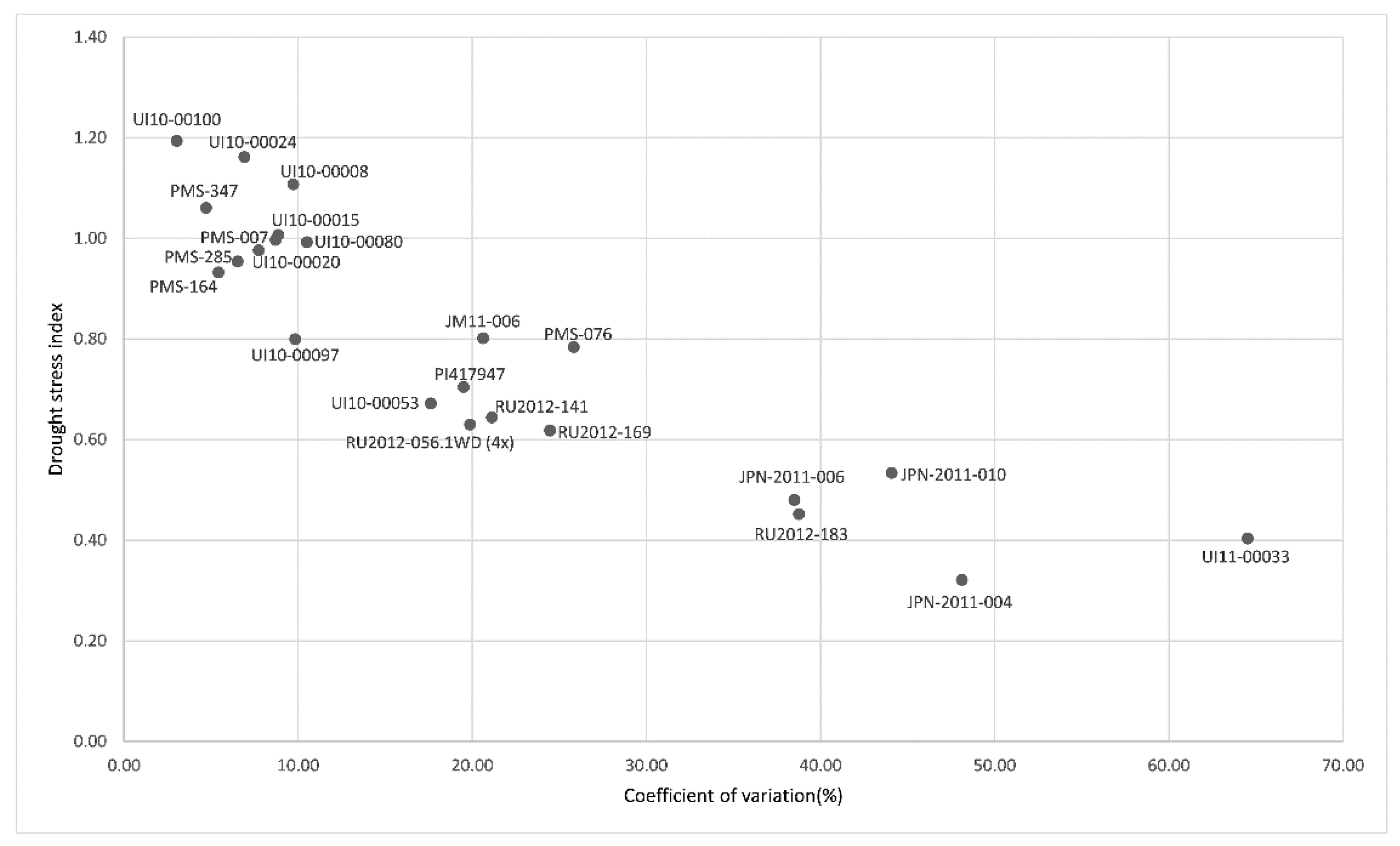
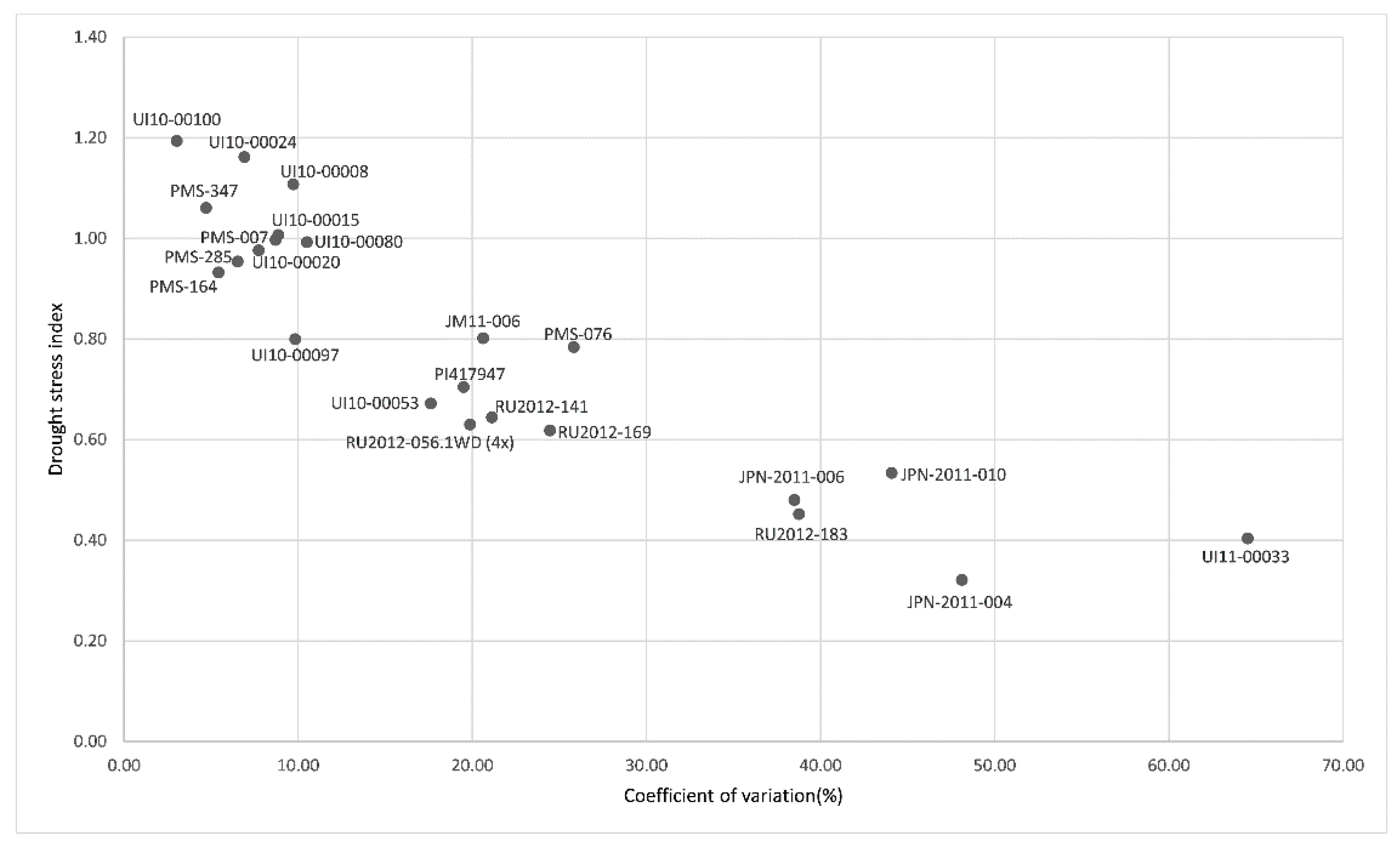
3.3. Performance of Genotypes under Dry-Down Experiment in BYU Screening Experiment
3.4. Performance of Genotypes under Fixed Drought Level with Automated Irrigation System in the HU Precise-Comparison Experiment
3.5. Drought Recovery Capacity of Miscanthus Genotypes of Post-Drought Recovery Experiment in BYU
4. Discussion
4.1. Comparison of Different Drought Treatment Methods for Evaluation
4.2. Characteristics of Drought Stress in Miscanthus spp.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morrow, W.R., III; Gopal, A.; Fitts, G.; Lewis, S.; Dale, L.; Masanet, E. Feedstock loss from drought is a major economic risk for biofuel producers. Biomass Bioenergy 2014, 69, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Deng, X. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, F.; Liu, L.; Zhu, S. Physiological responses of Leucaena leucocephala seedlings to drought stress. Procedia Eng. 2012, 28, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, X.; Tran, H.; Shan, L.; Kim, J.; Childs, K.; Ervin, E.H.; Frazier, T.; Zhao, B. Assessment of drought tolerance of 49 switchgrass (Panicum virgatum) genotypes using physiological and morphological parameters. Biotechnol. Biofuels 2015, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Trenberth, K.E.; Dai, A.; Schrier, G.; van der Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Sala, O.E.; Milchunas, D.G.; Lathrop, R.W. Root dynamics of Bouteloua gracilis during short-term recovery from drought. Funct. Ecol. 1987, 1, 117–124. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Panda, S.K.; Dutta, B.K. CaCl2 improves post-drought recovery potential in Camellia sinensis (L) O. Kuntze. Plant Cell Rep. 2011, 30, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zilinskas, B.A. Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought. Plant J. 1994, 5, 397–405. [Google Scholar] [CrossRef]
- Blackman, C.J.; Li, X.; Choat, B.; Rymer, P.D.; DeKauwe, M.G.; Duursma, R.A.; Tissue, D.A.; Medlyn, B.E. Desiccation time during drought is highly predictable across species of Eucalyptus from contrasting climates. New Phytol. 2019, 224, 632–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergsten, S.J.; Stewart, J.R. Measurement of the influence of low water availability on the productivity of Agave weberi cultivated under controlled irrigation. Can. J. Plant Sci. 2013, 94, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Nemali, K.S.; van Iersel, M.W. An automated system for controlling drought stress and irrigation in potted plants. Sci. Hortic. 2006, 110, 292–297. [Google Scholar] [CrossRef]
- Heaton, E.A.; Dohleman, F.G.; Long, S.P. Meeting US biofuel goals with less land: The potential of Miscanthus. Glob. Chang. Biol. 2008, 14, 2000–2014. [Google Scholar] [CrossRef]
- Toma, Y.; Fernandez, F.G.; Nishuwaki, A.; Yamada, T.; Bollero, G.; Stewart, J.R. Aboveground plant biomass, carbon, and nitrogen dynamics before and after burning in a seminatural grassland of Miscanthus sinensis in Kumamoto, Japan. Glob. Chang. Biol. Bioenergy 2010, 2, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.V.; Brummer, J.E.; Głowacka, K.; Hall, M.C.; Heo, K.; Peng, J.; Yamada, T.; Yoo, J.H.; Yu, C.Y.; Zhao, H.; et al. A footprint of past climate change on the diversity and population structure of Miscanthus sinensis. Ann. Bot. 2014, 114, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.V.; Jin, X.; Petersen, K.K.; Anzoua, K.G.; Bagmet, L.; Chebukin, P.; Deuter, M.; Dzyubenko, E.; Dzyubenko, N.; Heo, K.; et al. Population structure of Miscanthus sacchariflorus reveals two major polyploidization events, tetraploid-mediated unidirectional introgression from diploid M. sinensis, and diversity centred around the Yellow Sea. Ann. Bot. 2019, 124, 731–748. [Google Scholar] [CrossRef]
- Heaton, E.; Voigt, T.; Long, S.P. A quantitative review comparing the yields of two candidate C4 perennial biomass crops in relation to nitrogen, temperature and water. Biomass Bioenergy 2004, 27, 21–30. [Google Scholar] [CrossRef]
- Mann, J.J.; Barney, J.N.; Kyser, G.B.; DiTomaso, J.M. Root system dynamics of Miscanthus × giganteus and Panicum virgatum in response to rainfed and irrigated conditions in California. Bioenergy Res. 2013, 6, 678–687. [Google Scholar] [CrossRef]
- Joo, E.; Hussain, M.Z.; Zeri, M.; Masters, M.D.; Miller, J.N.; Gomez-Casanovas, N.; Bernacchi, C.J. The influence of drought and heat stress on long-term carbon fluxes of bioenergy crops grown in the Midwestern USA. Plant Cell Environ. 2016, 39, 1928–1940. [Google Scholar] [CrossRef]
- Van der Weijde, T.; Huxley, L.M.; Hawkins, S.; Sembiring, E.H.; Farrar, K.; Dolstra, O.; Visser, R.G.F.; Trindade, L.M. Impact of drought stress on growth and quality of miscanthus for biofuel production. Glob. Chang. Biol. Bioenergy 2017, 9, 770–782. [Google Scholar] [CrossRef] [Green Version]
- Vanloocke, A.; Bernacchi, C.J.; Twine, T.E. The impacts of Miscanthus × giganteus production on the Midwest US hydrologic cycle. Glob. Chang. Biol. Bioenergy 2010, 2, 180–191. [Google Scholar] [CrossRef]
- Ings, J.; Mur, L.A.J.; Robson, P.R.H.; Bosch, M. Physiological and growth responses to water deficit in the bioenergy crop Miscanthus × giganteus. Front. Plant Sci. 2013, 4, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerson, R.; Hoover, A.; Ray, A.; Lacey, J.; Cortez, M.; Payne, C.; Karlen, D.; Birrell, S.; Laird, D.; Kallenbach, R.; et al. Drought effects on composition and yield for corn stover, mixed grasses, and Miscanthus as bioenergy feedstocks. Biofuels 2014, 5, 275–291. [Google Scholar] [CrossRef] [Green Version]
- Kato, M.; Kobayashi, K.; Ogiso, E.; Yokoo, M. Photosynthesis and dry-matter production during ripening stage in a female-sterile line of rice. Plant Prod. Sci. 2004, 7, 184–188. [Google Scholar] [CrossRef]
- Takai, T.; Kondo, M.; Yano, M.; Yamamoto, T. A quantitative trait locus for chlorophyll content and its association with leaf photosynthesis in rice. Rice 2010, 3, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Namias, J. Nature and possible causes of the northeastern United States drought during 1962–1965. Mon. Weather. Rev. 1966, 94, 543–554. [Google Scholar] [CrossRef]
- Cornic, G.; Papgeorgiou, I.; Louason, G. Effect of a rapid and a slow drought cycle followed by rehydration on stomatal and non-stomatal components of leaf photosynthesis in Phaseolus vulgaris L. J. Plant Physiol. 1987, 126, 309–318. [Google Scholar] [CrossRef]
- Ganjeali, A.; Porsa, H.; Bagheri, A. Assessment of Iranian chickpea (Cicer arietinum L.) germplasms for drought tolerance. Agric. Water Manag. 2011, 98, 1477–1484. [Google Scholar] [CrossRef]
- Li, C.-N.; Yang, L.-T.; Srivastava, M.K.; Li, Y.-R. Foliar application of abscisic acid improves drought tolerance of sugarcane plant under severe water stress. Int. J. Agric. Innov. Res. 2014, 3, 101–107. [Google Scholar]
- Nazari, L.; Pakniyat, H. Assessment of drought tolerance in barley genotypes. J. Appl. Sci. 2010, 10, 151–156. [Google Scholar] [CrossRef]
- Percival, G.C.; Keary, I.P.; Al-Habsi, S. An assessment of the drought tolerance of Fraxinus genotypes for urban landscape plantings. Urban For. Urban Green. 2006, 5, 17–27. [Google Scholar] [CrossRef]
- Rohollahi, I.; Khoshkholghsima, N.; Nagano, H.; Hoshino, Y.; Yamada, T. Respiratory burst oxidase-D Expression and Biochemical Responses in Festuca arundinacea under Drought Stress. Crop Sci. 2018, 58, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Dracup, J.A.; Lee, K.S.; Paulson, E.G. On the Definition of Droughts. Water Resour. Res. 1980, 16, 297–302. [Google Scholar] [CrossRef]
- Kim, J.; van Iersel, M.W. Slowly developing drought stress increases photosynthetic acclimation of Catharanthus roseus. Physiol. Plant. 2011, 143, 166–177. [Google Scholar] [CrossRef]
- Tamura, K.; Uwatoko, N.; Yamashita, H.; Fujimori, M.; Akiyama, Y.; Shoji, A.; Sanada, Y.; Okumura, K.; Gau, M. Discovery of natural interspecific hybrids between Miscanthus sacchariflorus and Miscanthus sinensis in southern Japan: Morphological characterization, genetic structure, and origin. Bioenergy Res. 2016, 9, 315–325. [Google Scholar] [CrossRef]
- Smith, W.K. Temperatures of desert plants: Another perspective on the adaptability of leaf size. Science 1978, 201, 614–616. [Google Scholar] [CrossRef]
- Jangpromma, N.; Thammasirirak, S.; Jaisil, P.; Songsri, P. Effects of drought and recovery from drought stress on above ground and root growth, and water use efficiency in sugarcane (Saccharum officinarum L.). Aust. J. Crop Sci. 2012, 6, 1298–1304. [Google Scholar]
- Dougherty, R.F.; Quinn, L.D.; Voigt, T.B.; Barney, J.N. Response of naturalized and ornamental biotypes of Miscanthus sinensis to soil-moisture and shade stress. Northeast. Nat. 2015, 22, 372–386. [Google Scholar] [CrossRef]
- Linde-Laursen, I. Cytogenetic analysis of Miscanthus ‘Giganteus’, an interspecific hybrid. Hereditas 1993, 119, 297–300. [Google Scholar] [CrossRef]
- Alvarez, E.; Scheiber, S.M.; Beeson, R.C.; Sandrock, D.R. Drought tolerance responses of purple lovegrass and ‘Adagio’ maiden grass. HortScience 2007, 42, 1695–1699. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Hou, X.; Fan, X.; Wu, J.; Pan, Y. Drought tolerance analysis of Miscanthus sinensis ‘Gracillimu’ seedlings. Acta Prataculturae Sin. 2013, 22, 184–189. [Google Scholar] [CrossRef]
- Stavridou, E.; Webster, R.J.; Robson, P.R.H. Novel Miscanthus genotypes selected for different drought tolerance phenotypes show enhanced tolerance across combinations of salinity and drought treatments. Ann. Bot. 2019, 124, 653–674. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| HU Screening Experiment (2017, 2018) | BYU Screening Experiment (2019) | ||||
|---|---|---|---|---|---|
| Species | Accession | Type | Species | Accession | Type |
| M. sacchariflorus | JM11-006 | Wild | M. sacchariflorus | JM11-006 | Wild |
| M. sacchariflorus | JPN-2011-004 | Wild | M. sacchariflorus | JPN-2010-005 | Wild |
| M. sacchariflorus | JPN-2011-006 | Wild | M. sacchariflorus | JPN-2011-010 | Wild |
| M. sacchariflorus | JPN-2011-010 | Wild | M. sacchariflorus | UI11-00031 | Wild |
| M. sacchariflorus | PMS-076 | Wild | M. sinensis | PMS-007 | Wild |
| M. sacchariflorus | RU2012-056.1WD (4x) | Wild | M. sinensis | PMS-014 | Wild |
| M. sacchariflorus | RU2012-141 | Wild | M. sinensis | PMS-164 | Wild |
| M. sacchariflorus | RU2012-169 | Wild | M. sinensis | PMS-285 | Wild |
| M. sacchariflorus | RU2012-183 | Wild | M. sinensis | PMS-347 | Wild |
| M. sacchariflorus | UI10-00008 | Cultivar | M. sinensis | PMS-586 | Wild |
| M. sacchariflorus | UI11-00033 | Wild | M. sinensis | UI10-00048 | Cultivar |
| M. sinensis | PMS-164 | Wild | M. sinensis | UI10-00088 | Cultivar |
| M. sinensis | PMS-285 | Wild | M. sinensis | UI10-00092 | Wild |
| M. sinensis | PMS-347 | Wild | M. floridulus | PI417947 | Wild |
| M. sinensis | PMS-7 | Wild | |||
| M. sinensis var. condensatus | UI10-00015 | Wild | |||
| M. sinensis | UI10-00020 | Wild | |||
| M. sinensis | UI10-00024 | Cultivar | |||
| M. sinensis | UI10-00053 | Cultivar | |||
| M. sinensis | UI10-00080 | Cultivar | |||
| M. sinensis | UI10-00097 | Cultivar | |||
| M. sinensis | UI10-00100 | Cultivar | |||
| M. floridulus | PI417947 | Wild | |||
| ANOVA | |||||
|---|---|---|---|---|---|
| Source of variation | SS | df | MS | F | p-value |
| Between Groups | 0.0069 | 1 | 0.0069 | 0.1991 | 0.6650 |
| Within Groups | 0.3453 | 10 | 0.0345 | ||
| Total | 0.3522 | 11 | |||
| ANOVA | |||||
|---|---|---|---|---|---|
| Source of variation | SS | df | MS | F | p-value |
| Between Groups | 3.2866 | 1 | 3.2866 | 18.3031 | 0.0002 |
| Within Groups | 5.3870 | 30 | 0.1796 | ||
| Total | 8.6736 | 31 | |||
| Day 0 | Day 7 | Day 14 | Day 21 | Day 7 after Re-Watered | ||
|---|---|---|---|---|---|---|
| Soil Water Potential (mPa) | −0.096 | −0.14 | −2.6025 | −10.25 | 0.04 | |
| Species | Accession | Pn (μmol CO2 m−2 s−1) | ||||
| M. sacchariflorus | JM11-006 | 11.281 | 10.355 | 2.310 | NA | NA |
| M. sacchariflorus | JPN-2010-005 | 7.673 | 9.899 | 2.744 | NA | NA |
| M. sacchariflorus | JPN-2011-010 | 8.087 | 8.930 | 3.927 | NA | NA |
| M. sacchariflorus | UI11-00031 | 12.044 | 12.292 | 7.707 | 5.372 | 8.145 |
| M. floridulus | PI417947 | 3.961 | 5.595 | 3.192 | 3.006 | 2.774 |
| M. sinensis | PMS-007 | 6.268 | 8.006 | 5.824 | 3.488 | 3.367 |
| M. sinensis | PMS-014 | 10.724 | 12.436 | 6.394 | 1.655 | 11.899 |
| M. sinensis | PMS-164 | 6.294 | 11.962 | 3.107 | 7.393 | 7.330 |
| M. sinensis | PMS-285 | 7.613 | 8.364 | 6.810 | 6.484 | 5.121 |
| M. sinensis | PMS-347 | 8.624 | 10.411 | 2.919 | 1.886 | 5.144 |
| M. sinensis | PMS-586 | 5.148 | 9.832 | 2.051 | 1.438 | 6.569 |
| M. sinensis | UI10-00048 | 5.034 | 15.136 | 0.777 | NA | NA |
| M. sinensis | UI10-00088 | 4.418 | 5.081 | 4.312 | 1.160 | 3.275 |
| M. sinensis | UI10-00092 | 5.334 | 13.335 | 5.049 | NA | NA |
| Species | Accession | Origin | Genetic Clusters § | PC1 | PC2 | PC3 | Ranking Value | Rank |
|---|---|---|---|---|---|---|---|---|
| M. sinensis | PMS-285 | China | Yangtze-Qinling Msi | 2.2033 | −0.3101 | 0.4616 | 1.2647 | 1 |
| M. floridulus | PI417947 | Cultivar | SE China Msi | 1.5610 | 0.6848 | 0.1983 | 1.0543 | 2 |
| M. sinensis | UI10-00088 | Cultivar | C Japan Msi | 2.2217 | −1.0798 | 0.0110 | 1.0521 | 3 |
| M. sinensis | UI10-00092 | Cultivar | C Japan Msi | 1.8232 | −0.4588 | 0.4144 | 1.0117 | 4 |
| M. sinensis | PMS-007 | China | Yangtze-Qinling Msi | 1.7714 | −0.8379 | 0.8903 | 0.9825 | 5 |
| M. sinensis | PMS-347 | China | SE China Msi | −0.3521 | 2.5066 | 1.3918 | 0.5140 | 6 |
| M. sinensis | PMS-164 | China | Yangtze-Qinling Msi | 0.6587 | −0.1539 | −1.8284 | 0.0560 | 7 |
| M. sinensis | PMS-586 | China | Sichuan Msi | −0.6486 | 1.7557 | 0.0789 | −0.0098 | 8 |
| M. sacchariflorus | UI11-00031 | China | Yangtze diploids (ssp. lutarioriparius) Msa | −0.2352 | −0.1630 | −0.0439 | −0.1729 | 9 |
| M. sinensis | PMS-014 | China | Sichuan Msi | −0.3133 | −0.6493 | −0.3376 | −0.3596 | 10 |
| M. sacchariflorus | JPN-2011-010 | Japan | N Japan 4x Msa | −0.9108 | 0.0087 | −0.5782 | −0.6075 | 11 |
| M. sacchariflorus | JPN-2010-005 | Japan | N Japan 4x Msa | −1.6686 | 0.1401 | −1.2992 | −1.1262 | 12 |
| M. sacchariflorus | JM11-006 | Japan | S Japan 4x Msa | −1.9928 | 0.2420 | −0.9925 | −1.2422 | 13 |
| M. sinensis | UI10-00048 | Cultivar | S Japan Msi | −4.1179 | −1.6850 | 1.6335 | −2.4174 | 14 |
| Species | Accession | Origin | Genetic Clusters § | Leaf Width (cm) | Leaf Length (cm) | PC1 | PC2 | PC3 | Ranking Value | Rank |
|---|---|---|---|---|---|---|---|---|---|---|
| M. sinensis | PMS-007 | China | Yangtze-Qinling Msi | 2.0 | 60 | 0.71 | 3.03 | −0.57 | 1.232 | 1 |
| M. sinensis | PMS-285 | China | Yangtze-Qinling Msi | 1.1 | 56 | 0.93 | 1.02 | 0.04 | 0.749 | 2 |
| M. sacchariflorus | UI10-00008 | Cultivar | NEChina/Korea/Russia diploids Msa | 0.8 | 44 | 2.65 | −1.90 | 0.55 | 0.618 | 3 |
| M. sinensis | UI10-00020 | Cultivar | S Japan Msi | 0.4 | 18 | 0.89 | −0.09 | 0.68 | 0.450 | 4 |
| M. sinensis | PMS-164 | China | Yangtze-Qinling Msi | 1.1 | 25 | 1.29 | −0.58 | −0.36 | 0.333 | 5 |
| M. sinensis | UI10-00024 | Cultivar | S Japan Msi | 0.6 | 27 | −0.40 | 0.90 | 0.47 | 0.178 | 6 |
| M. sinensis | PMS-347 | China | SE China Msi | 1.8 | 48 | −0.04 | −0.76 | −0.52 | −0.333 | 7 |
| M. sinensis var. condensatus | UI10-00015 | Cultivar | C Japan Msi | 1.6 | 40 | −2.27 | 0.34 | 1.39 | −0.712 | 8 |
| M. sacchariflorus | JPN-2011-004 | Japan | S Japan 4x Msa | 1.8 | 55 | −1.35 | −0.79 | −1.80 | −1.085 | 9 |
| M. sacchariflorus | UI11-00033 | Japan | S Japan 4x Msa | 2.0 | 61 | −2.41 | −1.16 | 0.14 | −1.425 | 10 |
| Species | Accession | Origin | Genetic Clusters ‡ | PC1 | PC2 | PC3 | Ranking Value | Rank |
|---|---|---|---|---|---|---|---|---|
| M. sinensis | PMS-014 | China | Sichuan Msi | 4.3565 | −1.4954 | 0.2379 | 2.8970 | 1 |
| M. sinensis | PMS-586 | China | Sichuan Msi | 3.3939 | 0.4381 | 0.5461 | 2.5689 | 2 |
| M. sinensis | PMS-347 | China | SE China Msi | 2.7456 | 1.2399 | −1.0582 | 2.1484 | 3 |
| M. sinensis | UI10-00088 | Cultivar | C Japan Msi | 1.2360 | −0.6833 | 0.0990 | 0.7769 | 4 |
| M. sacchariflorus | UI11-00031 | China | Yangtze diploids (ssp. lutarioriparius) Msa | −0.2981 | 0.8091 | 0.6621 | −0.0294 | 5 |
| M. floridulus | PI417947 | Cultivar | SE China Msi | −0.9572 | 2.3607 | 0.7977 | −0.2173 | 6 |
| M. sinensis | PMS-164 | China | Yangtze-Qinling Msi | −0.5758 | 0.7901 | −0.0801 | −0.2786 | 7 |
| M. sinensis | PMS-007 | China | Yangtze-Qinling Msi | −0.5925 | −0.3455 | −0.4199 | −0.5167 | 8 |
| M. sinensis | PMS-285 | China | Yangtze-Qinling Msi | −0.9592 | −0.8056 | 0.0559 | −0.8369 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, T.-Y.; Nakashima, T.; Villanueva-Morales, A.; Stewart, J.R.; Sacks, E.J.; Yamada, T. Assessment of Drought Tolerance of Miscanthus Genotypes through Dry-Down Treatment and Fixed-Soil-Moisture-Content Techniques. Agriculture 2022, 12, 6. https://doi.org/10.3390/agriculture12010006
Weng T-Y, Nakashima T, Villanueva-Morales A, Stewart JR, Sacks EJ, Yamada T. Assessment of Drought Tolerance of Miscanthus Genotypes through Dry-Down Treatment and Fixed-Soil-Moisture-Content Techniques. Agriculture. 2022; 12(1):6. https://doi.org/10.3390/agriculture12010006
Chicago/Turabian StyleWeng, Tzu-Ya, Taiken Nakashima, Antonio Villanueva-Morales, J. Ryan Stewart, Erik J. Sacks, and Toshihiko Yamada. 2022. "Assessment of Drought Tolerance of Miscanthus Genotypes through Dry-Down Treatment and Fixed-Soil-Moisture-Content Techniques" Agriculture 12, no. 1: 6. https://doi.org/10.3390/agriculture12010006
APA StyleWeng, T.-Y., Nakashima, T., Villanueva-Morales, A., Stewart, J. R., Sacks, E. J., & Yamada, T. (2022). Assessment of Drought Tolerance of Miscanthus Genotypes through Dry-Down Treatment and Fixed-Soil-Moisture-Content Techniques. Agriculture, 12(1), 6. https://doi.org/10.3390/agriculture12010006