
Antifungal Activities of Ageratum conyzoides L. Extract against Rice Pathogens Pyricularia oryzae Cavara and Rhizoctonia solani Kühn


 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and General Methods
2.2. Extraction and Solvent Partition of A. conyzoides
2.3. Isolation of Antifungal Compounds from ACE
2.4. Assay for Inhibitory Activities against Mycelial Growth of P. oryzae and R. solani In Vitro
2.5. Identification of Antifungal Compounds
2.6. Field Trial Test for ACE on Suppressing Rice Blast Disease
2.7. Statistical Analysis
3. Results
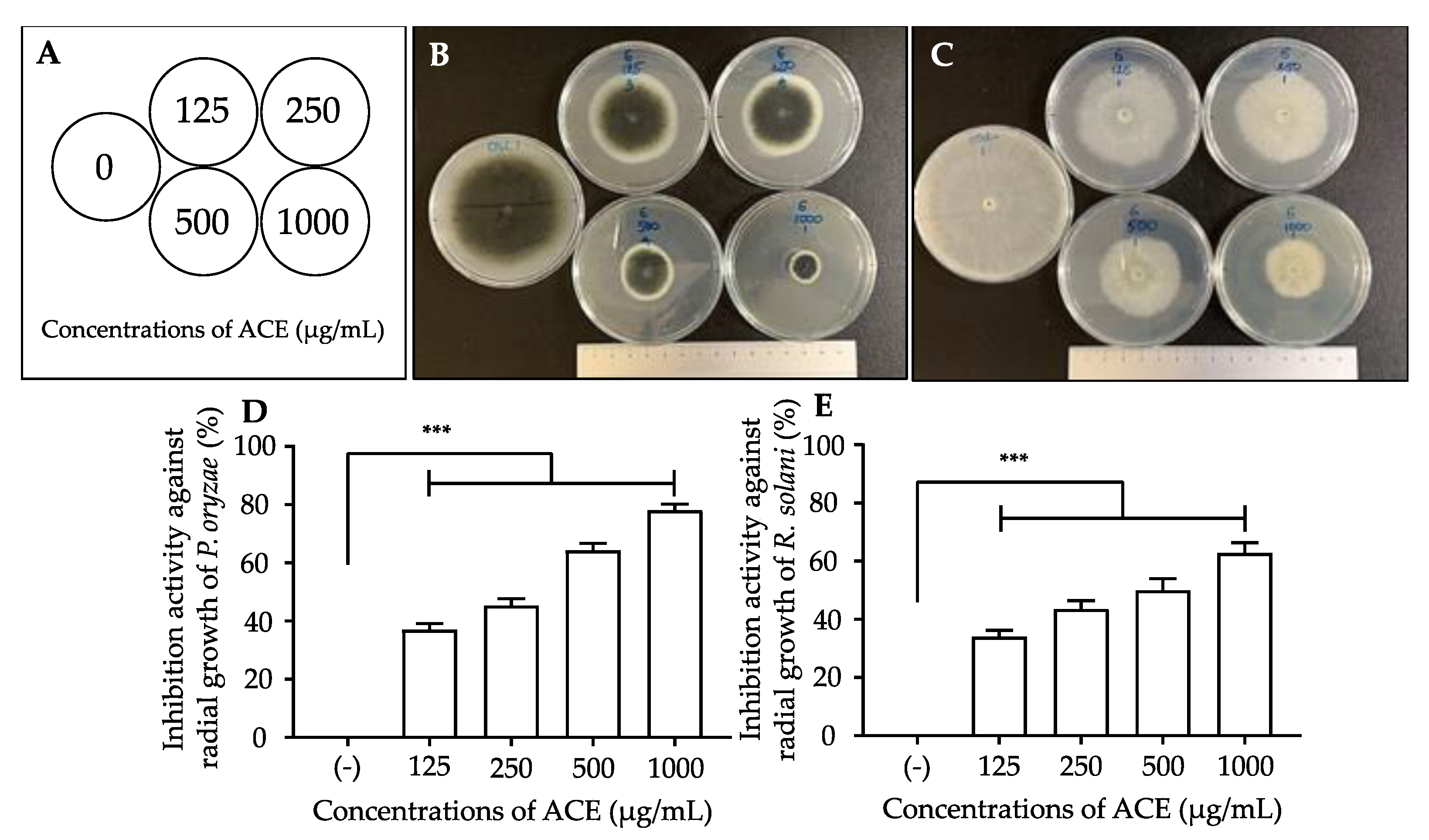
3.1. Effects of A. conyzoides Ethanol Extract (ACE) on the Mycelial Growth of P. oryzae and R. solani
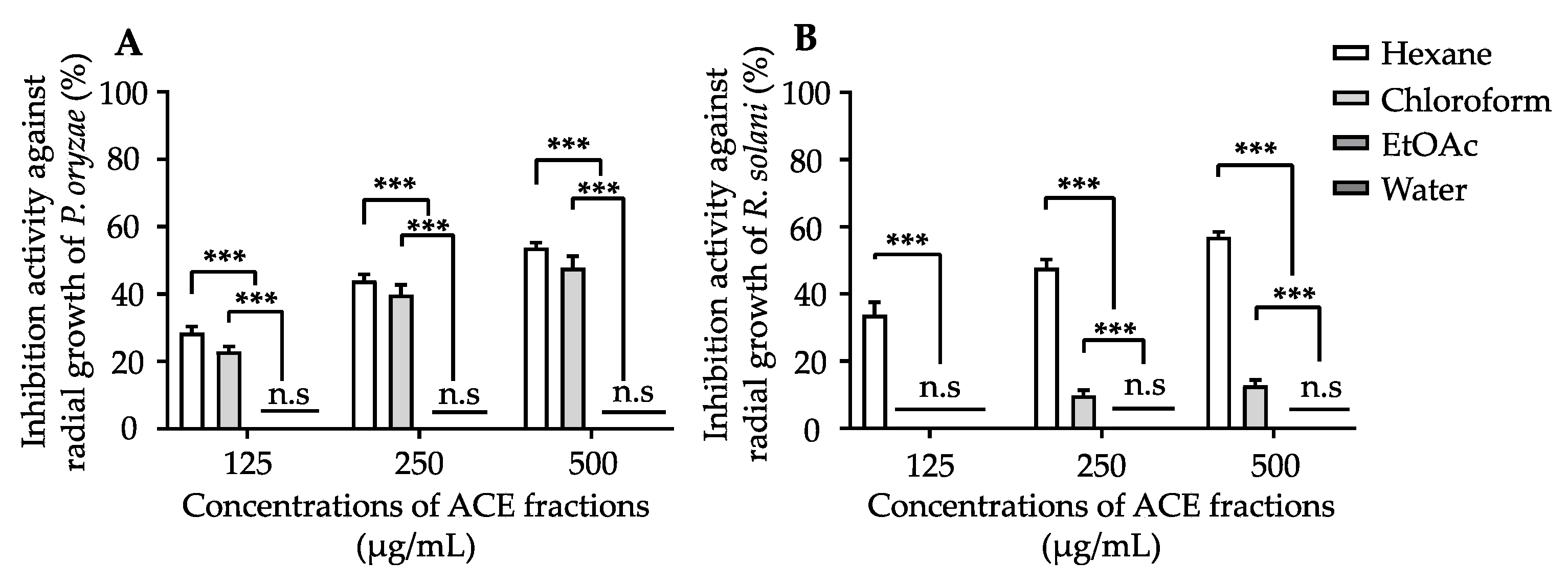
3.2. Isolation of Antifungal Compounds from ACE
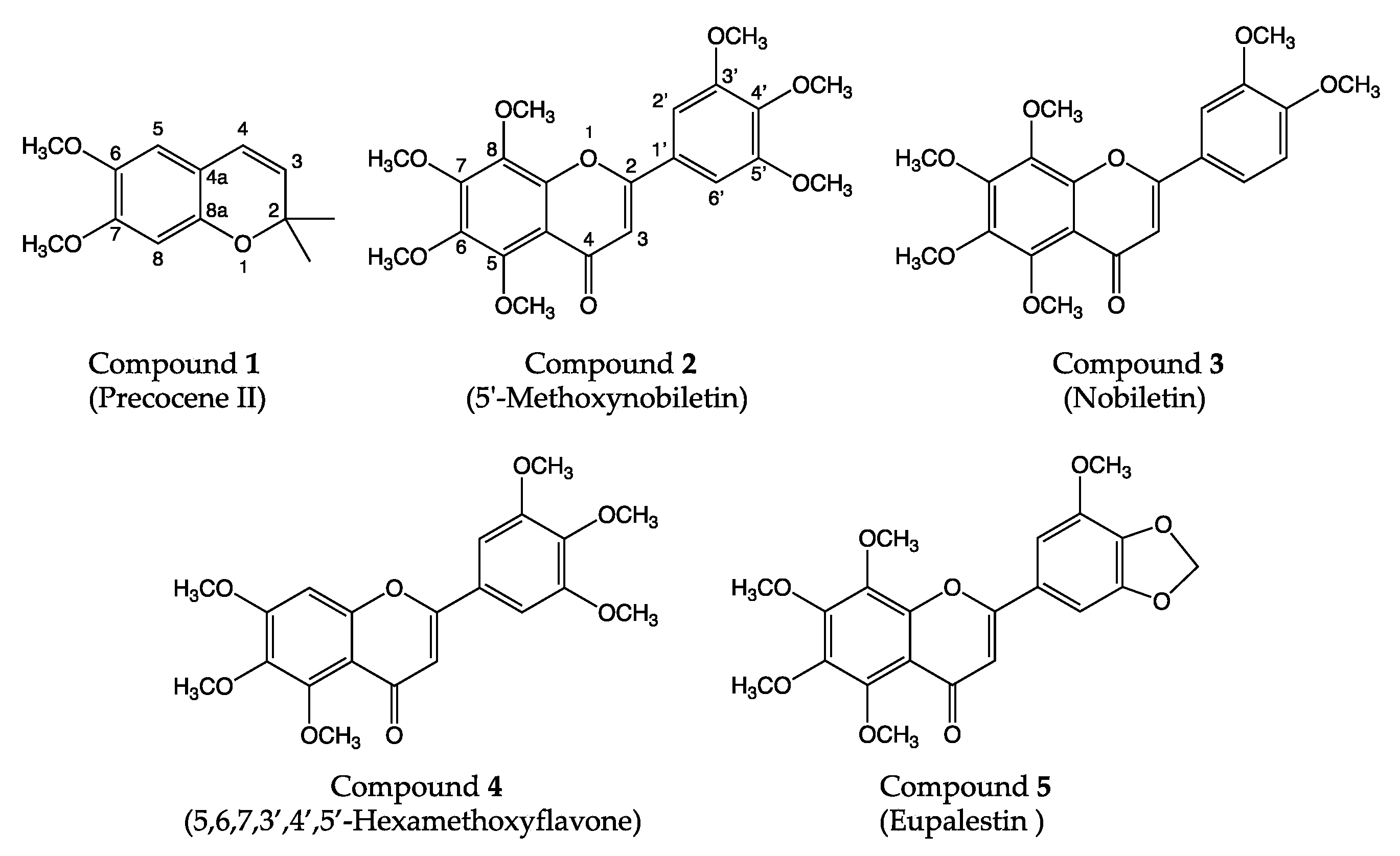
3.3. Identification of Antifungal Compounds
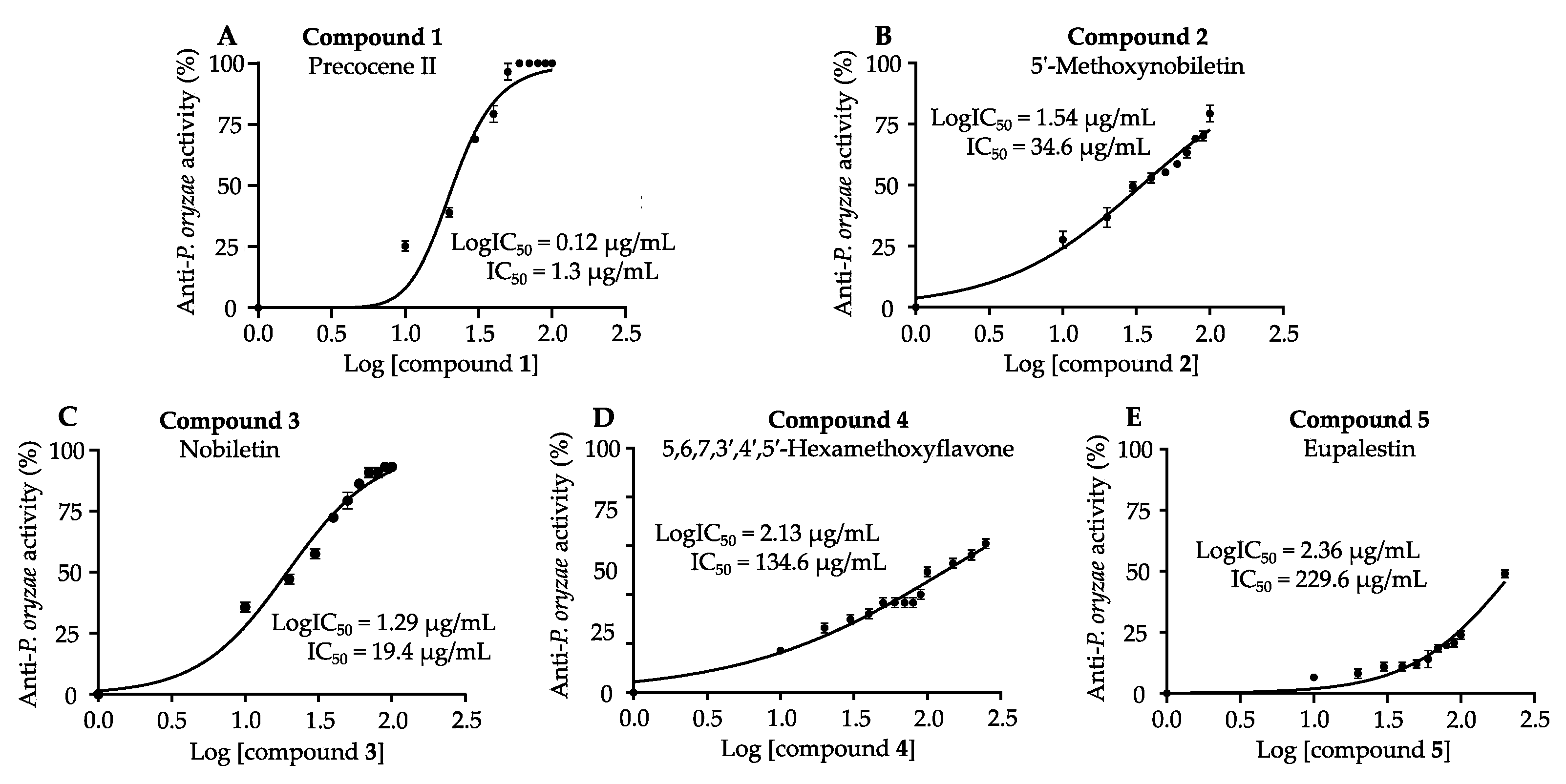
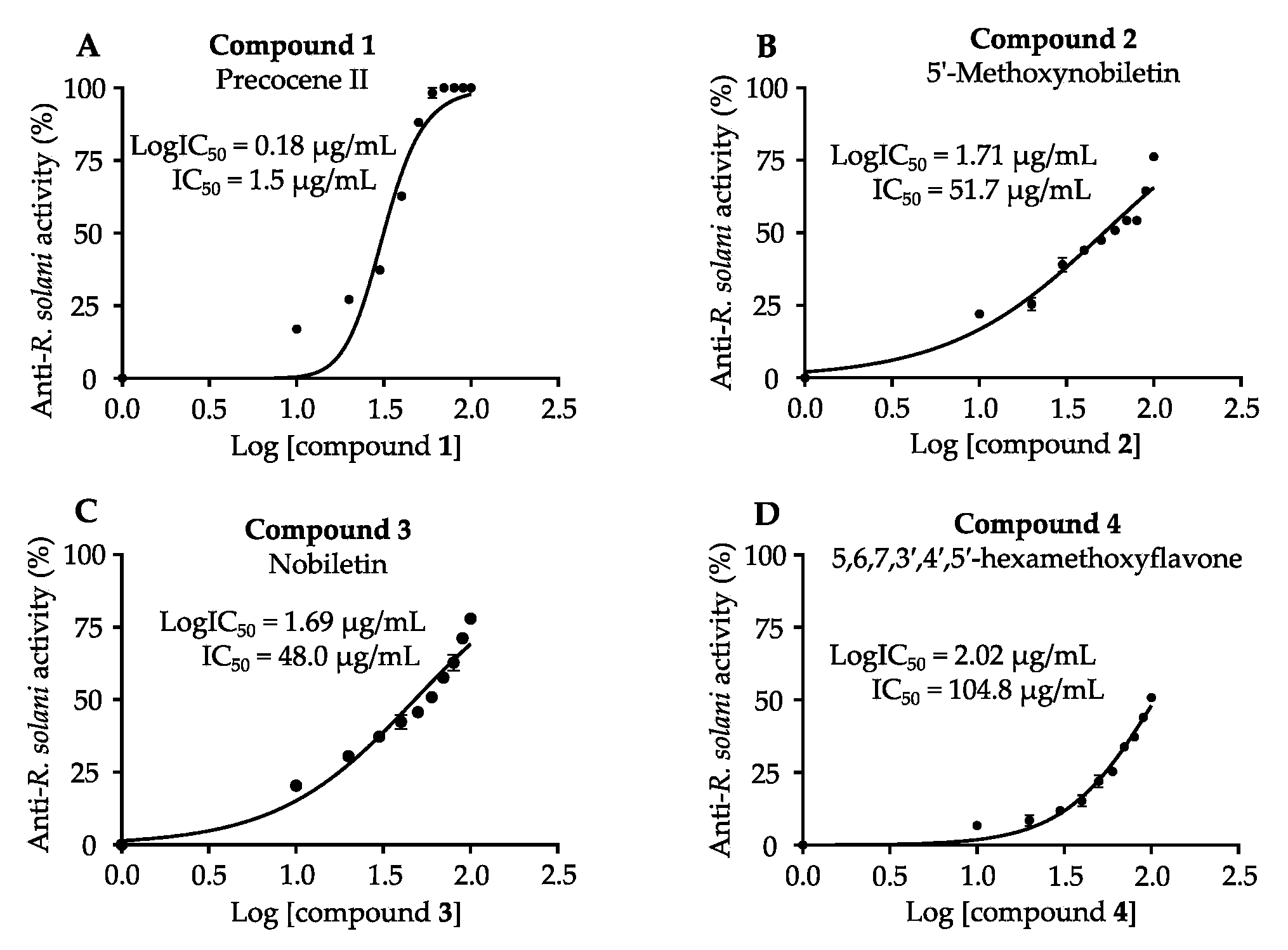
3.4. Antifungal Activities of Isolated Compounds against P. oryzae and R. solani
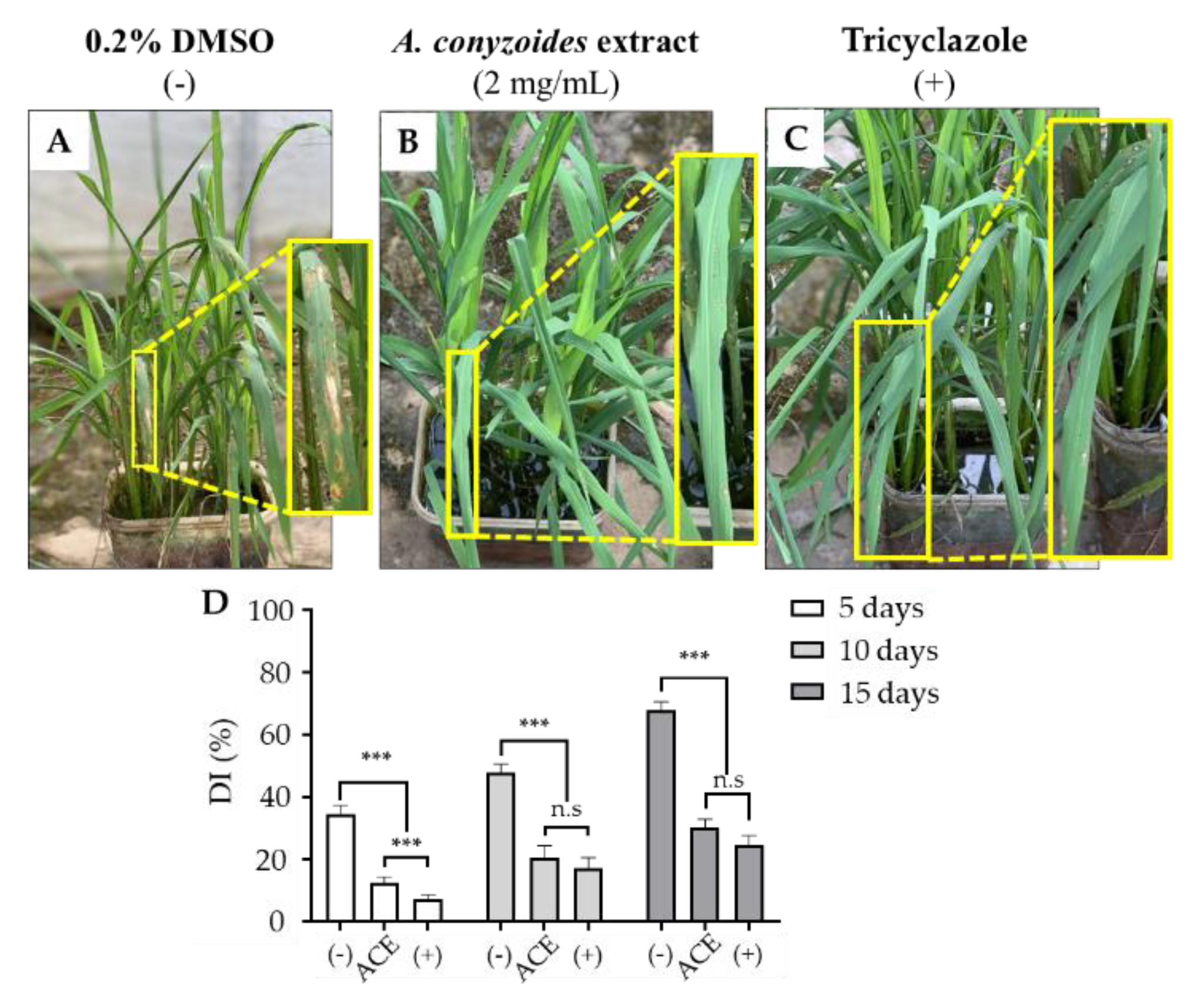
3.5. Field Trial Test for Protective Effect of ACE on Rice Crop against P. oryzae Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, N.; Xu, R.; Duan, P.; Li, Y. Control of grain size in rice. Plant Reprod. 2018, 31, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Gross, B.L.; Zhao, Z. Archaeological and genetic insights into the origins of domesticated rice. Proc. Natl. Acad. Sci. USA 2014, 111, 6190–6197. [Google Scholar] [CrossRef] [PubMed]
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for global nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef]
- Ou, S.H. Rice Diseases, 2nd ed.; Commonwealth Mycological Institute: Egham, UK, 1985; ISBN 0-85198-545-9. [Google Scholar]
- Jiang, N.; Yan, J.; Liang, Y.; Shi, Y.; He, Z.; Wu, Y.; Zeng, Q.; Liu, X.; Peng, J. Resistance Genes and their Interactions with Bacterial Blight/Leaf Streak Pathogens (Xanthomonas oryzae) in Rice (Oryza sativa L.)—An Updated Review. Rice 2020, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Groth, D.E. Rice Diseases and Disorders in Louisiana; LSU Agricultural Experiment Station Reports; LSU Agricultural Center: Baton Rouge, LA, USA, 1991. [Google Scholar]
- Shahriar, S.A.; Imtiaz, A.A.; Hossain, M.B.; Husna, A.; Eaty, M.N.K. Review: Rice Blast Disease. Annu. Res. Rev. Biol. 2020, 35, 50–64. [Google Scholar] [CrossRef]
- Asibi, A.E.; Chai, Q.; Coulter, J.A. Rice blast: A disease with implications for global food security. Agronomy 2019, 9, 451. [Google Scholar] [CrossRef]
- Ashik Iqbal Khan, M.; Rejwan Bhuiyan, M.; Hossain, M.S.; Pratim Sen, P.; Ara, A.; Abubakar Siddique, M.; Ansar Ali, M. Neck blast disease influences grain yield and quality traits of aromatic rice. Comptes Rendus Biol. 2014, 337, 635–641. [Google Scholar] [CrossRef]
- Law, J.W.F.; Ser, H.L.; Khan, T.M.; Chuah, L.H.; Pusparajah, P.; Chan, K.G.; Goh, B.H.; Lee, L.H. The potential of streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef]
- Xia, Y.; Fei, B.; He, J.; Zhou, M.; Zhang, D.; Pan, L.; Li, S.; Liang, Y.; Wang, L.; Zhu, J.; et al. Transcriptome analysis reveals the host selection fitness mechanisms of the Rhizoctonia solani AG1IA pathogen. Sci. Rep. 2017, 7, 10120. [Google Scholar] [CrossRef]
- Anderson, J.P.; Sperschneider, J.; Win, J.; Kidd, B.; Yoshida, K.; Hane, J.; Saunders, D.G.O.; Singh, K.B. Comparative secretome analysis of Rhizoctonia solani isolates with different host ranges reveals unique secretomes and cell death inducing effectors. Sci. Rep. 2017, 7, 10410. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Elazegui, F.A.; Castilla, N.P.; Teng, P.S. Rice pest constraints in tropical Asia: Quantification of yield losses due to rice pests in a range of production situations. Plant Dis. 2000, 84, 357–369. [Google Scholar] [CrossRef]
- Jamali, H.; Sharma, A.; Roohi; Srivastava, A.K. Biocontrol potential of Bacillus subtilis RH5 against sheath blight of rice caused by Rhizoctonia solani. J. Basic Microbiol. 2020, 60, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Li, Z.; Liu, J.; Shen, Z.; Gao, G.; Zhang, Q.; He, Y. Development and evaluation of improved lines with broad-spectrum resistance to rice blast using nine resistance genes. Rice 2019, 12, 29. [Google Scholar] [CrossRef]
- Hubert, J.; Mabagala, R.B.; Mamiro, D.P. Efficacy of Selected Plant Extracts against Pyricularia grisea. It is Causal Agent of Rice Blast Disease. Am. J. Plant Sci. 2015, 06, 602–611. [Google Scholar] [CrossRef]
- Khoa, N.D.; Thúy, P.T.H.; Thuy, T.T.T.; Collinge, D.B.; Jørgensen, H.J.L. Disease-reducing effect of Chromolaena odorata extract on sheath blight and other rice diseases. Phytopathology 2011, 101, 231–240. [Google Scholar] [CrossRef]
- Miller, J.D. Fungi and mycotoxins in grains: Implication for stored products research. J. Stored Prod. Res. 1995, 31, 1–16. [Google Scholar] [CrossRef]
- Ardalani, H.; Anam, S.; Kromphardt, K.J.K.; Staerk, D.; Kongstad, K.T. Coupling Microplate-Based Antibacterial Assay with Liquid Chromatography for High-Resolution Growth Inhibition Profiling of Crude Extracts: Validation and Proof-of-Concept Study with Staphylococcus aureus. Molecules 2021, 26, 1550. [Google Scholar] [CrossRef]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef]
- Gwinn, K.D. Bioactive Natural Products in Plant Disease Control, 1st ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2018; Volume 56, ISBN 9780444640581. [Google Scholar]
- Hao, L.; Wang, Y.; Chen, X.; Zheng, X.; Chen, S.; Li, S.; Zhang, Y.; Xu, Y. Exploring the potential of natural products from mangrove rhizosphere bacteria as biopesticides against plant diseases. Plant Dis. 2019, 103, 2925–2932. [Google Scholar] [CrossRef]
- Naqvi, S.A.H.; Umar, U.; Hasnain, A.; Rehman, A.; Perveen, R. Effect of Botanical Extracts: A Potential Biocontrol Agent for Xanthomonas oryzae pv. oryzae, Causing Bacterial Leaf Blight Disease of Rice. Pakistan J. Agric. Res. 2018, 32, 59–72. [Google Scholar] [CrossRef]
- Govindappa, M.; Umesha, S.; Lokesh, S. Adhatoda vasica leaf extract induces resistance in rice against bacterial leaf blight disease (Xanthomonas oryzae pv. oryzae). Int. J. Plant Physiol. Biochem. 2011, 3, 6–14. [Google Scholar]
- Amadioha, A.C. Controlling rice blast in vitro and in vivo with extracts of Azadirachta indica. Crop Prot. 2000, 19, 287–290. [Google Scholar] [CrossRef]
- Okunade, A.L. Ageratum conyzoides L. (Asteraceae). Fitoterapia 2002, 73, 1–16. [Google Scholar] [CrossRef]
- Kotta, J.C.; Lestari, A.B.S.; Candrasari, D.S.; Hariono, M. Medicinal Effect, in Silico Bioactivity Prediction, and Pharmaceutical Formulation of Ageratum conyzoides L.: A Review. Scientifica 2020, 2020, 6420909. [Google Scholar] [CrossRef] [PubMed]
- Ngo, M.T.; Han, J.W.; Yoon, S.; Bae, S.; Kim, S.Y.; Kim, H.; Choi, G.J. Discovery of new triterpenoid saponins isolated from maesa japonica with antifungal activity against rice blast fungus Magnaporthe oryzae. J. Agric. Food Chem. 2019, 67, 7706–7715. [Google Scholar] [CrossRef]
- Lee, Y. Lysimachia foenum-graecum herba extract, a novel biopesticide, inhibits ABC transporter genes and mycelial growth of Magnaporthe oryzae. Plant Pathol. J. 2016, 32, 8–15. [Google Scholar] [CrossRef]
- Mehta, V.; Kumar, S. Influence of different plant powders as grain protectants on Sitophilus oryzae (L.) (Coleoptera: Curculionidae) in stored wheat. J. Food Prot. 2020, 83, 2167–2172. [Google Scholar] [CrossRef]
- Kamboj, A.; Saluja, A. Ageratum conyzoides L.: A review on its phytochemical and pharmacological profile. Int. J. Green Pharm. 2008, 2, 59. [Google Scholar] [CrossRef]
- Ming, L.C. Ageratum conyzoides: A Tropical Source of Medicinal and Agricultural Products. In Perspectives on New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 1999; pp. 469–473. [Google Scholar]
- Yang, C.; Hamel, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of Action and Possible Impact on Nontarget Microorganisms. ISRN Ecol. 2011, 2011, 1–8. [Google Scholar] [CrossRef]
- Upadhyay, O.P.; Kumar, K.; Tiwari, R.K. Ethnobotanical study of skin treatment uses of medicinal plants of Bihar. Pharm. Biol. 1998, 36, 167–172. [Google Scholar] [CrossRef]
- Chauhan, A.; Rijhwani, S. A comprehensive review on phytochemistry of Ageratum conyzoides Linn. (Goat weed). Int. J. Eng. Technol. Manag. Appl. Sci. 2015, 3, 348–358. [Google Scholar]
- Kong, C.; Hu, F.; Xu, X.; Zhang, M.; Liang, W. Volatile allelochemicals in the Ageratum conyzoides intercropped citrus orchard and their effects on mites Amblyseius newsami and Panonychus citri. J. Chem. Ecol. 2005, 31, 2193–2203. [Google Scholar] [CrossRef]
- Kong, C.; Hu, F.; Xu, X. Allelopathic potential and chemical constituents of volatiles from Ageratum conyzoides under stress. J. Chem. Ecol. 2002, 28, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Astiti1, N.P.A.; Suprapta, D.N. Antifungal Activity of Teak (Tectona grandis L. F.) Leaf Extract Against Arthrinium phaeosperum (Corda) M.B. Ellis. The Cause of Wood decay on Albizia falcataria (L.) Fosberg. J. Int. Soc. Southeast Asian Agric. Sci. 2012, 18, 62–69. [Google Scholar]
- IRRI. Standard Evaluation System for Rice; International Rice Research Institute: Los Baños, Philippines, 2013; 55p. [Google Scholar]
- González, A.G.; Aguiar, Z.E.; Grillo, T.A.; Luis, J.G.; Rivera, A.; Calle, J. Chromenes from Ageratum conyzoides. Phytochemistry 1991, 30, 1137–1139. [Google Scholar] [CrossRef]
- Le-Van, N.; Van Cuong Pham, T. Two new flavones from Eupatorium coelestinum. Phytochemistry 1979, 18, 1859–1861. [Google Scholar] [CrossRef]
- Nour, A.M.M.; Khalid, S.A.; Kaiser, M.; Brun, R.; Abdalla, W.E.; Schmidt, T.J. The antiprotozoal activity of methylated flavonoids from Ageratum conyzoides L. J. Ethnopharmacol. 2010, 129, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Montanari, A.M.; Widmer, W.W. Two New Polymethoxylated Flavones, a Class of Compounds with Potential Anticancer Activity, Isolated from Cold Pressed Dancy Tangerine Peel Oil Solids. J. Agric. Food Chem. 1997, 45, 364–368. [Google Scholar] [CrossRef]
- Chen, C.C.; Chen, Y.P.; Hsu, H.Y.; Chen, Y.L. New Flavones from Bauhinia championii BENTH. Chem. Pharm. Bull. 1984, 32, 166–169. [Google Scholar] [CrossRef]
- Javed, S.; Bashir, U. Antifungal activity of different extracts of Ageratum conyzoides for the management of Fusarium solani. Afr. J. Biotechnol. 2012, 11, 11022–11029. [Google Scholar] [CrossRef]
- Iqbal, M.C.M.; Jayasinghe, U.L.B.; Herath, H.M.T.B.; Wijesekara, K.B.; Fujimoto, Y. A fungistatic chromene from Ageratum conyzoides. Phytoparasitica 2004, 32, 119–126. [Google Scholar] [CrossRef]
- Esper, R.H.; Gonçalez, E.; Felicio, R.C.; Felicio, J.D. Fungicidal activity and constituents of Ageratum conyzoides essential oil from three regions in São Paulo state, Brazil. Arq. Inst. Biol. 2015, 82, 1–4. [Google Scholar] [CrossRef]
- Ortuño, A.; Del Rio, J.A. Role of Citrus Phenolic Compounds in the Resistance Mechanism against Pathogenic Fungi. Tree For. Sci. Biotechnol. 2009, 3, 49–53. [Google Scholar]
- Moreira, M.D.; Picanço, M.C.; Barbosa, L.C.A.; Guedes, R.N.C.; Barros, E.C.; Campos, M.R. Compounds from Ageratum conyzoides: Isolation, structural elucidation and insecticidal activity. Pest Manag. Sci. 2007, 63, 615–621. [Google Scholar] [CrossRef]
- Kong, C.; Liang, W.; Hu, F.; Xu, X.; Wang, P.; Jiang, Y.; Xing, B. Allelochemicals and their transformations in the Ageratum conyzoides intercropped citrus orchard soils. Plant Soil 2004, 264, 149–157. [Google Scholar] [CrossRef]
- Wu, T.; Cheng, D.; He, M.; Pan, S.; Yao, X.; Xu, X. Antifungal action and inhibitory mechanism of polymethoxylated flavones from Citrus reticulata Blanco peel against Aspergillus niger. Food Control 2014, 35, 354–359. [Google Scholar] [CrossRef]
- Almada-Ruiz, E.; Martínez-Téllez, M.Á.; Hernández-Álamos, M.M.; Vallejo, S.; Primo-Yúfera, E.; Vargas-Arispuro, I. Fungicidal potential of methoxylated flavones from citrus for in vitro control of Colletotrichum gloeosporioides, causal agent of anthracnose disease in tropical fruits. Pest Manag. Sci. 2003, 59, 1245–1249. [Google Scholar] [CrossRef]
- Tomas-Barberán, F.A.; Msonthi, J.D.; Hostettmann, K. Antifungal epicuticular methylated flavonoids from Helichrysum nitens. Phytochemistry 1988, 27, 753–755. [Google Scholar] [CrossRef]
- Santra, H.K.; Banerjee, D. Natural Products as Fungicide and Their Role in Crop Protection. In Natural Bioactive Products in Sustainable Agriculture; Singh, J., Yadav, A.N., Eds.; Springer: Singapore, 2020; pp. 131–219. ISBN 978-981-15-3023-4. [Google Scholar]
- Chahal, R.; Nanda, A.; Akkol, E.K.; Sobarzo-sánchez, E.; Arya, A.; Kaushik, D.; Dutt, R.; Bhardwaj, R.; Rahman, M.H.; Mittal, V. Ageratum conyzoides L. and its secondary metabolites in the management of different fungal pathogens. Molecules 2021, 26, 2933. [Google Scholar] [CrossRef]
- Bioscience Solutions-Indepent Crop Protection R&D Consulting. Available online: https://biocomm.eu/2017/12/17/fungicide-mode-action-labcoat-guide-pesticides-biopesticides/ (accessed on 11 November 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (µg/mL) | |
|---|---|---|
| Anti-P. oryzae | Anti-R. solani | |
| 1. Precocene II | 1.3 ± 0.02 | 1.5 ± 0.2 |
| 2. 5′-Methoxynobiletin | 34.6 ± 1.5 | 51.7 ± 1.7 |
| 3. Nobiletin | 19.4 ± 1.3 | 48.0 ± 1.5 |
| 4. 5,6,7,3′,4′,5′-Hexamethoxyflavone | 134.6 ± 2.1 | 104.8 ± 2.0 |
| 5. Eupalestin | 229.6 ± 2.3 | N.D. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, C.C.; Nguyen, T.Q.C.; Kanaori, K.; Binh, T.D.; Dao, X.H.T.; Vang, L.V.; Kamei, K. Antifungal Activities of Ageratum conyzoides L. Extract against Rice Pathogens Pyricularia oryzae Cavara and Rhizoctonia solani Kühn. Agriculture 2021, 11, 1169. https://doi.org/10.3390/agriculture11111169
Nguyen CC, Nguyen TQC, Kanaori K, Binh TD, Dao XHT, Vang LV, Kamei K. Antifungal Activities of Ageratum conyzoides L. Extract against Rice Pathogens Pyricularia oryzae Cavara and Rhizoctonia solani Kühn. Agriculture. 2021; 11(11):1169. https://doi.org/10.3390/agriculture11111169
Chicago/Turabian StyleNguyen, Cuong C., Thanh Q. C. Nguyen, Kenji Kanaori, Tran Duy Binh, Xuyen H. T. Dao, Le Van Vang, and Kaeko Kamei. 2021. "Antifungal Activities of Ageratum conyzoides L. Extract against Rice Pathogens Pyricularia oryzae Cavara and Rhizoctonia solani Kühn" Agriculture 11, no. 11: 1169. https://doi.org/10.3390/agriculture11111169
APA StyleNguyen, C. C., Nguyen, T. Q. C., Kanaori, K., Binh, T. D., Dao, X. H. T., Vang, L. V., & Kamei, K. (2021). Antifungal Activities of Ageratum conyzoides L. Extract against Rice Pathogens Pyricularia oryzae Cavara and Rhizoctonia solani Kühn. Agriculture, 11(11), 1169. https://doi.org/10.3390/agriculture11111169