
Bacillus velezensis T149-19 and Bacillus safensis T052-76 as Potential Biocontrol Agents against Foot Rot Disease in Sweet Potato

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Fungal Strains
2.2. Bacterial DNA Extraction
2.3. Whole-Genome Sequencing (WGS), De Novo Genome Assembly and Sequence Analyses
2.4. Search for Antimicrobial Substances Coding Genes
2.5. Pot Experiments in Greenhouse
2.6. Plant Health Analyses
2.7. DNA Extraction for Molecular Analysis
2.8. Polymerase Chain Reaction (PCR) Amplification and Genetic Fingerprinting Analyses
2.9. Quantitative Real-Time PCR
3. Results
3.1. Genomes and Functional Annotation
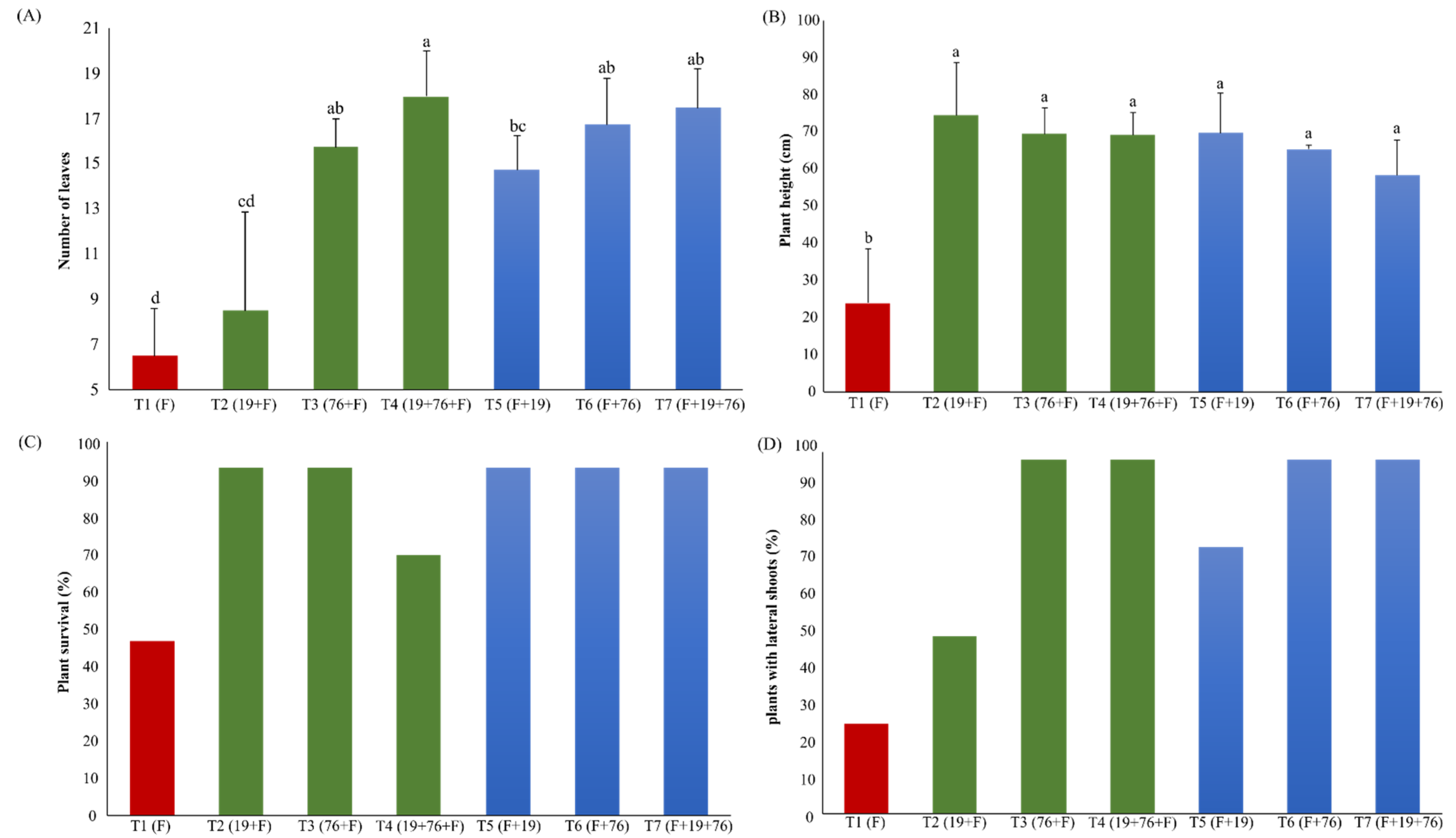
3.2. Pot Experiments in Greenhouse
3.2.1. Plant Health
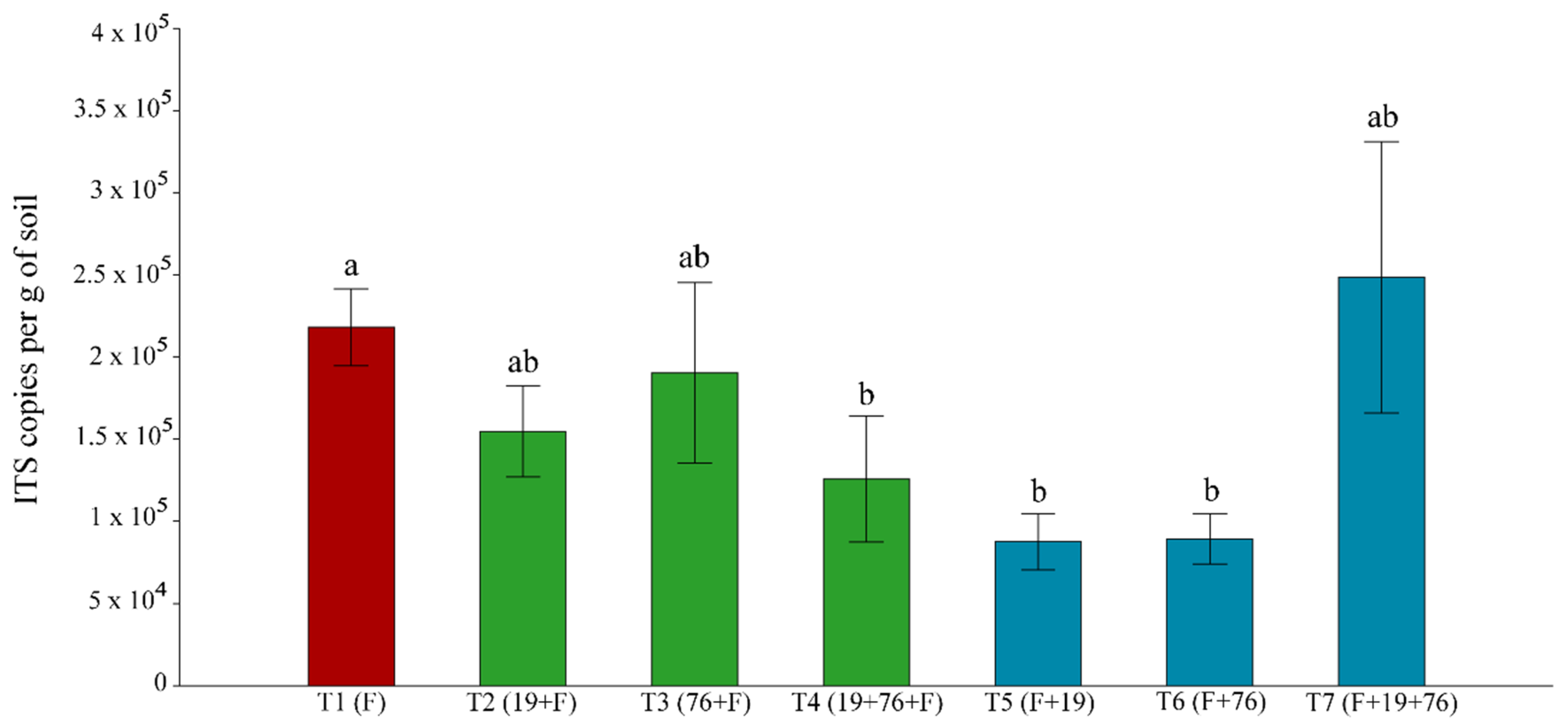
3.2.2. Total Fungal Community Quantification Using Real-Time PCR (qPCR)
3.2.3. Response of the Fungal Communities to the Inoculation of Strains T052-76 and T149-19
3.2.4. Non-Metric Multidimensional Scaling (NMDS) Ordination of PCR-Denaturing Gradient Gel Electrophoresis (DGGE) Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- CIP. 2021. Available online: https://cipotato.org/sweetpotato/ (accessed on 15 March 2021).
- Kays, S.J. Sweetpotato production worldwide: Assessment, trends and the future. Acta Hortic. 2005, 670, 19–25. [Google Scholar] [CrossRef]
- Zengzhi, S.; Bing, D.; Jinxi, H.; Shaozhen, H.; Qingchang, L.; Hong, Z. A genome-wide BAC-end sequence survey provides first insights into sweet potato (Ipomoea batatas (L.) Lam.) genome composition. BMC Genom. 2016, 17, 945. [Google Scholar]
- FAOSTAT—Statistics Database. Available online: http://www.faostat.fao.org (accessed on 15 March 2021).
- Silva, J.B.C.; Lopes, C.A.; Magalhães, J.S. Batata-Doce (Ipomoea Batatas), In Sistemas De Produção Nº 6, Versão Eletrônica; Embrapa Hortaliças: Brasília, Brazil, 2008. [Google Scholar]
- Mussoline, W.A.; Wilkie, A.C. Feed and fuel: The dual-purpose advantage of an industrial sweetpotato. J. Sci. Food Agric. 2016, 97, 1567–1575. [Google Scholar] [CrossRef]
- El Sheikha, A.F.; Ray, R.C. Potential impacts of bio-processing of sweet potato: Review. Crit. Rev. Food Sci. Nutr. 2017, 57, 455–471. [Google Scholar] [CrossRef]
- Williams, R.; Soares, F.; Pereira, L.; Belo, B.; Soares, A.; Setiawan, A.; Browne, M.; Nesbitt, H.; Erskine, W. Sweet potato can contribute to both nutritional and food security in Timor-Leste. Field Crop. Res. 2013, 146, 38–43. [Google Scholar] [CrossRef]
- Lopes, U.P.; Michereff, S.J. Desafios Do Manejo De Doenças Radiculares Causadas Por Fungos; Editora Universitária da UFRPE: Pernambuco, Brazil, 2018. [Google Scholar]
- Mikhail, M.S.; Abdel-Alim, A.I.; Abd-El-Khair, H.; Abdel-Gaied, T.G.; Shereen, A.M. Host range and total cellular protein fingerprint of soft rot Erwinia isolated from some vegetables in Egypt. Plant Arch. 2019, 19, 295–306. [Google Scholar]
- Jiang, L.; Jeong, J.C.; Lee, J.-S.; Park, J.M.; Yang, J.W.; Lee, M.H.; Choi, S.H.; Kim, C.Y.; Kim, D.-H.; Kim, S.W.; et al. Potential of Pantoea dispersa as an effective biocontrol agent for black rot in sweet potato. Sci. Rep. 2019, 9, 16354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, R.B.; Amaro, G.B.; Ribeiro, M.C.V.; Pinheiro, J.B. Reação de genótipos de batata-doce ao mal-do-pé (Plenodomus destruens). In Boletim de Pesquisa e Desenvolvimento, Nº 122; Embrapa Hortaliças: Brasília, Brazil, 2014. [Google Scholar]
- Pereira, R.B.; Fernandes, F.R.; Pinheiro, J.B. Recomendações Para Manejo Da Podridão-Do-Pé Em Batata-Doce. In Comunicado Técnico Nº 79; Embrapa Hortaliças: Brasília, Brazil, 2011. [Google Scholar]
- Gai, Y.; Ma, H.; Chen, X.; Zheng, J.; Chen, H.; Li, H. Stem blight, foot rot and storage tuber rot of sweet potato caused by Plenodomus destruens in China. J. Gen. Plant Pathol. 2016, 82, 181–185. [Google Scholar] [CrossRef]
- Lopes, C.A.; Silva, J.B.C. Management measures to control foot rot of sweet potato caused by Plenodomus destruens. Int. J. Pest Manag. 1993, 39, 72–74. [Google Scholar] [CrossRef]
- Okada, Y.; Kobayashi, A.; Tabuchi, H.; Kuranouchi, T. Review of major sweetpotato pests in Japan, with information on resistance breeding programs. Breed. Sci. 2017, 67, 73–82. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Tariq, M.; Khan, A.; Asif, M.; Khan, F.; Ansari, T.; Shariq, M.; Siddiqui, M.A. Biological control: A sustainable and practical approach for plant disease management. Acta Agric. Scand. B Soil Plant Sci. 2020, 70, 507–524. [Google Scholar] [CrossRef]
- Qiao, J.Q.; Wu, H.J.; Huo, R.; Gao, X.W.; Borriss, R. Stimulation of plant growth and biocontrol by Bacillus amyloliquefaciens subsp. plantarum FZB42 engineered for improved action. Chem. Biol. Technol. Agric. 2014, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Wu, H.-J.; Qiao, J.; Gao, X.; Borriss, R. Novel routes for improving biocontrol activity of Bacillus based bioinoculants. Front. Microbiol. 2015, 6, 1395. [Google Scholar] [CrossRef] [Green Version]
- Marques, J.M.; da Silva, T.F.; Vollú, R.E.; Blank, A.F.; Ding, G.-C.; Seldin, L.; Smalla, K. Plant age and genotype affect the bacterial community composition in the tuber rhizosphere of field-grown sweet potato plants. FEMS Microbiol. Ecol. 2014, 88, 424–435. [Google Scholar] [CrossRef]
- Marques, J.M.; da Silva, T.F.; Vollú, R.E.; de Lacerda, J.R.M.; Blank, A.F.; Smalla, K.; Seldin, L. Bacterial endophytes of sweet potato tuberous roots affected by the plant genotype and growth stage. Appl. Soil Ecol. 2015, 96, 273–281. [Google Scholar] [CrossRef]
- Mateus, J.R.; Marques, J.M.; Dal’Rio, I.; Vollú, R.E.; Coelho, M.R.R.; Seldin, L. Response of the microbial community associated with sweet potato (Ipomoea batatas) to Bacillus safensis and Bacillus velezensis strains. Antonie Van Leeuwenhoek 2019, 112, 501–512. [Google Scholar] [CrossRef]
- Seldin, L.; Rosado, A.S.; Cruz, D.W.; Nobrega, A.; van Elsas, J.D.; Paiva, E. Comparison of Paenibacillus azotofixans strains isolated from rhizoplane, rhizosphere and non-rhizosphere soil from maize planted in two different Brazilian soils. Appl. Environ. Microbiol. 1998, 64, 3860–3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seldin, L.; Dubnau, D. DNA homology among Bacillus polymyxa, Bacillus macerans, Bacillus azotofixans and other nitrogen fixing Bacillus strains. Int. J. Syst. Bacteriol. 1985, 35, 151–154. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; Dejongh, M.; Disz, T.; Edwards, R.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.; Gerdes, S.; Parrello, B.D.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 1, 9. [Google Scholar]
- Smit, E.; Leeflang, P.; Glandorf, B.; Van Elsas, J.D.; Wernars, K. Analysis of fungal diversity in the wheat rhizosphere by sequencing of cloned PCR-amplified genes encoding 18S rRNA and temperature gradient gel electrophoresis. Appl. Environ. Microbiol. 1999, 65, 2614–2621. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Analysis of phylogenetic relationships by amplification and direct sequencing of ribosomal RNA genes. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes: Application to the identification of mycorrhiza and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Heuer, H.; Wieland, J.; Schonfeld, J.; Schonwalder, A.; Gomes, N.C.M.; Smalla, K. Bacterial community profiling using DGGE or TGGE analysis. In Environmental Molecular Microbiology: Protocols and Applications Horizon; Rouchelle, P., Ed.; Horizon Scientific Press: Wymondham, Norfolk, UK, 2001; pp. 177–190. [Google Scholar]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jackson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Fira, D.; Dimkic, I.; Beric, T.; Lozo, J.; Stankovic, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Jacques, P. Surfactin and other lipopeptides from Bacillus spp. In Microbiology; Soberón-Chávez, G., Ed.; Springer: Berlin, Germany, 2011; Volume 20. [Google Scholar]
- Berić, T.; Kojić, M.; Stanković, S.; Topisirović, L.; Degrassi, G.; Myers, M.; Venturi, V.; Fira, D. Antimicrobial activity of Bacillus sp. natural isolates and their potential use in the biocontrol of phytopathogenic bacteria. Food Technol. Biotechnol. 2012, 50, 25–31. [Google Scholar]
- Grady, E.N.; MacDonald, J.; Ho, M.T.; Weselowski, B.; McDowell, T.; Solomon, O.; Renaud, J.; Yuan, Z.C. Characterization and complete genome analysis of the surfactin-producing, plant-protecting bacterium Bacillus velezensis 9D. BMC Microbiol. 2019, 19, 5. [Google Scholar] [CrossRef]
- Galindo, E.; Serrano-Carreón, L.; Gutiérrez, C.; Allende, R.; Balderas, K.; Patiño, M.; Trejo, M.; Wong, M.; Rayo, E.; Isauro, D.; et al. The challenges of introducing a new biofungicide to the market: A case study. Elect. J. Biotechnol. 2013, 16, 5. [Google Scholar] [CrossRef]
- Balderas-Ruíz, K.A.; Bustos, P.; Santamaria, R.I.; González, V.; Cristiano-Fajardo, S.A.; Barrera-Ortíz, S.; Mezo-Villalobos, M.; Aranda-Ocampo, S.; Guevara-García, A.; Galindo, E.; et al. Bacillus velezensis 83 a bacterial strain from mango phyllosphere, useful for biological control and plant growth promotion. AMB Express 2020, 10, 163. [Google Scholar] [CrossRef]
- Cui, L.; Yang, C.; Wei, L.; Li, T.; Chen, X. Isolation and identification of an endophytic bacteria Bacillus velezensis 8-4 exhibiting biocontrol activity against potato scab. Biol. Control 2020, 141, 104156. [Google Scholar] [CrossRef]
- Rong, S.; Xu, H.; Li, L.; Chen, R.; Gao, X.; Xu, Z. Antifungal activity of endophytic Bacillus safensis B21 and its potential application as a biopesticide to control rice blast. Pestic. Biochem. Physiol. 2020, 162, 69–77. [Google Scholar] [CrossRef]
- Zhu, Z.; Peng, Q.; Man, Y.; Li, Z.; Zhou, X.; Bai, L.; Peng, D. Analysis of the antifungal properties of Bacillus velezensis B-4 through a bioassay and complete-genome sequencing. Front. Genet. 2020, 11, 703. [Google Scholar] [CrossRef]
- Teixeira, G.M.; Mosela, M.; Nicoletto, M.L.A.; Ribeiro, R.A.; Hungria, M.; Youssef, K.; Higashi, A.Y.; Mian, S.; Ferreira, A.S.; Gonçalves, L.S.A.; et al. Genomic insights into the antifungal activity and plant growth-promoting ability in Bacillus velezensis CMRP. Front. Microbiol. 2021, 11, 618415. [Google Scholar] [CrossRef]
- Chen, L.; Heng, J.; Qin, S.; Bian, K. A comprehensive understanding of the biocontrol potential of Bacillus velezensis LM2303 against Fusarium head blight. PLoS ONE 2018, 13, e0198560. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Garcia, A.; Romero, D.; De Vicente, A. Plant protection and growth stimulation by microorganisms: Biotechnological applications of Bacilli in agriculture. Curr. Opin. Biotechnol. 2011, 22, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of bacillomycin- and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soilborne plant pathogens. Agric. Food Chem. 2012, 60, 2976–2981. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Geng, X.B.; Ji, P.S.; Pan, C.Q.; Wei, S. Isolation and evaluation of a Bacillus methylotrophicus strain for control of corn stalk rot. Biocontrol Sci. Technol. 2016, 26, 727–773. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Cluster (Size in nt) | Similarity (%) a | Access Number MIBiG b | Coding for | Related Genes |
|---|---|---|---|---|
| 88,273 | 100 | BGC0001103 | Mycosubtilin | fenF, mycA, mycB and mycC |
| 97,565 | 100 | BGC0001089 | Bacillaene | bae genes (I, J, L, M, N) |
| 88,300 | 100 | BGC0000181 | Macrolactin | pks and pdhA |
| 51,787 | 100 | BGC0000309 | Bacillibactin | dhb, yui, among others |
| 41,419 | 100 | BGC0001184 | Bacilysin | ywfA and bacA, B, C, D, E and F |
| 22,182 | 91 | BGC0000569 | Plantazolicin | ptn genes |
| 25,256 | 39 | BGC0000433 | Surfactin | srfAB, srfAC and srfAD |
| 160,188 | 43 | BGC0000433 | Surfactin | srfAA and other accessory genes |
| 9108 | 8 | BGC0000433 | Surfactin | srfAA and srfAB |
| 13,008 | 20 | BGC0001095 | Fengycin | fenA, fenB and fenC |
| 9205 | 13 | BGC0001095 | Fengycin | fenC and fenD |
| 49,123 | 30 | BGC0000407 | Plipastatin | ppsA and other accessory genes |
| 33,892 | 66 | BGC0000176 | Dificidine | dif genes (A, B, C, D, E, F, G, I, J, K) |
| Gene Cluster (Size in nt) | Similarity (%) a | Access Number MIBiG b | Coding for | Related Genes |
|---|---|---|---|---|
| 41,422 | 85 | BGC0001184 | Bacylisin | bac and ywf genes |
| 27,830 | 50 | BGC0000381 | Lichenysin | hyuA and B, putA, B and C and lchAA genes |
| 49,710 | 53 | BGC0000309 | Bacillibactin | dhb genes |
| 28,413 | 53 | BGC0001103 | Fengycin | yng genes |
| 44,186 | 39 | BGC0000433 | Surfactin | srfA and ycx genes, and others |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateus, J.R.; Dal’Rio, I.; Jurelevicius, D.; da Mota, F.F.; Marques, J.M.; Ramos, R.T.J.; da Costa da Silva, A.L.; Gagliardi, P.R.; Seldin, L. Bacillus velezensis T149-19 and Bacillus safensis T052-76 as Potential Biocontrol Agents against Foot Rot Disease in Sweet Potato. Agriculture 2021, 11, 1046. https://doi.org/10.3390/agriculture11111046
Mateus JR, Dal’Rio I, Jurelevicius D, da Mota FF, Marques JM, Ramos RTJ, da Costa da Silva AL, Gagliardi PR, Seldin L. Bacillus velezensis T149-19 and Bacillus safensis T052-76 as Potential Biocontrol Agents against Foot Rot Disease in Sweet Potato. Agriculture. 2021; 11(11):1046. https://doi.org/10.3390/agriculture11111046
Chicago/Turabian StyleMateus, Jackeline Rossetti, Isabella Dal’Rio, Diogo Jurelevicius, Fabio Faria da Mota, Joana Montezano Marques, Rommel Thiago Juca Ramos, Artur Luiz da Costa da Silva, Paulo Roberto Gagliardi, and Lucy Seldin. 2021. "Bacillus velezensis T149-19 and Bacillus safensis T052-76 as Potential Biocontrol Agents against Foot Rot Disease in Sweet Potato" Agriculture 11, no. 11: 1046. https://doi.org/10.3390/agriculture11111046
APA StyleMateus, J. R., Dal’Rio, I., Jurelevicius, D., da Mota, F. F., Marques, J. M., Ramos, R. T. J., da Costa da Silva, A. L., Gagliardi, P. R., & Seldin, L. (2021). Bacillus velezensis T149-19 and Bacillus safensis T052-76 as Potential Biocontrol Agents against Foot Rot Disease in Sweet Potato. Agriculture, 11(11), 1046. https://doi.org/10.3390/agriculture11111046