Humic Substances Improve Vegetable Seedling Quality and Post-Transplant Yield Performance under Stress Conditions


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growing Media, and Amendment Treatments
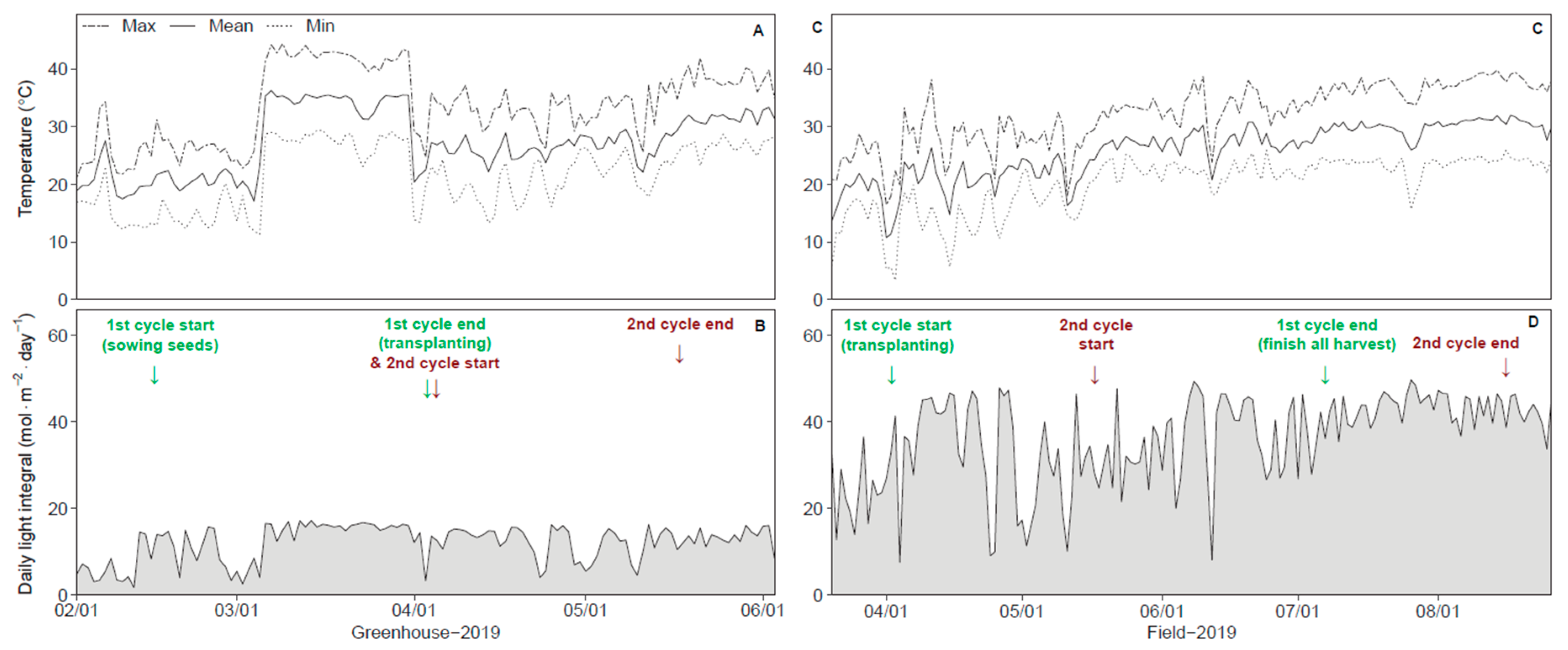
2.2. Growth Environments and Stress Treatments
2.3. Seedling and Transplant Quality Evaluation and Yield Performance
2.4. Statistical Analysis
3. Results
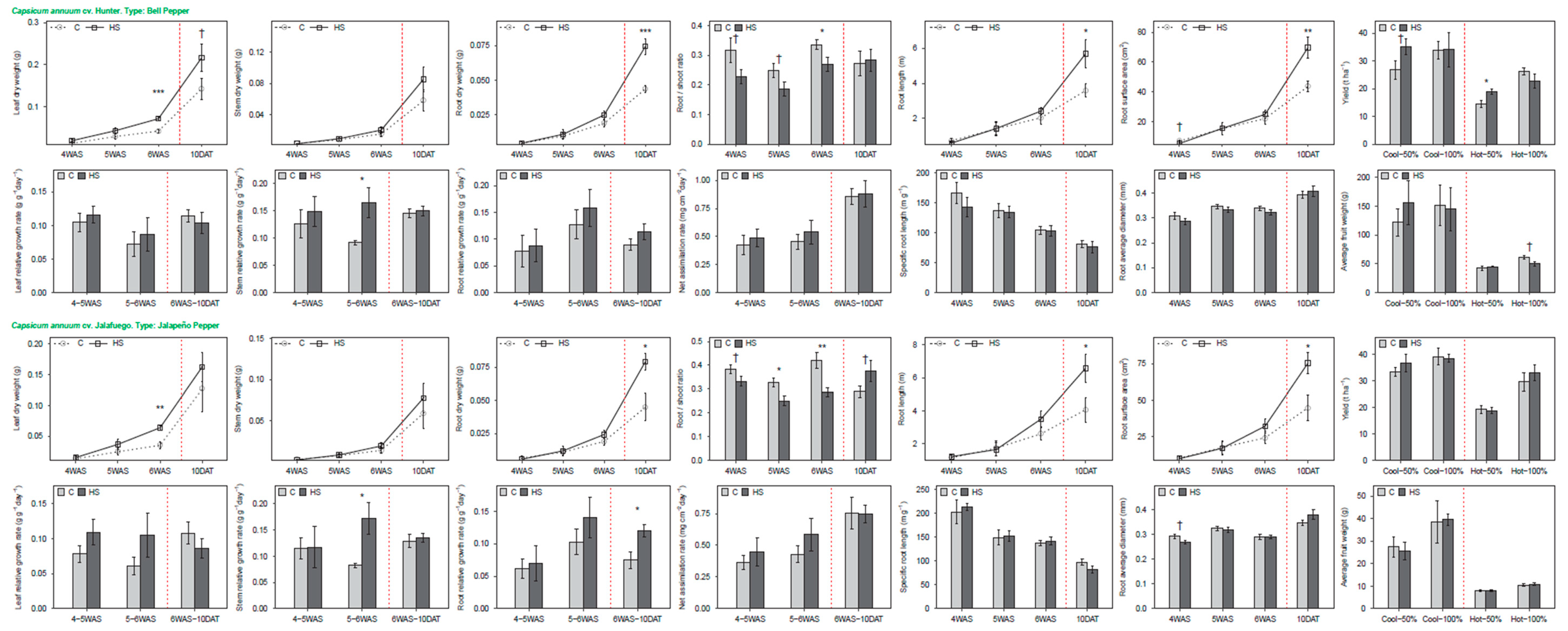
3.1. Pepper
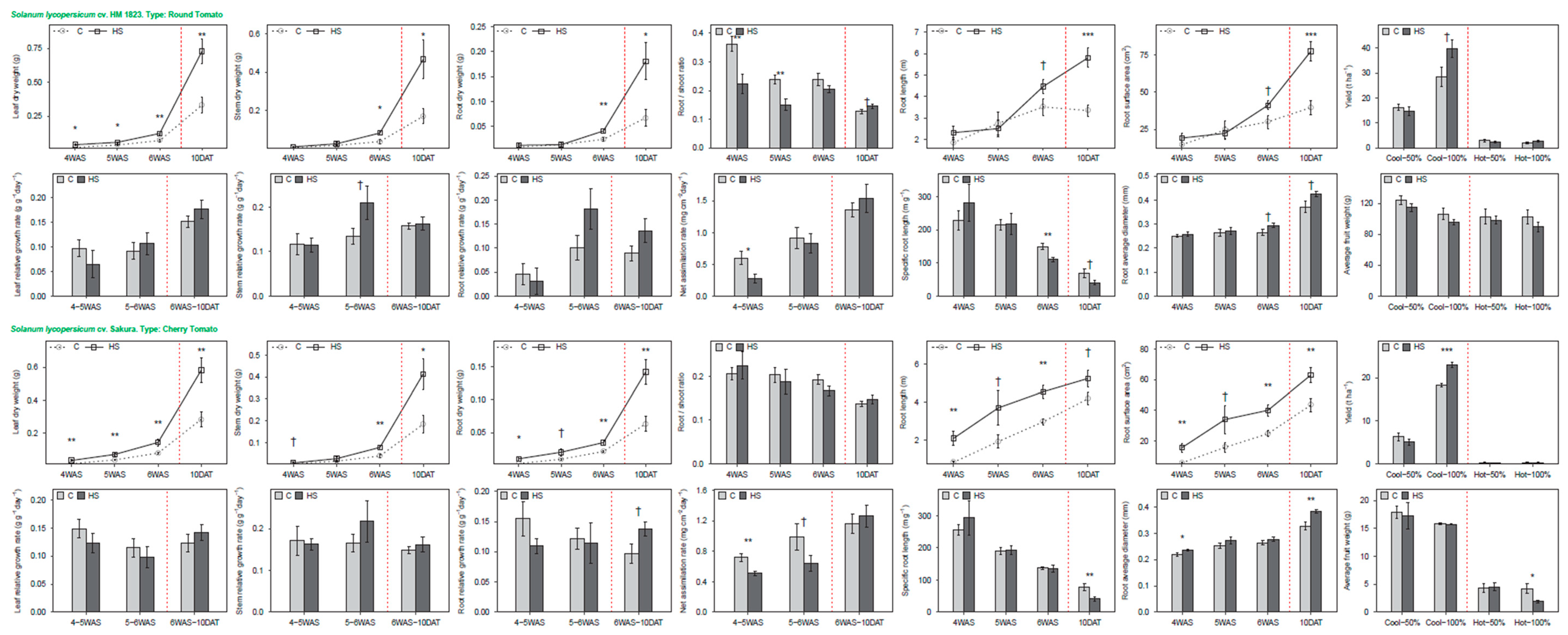
3.2. Tomato
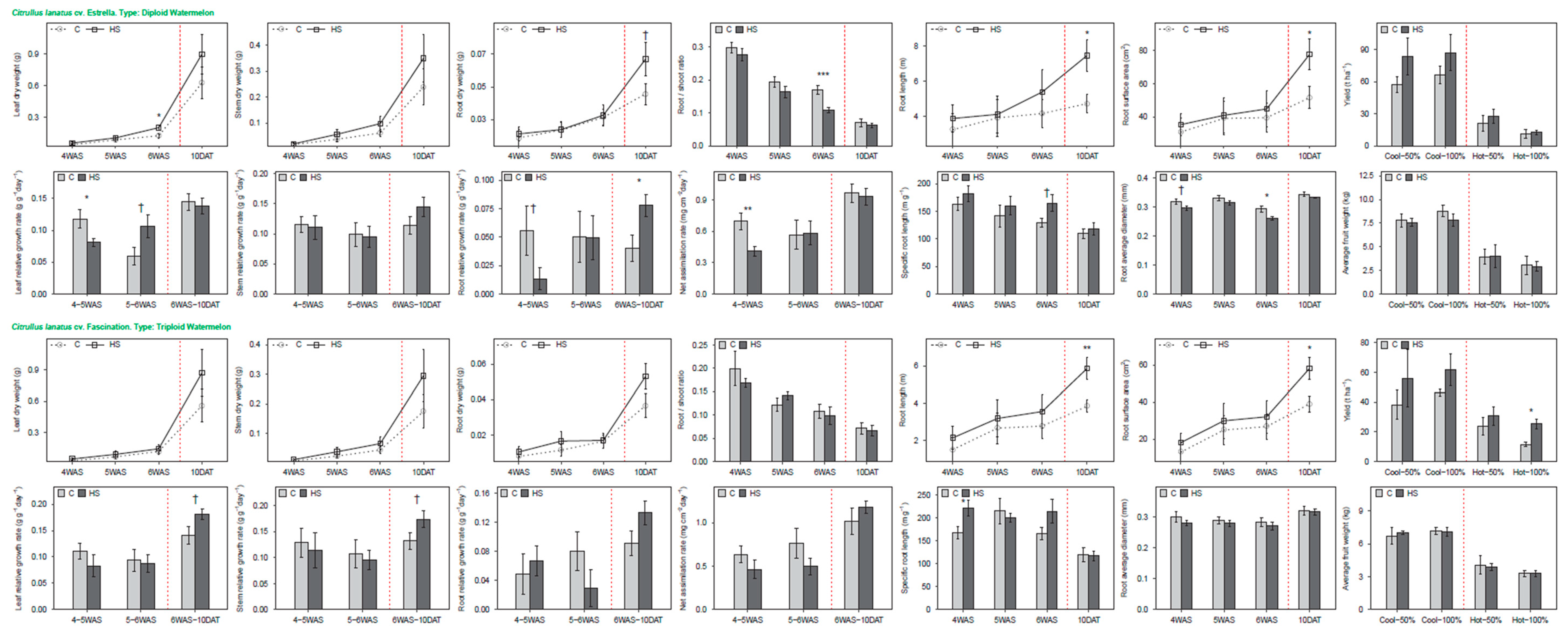
3.3. Watermelon
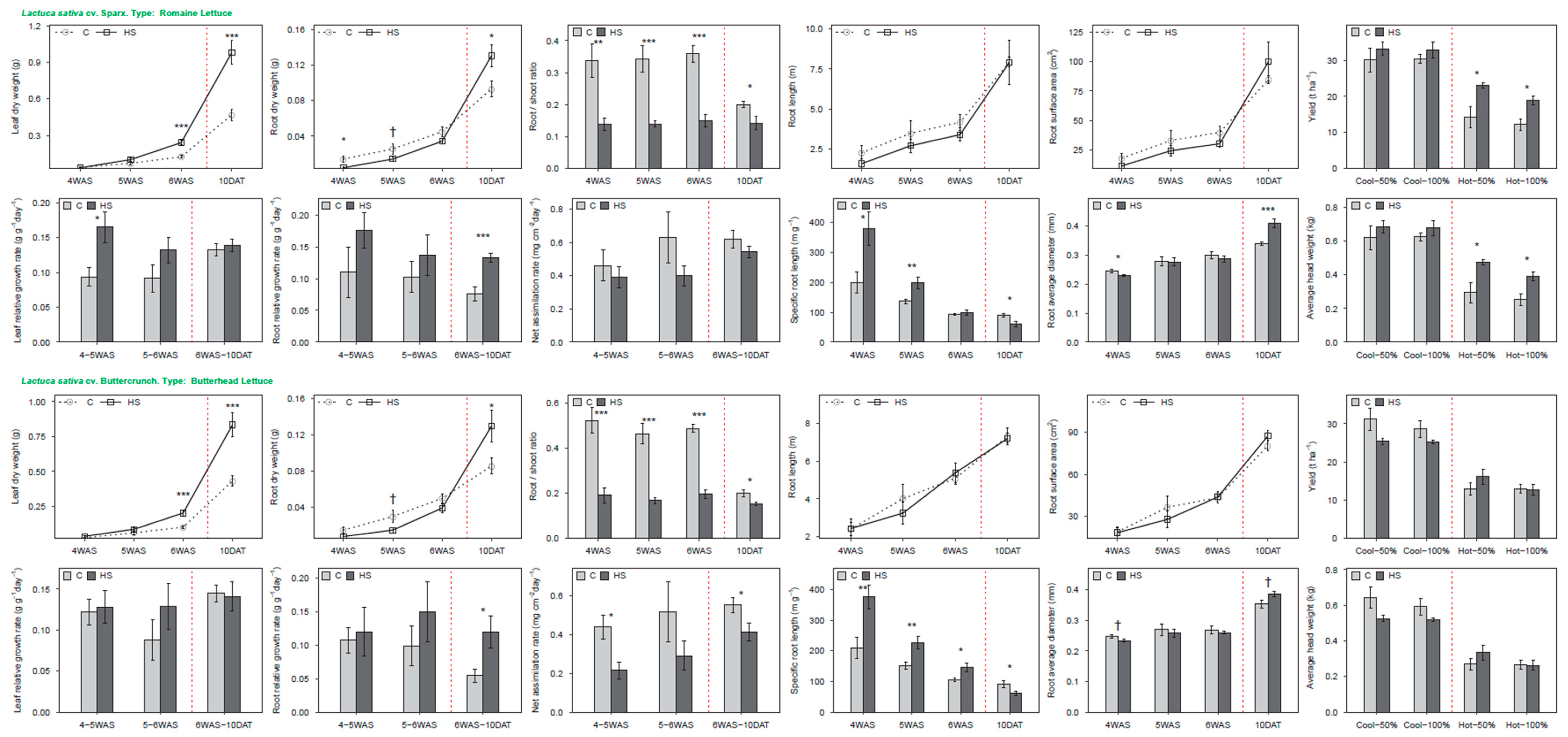
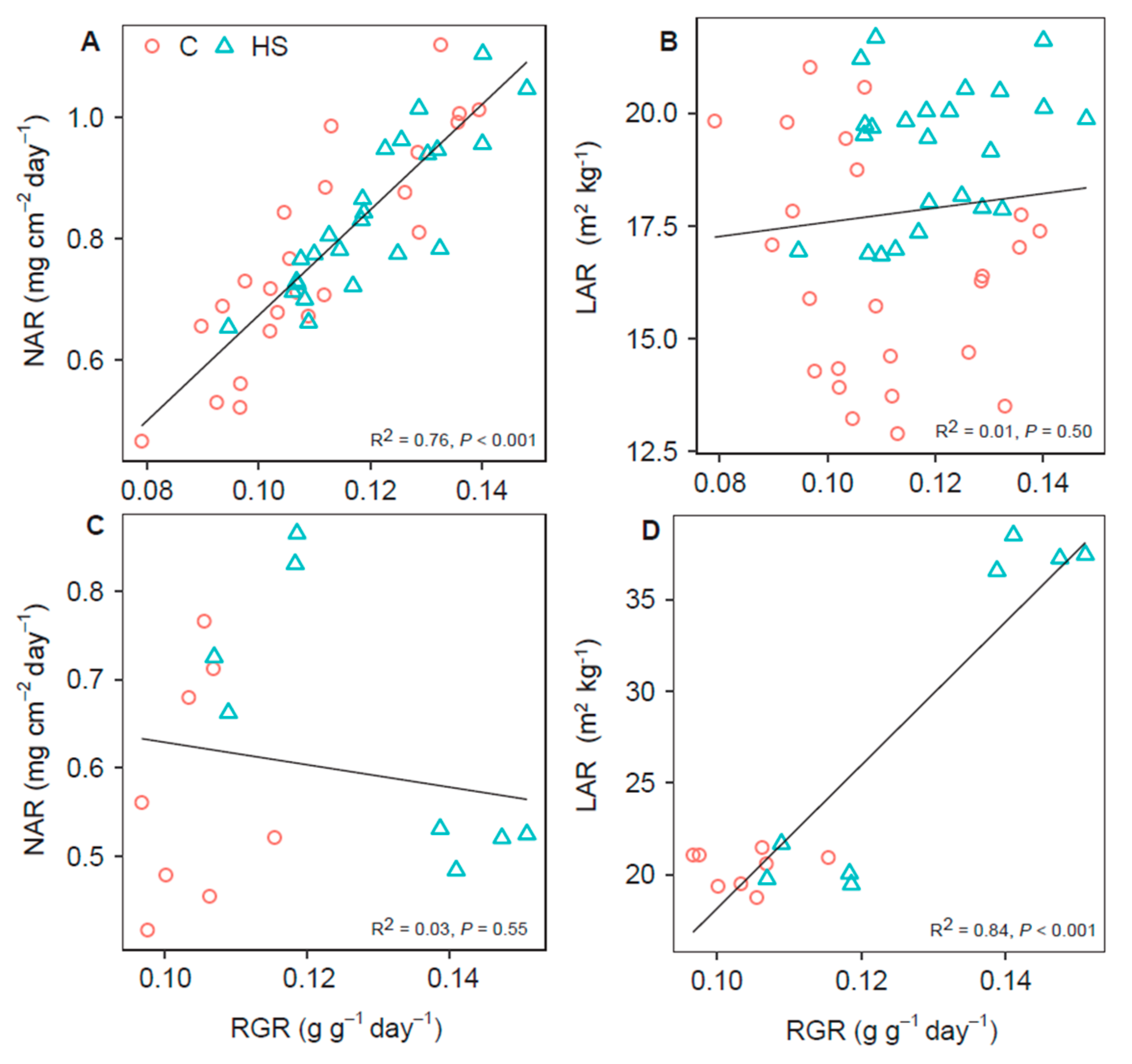
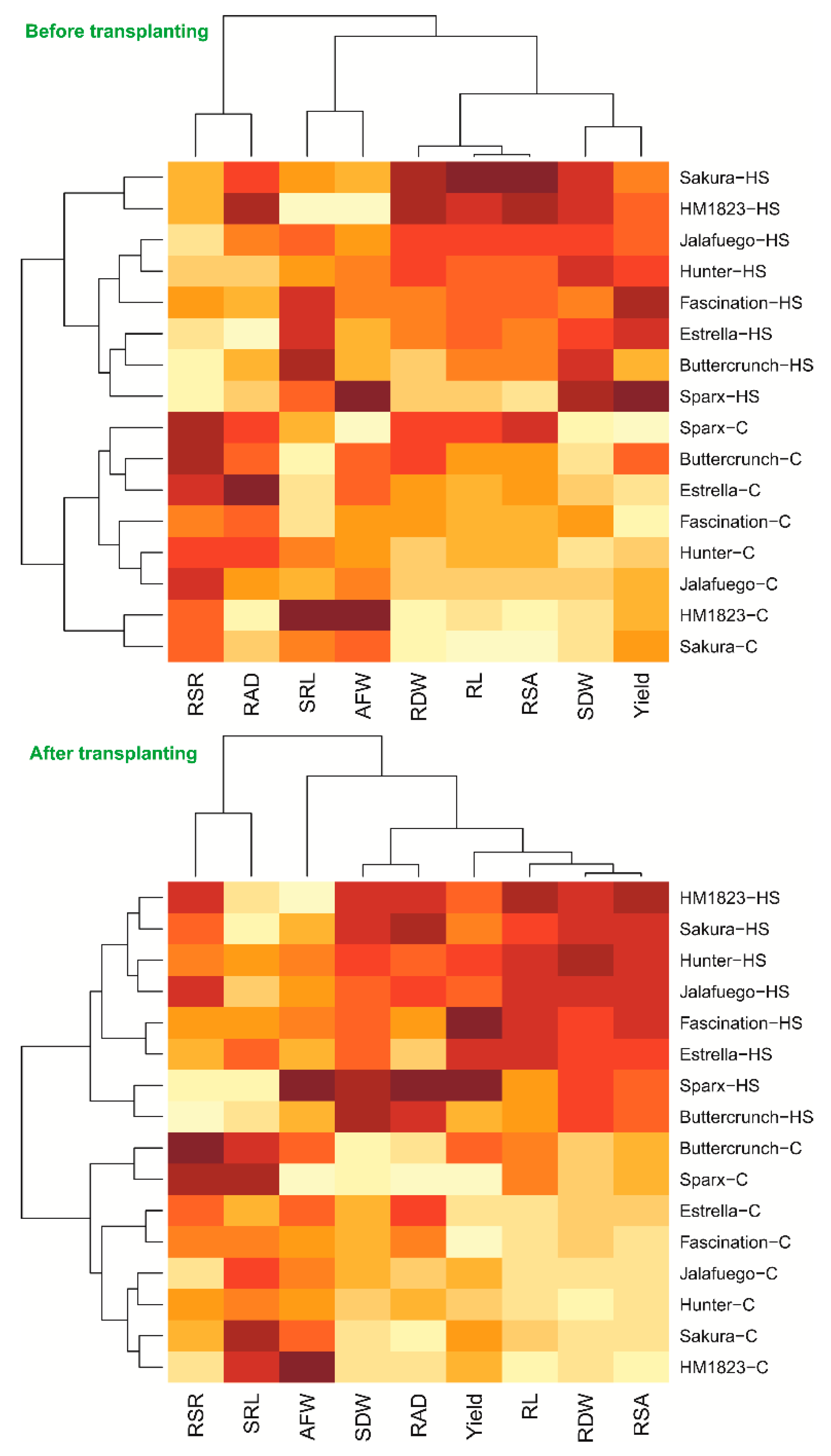
3.4. Lettuce
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leskovar, D.I. Transplanting. In The Physiology of Vegetable Crops, 2nd ed.; Wien, H.C., Stützel, H., Eds.; CABI: Oxfordshire, UK, 2020; pp. 31–60. [Google Scholar]
- Kerbiriou, P.J.; Stomph, T.J.; Van Bueren, E.T.L.; Struik, P.C. Influence of Transplant Size on the Above- and Below-Ground Performance of Four Contrasting Field-Grown Lettuce Cultivars. Front. Plant Sci. 2013, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhong, Q.; Li, Y.; Li, G.-H.; Ding, Y.; Wang, S.; Liu, Z.; Tang, S.; Ding, C.; Chen, L. Triacontanol Reduces Transplanting Shock in Machine-Transplanted Rice by Improving the Growth and Antioxidant Systems. Front. Plant Sci. 2016, 7, 501. [Google Scholar] [CrossRef] [PubMed]
- Russo, V.M. Biological Amendment, Fertilizer Rate, and Irrigation Frequency for Organic Bell Pepper Transplant Production. HortScience 2006, 41, 1402–1407. [Google Scholar] [CrossRef]
- Leskovar, D.I.; Stoffella, P.J. Vegetable Seedling Root Systems: Morphology, Development, and Importance. HortScience 1995, 30, 1153–1159. [Google Scholar] [CrossRef]
- Soundy, P.; Cantliffe, D.J.; Hochmuth, G.J.; Stoffella, P.J. Management of Nitrogen and Irrigation in Lettuce Transplant Production affects Transplant Root and Shoot Development and Subsequent Crop Yields. HortScience 2005, 40, 607–610. [Google Scholar] [CrossRef]
- Leskovar, D.I.; Othman, Y.A. Low Nitrogen Fertigation Promotes Root Development and Transplant Quality in Globe Artichoke. HortScience 2016, 51, 567–572. [Google Scholar] [CrossRef]
- Leskovar, D.I. Root and Shoot Modification by Irrigation. HortTechnology 1998, 8, 510–514. [Google Scholar] [CrossRef]
- Leskovar, D.I.; Vavrina, C.S. Onion Growth and Yield are Influenced by Transplant Tray Cell Size and Age. Sci. Hortic. 1999, 80, 133–143. [Google Scholar] [CrossRef]
- Hernández, R.; Eguchi, T.; Deveci, M.; Kubota, C. Tomato Seedling Physiological Responses Under Different Percentages of Blue and Red Photon Flux Ratios Using Leds and Cool White Fluorescent Lamps. Sci. Hortic. 2016, 213, 270–280. [Google Scholar] [CrossRef]
- Pascual, J.; Ceglie, F.; Tüzel, Y.; Koller, M.; Koren, A.; Hitchings, R.; Tittarelli, F. Organic Substrate for Transplant Production in Organic Nurseries. A Review. Agron. Sustain. Dev. 2018, 38, 35. [Google Scholar] [CrossRef]
- Jack, A.L.H.; Rangarajan, A.; Culman, S.W.; Sooksa-Nguan, T.; Thies, J.E. Choice of Organic Amendments in Tomato Transplants has Lasting Effects on Bacterial Rhizosphere Communities and Crop Performance in the Field. Appl. Soil Ecol. 2011, 48, 94–101. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and Fulvic Acids as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Hartwigsen, J.A.; Evans, M.R. Humic Acid Seed and Substrate Treatments Promote Seedling Root Development. HortScience 2000, 35, 1231–1233. [Google Scholar] [CrossRef]
- Türkmen, Ö.; Dursun, A.; Turan, M.; Erdinç, Ç. Calcium and Humic Acid affect Seed Germination, Growth, and Nutrient Content of Tomato (Lycopersicon esculentum L.) Seedlings under Saline Soil Conditions. Acta Agric. Scand. Sect. B Plant Soil Sci. 2004, 54, 168–174. [Google Scholar] [CrossRef]
- Osman, A.S.; Rady, M.M. Effect of Humic Acid as an Additive to Growing Media to Enhance the Production of Eggplant and Tomato Transplants. J. Hortic. Sci. Biotechnol. 2014, 89, 237–244. [Google Scholar] [CrossRef]
- Brooks, S.D.; DeMott, P.J.; Kreidenweis, S.M. Water Uptake by Particles Containing Humic Materials and Mixtures of Humic Materials with Ammonium Sulfate. Atmos. Environ. 2004, 38, 1859–1868. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, H.; Wang, H.; Xu, L.; Samuel, B.; Chang, J.; Liu, F.; Chen, H. Molecular Structure-Reactivity Correlations of Humic Acid and Humin Fractions from a Typical Black Soil for Hexavalent Chromium Reduction. Sci. Total Environ. 2019, 651, 2975–2984. [Google Scholar] [CrossRef]
- Gerke, J. Concepts and Misconceptions of Humic Substances as the Stable Part of Soil Organic Matter: A Review. Agronomy 2018, 8, 76. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements; Food and Agriculture Organization: Rome, Italy, 1998. [Google Scholar]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; Dezonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the Next Generation of Scientific Image Data. Bmc Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing. Available online: https://www.R-project.org (accessed on 11 September 2019).
- Zaller, J.G. Vermicompost in Seedling Potting Media can affect Germination, Biomass Allocation, Yields and Fruit Quality of Three Tomato Varieties. Eur. J. Soil Boil. 2007, 43, S332–S336. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Bassel, G.W. Seed Vigour and Crop Establishment: Extending Performance Beyond Adaptation. J. Exp. Bot. 2015, 67, 567–591. [Google Scholar] [CrossRef]
- Luo, Y.; Liang, J.; Zeng, G.; Chen, M.; Mo, D.; Li, G.; Zhang, D. Seed Germination Test for Toxicity Evaluation of Compost: Its Roles, Problems and Prospects. Waste Manag. 2018, 71, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Ninnemann, H.; Fischer, K.; Brendler, E.; Liebner, M.; Rosenau, T.; Liebner, F. Characterisation of Humic Matter Fractions Isolated from Ammonoxidised Miocene Lignite. J. Biobased Mater. Bioenergy 2011, 5, 241–252. [Google Scholar] [CrossRef]
- Zherebker, A.; Kostyukevich, Y.I.; Kononikhin, A.S.; Nikolaev, E.N.; Perminova, I.V. Molecular Compositions of Humic Acids Extracted from Leonardite and Lignite as Determined by Fourier Transform Ion Cyclotron Resonance Mass Spectrometry. Mendeleev Commun. 2016, 26, 446–448. [Google Scholar] [CrossRef]
- Francioso, O.; Sanchez-Cortes, S.; Tugnoli, V.; Marzadori, C.; Ciavatta, C. Spectroscopic Study (DRIFT, SERS and 1H NMR) of Peat, Leonardite and Lignite Humic Substances. J. Mol. Struct. 2001, 565, 481–485. [Google Scholar] [CrossRef]
- Houghton, J.; Thompson, K.; Rees, M. Does Seed Mass Drive the Differences in Relative Growth Rate Between Growth Forms? Proc. R. Soc. B Boil. Sci. 2013, 280, 20130921. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P.; Hunt, R. Relative Growth-Rate: Its Range and Adaptive Significance in a Local Flora. J. Ecol. 1975, 63, 393. [Google Scholar] [CrossRef]
- Poorter, H.; Remkes, C. Leaf Area Ratio and Net Assimilation Rate of 24 Wild Species Differing in Relative Growth Rate. Oecologia 1990, 83, 553–559. [Google Scholar] [CrossRef]
- Shipley, B. Net Assimilation Rate, Specific Leaf Area and Leaf Mass Ratio: Which is Most Closely Correlated with Relative Growth Rate? A Meta-Analysis. Funct. Ecol. 2006, 20, 565–574. [Google Scholar] [CrossRef]
- Medek, D.E.; Ball, M.; Schortemeyer, M. Relative Contributions of Leaf Area Ratio and Net Assimilation Rate to Change in Growth Rate Depend on Growth Temperature: Comparative Analysis of Subantarctic and Alpine Grasses. New Phytol. 2007, 175, 290–300. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Heuvelink, E.; Goudriaan, J. Modelling Biomass Production and Yield of Horticultural Crops: A Review. Sci. Hortic. 1998, 74, 83–111. [Google Scholar] [CrossRef]
- Mašková, T.; Herben, T. Root:Shoot Ratio in Developing Seedlings: How Seedlings Change their Allocation in Response to Seed Mass and Ambient Nutrient Supply. Ecol. Evol. 2018, 8, 7143–7150. [Google Scholar] [CrossRef] [PubMed]
- Kubota, C. Growth, Development, Transpiration and Translocation as Affected by Abiotic Environmental Factors. In Plant Factory; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 151–164. [Google Scholar]
- Ryser, P. The Mysterious Root Length. Plant Soil 2006, 286, 1–6. [Google Scholar] [CrossRef]
- Yano, K.; Kume, T. Root Morphological Plasticity for Heterogeneous Phosphorus Supply in Zea mays L. Plant Prod. Sci. 2005, 8, 427–432. [Google Scholar] [CrossRef]
- Li, H.; Ma, Q.; Li, H.; Zhang, F.; Rengel, Z.; Shen, J. Root Morphological Responses to Localized Nutrient Supply Differ Among Crop Species with Contrasting Root Traits. Plant Soil 2013, 376, 151–163. [Google Scholar] [CrossRef]
- Kramer-Walter, K.; Bellingham, P.; Millar, T.; Smissen, R.; Richardson, S.J.; Laughlin, D.C. Root Traits are Multidimensional: Specific Root Length is Independent from Root Tissue Density and the Plant Economic Spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Yoneda, Y. Local Temperature Control in Greenhouse Vegetable Production. Hortic. J. 2019, 88, 305–314. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Staggenborg, S.A.; Ristic, Z. Impacts of Drought and/or Heat Stress on Physiological, Developmental, Growth, and Yield Processes of Crop Plants. In Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes; Ahuja, L.R., Reddy, V.R., Saseendran, S.A., Yu, Q., Eds.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2008; pp. 301–355. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A Vegetal Biopolymer-Based Biostimulant Promoted Root Growth in Melon While Triggering Brassinosteroids and Stress-Related Compounds. Front. Plant Sci. 2018, 9, 11. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size Distribution (%) | TP 1 | AS 2 | CWHC 3 | BD 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| <0.25 mm | 0.25–0.50 | 0.50–1.00 | 1.00–2.00 | 2.00–2.80 | >2.80 | (%) | (%) | (%) | (g/cm3) | |
| CM | 0.2 | 1.1 | 20.0 | 55.3 | 4.9 | 18.5 | 58.6 | 2.9 | 55.7 | 0.07 |
| HS | 23.4 | 28.4 | 29.0 | 18.0 | 1.1 | 0.1 | 65.8 | 5.3 | 60.5 | 0.61 |
| pH | EC 1 | OC 2 | Total N | P | K | Ca | Mg | S | Na | Fe | Zn | Mn | Cu | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (dS/m) | (%) | (%) | (mg/kg) | |||||||||||
| CM | 5.5 | 0.08 | 41.5 | 0.63 | 1.85 | 7.52 | 938 | 123 | 14.82 | 25.70 | 19.43 | 4.56 | 10.44 | 0.72 |
| HS | 7.4 | 0.44 | 65.7 | 5.48 | 2.90 | 56.22 | 451 | 334 | 46.52 | 452.18 | 4.18 | 0.04 | 4.59 | 0.04 |
| FS | 8.0 | 0.29 | 2.0 | N/A 3 | 47.03 | 801.68 | 11355 | 206 | 19.05 | 4.13 | 2.99 | 0.78 | 15.29 | 0.73 |
| ANOVA | Pepper | Tomato | Watermelon | Lettuce | ||||
|---|---|---|---|---|---|---|---|---|
| Bell | Jalapeño | Round | Cherry | Diploid | Triploid | Romaine | Butterhead | |
| Yield | ||||||||
| S | *** | *** | *** | *** | *** | *** | *** | *** |
| IR | ** | *** | *** | *** | NS | NS | NS | NS |
| A | NS | NS | † | * | * | * | ** | NS |
| S × IR | NS | * | *** | *** | NS | NS | NS | NS |
| S × A | NS | NS | † | * | NS | NS | NS | * |
| IR × A | * | NS | * | *** | NS | NS | NS | NS |
| S × IR × A | NS | NS | † | *** | NS | NS | NS | NS |
| AFW | ||||||||
| S | *** | *** | * | *** | *** | *** | *** | *** |
| IR | NS | ** | * | * | NS | NS | NS | NS |
| A | NS | NS | † | NS | NS | NS | ** | NS |
| S × IR | NS | † | NS | NS | NS | NS | NS | NS |
| S × A | NS | NS | NS | NS | NS | NS | NS | * |
| IR × A | NS | NS | NS | NS | NS | NS | NS | NS |
| S × IR × A | NS | NS | NS | NS | NS | NS | NS | NS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, K.; Leskovar, D.I. Humic Substances Improve Vegetable Seedling Quality and Post-Transplant Yield Performance under Stress Conditions. Agriculture 2020, 10, 254. https://doi.org/10.3390/agriculture10070254
Qin K, Leskovar DI. Humic Substances Improve Vegetable Seedling Quality and Post-Transplant Yield Performance under Stress Conditions. Agriculture. 2020; 10(7):254. https://doi.org/10.3390/agriculture10070254
Chicago/Turabian StyleQin, Kuan, and Daniel I. Leskovar. 2020. "Humic Substances Improve Vegetable Seedling Quality and Post-Transplant Yield Performance under Stress Conditions" Agriculture 10, no. 7: 254. https://doi.org/10.3390/agriculture10070254
APA StyleQin, K., & Leskovar, D. I. (2020). Humic Substances Improve Vegetable Seedling Quality and Post-Transplant Yield Performance under Stress Conditions. Agriculture, 10(7), 254. https://doi.org/10.3390/agriculture10070254