Evaluation of Mugwort (Artemisia vulgaris L.) Aqueous Extract as a Potential Bioherbicide to Control Amaranthus retroflexus L. in Maize




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Sampling and Extractions
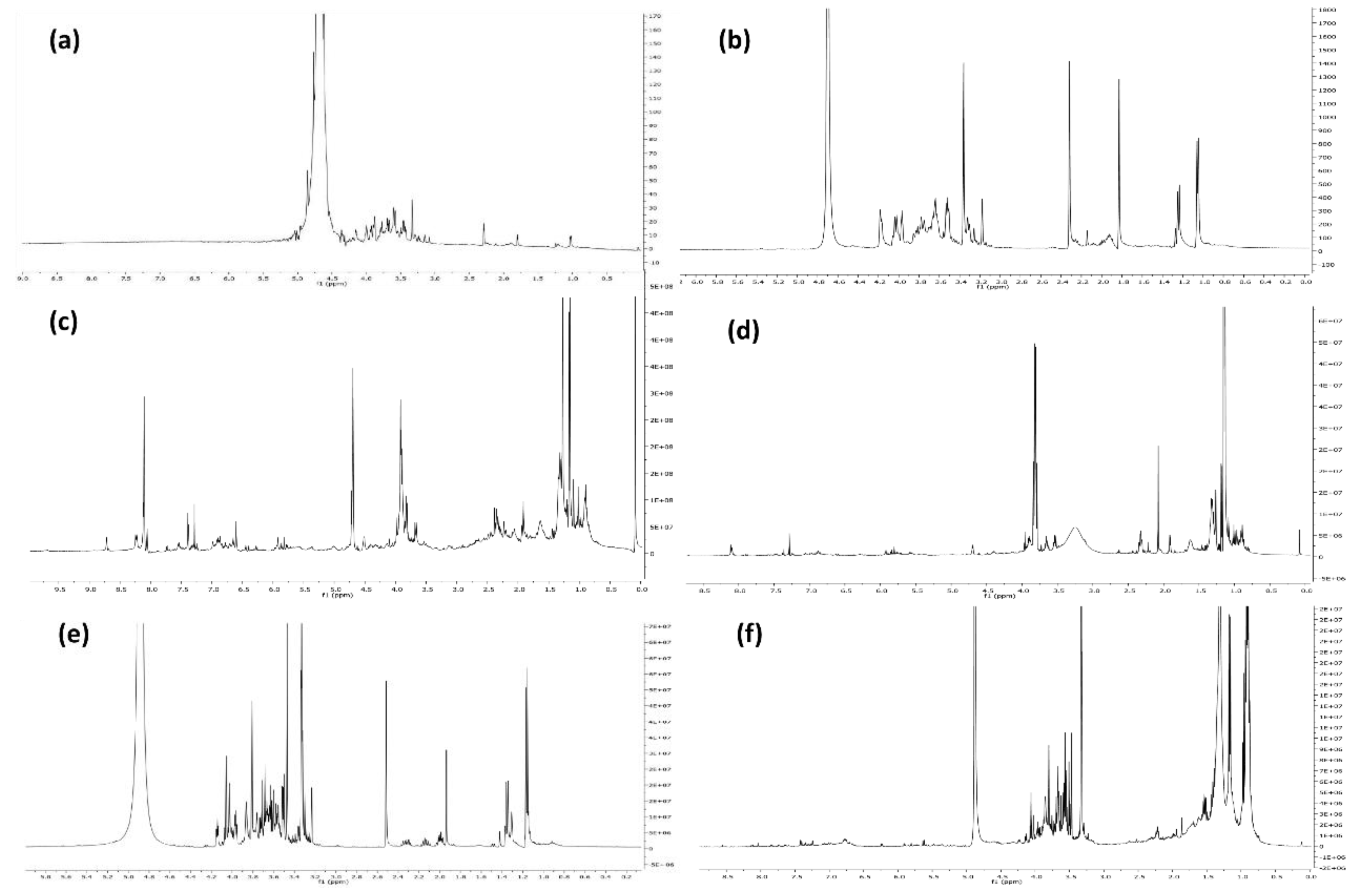
2.2. Extracts Chemical Characterization
2.3. Experiment 1: Petri Dish Bioassays
2.4. Experiment 2: Pot Culture Bioassays
2.5. Statistical Analysis
3. Results and Discussion
3.1. Qualitative Composition of the Extract
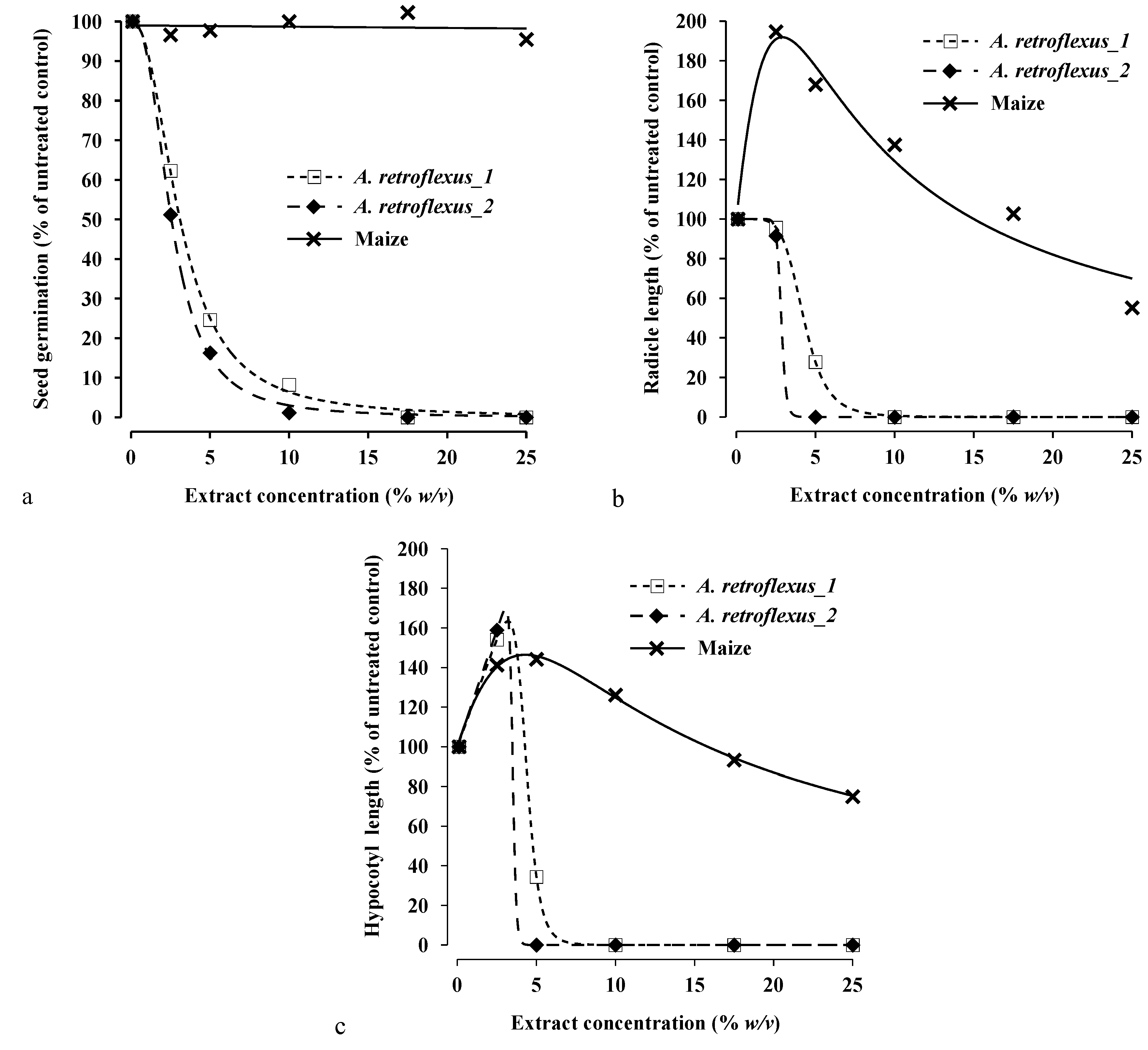
3.2. Exp.1: Effects of Extract on Seed Germination, Radicle and Hypocotyl Lengths of Seedling in Maize and Redroot Pigweed
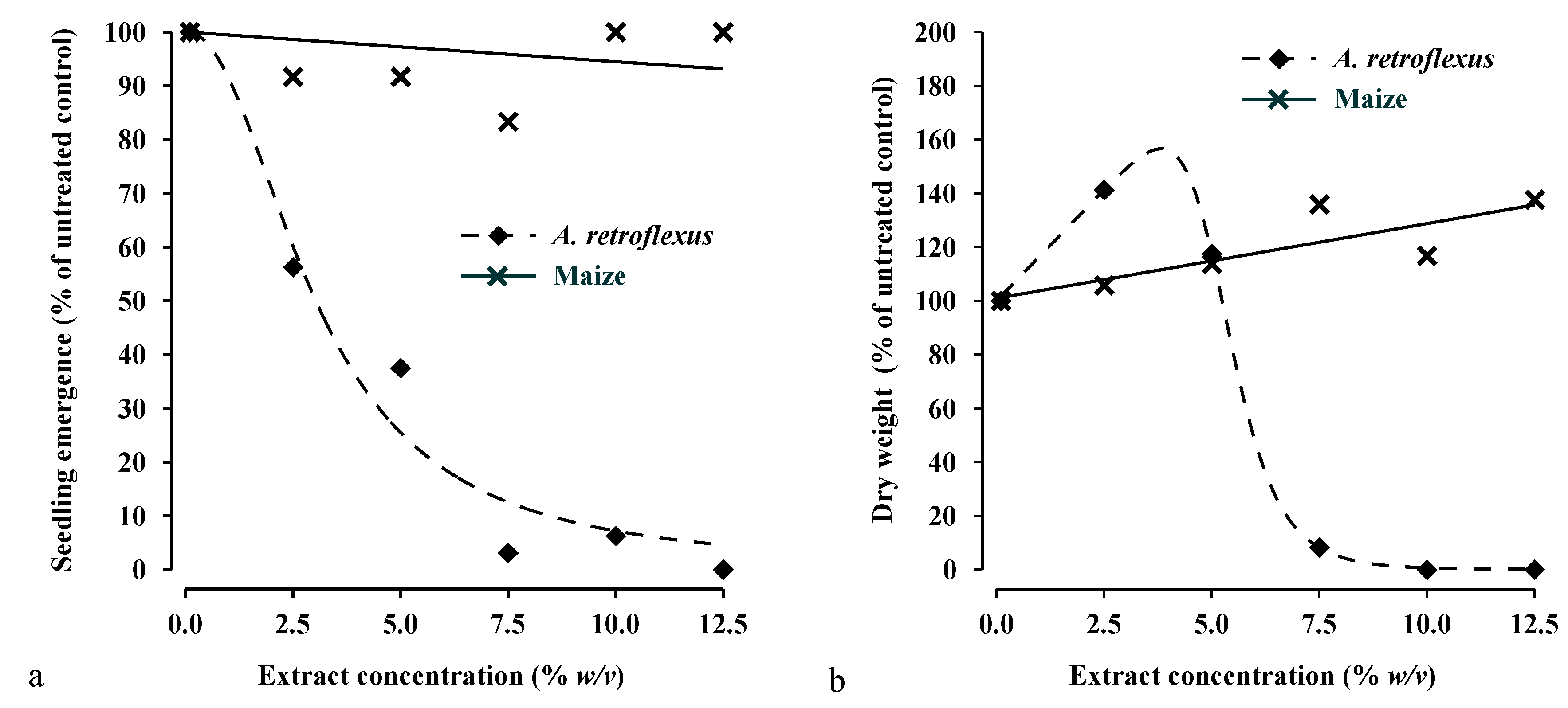
3.3. Exp.2: Effects of Extract on Seedlings Emergence and Plant Growth in Maize Andredroot Pigweed
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT–Crops Data. 2020. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 9 December 2020).
- World Data Atlas, World and Regional Statistics, National Data, Maps, Rankings: Maize Production Quantity. Knoema. 2020. Available online: https://knoema.com/atlas/topics/Agriculture/Crops-Production-Quantity-tonnes/Maize-production (accessed on 22 November 2020).
- Mhlanga, B.; Chauhan, B.S.; Thierfelder, C. Weed management in maize using crop competition: A review. Crop Prot. 2016, 88, 28–36. [Google Scholar] [CrossRef]
- European Commission. Cereals Market Situation Committee for the Common Organisation of Agricultural Markets, 29 October 2020. 2020. Available online: https://circabc.europa.eu/sd/a/df2b1c9e-56be-456e-b1da-4bc27e13de1a/cereals-balance-sheets-2017-18-and-forecast-2018-19.pdf (accessed on 22 November 2020).
- Rouf Shah, T.; Prasad, K.; Kumar, P. Maize—A potential source of human nutrition and health: A review. Cogent Food Agric. 2016, 2, 1166995. [Google Scholar] [CrossRef]
- Pannacci, E. Optimization of foramsulfuron doses for post-emergence weed control in maize (Zea mays L.). Span. J. Agric. Res. 2016, 14, e1005. [Google Scholar] [CrossRef]
- Gianessi, L.; Williams, A. High Maize Yields in Europe Depend on Herbicides. International Pesticide Benefits Case Study No. 14. Crop Protection Research Institute, CropLife Foundation. 2011. Available online: https://croplife.org/case-study/high-maize-yields-in-europe-depend-on-herbicides/ (accessed on 20 November 2020).
- Melander, B.; Rasmussen, I.A.; Barbéri, P. Integrating physical and cultural methods of weed control e examples from European research. Weed Sci. 2005, 53, 369–381. [Google Scholar] [CrossRef]
- Upadhyaya, M.K.; Blackshaw, R.E. Non-Chemical Weed Management. Principles, Concepts and Technology; CABI Press: Oxfordshire, UK, 2007; p. 239. [Google Scholar]
- Pannacci, E.; Tei, F. Effects of mechanical and chemical methods on weed control, weed seed rain and crop yield in maize, sunflower and soyabean. Crop Prot. 2014, 64, 51–59. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef]
- Novakoski, A.D.S.; Coelho, É.M.P.; Ravagnani, G.T.; Da Costa, A.C.P.R.; Rocha, S.A.; Zucareli, V.; Lopes, A.D. Allelopathic Potential of Plant Aqueous Mixtures on Euphorbia heterophylla. Agriculture 2020, 10, 449. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, NY, USA, 1984; p. 422. [Google Scholar]
- Weston, L.A.; Inderjit, S. Allelopathy: A potential tool in the development of strategies for biorational weed management. In Non-Chemical Weed Management–Principles, Concepts and Technology; Upadhyaya, M.K., Blackshaw, R.E., Eds.; CABI Press: Oxfordshire, UK, 2007; pp. 65–76. [Google Scholar]
- Findura, P.; Hara, P.; Szparaga, A.; Kocira, S.; Czerwińska, E.; Bartoš, P.; Nowak, J.; Treder, K. Evaluation of the Effects of Allelopathic Aqueous Plant Extracts, as Potential Preparations for Seed Dressing, on the Modulation of Cauliflower Seed Germination. Agriculture 2020, 10, 122. [Google Scholar] [CrossRef]
- Ehlert, K.A.; Mangold, J.M.; Engel, R.E. Integrating the herbicide imazapic and the fungal pathogen Pyrenophora semeniperda to control Bromus tectorum. Weed Res. 2014, 54, 418–424. [Google Scholar] [CrossRef]
- Pannacci, E.; Pettorossi, D.; Regni, L.; Tei, F. Allelopathic potential of mugwort (Artemisia vulgaris L.) to control the Italian ryegrass (Lolium multiflorum Lam.) in winter wheat. Allelopath. J. 2015, 36, 257–272. [Google Scholar]
- Pannacci, E.; Pettorossi, D.; Tei, F. Phytotoxic effects of aqueous extracts of sunflower on seed germination and growth of Sinapis alba L., Triticum aestivum L. and Lolium multiflorum Lam. Allelopath. J. 2013, 32, 23–36. [Google Scholar]
- Scavo, A.; Mauromicale, G. Integrated Weed Management in Herbaceous Field Crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowska, A. Allelochemicals as Bioherbicides—Present and Perspectives; InTech: London, UK, 2013; Chapter 20; pp. 517–542. Available online: http://cdn.intechopen.com/pdfs-wm/44466.pdf (accessed on 20 November 2020).
- Cai, X.; Gu, M. Bioherbicides in organic horticulture. Horticulturae 2016, 2, 3. [Google Scholar] [CrossRef]
- Bhadoria, P.B.S. Allelopathy: A natural way towards weed management. Am. J. Exp. Agric. 2011, 1, 7–20. [Google Scholar] [CrossRef]
- Ferreira, J.F.S.; Janick, J. Allelopathic Plants. XVI. Artemisia Species. Allelopath. J. 2004, 14, 167–176. [Google Scholar]
- Weston, L.A.; Barney, J.N.; DiTommaso, A. A review of the biology and ecology of three invasive perennials in New York State: Japanese knotweed (Polygonum cuspidatum), mugwort (Artemisia vulgaris) and pale swallow-wort (Vincetoxicum rossicum). Plant Soil 2005, 277, 53–69. [Google Scholar] [CrossRef]
- Duke, J.A.; Bogenshutz-Godwin, M.J.; du Cellier, M.J.; Duke, P.A. Handbook of Medicinal Herbs, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2002; p. 870. [Google Scholar]
- Marcinkeviciene, A.; Zita Kriauciuniene, Z.; Velicka, R.; Kosteckas, R.; Fujii, Y. Allelopathic effect of Artemisia vulgaris on winter wheat and winter oilseed rape. Fresen. Environ. Bull. 2018, 27, 727–732. [Google Scholar]
- Weston, L.A.; Duke, S.O. Weed and crop allelopathy. Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Barney, J.N.; Hay, A.G.; Weston, L.A. Isolation and characterization of allelopathic volatiles from mugwort (Artemisia vulgaris). J. Chem. Ecol. 2005, 31, 247–265. [Google Scholar] [CrossRef]
- Harker, K.N.; O’Donovan, J.T. Recent weed control, weed management, and integrated weed management. Weed Technol. 2013, 27, 1–11. [Google Scholar] [CrossRef]
- Sheibany, K.; Baghestani, M.A.; Atri, A. Competitive effects of redroot pigweed (Amaranthus retroflexus) on growth indices and yield of corn. Weed Biol. Manag. 2009, 9, 152–159. [Google Scholar] [CrossRef]
- Kenzevic, S.Z.; Weise, S.F.; Swanton, C.J. Interference of redroot pigweed (Amaranthus retroflexus) in corn (Zea mays L.). Weed Sci. 1994, 42, 568–573. [Google Scholar] [CrossRef]
- Bosnic, A.C.; Swanton, C.J. Influence of barnyard grass (Echinochloa crus-galli) time of emergence and density on corn (Zea mays). Weed Sci. 1997, 45, 276–282. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2020. Available online: http://www.weedscience.org (accessed on 10 November 2020).
- Stepanovic, S.; Datta, A.; Neilson, B.; Bruening, C.; Shapiro, C.; Gogos, G.; Knezevic, S.Z. Effectiveness of flame weeding and cultivation for weed control in organic maize. Biol. Agric. Hortic. 2016, 32, 47–62. [Google Scholar] [CrossRef]
- Cordeau, S.; Marion Triolet, M.; Wayman, S.; Steinberg, C.; Guillemin, J.-P. Bioherbicides: Dead in the water? A review of the existing products for integrated weed management. Crop Prot. 2016, 87, 44–49. [Google Scholar] [CrossRef]
- Masi, M.; Pannacci, E.; Santoro, E.; Zermane, N.; Superchi, S.; Evidente, A. Stoechanones a and b, phytotoxic copaane sesquiterpenoids isolated from Lavandula stoechas with potential herbicidal activity against Amaranthus retroflexus. J. Nat. Prod. 2020, 83, 1658–1665. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; XingXiang, G.; ZongJun, G.; Qing, W. A primary study on allelopathy of Artemisia vulgaris. Acta Pratacult. Sin. 2010, 19, 114–119. [Google Scholar]
- Kadioilu, I.; Yanar, Y. Allelopathic effects of plant extracts against seed germination of some weeds. Asian J. Plant Sci. 2004, 3, 472–475. [Google Scholar] [CrossRef]
- Inderjit, S.; Foy, C.L. Nature of interference mechanism of mugwort (Artemisia vulgaris). Weed Technol. 1999, 13, 176–182. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono-and Dicotyledonous Plants. BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany; Braunschweig, Germany, 2001; Available online: https://www.politicheagricole.it/flex/AppData/WebLive/Agrometeo/MIEPFY800/BBCHengl2001.pdf (accessed on 22 November 2020).
- Pannacci, E.; Onofri, A.; Covarelli, G. Biological activity, availability and duration of phytotoxicity for imazamox in four different soils of central Italy. Weed Res. 2006, 46, 243–250. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Streibig, J.C.; Rudemo, M.; Jensen, J.E. Dose-response curves and statistical models. In Herbicide Bioassay; Streibig, J.C., Kudsk, P., Eds.; CRC Press: Boca Raton, FL, USA, 1993; pp. 29–55. [Google Scholar]
- Brain, P.; Cousens, R. An Equation to Describe Dose Responses where there is Stimulation of Growth at Low Dose. Weed Res. 1989, 29, 93–96. [Google Scholar] [CrossRef]
- Pestemer, W.; Günther, P. Growth inhibition of plants as a bioassay for herbicide analysis. Chem. Plant Prot. 1995, 11, 219–231. [Google Scholar]
- Ritz, C.; Streibig, J.C. Bioassay Analysis using R. J. Stat. Softw. 2005, 12, 1–22. [Google Scholar] [CrossRef]
- Onofri, A.; Pannacci, E. Spreadsheet tools for biometry classes in crop science programmes. Commun. Biometry Crop. Sci. 2014, 9, 3–13. [Google Scholar]
- Pretsch, E.; Bühlmann, P.; Affolter, C. Structure Determination of Organic Compounds Tables of Spectral Data, 3rd ed.; Springer: Berlin, Germany, 2000; pp. 161–243. [Google Scholar]
- Berger, S.; Braun, S. 200 and More Basic NMR Experiments: A Practical Course, 1st ed.; Wiley-VCH: Weinheim, Germany, 2004; p. 854. [Google Scholar]
- Abiri, R.; Silva, A.L.M.; de Mesquita, L.S.S.; de Mesquita, J.W.C.; Atabaki, N.; de Almeida, E.B., Jr.; Shaharuddin, M.A.; Malik, S. Towards a better understanding of Artemisia vulgaris: Botany, phytochemistry, pharmacological and biotechnological potential. Food Res. Int. 2018, 109, 403–415. [Google Scholar] [CrossRef]
- Corrêa-Ferreira, M.L.; Verdan, M.H.; dos Reis Lívero, F.A.; Galuppo, L.F.; Telles, J.E.Q.; Stefanello, M.É.A.; Acco, A.; de Oliveira Petkowicz, C.L. Inulin-type fructan and infusion of Artemisia vulgaris protect the liver against carbon tetrachloride-induced liver injury. Phytomedicine 2017, 24, 68–76. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Lombardo, S.; Pesce, G.R.; Mauromicale, G. Allelopathic potential of leaf aqueous extracts from Cynara cardunculus L. on the seedling growth of two cosmopolitan weed species. Ital. J. Agron. 2019, 14, 1373. [Google Scholar] [CrossRef]
- Duke, S.O.; Cedergreen, N.; Velini, E.D.; Belz, R.G. Hormesis: Is it an important factor in herbicide use and allelopathy? Outlooks Pest Manag. 2006, 17, 29–33. [Google Scholar]
- Findura, P.; Kocira, S.; Hara, P.; Pawłowska, A.; Szparaga, A.; Kangalov, P. Extracts from Artemisia vulgaris L. in Potato Cultivation—Preliminary Research on Biostimulating Effect. Agriculture 2020, 10, 356. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Günther, P.; Rahman, A.; Pestemer, W. Quantitative bioassays for determining residues and availability to plants of sulphonylurea herbicides. Weed Res. 1989, 29, 141–146. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| pH Value | Organic Solvent | |||
|---|---|---|---|---|
| n-hexane | CH2Cl2 | EtOAc | n-butanol | |
| 7 | 1.7 mg | 17.2 mg | 23.5 mg | 20.3 mg |
| 2 | n.d. 1 | n.d. | 147.9 mg | n.d. |
| Species | Germination (%) | Radicle Lenght | Meso/Hypocotyl Length | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EC10 | EC50 | EC90 | EC10 | EC50 | EC90 | EC10 | EC50 | EC90 | ||||||||||
| A. retroflexus_1 | 1.2 | (0.06) | 3.1 | (0.06) | 8.1 | (0.37) | 2.9 | (0.03) | 4.2 | (0.02) | 6.2 | (0.03) | 4.4 | (4.00–4.55) | 4.8 | (4.58–4.84) | 5.7 | (5.45–6.10) |
| A. retroflexus_2 | 1.1 | (0.06) | 2.6 | (0.04) | 6.1 | (0.24) | 2.5 | (0.001) | 2.8 | (0.02) | 3.2 | (0.04) | 3.5 | (1.77–4.70) | 3.6 | (1.78–4.70) | 3.8 | (1.71–4.70) |
| Maize | >25 | >25 | >25 | 17.7 | (n.e.) | >25 | >25 | 18.8 | (3.50–31.56) | >25 | >25 | |||||||
| Species | Seedling Emergence | Plant Dry Weight | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EC10 | EC50 | EC90 | EC10 | EC50 | EC90 | |||||||
| A. retroflexus | 1.1 | (0.35) | 3.0 | (0.39) | 8.5 | (1.90) | 5.4 | (5.33–5.42) | 6.0 | (5.89–6.04) | 7.3 | (7.52–7.12) |
| Maize | >12.5 | >12.5 | >12.5 | >12.5 | >12.5 | >12.5 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pannacci, E.; Masi, M.; Farneselli, M.; Tei, F. Evaluation of Mugwort (Artemisia vulgaris L.) Aqueous Extract as a Potential Bioherbicide to Control Amaranthus retroflexus L. in Maize. Agriculture 2020, 10, 642. https://doi.org/10.3390/agriculture10120642
Pannacci E, Masi M, Farneselli M, Tei F. Evaluation of Mugwort (Artemisia vulgaris L.) Aqueous Extract as a Potential Bioherbicide to Control Amaranthus retroflexus L. in Maize. Agriculture. 2020; 10(12):642. https://doi.org/10.3390/agriculture10120642
Chicago/Turabian StylePannacci, Euro, Marco Masi, Michela Farneselli, and Francesco Tei. 2020. "Evaluation of Mugwort (Artemisia vulgaris L.) Aqueous Extract as a Potential Bioherbicide to Control Amaranthus retroflexus L. in Maize" Agriculture 10, no. 12: 642. https://doi.org/10.3390/agriculture10120642
APA StylePannacci, E., Masi, M., Farneselli, M., & Tei, F. (2020). Evaluation of Mugwort (Artemisia vulgaris L.) Aqueous Extract as a Potential Bioherbicide to Control Amaranthus retroflexus L. in Maize. Agriculture, 10(12), 642. https://doi.org/10.3390/agriculture10120642