Ancient Wheat Species (Triticum sphaerococcum Perc. and T. persicum Vav.) in Organic Farming: Influence of Sowing Density on Agronomic Traits, Pests and Diseases Occurrence, and Weed Infestation

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Crop Management
2.2. Experimental Design
2.3. Wheat Cultivars
2.4. Sampling and Measurements
2.4.1. Agronomic Traits
2.4.2. Pests Occurrence
2.4.3. Plant Diseases Occurrence
2.4.4. Weed Infestation
2.4.5. Statistical Analyses
3. Results
3.1. Agronomic Traits
3.2. Pests
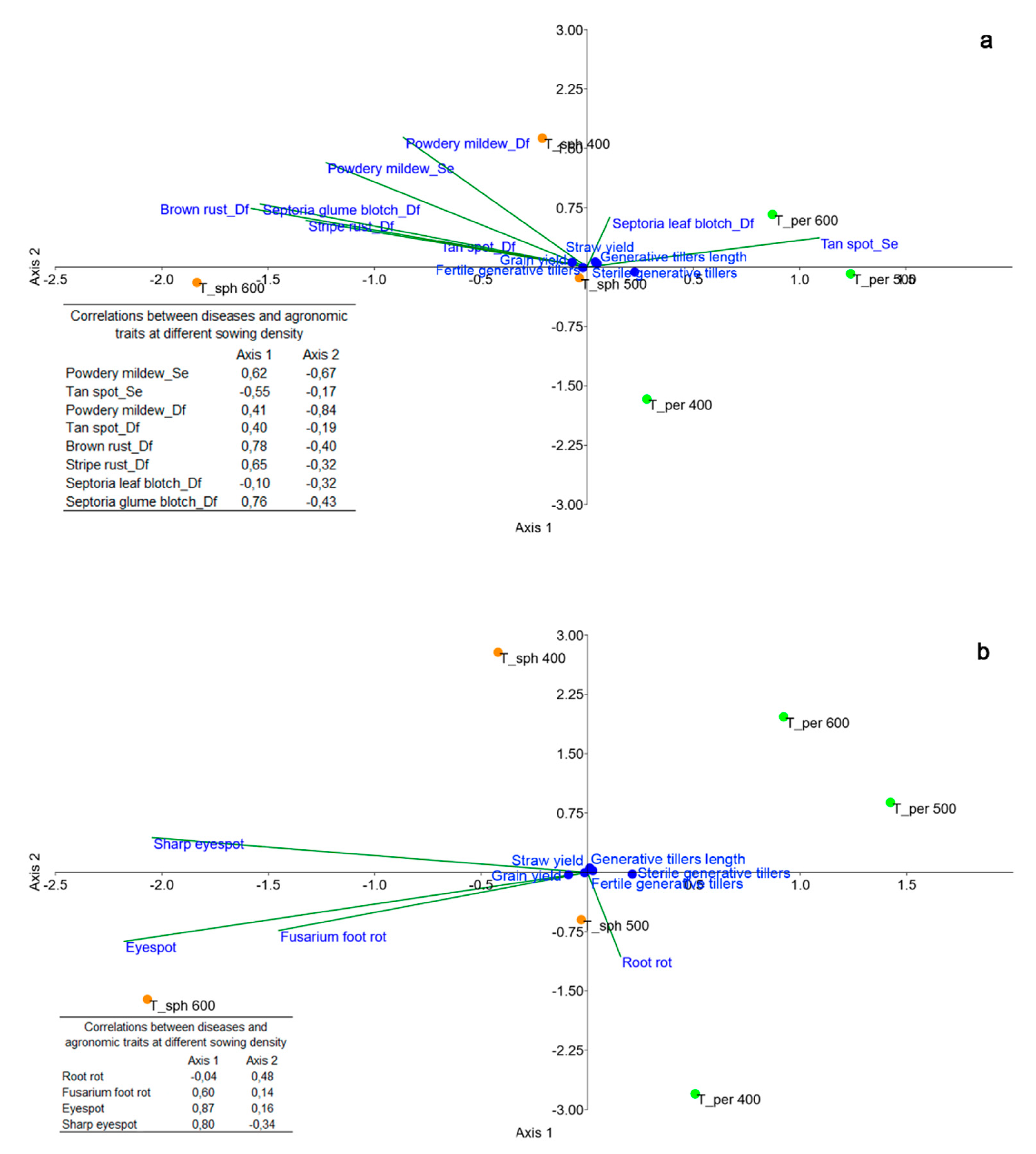
3.3. Diseases
3.4. Weeds
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- EUROSTAT. Available online: https://ec.europa.eu/eurostat (accessed on 10 March 2020).
- Bavec, F.; Bavec, M. Organic Production and Use of Alternative Crops; CRC/Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar]
- Kociszewski, K. The development of organic farming in Poland under the conditions of integration with the European Union. Econ. Environ. Stud. 2010, 10, 11–22. [Google Scholar]
- Arzani, A.; Ashraf, M. Cultivated ancient wheats (Triticum spp.): A Potential source of health-beneficial food products. Compr. Rev. Food Sci. Food Saf. 2017, 16, 477–488. [Google Scholar] [CrossRef]
- Szczepanek, M.; Prus, P.; Knapowski, T. The assessment of market demand for products obtained from primary wheat forms with increased nutritional value. In Proceedings of the 27th International Scientific Conference Agrarian Perspectives XXVII “Food Safety—FoodSecurity”, Prague, Czech Republic, 19–20 September 2018. [Google Scholar]
- Tyburski, J. Organic farming in Poland: Past, present and future perspectives. In Organic Agriculture: Sustainability, Markets, and Policies; OECD publishing: Paris, France, 2003; pp. 301–309. [Google Scholar]
- Suchowilska, E.; Wiwart, M.; Kandler, W.; Krska, R. A comparison of macro- and microelement concentrations in the whole grain of four Triticum species. Plant. Soil Environ. 2012, 58, 141–147. [Google Scholar] [CrossRef]
- Mori, N.; Ohta, S.; Chiba, H.; Takagi, T.; Niimi, Y.; Shinde, V.; Kajale, M.D.; Osada, T. Rediscovery of Indian dwarf wheat (Triticum aestivum L. ssp. sphaerococcum (Perc.) MK.) an ancient crop of the Indian subcontinent. Genet. Resour. Crop. Evol. 2013, 60, 1771–1775. [Google Scholar] [CrossRef]
- Matsuoka, Y. Evolution of polyploid triticum wheats under cultivation: The role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant. Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [PubMed]
- Josekutty, P.C. Defining the Genetic and Physiological Basis of Triticum sphaerococcum Perc. Master’s Thesis, University of Canterbury, Christchurch, New Zealand, 2008. [Google Scholar]
- Mosulishvili, M.; Bedoshvili, D.; Maisaia, I. A consolidated list of Triticum species and varieties of Georgia to promote repatriation of local diversity from foreign genebanks. Ann. Agrar. Sci. 2017, 15, 61–70. [Google Scholar] [CrossRef]
- Haque, M.A.; Takayama, A.; Watanabe, N.; Kuboyama, T. Cytological and genetic mapping of the gene for four-awned phenotype in Triticum carthlicum Nevski. Genet. Resour. Crop. Evol. 2010, 58, 1087–1093. [Google Scholar] [CrossRef]
- Zhuang, P.-P.; Ren, Q.-C.; Li, W.; Chen, G. Genetic diversity of persian wheat (Triticum turgidum ssp. carthlicum) Accessions by EST-SSR Markers. Am. J. Biochem. Mol. Biol. 2011, 1, 223–230. [Google Scholar] [CrossRef][Green Version]
- Brandsæter, L.O.; Mangerud, K.; Andersson, L.; Børresen, T.; Brodal, G.; Melander, B. Influence of mechanical weeding and fertilisation on perennial weeds, fungal diseases, soil structure and crop yield in organic spring cereals. Acta Agric. Scand. Sect. B Plant. Soil Sci. 2020, 70, 318–332. [Google Scholar] [CrossRef]
- Cyrkler-Degulis, M.; Bulińska-Radomska, Z. Yielding and healthiness of cultivars and populations of four winter wheat species under organic agriculture conditions. J. Res. Appl. Agric. Eng. 2006, 51, 17–21. [Google Scholar]
- Kolb, L.N.; Gallandt, E.R.; Mallory, E.B. Impact of spring wheat planting density, row spacing, and mechanical weed control on yield, grain protein, and economic return in maine. Weed Sci. 2012, 60, 244–253. [Google Scholar] [CrossRef]
- Stockmann, F.; Weber, E.A.; Merkt, N.; Schreiter, P.; Claupein, W.; Graeff-Hoenninger, S. Impact of row distance and seed density on grain yield, quality traits, and free asparagine of organically grown wheat. Agronomy 2019, 9, 713. [Google Scholar] [CrossRef]
- Beavers, R.L.; Hammermeister, A.M.; Frick, B.; Astatkie, T.; Martin, R.C. Spring wheat yield response to variable seeding rates in organic farming systems at different fertility regimes. Can. J. Plant. Sci. 2008, 88, 43–52. [Google Scholar] [CrossRef]
- Boström, U.; Anderson, L.E.; Wallenhammar, A.-C. Seed distance in relation to row distance: Effect on grain yield and weed biomass in organically grown winter wheat, spring wheat and spring oats. Field Crop. Res. 2012, 134, 144–152. [Google Scholar] [CrossRef]
- Kristensen, L.; Olsen, J.; Weiner, J. Crop density, sowing pattern, and nitrogen fertilization effects on weed suppression and yield in spring wheat. Weed Sci. 2008, 56, 97–102. [Google Scholar] [CrossRef]
- Lemerle, D.; Verbeek, B.; Cousens, R.; E Coombes, N. The potential for selecting wheat varieties strongly competitive against weeds. Weed Res. 1996, 36, 505–513. [Google Scholar] [CrossRef]
- Auskalniene, O.; Kadziene, G.; Jomantaite, B. The influence of crop density and sowing delay on weed germination in winter wheat. Julius-Kühn-Archiv 2018, 458, 306–309. [Google Scholar] [CrossRef]
- Haliniarz, M.; Kapeluszny, J. Assessment of the effect of sowing density on weed infestation and yields of three spring wheat cultivars. Acta Sci. Pol. Agric. 2012, 11, 13–25. [Google Scholar]
- Kaniuczak, Z.; Bereś, P. Occurrence and harmfulness of economically important cereals pests in ecological farms in Podkarpackie province in 2008–2010. J. Res. Appl. Agric. Eng. 2011, 56, 189–195. [Google Scholar]
- Kaniuczak, Z.; Bereś, P. The most harmful cereals pests in ecological farms of south-east Poland. J. Res. Appl. Agric. Eng. 2008, 53, 128–132. [Google Scholar]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases-a field perspective. Mol. Plant. Pathol. 2017, 19, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Ullah, M.; Ahmad, W.; Shah, S.U.A. The use of modern technologies to combat stripe rust in wheat. Rom. Biotechnol. Lett. 2020, 25, 1281–1288. [Google Scholar] [CrossRef]
- Lemańczyk, G. Effects of farming system, chemical control, fertilizer and sowing density on sharp eyespot and Rhizoctonia spp. in winter wheat. J. Plant. Prot. Res. 2012, 52, 381–396. [Google Scholar] [CrossRef]
- Baccar, R.; Fournier, C.; Dornbusch, T.; Andrieu, B.; Gouache, D.; Robert, C. Modelling the effect of wheat canopy architecture as affected by sowing density on Septoria tritici epidemics using a coupled epidemic–virtual plant model. Ann. Bot. 2011, 108, 1179–1194. [Google Scholar] [CrossRef]
- Sanguin, H.; Sarniguet, A.; Gazengel, K.; Moënne-Loccoz, Y.; Grundmann, G.L. Rhizosphere bacterial communities associated with disease suppressiveness stages of take-all decline in wheat monoculture. New Phytol. 2009, 184, 694–707. [Google Scholar] [CrossRef]
- Pradawne Ziarno. Available online: http://www.pradawneziarno.pl/ (accessed on 20 April 2020).
- Skrajda-Brdak, M.; Konopka, I.; Tańska, M.; Szczepanek, M.; Sadowski, T.; Rychcik, B. Low molecular phytochemicals of Indian dwarf (Triticum sphaerococcum Percival) and Persian wheat (T. carthlicum Nevski) grain. J. Cereal Sci. 2020, 91, 102887. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono-and Dicotyledonous Plants, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany, 2001; pp. 14–16. [Google Scholar]
- EPPO. Oulema spp. on cereals. Bull. OEPP/EPPO 2005, 35, 221–223.
- EPPO. Aphids on cereals. Bull. OEPP/EPPO 2007, 37, 29–32. [Google Scholar] [CrossRef]
- Walczak, F. Cereal Protection Guide; Wyd. IOR: Poznań, Poland, 2007; pp. 55–68. [Google Scholar]
- Bithell, S.L.; Butler, R.C.; McKay, A.C.; Cromey, M.G. Influences of crop sequence, rainfall and irrigation, on relationships between Gaeumannomyces graminis var. tritici and take-all in New Zealand wheat fields. Australas. Plant. Pathol. 2012, 42, 205–217. [Google Scholar] [CrossRef]
- Townsend, G.R.; Heuberger, J.W. Methods for estimating losses caused by diseases in fungicide experiments. Plant Dis. Rep. 1943, 27, 340–343. [Google Scholar]
- Douros, N.; Angelopoulou, F.; Papastylianou, P.; Papatheohari, Y.; Travlos, I.S.; Konstantas, A.; Bilalis, D. Building Organic Bridges, Proceedings of The 4th Isofar Scientific Conference at The Organic World Congress 2014, Istanbul, Turkey, 13–15 October 2014; Thünen: Braunschweig, Germany, 2014. [Google Scholar]
- Estefan, G.; Rolf Sommer, R.; Ryan, J. Methods of Soil, Plant, and Water Analysis: A Manual for the West Asia and North Africa Region, 3rd ed.; International Center for Agricultural Research in the Dry Areas (ICARDA): Beirut, Lebanon, 2013; p. 24. [Google Scholar]
- Berthet, P.; Gérard, G. A Statistical Study of Microdistribution of Oribatei (Acari) Part I. The Distribution Pattern. Oikos 1965, 16, 214. [Google Scholar] [CrossRef]
- Łomnicki, A. Introduction to Statistics for Naturalists, 4th ed.; PWN: Warszawa, Poland, 2010; p. 280. [Google Scholar]
- Stanisz, A. Easy Course of Statistic Using Statistica PL and Medicine Examples, 1. In Basic Statistic; StatSoft Polska: Kraków, Poland, 2006; p. 532. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electron. 2014, 1, 9. [Google Scholar]
- Hill, M.O.; Gauch, H.G. Detrended correspondence analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Leps, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; p. 269. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Science BV: Amsterdam, The Netherlands, 1998; p. 853. [Google Scholar]
- Legendre, P.; Oksanen, J.; Ter Braak, C.J.F. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Tokatlidis, I.S. Addressing the yield by density interaction is a prerequisite to bridge the yield gap of rain-fed wheat. Ann. Appl. Biol. 2014, 165, 27–42. [Google Scholar] [CrossRef]
- Poutala, R.; Korva, J.; Varis, E. Spring Wheat Cultivar Performance in Ecological and Conventional Cropping Systems. J. Sustain. Agric. 1993, 3, 63–84. [Google Scholar] [CrossRef]
- Lloveras, J.; Manent, J.; Viudas, J.; Lopez, A.; Santiveri, P. Seeding Rate Influence on Yield and Yield Components of Irrigated Winter Wheat in a Mediterranean Climate. Agron. J. 2004, 96, 1258–1265. [Google Scholar] [CrossRef]
- Tahir, S.A.; Khaliq, A.T.; Cheema, M.J.M. Evaluating the impact of seed rate and sowing dates on wheat productivity in semi-arid environment. Int. J. Agric. Biol. 2019, 22, 57–64. [Google Scholar] [CrossRef]
- Gałęzewski, M. Pests occurrence on spring cereal mixtures. Prog. Plant Prot./Post. Ochr. Roślin 2008, 48, 831–835. [Google Scholar]
- Beres, B.L.; Cárcamo, H.; Yang, R.-C.; Spaner, D. Integrating spring wheat sowing density with variety selection to manage wheat stem sawfly. Agron. J. 2011, 103, 1755–1764. [Google Scholar] [CrossRef]
- Lamparski, R.; Szczepanek, M. Entomophagous insects of the red fescue in the various condition of nitrogen fertilization. Prog. Plant Prot./Post. Ochr. Roślin 2008, 48, 357–361. [Google Scholar]
- Lemańczyk, G.; Piekarczyk, M. Effect of fertilization, chemical crop protection and sowing density on health status of winter wheat grown in short-time monoculture on light soil. Prog. Plant Prot./Post. Ochr. Roślin 2013, 53, 487–493. [Google Scholar]
- Robert, C.; Garin, G.; Abichou, M.; Houlès, V.; Pradal, C.; Fournier, C. Plant architecture and foliar senescence impact the race between wheat growth and Zymoseptoria tritici epidemics. Ann. Bot. 2018, 121, 975–989. [Google Scholar] [CrossRef] [PubMed]
- Finckh, M.R.; Gacek, E.S.; Czembor, H.J.; Wolfe, M.S. Host frequency and density effects on powdery mildew and yield in mixtures of barley cultivars. Plant. Pathol. 1999, 48, 807–816. [Google Scholar] [CrossRef]
- Beres, B.L.; Turkington, T.K.; Kutcher, H.R.; Irvine, B.; Johnson, E.N.; O’Donovan, J.T.; Harker, K.N.; Holzapfel, C.B.; Mohr, R.; Peng, G.; et al. Winter wheat cropping system response to seed treatments, seed size, and sowing density. Agron. J. 2016, 108, 1101–1111. [Google Scholar] [CrossRef]
- McDonald, B.A.; Mundt, C.C. How Knowledge of pathogen population biology informs management of septoria tritici blotch. Phytopathology 2016, 106, 948–955. [Google Scholar] [CrossRef]
- Morais, D.; Sache, I.; Suffert, F.; Laval, V. Is the onset of septoria tritici blotch epidemics related to the local pool of ascospores? Plant. Pathol. 2015, 65, 250–260. [Google Scholar] [CrossRef]
- Walters, D. Disease Control in Crops: Biological and Environmentally Friendly Approaches; John Wiley & Sons: Edinburgh, UK, 2009; pp. 1–226. [Google Scholar]
- Eken, C.; Bulut, S.; Genc, T.; Öztürk, A. Effects of different fertilizer sources and sowing density on root and crown rot disease agents of organic wheat. J. Agric. Fac. Gaziosmanpasa Univ. 2014, 31, 12–19. [Google Scholar] [CrossRef]
- Colbach, N.; Lucas, P.; Meynard, J.-M. Influence of Crop Management on take-all development and disease cycles on winter w heat. Phytopathology 1997, 87, 26–32. [Google Scholar] [CrossRef]
- Bhaskar, A.V.; Davies, W.; Cannon, N.; Conway, J. Weed manifestation under different tillage and legume undersowing in organic wheat. Biol. Agric. Hortic. 2014, 30, 253–263. [Google Scholar] [CrossRef]
- Feledyn-Szewczyk, B. The effectiveness of weed regulation methods in spring wheat cultivated in integrated, conventional and organic crop production systems. J. Plant. Prot. Res. 2012, 52, 486–493. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Location | N-NO3 | N-NH4 | pH | P | K | Mg | C-org. |
|---|---|---|---|---|---|---|---|
| mg kg−1 of Soil | (KCl) | mg kg−1 of Soil | % | ||||
| Kiełpin | 6.26 | 2.65 | 5.6 | 36.2 | 111.2 | 6.4 | 1.71 |
| Trzcińsk | 6.83 | 10.9 | 4.9 | 84.1 | 137.0 | 5.6 | 3.99 |
| Grabina Wielka | 8.64 | 9.99 | 5.9 | 64.5 | 182.6 | 5.7 | 1.63 |
| Location | 2019 | 1981–2010 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| March | April | May | June | July | March | April | May | June | July | |
| Temperature °C | ||||||||||
| Kiełpin | 4.9 | 9.0 | 11.3 | 20.4 | 17.4 | 2.0 | 7.2 | 12.5 | 15.1 | 17.6 |
| Trzcińsk | 5.8 | 8.6 | 11.9 | 20.0 | 18.0 | 1.9 | 7.1 | 12.4 | 15.3 | 18.0 |
| Grabina Wielka | 6.1 | 10.1 | 12.5 | 22.1 | 19.2 | 3.2 | 8.6 | 13.9 | 16.5 | 18.9 |
| Precipitation mm | ||||||||||
| Kiełpin | 46.0 | 3.0 | 69.4 | 63.3 | 39.5 | 41.3 | 32.9 | 57.2 | 62.7 | 70.3 |
| Trzcińsk | 44.0 | 2.0 | 65.0 | 63.0 | 59.0 | 30.0 | 30.0 | 54.0 | 67.0 | 73.0 |
| Grabina Wielka | 35.4 | 14.6 | 44.7 | 23.7 | 50.7 | 32,8 | 31.1 | 52.9 | 57.6 | 77.2 |
| Species | 1000-Grain Weight (g) | Germination Capacity (%) | Theoretical Crop Establishment (%) | Sowing Density (no m−2) | Seeding Rate (kg ha−1) |
|---|---|---|---|---|---|
| T. sphaerococcum | 31.3 | 94 | 90 | 400 | 148 |
| 500 | 185 | ||||
| 600 | 222 | ||||
| T. persicum | 30.2 | 84 | 90 | 400 | 160 |
| 500 | 200 | ||||
| 600 | 240 |
| Species | Sowing Density | Plant Density (no m−2) | Generative Tillers (no plant−1) | Vegetative Tillers (no plant−1) | Generative and Vegetative Tillers (no plant−1) |
|---|---|---|---|---|---|
| T. sphaerococcum | 400 | 355 ± 45 | 1.31 ± 0.12 | 0.77 ± 0.67 | 2.08 ± 0.43 |
| 500 | 439 ± 53 | 1.17 ± 0.11 | 0.67 ± 0.27 | 1.84 ± 0.35 | |
| 600 | 490 ± 103 | 1.21 ± 0.11 | 0.55 ± 0.21 | 1.77 ± 0.25 | |
| T. persicum | 400 | 342 ± 48 | 1.46 ± 0.28 | 0.71 ± 0.38 | 2.16 ± 0.61 |
| 500 | 455 ± 68 | 1.30 ± 0.19 | 0.85 ± 0.34 | 2.14 ± 0.43 | |
| 600 | 536 ± 42 | 1.32 ± 0.20 | 0.87 ± 0.30 | 2.19 ± 0.40 | |
| p-value | 0.271 | 0.913 | 0.068 | 0.275 | |
| T. sphaerococcum | 428 ± 90 | 1.23 ± 0.13 b 1 | 0.66 ± 0.31 b | 1.90 ± 0.37 b | |
| T. persicum | 444 ± 96 | 1.36 ± 0.23 a | 0.81 ± 0.34 a | 2.17 ± 0.47 a | |
| p-value | 0.290 | 0.002 | 0.031 | 0.003 |
| Species | Sowing Density | Grain Yield (Mg ha−1) | Straw Yield (Mg ha−1) | Fertile Generative Tillers (no m−2) | Sterile Generative Tillers (no m−2) | Total Generative Tillers (no m−2) | Generative Tillers Length (cm) |
|---|---|---|---|---|---|---|---|
| T.sphaerococcum | 400 | 2.70 ± 0.82 b 1 | 5.29 ± 2.77 | 410 ± 65 ab | 15.0 ± 12.7 | 425 ± 66 ab | 90.2 ± 12.7 |
| 500 | 2.83 ± 0.94 ab | 4.75 ± 2.07 | 427 ± 97 ab | 18.3 ± 10.2 | 446 ± 100 ab | 86.8 ± 11.4 | |
| 600 | 2.97 ± 1.00 a | 5.23 ± 2.20 | 488 ± 102 a | 12.0 ± 8.5 | 500 ± 102 a | 87.1 ± 11.4 | |
| T. persicum | 400 | 2.25 ± 0.79 c | 4.70 ± 1.95 | 453 ± 76 ab | 22.3 ± 15.7 | 475 ± 73 ab | 86.8 ± 5.8 |
| 500 | 2.09 ± 0.80 c | 5.16 ± 2.09 | 393 ± 70 b | 22.0 ± 14.8 | 415 ± 69 ab | 86.9 ± 6.6 | |
| 600 | 2.11 ± 0.76 c | 4.96 ± 1.66 | 381 ± 42 b | 19.1 ± 15.0 | 400 ± 53 b | 85.7 ± 6.7 | |
| p-value | 0.006 | 0.176 | 0.003 | 0.774 | 0.003 | 0.427 | |
| T. phaerococcum | 2.83 ± 0.91 a | 5.09 ± 2.31 | 442 ± 94 | 15.1 ± 10.6 b | 457 ± 94 | 88.0 ± 11.6 | |
| T. persicum | 2.15 ± 0.76 b | 4.94 ± 1.86 | 409 ± 70 | 21.1 ± 14.8 a | 430 ± 72 | 86.5 ± 6.2 | |
| p-value | <0.001 | 0.293 | 0.056 | 0.013 | 0.125 | 0.143 |
| Species | Sowing Density | Ear Length (cm) | Fertile Spikelets (no ear−1) | Sterile Spikelets (no ear−1) | Spikelets-Total (no ear−1) | Number of Grain per ear | 1000-Grain Weight (g) |
|---|---|---|---|---|---|---|---|
| T. sphaerococcum | 400 | 5.93 ± 0.52 | 13.1 ± 0.7 bc 1 | 5.8 ± 1.1 a | 18.9 ± 1.4 a | 25.1 ± 2.2 bc | 27.8 ± 3.1 |
| 500 | 6.13 ± 0.66 | 13.7 ± 1.2 ab | 5.2 ± 0.7 ab | 18.9 ± 1.6 a | 27.3 ± 2.4 ab | 28.3 ± 5.3 | |
| 600 | 6.33 ± 0.52 | 14.3 ± 1.1 a | 4.9 ± 0.6 b | 19.3 ± 1.0 a | 29.4 ± 3.0 a | 29.2 ± 4.8 | |
| T. persicum | 400 | 7.87 ± 0.98 | 12.4 ± 1.3 cd | 2.3 ± 0.4 c | 14.6 ± 1.4 b | 24.5 ± 4.3 c | 27.6 ± 4.2 |
| 500 | 8.02 ± 0.81 | 11.8 ± 1.4 d | 2.7 ± 0.3 c | 14.5 ± 1.4 b | 22.7 ± 3.0 c | 28.1 ± 3.7 | |
| 600 | 8.01 ± 0.54 | 11.8 ± 0.7 d | 2.3 ± 0.6 c | 14.1 ± 0.8 b | 24.0 ± 3.6 c | 27.6 ± 5.4 | |
| p-value | 0.460 | <0.001 | 0.001 | 0.012 | 0.001 | 0.266 | |
| T. sphaerococcum | 6.13 ± 0.58 b | 13.7 ± 1.1 a | 5.3 ± 0.9 a | 19.0 ± 1.5 a | 27.2 ± 3.2 a | 28.5 ± 4.4 | |
| T. persicum | 7.97 ± 0.78 a | 12.0 ± 1.2 b | 2.4 ± 0.5 b | 14.4 ± 1.2 b | 23.7 ± 3.4 b | 27.8 ± 4.4 | |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.094 |
| Species | Sowing Density | Oulema spp. Larvae | Aphididae | Oulema spp. Adults | Oulema spp. Larvae | Oulema spp. Larvae | Aphididae | Oulema spp. Adults | Oulema spp. Larvae |
|---|---|---|---|---|---|---|---|---|---|
| (no per 25 Shoots) | Damaged Plant Surface (%) | (no per 25 Shoots) | Damaged Plant Surface (%) | ||||||
| Flag Leaf Stage (BBCH 39) | Flowering (BBCH 65) | ||||||||
| T. sphaerococcum | 400 | 4.00 ± 3.28 a 1 | 0.25 ± 0.45 a | 2.83 ± 3.19 a | 3.25 ± 2.73 a | 0.17 ± 0.39 | 1.17 ± 1.19 | 5.08 ± 2.35 | 7.08 ± 6.26 a |
| 500 | 2.42 ± 1.98 b | 0.00 ± 0.00 b | 1.75 ± 1.71 b | 2.00 ± 1.60 b | 0.08 ± 0.29 | 1.00 ± 1.04 | 5.08 ± 2.19 | 5.58 ± 4.08 b | |
| 600 | 1.67 ± 0.78 b | 0.00 ± 0.00 b | 1.08 ± 0.67 b | 2.50 ± 0.90 b | 0.17 ± 0.39 | 1.08 ± 1.16 | 4.50 ± 1.93 | 4.92 ± 1.88 b | |
| T. persicum | 400 | 0.33 ± 0.49 c | 0.00 ± 0.00 b | 1.33 ± 1.15 b | 0.42 ± 0.67 c | 0.0 ± 0.0 | 0.50 ± 0.67 | 1.25 ± 0.62 | 1.67 ± 1.67 c |
| 500 | 0.08 ± 0.29 c | 0.08 ± 0.29 b | 1.00 ± 0.85 b | 0.0 ± 0.0 c | 0.0 ± 0.0 | 0.08 ± 0.29 | 0.58 ± 0.67 | 1.08 ± 1.16 c | |
| 600 | 0.33 ± 0.49 c | 0.00 ± 0.00 b | 1.08 ± 1.16 b | 0.08 ± 0.29 c | 0.0 ± 0.0 | 0.08 ± 0.29 | 0.58 ± 0.51 | 1.17 ± 0.83 c | |
| p-value | <0.001 | 0.026 | 0.001 | 0.042 | 0.762 | 0.484 | 0.169 | 0.045 | |
| T. sphaerococcum | 2.69 ± 2.40 a | 0.08 ± 0.28 | 1.89 ± 2.19 a | 2.58 ± 1.92 a | 0.14 ± 0.35 a | 1.08 ±1.11 a | 4.89 ± 2.12 a | 5.86 ± 4.42 a | |
| T. persicum | 0.25 ± 0.44 b | 0.03 ± 0.17 | 1.14 ± 1.05 b | 0.17 ± 0.45 b | 0.00 ± 0.00 b | 0.22 ± 0.48 b | 0.81 ± 0.67 b | 1.31 ± 1.26 b | |
| p-value | <0.001 | 0.310 | <0.001 | <0.001 | 0.020 | <0.001 | <0.001 | <0.001 | |
| Species | Sowing Density | Powdery Mildew | Tan Spot | Powdery Mildew | Tan Spot | Brown Rust | Stripe Rust | Septoria Leaf Blotch | Septoria Glume Blotch |
|---|---|---|---|---|---|---|---|---|---|
| Leaf or Ear Area with Disease Symptoms (%) | |||||||||
| Stem Elongation (BBCH 35–37) | Development of Fruit (BBCH 75–77) | ||||||||
| T. sphaerococcum | 400 | 26.8 ± 29.4 | 5.58 ± 3.84 b 1 | 1.00 ± 1.30 a | 7.67 ± 5.72 bc | 4.53 ± 5.00 | 0.31 ± 0.52 | 0.33 ± 0.62 | 9.78 ± 7.68 |
| 500 | 25.8 ± 28.2 | 4.33 ± 3.46 b | 0.61 ± 0.97 ab | 7.57 ± 5.42 bc | 4.48 ± 3.93 | 0.47 ± 0.78 | 0.44 ± 0.81 | 9.67 ± 7.33 | |
| 600 | 27.9 ± 29.5 | 4.96 ± 3.32 b | 0.63 ± 0.97 ab | 10.11 ± 7.42 a | 4.73 ± 4.33 | 0.33 ± 0.79 | 0.11 ± 0.17 | 9.92 ± 8.62 | |
| T. persicum | 400 | 22.5 ± 21.2 | 6.31 ± 5.60 b | 0.28 ± 0.54 b | 6.90 ± 8.66 c | 0.87 ± 0.74 | 0.0 ± 0.0 | 0.14 ± 0.26 | 0.0 ± 0.0 |
| 500 | 23.4 ± 20.6 | 6.44 ± 4.86 b | 0.40 ± 0.66 b | 9.26 ± 12.32 ab | 0.72 ± 0.54 | 0.0 ± 0.0 | 0.21 ± 0.41 | 0.0 ± 0.0 | |
| 600 | 26.8 ± 21.9 | 10.84 ± 9.34 a | 0.47 ± 0.62 b | 8.46 ± 11.02 abc | 0.69 ± 0.54 | 0.0 ± 0.0 | 0.11 ± 0.19 | 0.0 ± 0.0 | |
| p-value | 0.803 | <0.001 | 0.028 | 0.012 | 0.910 | 0.669 | 0.369 | 0.915 | |
| T. sphaerococcum | 26.8 ± 28.2 | 4.96 ± 3.48 b | 0.75 ± 1.07 a | 8.45 ± 6.18 | 4.58 ± 4.31 a | 0.37 ± 0.69 a | 0.30 ± 0.60 | 9.79 ± 7.67 a | |
| T. persicum | 24.3 ± 20.7 | 7.86 ± 7.02 a | 0.38 ± 0.60 b | 8.21 ± 10.51 | 0.76 ± 0.60 b | 0.00 ± 0.00 b | 0.15 ± 0.29 | 0.00 ± 0.00 b | |
| p-value | 0.195 | <0.001 | <0.001 | 0.598 | <0.001 | <0.001 | 0.054 | <0.001 | |
| Species | Sowing Density | Root Rot | Fusarium Foot Rot | Eyespot | Sharp Eyespot | ||||
|---|---|---|---|---|---|---|---|---|---|
| DI | % | DI | % | DI | % | DI | % | ||
| T. sphaerococcum | 400 | 7.00 ± 4.00 | 28.0 ± 16.0 | 1.17 ± 0.94 | 4.67 ± 3.75 | 9.7 ± 3.45 | 38.0 ± 14.2 | 0.17 ± 0.39 | 0.67 ± 1.56 |
| 500 | 8.25 ± 3.93 | 32.7 ± 15.9 | 2.33 ± 1.78 | 9.33 ± 7.10 | 10.3 ± 5.93 | 40.7 ± 22.6 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| 600 | 7.75 ± 3.96 | 31.0 ± 15.8 | 2.08 ± 2.50 | 8.33 ± 10.01 | 10.9 ± 6.68 | 42.7 ± 25.3 | 0.17 ± 0.39 | 0.67 ± 1.56 | |
| T. persicum | 400 | 8.75 ± 4.18 | 35.0 ± 16.7 | 0.75 ± 1.29 | 3.00 ± 5.15 | 8.2 ± 4.61 | 32.7 ± 18.4 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 500 | 6.83 ± 3.95 | 27.0 ± 15.3 | 1.33 ± 2.53 | 5.33 ± 10.14 | 5.5 ± 4.32 | 22.0 ± 17.3 | 0.0 ± 0.0 | 0.0 ± 0.0 | |
| 600 | 8.83 ± 4.73 | 35.3 ± 18.9 | 0.50 ± 0.67 | 2.00 ± 2.70 | 6.1 ± 4.70 | 24.3 ± 18.8 | 0.0 ± 0.0 | 0.0 ± 0.0 | |
| p-value | 0.283 | 0.278 | 0.431 | 0.431 | 0.235 | 0.235 | 0.232 | 0.232 | |
| T. sphaerococcum | 7.67 ± 3.88 | 30.6 ± 15.6 | 1.86 ± 1.87 a 1 | 7.44 ± 7.48 a | 10.3 ± 5.39 a | 40.4 ± 20.7 a | 0.11 ± 0.32 a | 0.44 ± 1.27 a | |
| T. persicum | 8.14 ± 4.28 | 32.4 ± 17.0 | 0.86 ± 1.68 b | 3.44 ± 6.70 b | 6.6 ± 4.56 b | 26.3 ± 18.3 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | |
| p-value | 0.580 | 0.577 | 0.008 | 0.008 | <0.001 | <0.001 | 0.018 | 0.018 | |
| Species | Sowing Density | Dry Matter of Weeds (g m−2) | Number of Weeds (no m−2) |
|---|---|---|---|
| T. sphaerococcum | 400 | 61.1 ± 46.3 | 57.9 ± 44.4 |
| 500 | 60.9 ± 46.2 | 49.0 ± 42.1 | |
| 600 | 57.4 ± 44.4 | 52.4 ± 41.9 | |
| T. persicum | 400 | 91.9 ± 78.0 | 59.2 ± 43.1 |
| 500 | 75.8 ± 68.4 | 51.0 ± 38.4 | |
| 600 | 62.4 ± 49.8 | 50.3 ± 38.2 | |
| p-value | 0.280 | 0.865 | |
| T. sphaerococcum | 59.8 ± 44.3 b 1 | 53.1 ± 41.7 | |
| T. persicum | 79.7 ± 65.7 a | 53.5 ± 39.0 | |
| p-value | 0.013 | 0.906 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczepanek, M.; Lemańczyk, G.; Lamparski, R.; Wilczewski, E.; Graczyk, R.; Nowak, R.; Prus, P. Ancient Wheat Species (Triticum sphaerococcum Perc. and T. persicum Vav.) in Organic Farming: Influence of Sowing Density on Agronomic Traits, Pests and Diseases Occurrence, and Weed Infestation. Agriculture 2020, 10, 556. https://doi.org/10.3390/agriculture10110556
Szczepanek M, Lemańczyk G, Lamparski R, Wilczewski E, Graczyk R, Nowak R, Prus P. Ancient Wheat Species (Triticum sphaerococcum Perc. and T. persicum Vav.) in Organic Farming: Influence of Sowing Density on Agronomic Traits, Pests and Diseases Occurrence, and Weed Infestation. Agriculture. 2020; 10(11):556. https://doi.org/10.3390/agriculture10110556
Chicago/Turabian StyleSzczepanek, Małgorzata, Grzegorz Lemańczyk, Robert Lamparski, Edward Wilczewski, Radomir Graczyk, Rafał Nowak, and Piotr Prus. 2020. "Ancient Wheat Species (Triticum sphaerococcum Perc. and T. persicum Vav.) in Organic Farming: Influence of Sowing Density on Agronomic Traits, Pests and Diseases Occurrence, and Weed Infestation" Agriculture 10, no. 11: 556. https://doi.org/10.3390/agriculture10110556
APA StyleSzczepanek, M., Lemańczyk, G., Lamparski, R., Wilczewski, E., Graczyk, R., Nowak, R., & Prus, P. (2020). Ancient Wheat Species (Triticum sphaerococcum Perc. and T. persicum Vav.) in Organic Farming: Influence of Sowing Density on Agronomic Traits, Pests and Diseases Occurrence, and Weed Infestation. Agriculture, 10(11), 556. https://doi.org/10.3390/agriculture10110556