Diet: A Specific Part of the Western Lifestyle Pack in the Asthma Epidemic



{kind=link}
Abstract
1. Introduction
2. Diet and the Parallel Epidemics of Obesity and Asthma
2.1. Role of Micronutrients and Macronutrients
2.2. Role of Food Groups and Dietary Patterns
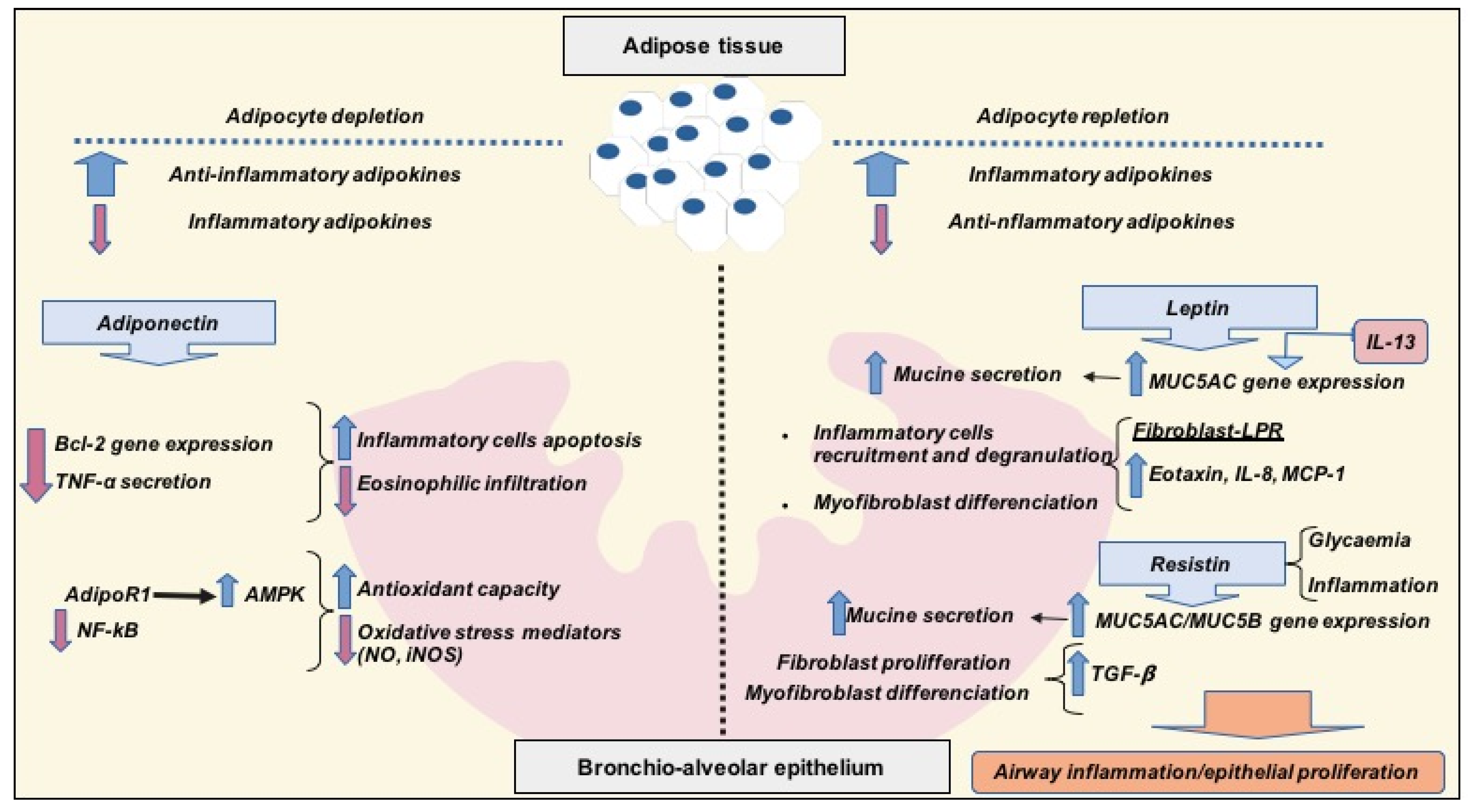
3. Obesity-Related Asthma and Interrelations with Diet, Inflammation and Adipose Tissue
3.1. Anti-Inflammatory Adipokines: Adiponectin
3.2. Pro-Inflammatory Adipokines: Leptin and Resistin
4. Concluding Remarks and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Christ, A.; Latz, E. The Western lifestyle has lasting effects on metaflammation. Nat. Rev. Immunol. 2019, 19, 267–268. [Google Scholar] [CrossRef]
- Periyalil, H.A.; Wood, L.G.; Wright, T.A.; Karihaloo, C.; Starkey, M.R.; Miu, A.S.; Baines, K.J.; Hansbro, P.M.; Gibson, P.G. Obese asthmatics are characterized by altered adipose tissue macrophage activation. Clin. Exp. Allergy 2018, 48, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Sin, D.D.; Sutherland, E.R. Obesity and the lung: 4. Obesity and asthma. Thorax 2008, 63, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- Rush, E.C.; Yan, M.R. Evolution not Revolution: Nutrition and Obesity. Nutrients 2017, 9, 519. [Google Scholar] [CrossRef] [PubMed]
- Braback, L.; Hjern, A.; Rasmussen, F. Trends in asthma, allergic rhinitis and eczema among Swedish conscripts from farming and non-farming environments. A nationwide study over three decades. Clin. Exp. Allergy 2004, 34, 38–43. [Google Scholar] [CrossRef]
- Haahtela, T.; Lindholm, H.; Bjorksten, F.; Koskenvuo, K.; Laitinen, L.A. Prevalence of asthma in Finnish young men. BMJ 1990, 301, 266–268. [Google Scholar] [CrossRef]
- Eder, W.; Ege, M.J.; von Mutius, E. The asthma epidemic. N. Engl. J. Med. 2006, 355, 2226–2235. [Google Scholar] [CrossRef]
- García-Marcos, L.; Quiros, A.B.; Hernández, G.G.; Guillen-Grima, F.; Díaz, C.G.; Ureña, I.C.; Peña, A.A.; Monge, R.B.; Suarez-Varela, M.M.; Varela, A.L.; et al. Stabilization of asthma prevalence among adolescents and increase among schoolchildren (ISAAC phases I and III) in Spain. Allergy 2004, 59, 1301–1307. [Google Scholar] [CrossRef]
- Asher, M.I.; Montefort, S.; Bjorksten, B.; Lai, C.K.; Strachan, D.P.; Weiland, S.K.; Williams, H. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC Phases One and Three repeat multicountry cross-sectional surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef]
- Hales, C.M.; Fryar, C.D.; Carroll, M.D.; Freedman, D.S.; Ogden, C.L. Trends in Obesity and Severe Obesity Prevalence in US Youth and Adults by Sex and Age, 2007–2008 to 2015–2016. JAMA 2018, 319, 1723–1725. [Google Scholar] [CrossRef]
- Ogden, C.L.; Fryar, C.D.; Hales, C.M.; Carroll, M.D.; Aoki, Y.; Freedman, D.S. Differences in Obesity Prevalence by Demographics and Urbanization in US Children and Adolescents, 2013–2016. JAMA 2018, 319, 2410–2418. [Google Scholar] [CrossRef] [PubMed]
- Hallstrand, T.S.; Fischer, M.E.; Wurfel, M.M.; Afari, N.; Buchwald, D.; Goldberg, J. Genetic pleiotropy between asthma and obesity in a community-based sample of twins. J. Allergy Clin. Immunol. 2005, 116, 1235–1241. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Thomsen, S.F.; Ulrik, C.S.; Kyvik, K.O.; Sørensen, T.I.; Posthyma, D.; Skadhauge, L.R.; Steffensen, I.; Backer, V. Association between obesity and asthma in a twin cohort. Allergy 2007, 62, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Marcos, L.; Canflanca, I.M.; Garrido, J.B.; Varela, A.L.; Garcia-Hernandez, G.; Guillen-Grima, F.; Gonzalez-Diaz, C.; Carbajal-Ureña, I.; Arnedo-Peña, A.; Busquets-Monge, R.M.; et al. Relationship of asthma and rhinoconjunctivitis with obesity, exercise and Mediterranean diet in Spanish schoolchildren. Thorax 2007, 62, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Marcos, L.; Castro-Rodriguez, J.A.; Weinmayer, G.; Panagiotakos, D.B.; Priftis, K.N.; Nagel, G. Influence of Mediterranean diet on asthma in children: A systematic review and meta-analysis. Pediatr. Allergy Immunol. 2013, 24, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Ellwood, P.; Asher, M.I.; Garcia-Marcos, L.; Williams, H.; Keil, U.; Robertson, C.; Nagel, G. Do fast foods cause asthma, rhinoconjunctivitis and eczema? Global findings from the International Study of Asthma and Allergies in Childhood (ISAAC) phase three. Thorax 2013, 68, 351–360. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Panagiotakos, D.B.; Hatziagorou, E.; Antonogeorgos, G.; Matziou, V.N.; Tsanakas, J.N.; Gratziou, C.; Tsabouri, S.; Priftis, K.N. Antioxidant foods consumption and childhood asthma and other allergic diseases: The Greek cohorts of the ISAAC II survey. Allergol. Immunopathol. (Madr) 2015, 43, 353–360. [Google Scholar] [CrossRef]
- Sahiner, U.M.; Birben, E.; Erzurum, S.; Sackesen, C.; Kalayci, O. Oxidative stress in asthma: Part of the puzzle. Pediatr. Allergy Immunol. 2018, 29, 789–800. [Google Scholar] [CrossRef]
- Wall, C.R.; Stewart, A.W.; Hancox, R.J.; Murphy, R.; Braithwaite, I.; Beasley, R.; Mitchell, E.A. Association between frequency counsumption of fruit, vegetables, nuts and pulses and BMI: Alayses of the International study of Asthma and Allergies in Childhood. Nutrients 2018, 10, 316. [Google Scholar] [CrossRef]
- Amati, F.; Hassounah, S.; Swaka, A. The Impact of Mediterranean Dietary Patterns During Pregnancy on Maternal and Offspring Health. Nutrients 2019, 11, 1098. [Google Scholar] [CrossRef]
- Forno, E.; Young, O.M.; Kumar, R.; Simhan, H.; Celedon, J.C. Maternal obesity in pregnancy, gestional weight gain and risk of childhood asthma. Pediatrics 2014, 134, e535–e546. [Google Scholar] [CrossRef] [PubMed]
- Tie, H.T.; Xia, Y.Y.; Zeng, Y.S.; Zhang, Y.; Dai, C.L.; Guo, J.J.; Zhao, Y. Risk of childhood overweight or obesity associated with excessive weight gain during pregnancy: A meta-analysis. Arch. Gynecol. Obstet. 2014, 289, 247–257. [Google Scholar] [CrossRef]
- Nagel, G.; Weinmayr, G.; Kleiner, A.; García-Marcos, L.; Strachan, D.P. Effect of diet on asthma and allergic sensitization in the International Studies on Allergies and Asthma in Childhood (ISAAC) Phase Two. Thorax 2010, 65, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Alwarith, J.; Kahleova, H.; Crosby, L.; Brooks, A.; Brandon, L.; Levin, S.M.; Barnard, N.D. The role of nutrition in asthma prevention and treatment. Nutr. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.B.; Martínez-González, M.A.; Fung, T.T.; Lichtenstein, A.H.; Forouhi, N.H. Food based dietary patterns and chronic disease prevention. BMJ 2018, 361, k2396. [Google Scholar] [CrossRef] [PubMed]
- Younas, H.; Vieira, M.; Gu, C.; Lee, R.; Shin, M.K.; Berger, S.; Loube, J.; Nelson, A.; Bevans-Fonti, S.; Zhong, Q.; et al. Caloric restriction prevents the development of airway hyperresponsiveness in mice on a high fat diet. Sci. Rep. 2019, 22, 279. [Google Scholar] [CrossRef]
- Castro Rodríguez, J.A.; Garcia Marcos, L. What is the effect of a Mediterranean diet on allergies and asthma in children? Front. Pediatr. 2017, 21, 72. [Google Scholar] [CrossRef]
- Brandhorst, S.; Longo, V.D. Dietary restrictions and nutrition in the prevention and treatment of cardiovascular disease. Circ. Res. 2019, 124, 952–965. [Google Scholar] [CrossRef]
- López-Guarnido, O.; Urquiza, N.; Saiz, M.; Lozano, D.; Rodrigo, L.; Pascual, M.; Lorente, J.A.; Alvarez-Cubero, M.J.; Rivas, A. Bioactive compounds of the Mediterranean diet and prostate cancer. Aging Male 2018, 21, 251–260. [Google Scholar] [CrossRef]
- Brigham, E.P.; Kolahdooz, F.; Hansel, N.; Breysse, P.N.; Davis, M.; Sharma, S.; Matsui, E.C.; Diette, G.; McCormack, M.C. Association between Western diet pattern and adult asthma: A focused review. Ann. Allergy Asthma Immunol. 2015, 114, 273–280. [Google Scholar] [CrossRef]
- Park, Y.; Subar, A.F.; Hollenbeck, A.; Schatzkin, A. Dietary fiber intake and mortality in the NIH-AARP diet and health study. Arch. Intern. Med. 2011, 171, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.J.; Fernandez, M.L. Dietary strategies to reduce metabolic syndrome. Rev. Endocr. Metab. Dis. 2013, 14, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Sarin, H.V.; Lee, H.; Jauhiainen, M.; Joensuu, A.; Borodulin, K.; Männistö, S.; Jin, Z.; Terwilliger, J.D.; Isola, V.; Ahtiainen, P.; et al. Substantial fat mass loss reduces low-grade inflammation and induces positive alteration in cardiometabolic factors in normal-weight individuals. Nat. Sci. Rep. 2019, 9, 3450. [Google Scholar] [CrossRef] [PubMed]
- Duwaerts, C.C.; Maher, J.J. Macronutrients and the adipose-liver axis in obesity and fatty liver. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 749–761. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Patimah, I.; Khazáai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef]
- Maggini, S.; Pierre, A.; Calder, P.C. Inmune function and micronutrient requirements change over the life course. Nutrients 2018, 10, 1531. [Google Scholar] [CrossRef]
- Jensen, M.K.; Koh-Banerjee, P.; Franz, M.; Sampson, L.; Gronbaek, M.; Rimm, E.B. Whole grains, bran and germ in relation to homocysteine and markers of glycemic control, lipids and inflammation. Am. J. Clin. Nutr. 2006, 83, 275–283. [Google Scholar] [CrossRef]
- Boulangé, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.E. Impact of the gut microbiota, obesity, and metabolic disease. Genome Med. 2016, 8, 42. [Google Scholar] [CrossRef]
- DeChristopher, L.R.; Tucker, K.L. Excess of free fructose, high-fructose corn syrup and adult asthma: The Framingham Offspring Cohort. Br. J. Nutr. 2018, 119, 1157–1167. [Google Scholar] [CrossRef]
- Botchlett, R.; Chaodong, W. Diet composition for the management of obesity and obesity-related disorders. J. Diabetes Mellit. Metab. Syndr. 2018, 3, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Wendel, S.G.; Baffi, C.; Holguin, F. Fatty acids, inflammation and asthma. J. Allergy Clin. Inmunol. 2014, 133, 1255–1264. [Google Scholar] [CrossRef]
- Sureda, A.; Bibiloni, M.M.; Julibert, A.; Bouzas, C.; Argelich, E.; Llompart, I.; Pons, A.; Tur, J.A. Adherence to the Mediterranean Diet and inflammatory markers. Nutrients 2018, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Li, Q.; Scott, H.A.; Rutting, S.; Berthon, B.S.; Gibson, P.G.; Hansbro, P.M.; Williams, E.; Horvat, J.; Simpson, J.L.; et al. Saturated fatty acids, obesity, and the nucleotide oligomerization domain-like receptor protein 3 (NLRP3) inflammasome in asthmatic patients. J. Allergy Clin. Inmunol. 2019, 143, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Shivappa, N.; Berthon, B.S.; Gibson, P.G.; Hebert, J.R. Dietary inflammatory index is related to asthma risk, lung function and systemic inflammation in asthma. Clin. Exp. Allergy 2015, 45, 177–183. [Google Scholar] [CrossRef]
- Kim, J.H.; Elwood, P.E.; Asher, M.I. Diet and asthma: Looking back, moving forward. Respir. Res. 2009, 10, 49. [Google Scholar] [CrossRef]
- Dandona, P.; Ghanim, H.; Chaudhuri, A.; Dhindsa, S.; Kim, S.S. Macronutrient intake induces oxidative and inflammatory stress: Potential relevance to atherosclerosis and insulin resistance. Exp. Mol. Med. 2010, 42, 245–253. [Google Scholar] [CrossRef]
- Sofi, F.; Macchi, C.; Abbate, R.; Gensini, G.F.; Casini, A. Mediterranean diet and health. Biofactors 2013, 39, 335–342. [Google Scholar] [CrossRef]
- Te Morenga, L.; Montez, J.M. Health effects of saturated and trans-fatty acid intake in children and adolescents: Systematic review and meta-analysis. PLoS ONE 2017, 12, e0186672. [Google Scholar] [CrossRef]
- Sievert, K.; Lawrence, M.; Naika, A.; Baker, P. Processed foods and nutrition transition in the Pacific: Regional trends, patterns and food system drivers. Nutrients 2019, 11, 1328. [Google Scholar] [CrossRef]
- WHO. Essential Nutrition Actions: Mainstreaming Nutrition throughout the Life-Course; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-151585-6. [Google Scholar]
- Li, Z.; Rava, M.; Bédard, A.; Dumas, O.; Garcia-Aymerich, J.; Leynaert, B.; Pison, C.; Le Moual, N.; Romieu, I.; Siroux, V.; et al. Cured meat intake is associated with worsening asthma symptoms. Thorax 2017, 72, 206–212. [Google Scholar] [CrossRef]
- Brathwaite, N.; Fraser, H.S.; Modeste, N.; Broome, H.; King, R. Obesity, diabetes, hypertension, and vegetarian status among Seventh-Day Adventists in Barbados: Preliminary results. Ethn. Dis. 2003, 13, 34–39. [Google Scholar]
- Andrianasolo, R.; Kesse-Guyot, E.; Moufidath, A.; Hercberg, S.; Galan, P.; Varraso, R. Association between cured meat intake and asthma symptoms. Eur. Respir. J. 2018, 52, PA1149. [Google Scholar]
- Statovci, D.; Aguilera, M.; MacSharry, J.; Melgar, S. The impact of Western diet and nutrients on the microbiota and immune response at mucosal interfaces. Front. Immunol. 2017, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, P.E.; Hawrylowicz, C.M. Vitamin D in asthma. Mechanisms of action and considerations for clinical trials. CHEST 2018, 153, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Scott, H.A.; Gibson, P.G.; Garg, M.L.; Wood, L.G. Airway inflammation is augmented by obesity and fatty acids in asthma. Eur. Respir. J. 2011, 38, 594–602. [Google Scholar] [CrossRef]
- Umetsu, D.T. Mechanisms by which obesity impacts upon asthma. Thorax 2017, 72, 174–177. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Vilariño-García, T.; Fernández-Riejos, P.; Martín-González, J.; Segura-Egea, J.J.; Sánchez-Margalet, V. Role of leptin as a link between metabolism and the immune system. Cytokine Growth Factor Rev. 2017, 35, 71–84. [Google Scholar] [CrossRef]
- Scott, H.A.; Gibson, P.G.; Garg, M.L.; Pretto, J.J.; Morgan, P.J.; Callister, R.; Wood, L.G. Relationship between body composition, inflammation and lung function in overweight and obese asthma. Respir. Res. 2012, 13, 10. [Google Scholar] [CrossRef]
- Dixon, A.A.; Poynter, M.E. Mechanisms of asthma in obesity: Pleiotropic aspects of obesity produce distinct asthma phenotypes. Am. J. Respir. Cell Mol. Biol. 2016, 54, 601–608. [Google Scholar] [CrossRef]
- Frati, F.; Salvatori, C.; Incorvaia, C.; Bellucci, A.; Di Cara, G.; Marcucci, F.; Esposito, S. The role of the microbiome in asthma: The gut-lung axis. Int. J. Mol. Sci. 2019, 20, 123. [Google Scholar] [CrossRef] [PubMed]
- Samir, P.; Malireddi, S.; Kanneganti, T.D. Food for training- Western diet and inflammatory memory. Cell Metab. 2018, 27, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, P. The role of adipokines in chronic inflammation. Immunotargets Ther. 2016, 5, 47–56. [Google Scholar] [CrossRef]
- Ambroszkiewicz, J.; Chełchowska, M.; Rowicka, G.; Klemarczyk, W.; Strucińska, M.; Gajewska, J. Anti-Inflammatory and Pro-Inflammatory Adipokine Profiles in Children on vegetarian and Omnivorous Diets. Nutrients 2018, 10, 1241. [Google Scholar] [CrossRef]
- Peake, P.W.; Kriketos, A.D.; Campbell, L.V.; Shen, Y.; Charlesworth, J.A. The metabolism of isoforms of human adiponectin: Studies in human subjects and in experimental animals. Eur. J. Endocrinol. 2005, 153, 409–417. [Google Scholar] [CrossRef]
- Salehi-Abargouei, A.; Izadi, V.; Azadbakht, L. The effect of low calorie diet on adiponectin concentration: A systematic review and meta-analysis. Horm. Metab. Res. 2015, 47, 549–555. [Google Scholar] [CrossRef]
- Zhu, N.; Pankow, J.S.; Ballantyne, C.M.; Couper, D.; Hoogeveen, R.C.; Pereira, M.; Duncan, B.B.; Schmidt, M.I. High-molecular-weight adiponectin and the risk of type 2 diabetes in the ARIC study. J. Clin. Endocrinol. Metab. 2010, 95, 5097–5104. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Hara, K.; Kamura, Y.; Honoki, H.; Fujisaka, S.; Ishiki, M.; Usui, I.; Yagi, K.; Fukushima, Y.; Takano, A.; et al. Ratio of low molecular weight serum adiponectin to the total adiponectin value is associated with type 2 diabetes through its relation to increasing insulin resistance. PLoS ONE 2018, 13, e0192609. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liang, S.; Que, S.; Zhou, L.; Zheng, S.; Mardinoglu, A. Meta-analysis of adiponectin as a biomarker for the detection of metabolic syndrome. Front. Physiol. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Daniele, A.; Scudiero, O.; Ludovica-Monaco, M.; Roviezzo, F.; D’Agostino, B.; Mazzarella, G.; Bianco, A. Adiponectin in asthma: Implications for phenotyping. Curr. Protein Pept. Sci. 2015, 16, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Denzel, M.S.; Scimia, M.C.; Zumstein, P.M.; Walsh, K.; RuizLozano, P.; Ranscht, B. T-cadherin is critical for adiponectin mediated cardioprotection in mice. J. Clin. Investig. 2010, 120, 4342–4352. [Google Scholar] [CrossRef] [PubMed]
- Parker-Duffen, J.L.; Nakamura, K.; Silver, M.; Kikuchi, R.; Tigges, U.; Yoshida, S.; Denzel, M.S.; Ranscht, B.; Walsh, K. T-cadherin is essential for adiponectin-mediated revascularization. J. Biol. Chem. 2013, 288, 24886–24897. [Google Scholar] [CrossRef] [PubMed]
- Kalisz, M.; Baranowska, B.; Wolińska-Witort, E.; Mączewski, M.; Mackiewicz, U.; Tułacz, D.; Gora, M.; Martynska, L.; Bik, W. Total and high molecular weight adiponectin levels in the rat model of post-myocardial infarction heart failure. J. Physiol. Pharmacol. 2015, 66, 673–680. [Google Scholar]
- Jaswal, S.; Saini, V.; Kaur, J.; Gupta, S.; Kaur, H.; Garg, K. Association of Adiponectin with Lung Function Impairment and Disease Severity in Chronic Obstructive Pulmonary Disease. Int. J. Appl. Basic. Med. Res. 2018, 8, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Otero, M.; Lago, R.; Gomez, R.; Lago, F.; Dieguez, C.; Gómez-Reino, J.J.; Gualillo, O. Changes in plasma levels of fat-derived hormones adiponectin, leptin, resistin and visfatin in patients with rheumatoid arthritis. Ann. Rheum. Dis. 2006, 65, 1198–1201. [Google Scholar] [CrossRef] [PubMed]
- Kirdar, S.; Serter, M.; Ceylan, E.; Sener, A.G.; Kavak, T.; Karadağ, F. Adiponectin as a biomarker of systemic inflammatory response in smoker patients with stable and exacerbation phases of chronic obstructive pulmonary disease. Scand. J. Clin. Lab. Investig. 2009, 69, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.; Shore, S.A. Adiponectin, leptin, and resistin in asthma: Basic mechanisms through population studies. J. Allergy 2013, 2013, e785835. [Google Scholar] [CrossRef]
- Ma, C.; Wang, Y.; Xue, M. Correlations of severity of asthma in children with body mass index, adiponectin and leptin. J. Clin. Lab. Anal. 2019, 33, e22915. [Google Scholar] [CrossRef]
- Wahab, A.; Maarafiya, M.M.; Ashraf Soliman, A.; Noura, B.M.; Younes, N.B.M.; Chandra, P. Serum Leptin and Adiponectin Levels in Obese and Nonobese Asthmatic School Children in relation to Asthma Control. J. Allergy 2013, 2013, e654104. [Google Scholar]
- Yuksel, H.; Sogut, A.; Yilmaz, O.; Onur, E.; Dinc, G. Role of adipokines and hormones of obesity in childhood asthma. Allergy Asthma Immunol. Res. 2012, 4, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Chen, X.; Chong, L.; Kong, L.; Wen, S.; Zhang, H.; Zhang, W.; Li, C. Adiponectin alleviates exacerbation of airway inflammation and oxidative stress in obesity-related asthma mice partly through AMPK signaling pathway. Int. Immunopharmacol. 2019, 67, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Heisler, L.K.; Lam, D.D. An appetite for life: Brain regulation of hunger and satiety. Curr. Opin. Pharmacol. 2017, 37, 100–106. [Google Scholar] [CrossRef]
- Otero, M.; Lago, R.; Gomez, R.; Dieguez, C.; Lago, F.; Gomez-Reino, J.; Gualillo, O. Towards a pro-inflammatory and immunomodulatory emerging role of leptin. Rheumatology 2006, 45, 944–950. [Google Scholar] [CrossRef]
- Frühbeck, G. Intracellular signalling pathways activated by leptin. Biochem. J. 2006, 393, 7–20. [Google Scholar] [CrossRef]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gomez-Reino, J.J.; Mera, A.; Lago, F.; Gomez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.R.; Lazar, M.A. Human resistin: Found in translation from mouse to man. Trends Endocrinol. Metab. 2011, 22, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, H.C.; Kwon, Y.W.; Lee, S.E.; Cho, Y.; Kim, J.; Lee, S.; Kim, J.Y.; Lee, J.; Yang, H.M.; et al. Adenylyl cyclase-associated protein 1 is a receptor for human resistin and mediates inflammatory actions of human monocytes. Cell Metab. 2014, 4, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Park, H.K.; Kwak, M.K.; Kim, H.J.; Ahima, R.S. Linking resistin, inflammation, and cardiometabolic diseases. Korean J. Intern. Med. 2017, 32, 239–247. [Google Scholar] [CrossRef]
- Sood, A.; Ford, E.S.; Camargo, C.A. Association between leptin and asthma in adults. Thorax 2006, 61, 300–305. [Google Scholar] [CrossRef]
- Guler, N.; Kirerleri, E.; Ones, U.; Tamay, Z.; Salmayenli, N.; Darendeliler, F. Leptin: Does it have any role in childhood asthma? J. Allergy Clin. Immunol. 2004, 114, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Bodini, A.; Tenero, L.; Sandri, M.; Maffeis, C.; Piazza, M.; Zanoni, L.; Peroni, D.; Boner, A.; Piacentini, G. Serum and exhaled breath condensate leptin levels in asthmatic and obesity children: A pilot study. J. Breath Res. 2017, 11, 046005. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Wang, J.; Zhang, Y.; Wang, Y.; Sun, L.; Han, W. Leptin positively regulates MUC5AC production and secretion induced by interleukin-13 in human bronchial epithelial cells. Biochem Biophys. Res. Commun. 2017, 493, 979–984. [Google Scholar]
- Watanabe, K.; Suzukawa, M.; Arakawa, S.; Kobayashi, K.; Igarashi, S.; Tashimo, H.; Nagai, H.; Tohma, S.; Nagase, T.; Ohta, K. Leptin enhances cytokine/chemokine production by normal lung fibroblasts by binding to leptin receptor. Allergol. Int. 2019, 68, S3–S8. [Google Scholar] [CrossRef]
- Chong, L.; Liu, L.; Zhu, L.; Li, H.; Shao, Y.; Zhang, H.; Yu, G. Expression Levels of Predominant Adipokines and Activations of STAT3, STAT6 in an Experimental Mice Model of Obese Asthma. Iran. J. Allergy Asthma Immunol. 2019, 18, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, D.; Scott, H.; MacDonald-Wicks, L.; Gibson, P.G.; Wood, L.G. Resistin is a predictor of asthma risk and resistin: Adiponectin ratio is a negative predictor of lung function in asthma. Clin. Exp. Allergy 2016, 46, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.L.; Yin, L.J.; Sharma, S.; Kierstein, S.; Wu, H.F.; Eid, G.; Haczku, A.; Corrigan, C.J.; Ying, S. Resistin-like molecule-β (RELM-β) targets airways fibroblasts to effect remodelling in asthma: From mouse to man. Clin. Exp. Allergy 2015, 45, 940–952. [Google Scholar] [CrossRef]
- Kwak, S.; Kim, Y.D.; Na, H.G.; Bae, C.H.; Song, S.Y.; Choi, Y.S. Resistin upregulates MUC5AC/B mucin gene expression in human airway epithelial cells. Biochem. Biophys. Res. Commun. 2018, 499, 655–661. [Google Scholar] [CrossRef]
- Lachowicz-Scroggins, M.E.; Yuan, S.; Kerr, S.C.; Dunican, E.M.; Yu, M.; Carrington, S.D.; Fahy, J.V. Abnormalities in MUC5AC and MUC5B protein in airway mucus in asthma. Am. J. Respir. Crit. Care Med. 2016, 194, 1296–1299. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frontela-Saseta, C.; González-Bermúdez, C.A.; García-Marcos, L. Diet: A Specific Part of the Western Lifestyle Pack in the Asthma Epidemic. J. Clin. Med. 2020, 9, 2063. https://doi.org/10.3390/jcm9072063
Frontela-Saseta C, González-Bermúdez CA, García-Marcos L. Diet: A Specific Part of the Western Lifestyle Pack in the Asthma Epidemic. Journal of Clinical Medicine. 2020; 9(7):2063. https://doi.org/10.3390/jcm9072063
Chicago/Turabian StyleFrontela-Saseta, Carmen, Carlos A. González-Bermúdez, and Luis García-Marcos. 2020. "Diet: A Specific Part of the Western Lifestyle Pack in the Asthma Epidemic" Journal of Clinical Medicine 9, no. 7: 2063. https://doi.org/10.3390/jcm9072063
APA StyleFrontela-Saseta, C., González-Bermúdez, C. A., & García-Marcos, L. (2020). Diet: A Specific Part of the Western Lifestyle Pack in the Asthma Epidemic. Journal of Clinical Medicine, 9(7), 2063. https://doi.org/10.3390/jcm9072063