The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control

 ,
, 

 and
and 
Abstract
1. Introduction
2. History of Coronavirus Outbreaks
3. Coronavirus Taxonomy
4. SARS-CoV-2 Genome Organization
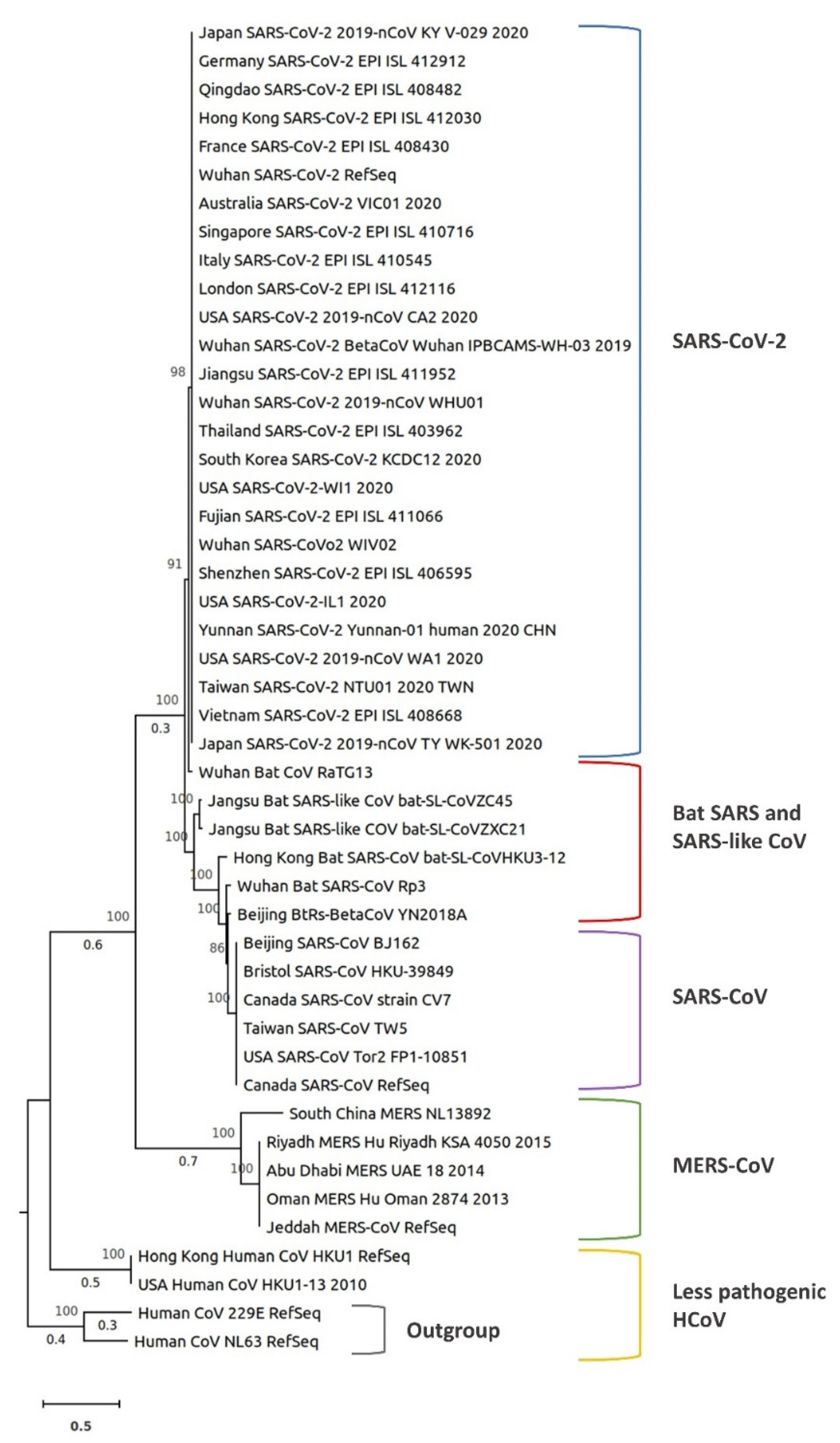
5. Comparative Phylogenetic Analysis of SARS-CoV-2
6. SARS-CoV-2 Mutations and Their Effects
7. Genetic Diversity of SARS-CoV-2
8. Epidemiology of COVID-19
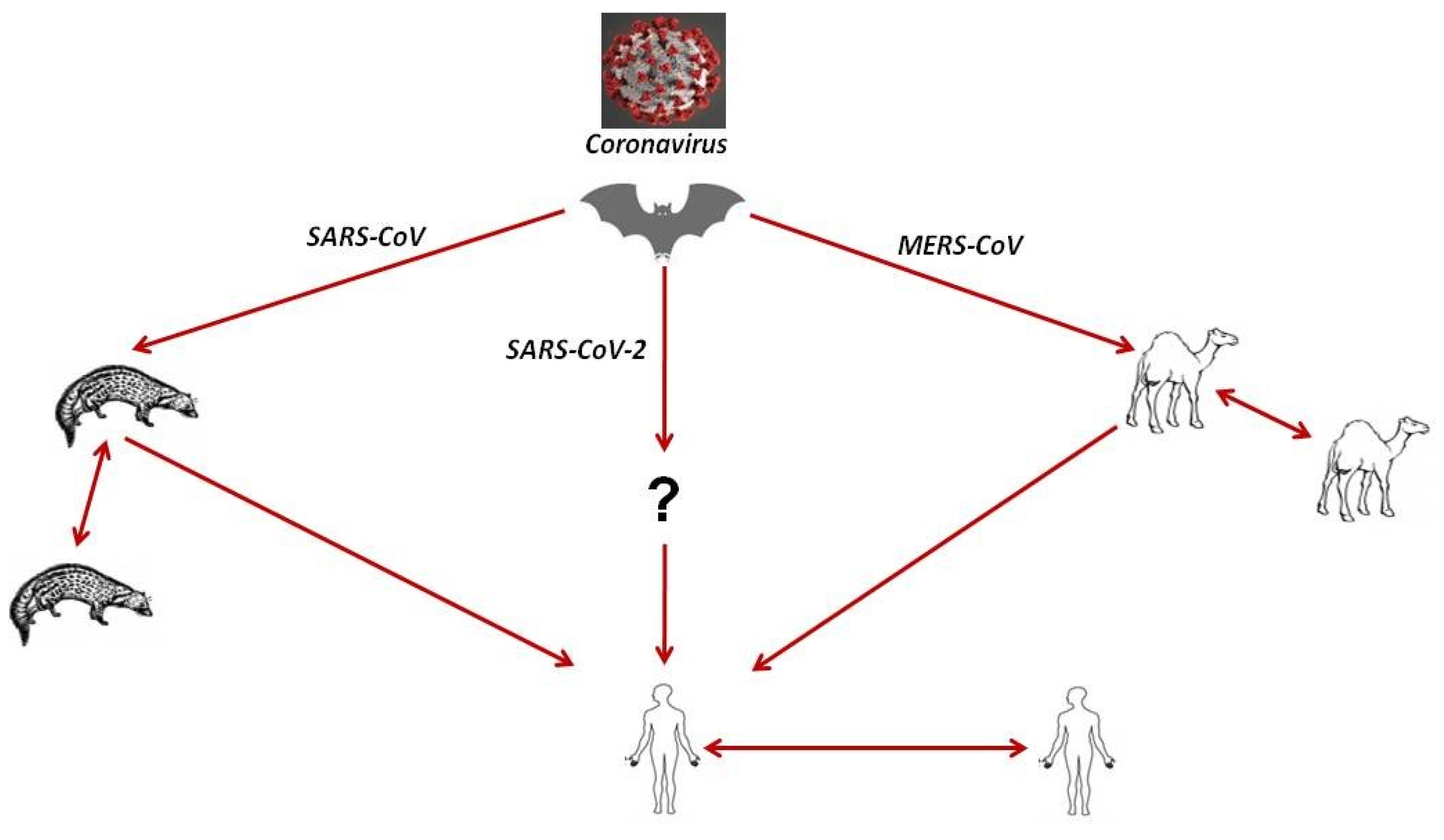
8.1. Source of Infection and Evolution of SARS-CoV-2
8.2. Mode of SARS-CoV-2 Transmission to Humans: Transmission Dynamics and Virus Persistence
8.3. Risk Factors for SARS-CoV-2 Infection and Their Assessment
8.4. Clinical Characteristics and Susceptibility of SARS-CoV-2 Infection in Humans
9. Diagnosis of SARS-CoV-2 Infection
9.1. Epidemiological History
9.2. Laboratory Diagnosis
9.3. Virus Detection
9.4. Serological and Molecular Diagnosis
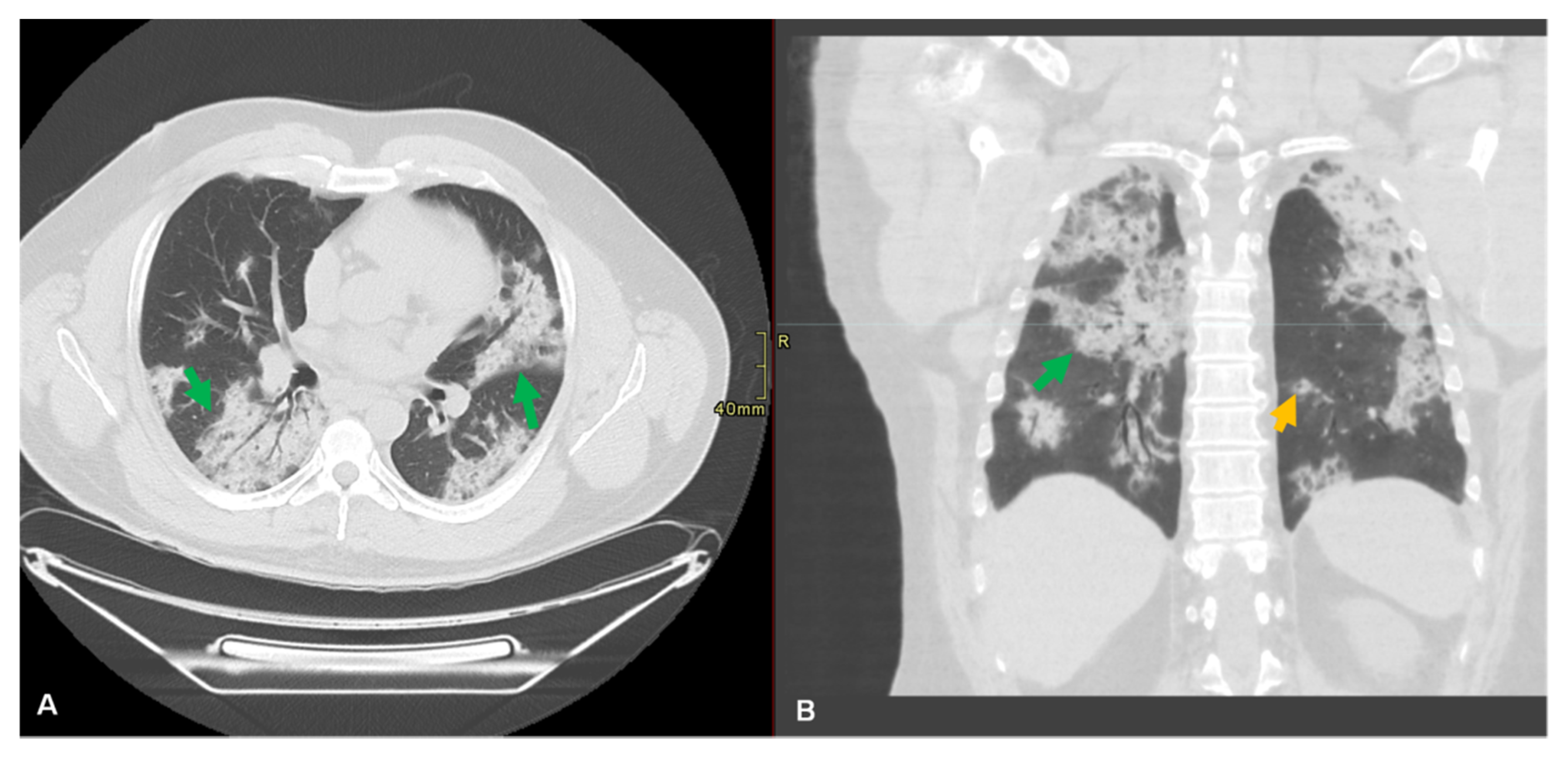
9.5. Radiological Diagnosis
10. Control and Treatment of COVID-19 Infection
11. Vaccination
12. Preventive Measures to Control the SARS-CoV-2 Spread
13. Challenges to Control SARS-CoV-2 and the Role of the One-Health Approach in Disease Control
14. Conclusions and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Decaro, N.; Mari, V.; Elia, G.; Addie, D.D.; Camero, M.; Lucente, M.S.; Martella, V.; Buonavoglia, C. Recombinant canine coronaviruses in dogs, Europe. Emerg. Infect. Dis. 2010, 16, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Peng, Y.; Huang, B.; Ding, X.; Wang, X.; Niu, P.; Meng, J.; Zhu, Z.; Zhang, Z.; Wang, J. Genome composition and divergence of the novel coronavirus (2019-nCoV) originating in China. Cell Host Microbe 2020, 27, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Zheng, B.; He, Y.; Liu, X.; Zhuang, Z.; Cheung, C.; Luo, S.; Li, P.; Zhang, L.; Guan, Y. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Meyer, B.; Müller, M. Supplement to: Transmission of MERS-coronavirus in household contacts. N. Engl. J. Med. 2014, 371, 1–10. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef]
- WHO. Summary of Probable SARS Cases with Onset of Illness from 1 November 2002 to 31 July 2003. Available online: https://www.who.int/csr/sars/country/table2004_04_21/en/ (accessed on 10 March 2020).
- WHO. MERS Situation Update, January 2020. Available online: http://www.emro.who.int/health-topics/mers-cov/mers-outbreaks.html (accessed on 11 March 2020).
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Chan, J.F.; Yuan, S.; Kok, K.H.; To, K.K.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.; Poon, R.W.; et al. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: A study of a family cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef]
- Paraskevis, D.; Kostaki, E.G.; Magiorkinis, G.; Panayiotakopoulos, G.; Sourvinos, G.; Tsiodras, S. Full-genome evolutionary analysis of the novel corona virus (2019-nCoV) rejects the hypothesis of emergence as a result of a recent recombination event. Infect. Genet. Evol. 2020, 79, 104212. [Google Scholar] [CrossRef]
- Rothe, C.; Schunk, M.; Sothmann, P.; Bretzel, G.; Froeschl, G.; Wallrauch, C.; Zimmer, T.; Thiel, V.; Janke, C.; Guggemos, W. Transmission of 2019-nCoV infection from an asymptomatic contact in Germany. N. Engl. J. Med. 2020, 382, 970–971. [Google Scholar] [CrossRef]
- Chan, J.F.; Kok, K.H.; Zhu, Z.; Chu, H.; To, K.K.; Yuan, S.; Yuen, K.Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; You, Z.; Wang, Q.; Zhou, Z.-J.; Qiu, Y.; Luo, R.; Ge, X.-Y. The epidemic of 2019-novel-coronavirus (2019-nCoV) pneumonia and insights for emerging infectious diseases in the future. Microb. Infect. 2020, 22, 80–85. [Google Scholar] [CrossRef] [PubMed]
- WHO. Clinical Management of Severe Acute Respiratory Infection When Novel Coronavirus (nCoV) Infection Is Suspected. Available online: https://www.who.int/publications-detail/clinical-management-of-severe-acute-respiratory-infection-when-novel-coronavirus-(ncov)-infection-is-suspected (accessed on 17 March 2020).
- Cowling, B.J.; Leung, G.M. Epidemiological research priorities for public health control of the ongoing global novel coronavirus (2019-nCoV) outbreak. Euro Surveill. 2020, 25, 2000110. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular evolution of human coronavirus genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef]
- Beaudette, F.R.; Hudson, C.B. Cultivation of the virus of infectious bronchitis. J. Am. Vet. Med. A. 1937, 90, 51–56. [Google Scholar]
- Cheever, F.S.; Daniels, J.B.; Pappenheimer, A.M.; Bailey, O.T. A murine virus (JHM) causing disseminated encephalomyelitis with extensive destruction of myelin: I. Isolation and biological properties of the virus. J. Exp. Med. 1949, 90, 181–194. [Google Scholar] [CrossRef]
- Kahn, J.S.; McIntosh, K. History and recent advances in coronavirus discovery. Pediatr. Infect. Dis. J. 2005, 24, S223–S227. [Google Scholar] [CrossRef]
- Tyrrell, D.; Bynoe, M. Cultivation of viruses from a high proportion of patients with colds. Lancet 1966, 1, 76–77. [Google Scholar] [CrossRef]
- Hamre, D.; Procknow, J.; Medicine. A new virus isolated from the human respiratory tract. Proc. Soc. Exp. Biol. Med. 1966, 121, 190–193. [Google Scholar] [CrossRef]
- Tyrrell, D.; Almeida, J.; Cunningham, C.; Dowdle, W.; Hofstad, M.; McIntosh, K.; Tajima, M.; Zakstelskaya, L.Y.; Easterday, B.; Kapikian, A. Coronaviridae. Intervirology 1975, 5, 76–82. [Google Scholar] [CrossRef]
- McIntosh, K.; Becker, W.B.; Chanock, R.M. Growth in suckling-mouse brain of “IBV-like” viruses from patients with upper respiratory tract disease. Proc. Natl. Acad. Sci. USA 1967, 58, 2268–2273. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M.; Perlman, S.; Anderson, L. Coronaviridae. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 1, pp. 1305–1318. [Google Scholar]
- Hu, B.; Zeng, L.-P.; Yang, X.-L.; Ge, X.-Y.; Zhang, W.; Li, B.; Xie, J.-Z.; Shen, X.-R.; Zhang, Y.-Z.; Wang, N. Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS Pathog. 2017, 13, e1006698. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.M.; Van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Hui, K.P.; Perera, R.A.; Miguel, E.; Niemeyer, D.; Zhao, J.; Channappanavar, R.; Dudas, G.; Oladipo, J.O.; Traoré, A. MERS coronaviruses from camels in Africa exhibit region-dependent genetic diversity. Proc. Natl. Acad. Sci. USA 2018, 115, 3144–3149. [Google Scholar] [CrossRef]
- Lau, S.K.; Li, K.S.; Tsang, A.K.; Lam, C.S.; Ahmed, S.; Chen, H.; Chan, K.-H.; Woo, P.C.; Yuen, K.-Y. Genetic characterization of Betacoronavirus lineage C viruses in bats reveals marked sequence divergence in the spike protein of pipistrellus bat coronavirus HKU5 in Japanese pipistrelle: Implications for the origin of the novel Middle East respiratory syndrome coronavirus. J. Virol. 2013, 87, 8638–8650. [Google Scholar]
- Dudas, G.; Rambaut, A. MERS-CoV recombination: Implications about the reservoir and potential for adaptation. Virus Evol. 2016, 2. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, D.; Shi, W.; Lu, R.; Wang, W.; Zhao, Y.; Deng, Y.; Zhou, W.; Ren, H.; Wu, J. Origin and possible genetic recombination of the Middle East respiratory syndrome coronavirus from the first imported case in China: Phylogenetics and coalescence analysis. mBio 2015, 6, e01280-15. [Google Scholar] [CrossRef]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Luo, C.-M.; Wang, N.; Yang, X.-L.; Liu, H.-Z.; Zhang, W.; Li, B.; Hu, B.; Peng, C.; Geng, Q.-B.; Zhu, G.-J. Discovery of novel bat coronaviruses in south China that use the same receptor as Middle East respiratory syndrome coronavirus. J. Virol. 2018, 92, e00116-18. [Google Scholar] [CrossRef]
- Weiss, S.R.; Navas-Martin, S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef]
- Lai, M.M.; Cavanagh, D. The molecular biology of coronaviruses. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 1997; Volume 48, pp. 1–100. [Google Scholar]
- Tyrrell, D.; Bynoe, M. Cultivation of a novel type of common-cold virus in organ cultures. Br. Med. J. 1965, 1, 1467–1470. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Günther, S.; Preiser, W.; Van Der Werf, S.; Brodt, H.-R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, K.; Shi, Z.-L.; Zhou, P. Bat Coronaviruses in China. Viruses 2019, 11, 210. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, H.; Zhao, J.; Zhong, Q.; Jin, J.-H.; Zhang, G.-Z. Evolution of infectious bronchitis virus in China over the past two decades. J. Gen. Virol. 2016, 97, 1566–1574. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Halpin, R.; Wang, S.; Ghedin, E.; Spiro, D.J.; Saif, L.J. Molecular characterization of a new species in the genus Alphacoronavirus associated with mink epizootic catarrhal gastroenteritis. J. Gen. Virol. 2011, 92, 1369–1379. [Google Scholar] [CrossRef]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 2018, 556, 255–258. [Google Scholar] [CrossRef]
- GISAID. Global Initiative on Sharing All Influenza Data. Available online: https://www.gisaid.org/ (accessed on 14 April 2020).
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef]
- Ceraolo, C.; Giorgi, F.M. Genomic variance of the 2019-nCoV coronavirus. J. Med. Virol. 2020, 92, 522–528. [Google Scholar] [CrossRef]
- Angeletti, S.; Benvenuto, D.; Bianchi, M.; Giovanetti, M.; Pascarella, S.; Ciccozzi, M. COVID-2019: The role of the nsp2 and nsp3 in its pathogenesis. J. Med. Virol. 2020, in press. [Google Scholar] [CrossRef]
- Krichel, B.; Falke, S.; Hilgenfeld, R.; Redecke, L.; Uetrecht, C. Processing of the SARS-CoV pp1a/ab nsp7-10 region. Biochem. J. 2019, 477, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Elena, S.F.; Sanjuán, R. Adaptive value of high mutation rates of RNA viruses: Separating causes from consequences. J. Virol. 2005, 79, 11555–11558. [Google Scholar] [CrossRef] [PubMed]
- Ferron, F.; Subissi, L.; De Morais, A.T.S.; Le, N.T.T.; Sevajol, M.; Gluais, L.; Decroly, E.; Vonrhein, C.; Bricogne, G.; Canard, B. Structural and molecular basis of mismatch correction and ribavirin excision from coronavirus RNA. Proc. Natl. Acad. Sci. USA 2018, 115, E162–E171. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Lee, P.; Tsang, A.K.; Yip, C.C.; Tse, H.; Lee, R.A.; So, L.-Y.; Lau, Y.-L.; Chan, K.-H.; Woo, P.C. Molecular epidemiology of human coronavirus OC43 reveals evolution of different genotypes over time and recent emergence of a novel genotype due to natural recombination. J. Virol. 2011, 85, 11325–11337. [Google Scholar] [CrossRef]
- Makino, S.; Keck, J.G.; Stohlman, S.A.; Lai, M. High-frequency RNA recombination of murine coronaviruses. J. Virol. 1986, 57, 729–737. [Google Scholar] [CrossRef]
- Zhang, X.; Yap, Y.; Danchin, A. Testing the hypothesis of a recombinant origin of the SARS-associated coronavirus. Arch. Virol. 2005, 150, 1–20. [Google Scholar] [CrossRef]
- Zhang, Z.; Shen, L.; Gu, X. Evolutionary dynamics of MERS-CoV: Potential recombination, positive selection and transmission. Sci. Rep. 2016, 6, 25049. [Google Scholar] [CrossRef]
- Lassmann, T. Kalign 3: Multiple sequence alignment of large datasets. Bioinformatics 2019, 36, 1928–1929. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor recognition by novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS. J. Virol. 2020, 94, e00127-20. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-J.; Walls, A.C.; Wang, Z.; Sauer, M.M.; Li, W.; Tortorici, M.A.; Bosch, B.-J.; DiMaio, F.; Veesler, D. Structures of MERS-CoV spike glycoprotein in complex with sialoside attachment receptors. Nat. Struct. Mol. Biol. 2019, 26, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, in press. [Google Scholar] [CrossRef]
- Andersen, K.; Rambaut, A.; Lipkin, W.; Holmes, E.C.; Garry, R. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef]
- Yount, B.; Roberts, R.S.; Sims, A.C.; Deming, D.; Frieman, M.B.; Sparks, J.; Denison, M.R.; Davis, N.; Baric, R.S. Severe acute respiratory syndrome coronavirus group-specific open reading frames encode nonessential functions for replication in cell cultures and mice. J. Virol. 2005, 79, 14909–14922. [Google Scholar] [CrossRef]
- Khan, S.; Fielding, B.C.; Tan, T.H.; Chou, C.-F.; Shen, S.; Lim, S.G.; Hong, W.; Tan, Y.-J. Over-expression of severe acute respiratory syndrome coronavirus 3b protein induces both apoptosis and necrosis in Vero E6 cells. Virus Res. 2006, 122, 20–27. [Google Scholar] [CrossRef]
- Kopecky-Bromberg, S.A.; Martínez-Sobrido, L.; Frieman, M.; Baric, R.A.; Palese, P. Severe acute respiratory syndrome coronavirus open reading frame (ORF) 3b, ORF 6, and nucleocapsid proteins function as interferon antagonists. J. Virol. 2007, 81, 548–557. [Google Scholar] [CrossRef]
- Phan, T. Genetic diversity and evolution of SARS-CoV-2. Infect. Genet. Evol. 2020, 81, 104260. [Google Scholar] [CrossRef]
- CDC. Coronavirus Disease 2019 (COVID-19) Situation Summary. Available online: https://www.cdc.gov/coronavirus/2019-nCoV/summary.html#risk-assessment (accessed on 3 April 2020).
- WHO. Coronavirus Disease (COVID-2019) Situation Reports-48. Available online: https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200308-sitrep-48-covid-19.pdf?sfvrsn=16f7ccef_4 (accessed on 10 March 2020).
- WHO. Rolling Updates on Coronavirus Disease (COVID-19). Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/events-as-they-happen (accessed on 14 April 2020).
- WHO. Coronavirus Disease (COVID-2019) Situation Reports. Available online: https://www.who.int/docs/default-source/coronaviruse/situation-%E2%80%8Ereports/20200304-sitrep-44-covid-19.pdf?sfvrsn=783b4c9d_2%E2%80%8E (accessed on 8 March 2020).
- WHO. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19-11 March 2020. Available online: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 12 March 2020).
- Helmy, Y.A.; El-Adawy, H.; Abdelwhab, E.M. A comprehensive review of common bacterial, parasitic and viral zoonoses at the human-animal interface in Egypt. Pathogens 2017, 6, 33. [Google Scholar] [CrossRef]
- Ji, W.; Wang, W.; Zhao, X.; Zai, J.; Li, X. Cross-species transmission of the newly identified coronavirus 2019-nCoV. J. Med. Virol. 2020, 92, 433–440. [Google Scholar] [CrossRef] [PubMed]
- ECDC. Outbreak of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Increased Transmission Beyond China—Fourth Update. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/SARS-CoV-2-risk-assessment-14-feb-2020.pdf (accessed on 26 February 2020).
- Liu, S.L.; Saif, L.J.; Weiss, S.R.; Su, L. No credible evidence supporting claims of the laboratory engineering of SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 505–507. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS–coronavirus 2. Science 2020. [Google Scholar] [CrossRef] [PubMed]
- CDC. What You Need to Know about Coronavirus Disease 2019 (COVID-19). Available online: https://www.cdc.gov/coronavirus/2019-ncov/downloads/2019-ncov-factsheet.pdf (accessed on 6 April 2020).
- Lai, C.C.; Shih, T.P.; Ko, W.C.; Tang, H.J.; Hsueh, P.R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and corona virus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef]
- Holshue, M.L.; DeBolt, C.; Lindquist, S.; Lofy, K.H.; Wiesman, J.; Bruce, H.; Spitters, C.; Ericson, K.; Wilkerson, S.; Tural, A.; et al. First case of 2019 novel coronavirus in the United States. N. Engl. J. Med. 2020, 382, 929–936. [Google Scholar] [CrossRef]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and its inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef]
- Chen, Z.M.; Fu, J.F.; Shu, Q.; Chen, Y.H.; Hua, C.Z.; Li, F.B.; Lin, R.; Tang, L.F.; Wang, T.L.; Wang, W.; et al. Diagnosis and treatment recommendations for pediatric respiratory infection caused by the 2019 novel coronavirus. World J. Pediatr. 2020. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- WHO. Coronavirus Disease (COVID-19) Technical Guidance: Laboratory Testing for 2019-nCoV in Humans. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/technical-guidance/laboratory-guidance (accessed on 12 March 2020).
- Bai, Y.; Yao, L.; Wei, T.; Tian, F.; Jin, D.Y.; Chen, L.; Wang, M. Presumed asymptomatic carrier transmission of COVID-19. JAMA 2020, 323, 1406–1407. [Google Scholar] [CrossRef]
- WHO. Coronavirus. Available online: https://www.who.int/health-topics/coronavirus#tab=tab_3 (accessed on 14 April 2020).
- NCPERE. The epidemiological characteristics of an outbreak of 2019 novel coronavirus diseases (COVID-19) in China. Zhonghua Liu Xing Bing Xue Za Zhi 2020, 41, 145–151. [Google Scholar] [CrossRef]
- Zhang, W.; Du, R.-H.; Li, B.; Zheng, X.-S.; Yang, X.-L.; Hu, B.; Wang, Y.-Y.; Xiao, G.-F.; Yan, B.; Shi, Z.-L. Molecular and serological investigation of 2019-nCoV infected patients: Implication of multiple shedding routes. Emerg. Microbes Infect. 2020, 9, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S. Another decade, another coronavirus. N. Engl. J. Med. 2020, 382, 760–762. [Google Scholar] [CrossRef] [PubMed]
- Shirato, K.; Nao, N.; Katano, H.; Takayama, I.; Saito, S.; Kato, F.; Katoh, H.; Sakata, M.; Nakatsu, Y.; Mori, Y.; et al. Development of genetic diagnostic methods for novel coronavirus 2019 (nCoV-2019) in Japan. Jpn. J. Infect. Dis. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brunink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro. Surveill. 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Outbreak of novel coronavirus (COVID-19): What is the role of radiologists? Eur. Radiol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Ye, T.; Sun, P.; Gui, S.; Liang, B.; Li, L.; Zheng, D.; Wang, J.; Hesketh, R.L.; Yang, L.; et al. Time course of lung changes on chest CT during recovery from 2019 novel coronavirus (COVID-19) pneumonia. Radiology 2020, in press. [Google Scholar] [CrossRef]
- Ryu, S.; Chun, B. An interim review of the epidemiological characteristics of 2019 novel coronavirus. Epidemiol. Health 2020, 42, e2020006. [Google Scholar] [CrossRef]
- Chung, M.; Bernheim, A.; Mei, X.; Zhang, N.; Huang, M.; Zeng, X.; Cui, J.; Xu, W.; Yang, Y.; Fayad, Z.A. CT imaging features of 2019 novel coronavirus (2019-nCoV). Radiology 2020, 295, 202–207. [Google Scholar] [CrossRef]
- Song, F.; Shi, N.; Shan, F.; Zhang, Z.; Shen, J.; Lu, H.; Ling, Y.; Jiang, Y.; Shi, Y. Emerging 2019 novel coronavirus (2019-nCoV) pneumonia. Radiology 2020, 295, 210–217. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Xu, J.; Cao, B. Potential antiviral therapeutics for 2019 novel coronavirus. Zhonghua Jie He He Hu Xi Za Zhi 2020, 43, E002. [Google Scholar]
- Zylka-Menhorn, V.; Grunert, G. Coronavirus 2019-nCoV: Der Steckbrief des Virus ist im Fluss. Deutsches Ärzteblatt Jg. 117, Heft 6. Dtsch. Arztebl. 2020, 117, A-250/B-219/C-215. [Google Scholar]
- Han, W.; Quan, B.; Guo, Y.; Zhang, J.; Lu, Y.; Feng, G.; Wu, Q.; Fang, F.; Cheng, L.; Jiao, N.; et al. The course of clinical diagnosis and treatment of a case infected with coronavirus disease 2019. J. Med. Virol. 2020, 92, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Jeon, S.; Shin, H.Y.; Kim, M.J.; Seong, Y.M.; Lee, W.J.; Choe, K.W.; Kang, Y.M.; Lee, B.; Park, S.J. Case of the index patient who caused tertiary transmission of COVID-19 infection in Korea: The application of lopinavir/ritonavir for the treatment of COVID-19 infected pneumonia monitored by quantitative RT-PCR. J. Korean Med. Sci. 2020, 35, e79. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xu, Y.; Yang, Q.; Chen, J.; Zhang, T.; Li, Z.; Guo, C.; Chen, H.; Wu, H.; Li, N. Efficacy and biological safety of lopinavir/ritonavir based anti-retroviral therapy in HIV-1-infected patients: A meta-analysis of randomized controlled trials. Sci. Rep. 2015, 5, 8528. [Google Scholar] [CrossRef]
- Al-Tawfiq, J.A.; Momattin, H.; Dib, J.; Memish, Z.A. Ribavirin and interferon therapy in patients infected with the Middle East respiratory syndrome coronavirus: An observational study. Int. J. Infect. Dis. 2014, 20, 42–46. [Google Scholar] [CrossRef]
- Arabi, Y.M.; Shalhoub, S.; Mandourah, Y.; Al-Hameed, F.; Al-Omari, A.; Al Qasim, E.; Jose, J.; Alraddadi, B.; Almotairi, A.; Al Khatib, K.; et al. Ribavirin and interferon therapy for critically ill patients with Middle East respiratory syndrome: A multicenter observational study. Clin. Infect. Dis. 2019, in press. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, Y.; Zhang, F.; Wang, Q.; Li, T.; Liu, Z.; Wang, J.; Qin, Y.; Zhang, X.; Yan, X.; et al. The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The experience of clinical immunologists from China. Clin. Immunol. 2020, 214, 108393. [Google Scholar] [CrossRef]
- Bradley, K.C.; Finsterbusch, K.; Schnepf, D.; Crotta, S.; Llorian, M.; Davidson, S.; Fuchs, S.Y.; Staeheli, P.; Wack, A. Microbiota-driven tonic interferon signals in lung stromal cells protect from influenza virus infection. Cell Rep. 2019, 28, 245–256.e244. [Google Scholar] [CrossRef]
- Dai, M.; Liu, Y.; Chen, W.; Buch, H.; Shan, Y.; Chang, L.; Bai, Y.; Shen, C.; Zhang, X.; Huo, Y. Rescue fecal microbiota transplantation for antibiotic-associated diarrhea in critically ill patients. Crit. Care 2019, 23, 324. [Google Scholar] [CrossRef]
- Ding, X.; Li, Q.; Li, P.; Zhang, T.; Cui, B.; Ji, G.; Lu, X.; Zhang, F. Long-Term safety and efficacy of fecal microbiota transplant in active ulcerative colitis. Drug Saf. 2019, 42, 869–880. [Google Scholar] [CrossRef]
- Domínguez-Díaz, C.; García-Orozco, A.; Riera-Leal, A.; Padilla-Arellano, J.R.; Fafutis-Morris, M. Microbiota and its role on viral evasion: Is it with us or against us? Front. Cell. Infect. Microbiol. 2019, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Tang, Q.-L.; Shang, Y.-X.; Liang, S.-B.; Yang, M.; Robinson, N.; Liu, J.-P. Can Chinese medicine be used for prevention of corona virus disease 2019 (COVID-19)? A review of historical classics, research evidence and current prevention programs. Chin. J. Integr. Med. 2020, 26, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.-R.; Lin, C.-S.; Lai, H.-C.; Lin, Y.-P.; Wang, C.-Y.; Tsai, Y.-C.; Wu, K.-C.; Huang, S.-H.; Lin, C.-W. Antiviral activity of Sambucus FormosanaNakai ethanol extract and related phenolic acid constituents against human coronavirus NL63. Virus Res. 2019, 273, 197767. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.M.; Cheng, V.C.; Hung, I.F.; Wong, M.M.; Chan, K.H.; Chan, K.S.; Kao, R.Y.; Poon, L.L.; Wong, C.L.; Guan, Y.; et al. Role of lopinavir/ritonavir in the treatment of SARS: Initial virological and clinical findings. Thorax 2004, 59, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ding, Y.; Yang, C.; Li, R.; Du, Q.; Hao, Y.; Li, Z.; Jiang, H.; Zhao, J.; Chen, Q.; et al. Inhibition of the infectivity and inflammatory response of influenza virus by Arbidol hydrochloride in vitro and in vivo (mice and ferret). Biomed. Pharmacother. 2017, 91, 393–401. [Google Scholar] [CrossRef]
- Furuta, Y.; Gowen, B.B.; Takahashi, K.; Shiraki, K.; Smee, D.F.; Barnard, D.L. Favipiravir (T-705), a novel viral RNA polymerase inhibitor. Antiviral Res. 2013, 100, 446–454. [Google Scholar] [CrossRef]
- Furuta, Y.; Komeno, T.; Nakamura, T. Favipiravir (T-705), a broad spectrum inhibitor of viral RNA polymerase. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 449–463. [Google Scholar] [CrossRef]
- Janowski, A.B.; Dudley, H.; Wang, D. Antiviral activity of ribavirin and favipiravir against human astroviruses. J. Clin. Virol. 2020, 123, 104247. [Google Scholar] [CrossRef]
- Gao, J.; Li, J.; Shao, X.; Jin, Y.; Lü, X.W.; Ge, J.F.; Huang, Y.; Zhang, L.; Chen, L. Antiinflammatory and immunoregulatory effects of total glucosides of Yupingfeng powder. Chin. Med. J. 2009, 122, 1636–1641. [Google Scholar]
- Sheahan, T.P.; Sims, A.C.; Leist, S.R.; Schäfer, A.; Won, J.; Brown, A.J.; Montgomery, S.A.; Hogg, A.; Babusis, D.; Clarke, M.O.; et al. Comparative therapeutic efficacy of remdesivir and combination lopinavir, ritonavir, and interferon beta against MERS-CoV. Nat. Commun. 2020, 11, 222. [Google Scholar] [CrossRef]
- Lathouwers, E.; Wong, E.Y.; Luo, D.; Seyedkazemi, S.; De Meyer, S.; Brown, K. HIV-1 resistance rarely observed in patients using darunavir once-daily regimens across clinical studies. HIV Clin. Trials 2017, 18, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Mifsud, E.J.; Tilmanis, D.; Oh, D.Y.; Ming-Kay Tai, C.; Rossignol, J.F.; Hurt, A.C. Prophylaxis of ferrets with nitazoxanide and oseltamivir combinations is more effective at reducing the impact of influenza a virus infection compared to oseltamivir monotherapy. Antiviral Res. 2020, 176, 104751. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Allen Chan, K.C.; Hui, D.S.; Ng, E.K.; Wu, A.; Chiu, R.W.; Wong, V.W.; Chan, P.K.; Wong, K.T.; Wong, E.; et al. Effects of early corticosteroid treatment on plasma SARS-associated Coronavirus RNA concentrations in adult patients. J. Clin. Virol. 2004, 31, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, Z.; Zhao, F.; Yang, Y.; Li, J.; Yuan, J.; Wang, F.; Li, D.; Yang, M.; Xing, L.; et al. Treatment of 5 critically ill patients with COVID-19 with convalescent plasma. JAMA 2020, in press. [Google Scholar] [CrossRef]
- Jawhara, S. Could Intravenous Immunoglobulin Collected from Recovered Coronavirus Patients Protect against COVID-19 and Strengthen the Immune System of New Patients? Int. J. Mol. Sci. 2020, 21, 2272. [Google Scholar] [CrossRef]
- Atluri, S.; Manchikanti, L.; Hirsch, J. Expanded umbilical cord mesenchymal stem cells (UC-MSCs) as a therapeutic strategy in managing critically ill COVID-19 patients: The case for compassionate use. Pain Phys. 2020, 23, E71–E83. [Google Scholar]
- Clay, L.; Druce, J.; Catton, M.; Jans, D.; Wagstaff, K. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antiviral Res. 2020, in press. [Google Scholar] [CrossRef]
- Chen, W.-H.; Strych, U.; Hotez, P.; Bottazzi, M.E. The SARS-CoV-2 vaccine pipeline: An overview. Curr. Trop. Med. Rep. 2020, in press. [Google Scholar] [CrossRef]
- Phan, L.T.; Nguyen, T.V.; Luong, Q.C.; Nguyen, T.V.; Nguyen, H.T.; Le, H.Q.; Nguyen, T.T.; Cao, T.M.; Pham, Q.D. Importation and human-to-human transmission of a novel coronavirus in Vietnam. N. Engl. J. Med. 2020, 382, 872–874. [Google Scholar] [CrossRef]
- Neiderud, C.-J. How urbanization affects the epidemiology of emerging infectious diseases. Infect. Ecol. Epidemiol. 2015, 5, 27060. [Google Scholar] [CrossRef]
- WHO. Infection Prevention and Control during Health Care When Novel Coronavirus (nCoV) Infection Is Suspected Interim Guidance. Available online: https://www.who.int/publications-detail/infection-prevention-and-control-during-health-care-when-novel-coronavirus-(ncov)-infection-is-suspected-20200125 (accessed on 17 March 2020).
- Kock, R.A.; Karesh, W.B.; Veas, F.; Velavan, T.P.; Simons, D.; Mboera, L.E.; Dar, O.; Arruda, L.B.; Zumla, A. 2019-nCoV in context: Lessons learned? Lancet Planet. Health 2020, 4, E87–E88. [Google Scholar] [CrossRef]
- Daszak, P.; Olival, K.J.; Li, H. A strategy to prevent future pandemics similar to the 2019-nCoV outbreak. Biosaf. Health 2020, 2, 6–8. [Google Scholar] [CrossRef]
- Fawzy, M.; Helmy, Y.A. The one health approach is necessary for the control of Rift Valley fever infections in Egypt: A comprehensive review. Viruses 2019, 11, 139. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Subfamily | Genus | Subgenus | Species | Abbreviations | Accession Numbers | Reservoir Host | Zoonotic [39] |
|---|---|---|---|---|---|---|---|---|
| Coronaviridae | Letovirinae | Alphaletovirus | Milecovirus | Microhyla letovirus 1 | No | |||
| Orthocoronavirinae | Alphacoronavirus | Colacovirus | Bat coronavirus CDPHE15 | BtCoV-CDPHE15 | NC_022103.1 | Bats | No | |
| Decacovirus | Bat coronavirus HKU10 | BtCoV-HKU10 | NC_018871.1 | Bats | No | |||
| Rhinolophus ferrumeguinum alphacoronavirus HuB-2013 | BtRfCoV-HuB13 | KJ473807.1 | Bats | No | ||||
| Duvinacovirus | Human coronavirus 229E | HCoV-229E | NC_002645.1 | Human | No | |||
| Luchacovirus | Lucheng Rn rat coronavirus | LRNV | NC_032730.1 | Rats | No | |||
| Minacovirus | Ferret coronavirus | FRCoV | NC_030292.1 | Ferrets | No | |||
| Mink coronavirus 1 | MCoV | NC_023760.1 | Minks | No | ||||
| Minunacovirus | Miniopetrus batcoronavirus 1 | BtMiCoV-1 | EU420138.1 | Bats | No | |||
| Miniopetrus batcoronavirus HKU8 | BtMiCoV-HKU8 | EU420139.1 | Bats | No | ||||
| Mytacovirus | Myotis ricketti alphacoronavirus Sax-2011 | BtMy-Sax11 | NC_028811.1 | Bats | No | |||
| Nyctacovirus | Nyctalus velutinus alphacoronavirus SC-2013 | BtNy-Sc13 | NC_028833.1 | Bats | No | |||
| Pedacovirus | Porcine epidemic diarrhea virus | PEDV | NC_003436.1 | Pigs | No | |||
| Scotophilus bat coronavirus 512 | BtScCoV-512 | NC_009657.1 | Bats | |||||
| Rhinacovirus | Rhinolophus bat coronavirus HKU2 | BtRhCoV-HKU2 (SADS) | NC_009988.1 | Bats and pigs | Yes | |||
| Setracovirus | Human coronavirus NL63 | HCoV-NL63 | NC_005831.2 | Human | No | |||
| NL63-related bat coronavirus strain BtKYNL63-9b | BtKYNL63 | NC_032107.1 | Bats | No | ||||
| Tegacovirus | Alphacoronavirus 1 | TGEV CCoV FeCoV | NC_038861.1 KP_849472.1 JQ_408980.1 | Porcines, canines, felines | No No No | |||
| Betacoronavirus | Embecovirus | Betacoronavirus 1 | HCoV-OC43 BCoV ECoV | NC_006213.1 NC_003045.1 EF_446615.1 | Human Bovines Equines | No No No | ||
| China Rattus coronavirus HKU24 | RtCoV-HKU24 | NC_026011.1 | Rats | No | ||||
| Human coronavirus HKU1 | HCoV-HKU1 | NC_006577.2 | Human | No | ||||
| Murine coronavirus | MHV | NC_001846.1 | Mouse | No | ||||
| Rabbit coronavirus HKU14 | RbCoV HKU14 | JN_874559 | Rabbits | No | ||||
| Hibecovirus | Bat Hp-betacoronavirus Zhejiang2013 | BtHpCoV-ZJ13 | NC_025217.1 | Bats | No | |||
| Merbecovirus | Hedgehog coronavirus 1 | EriCoV-1 | NC_039207.1 | Hedgehog | No | |||
| Middle East respiratory syndrome-related coronavirus | MERSr-CoV | NC_019843.3 | Human, camels, and bats | Yes | ||||
| Pipistrellus bat coronavirus HKU5 | BtPiCoV-HKU5 | NC_009020.1 | Bats | No | ||||
| Tylonycteris bat coronavirus HKU4 | BtTyCoV-HKU4 | NC_009019.1 | Bats | No | ||||
| Nobecovirus | Roussetus bat coronavirus GCCDC1 | BtEoCoV-GCCDC1 | NC_030886.1 | Bats | No | |||
| Roussetus bat coronavirus HKU9 | BtRoCoV-HKU9 | MG762674.1 | Bats | No | ||||
| Sarbecovirus | Severe acute respiratory syndrome-related coronavirus | SARSr-CoV | NC_004718.3 | Human, palm civets, and bats | Yes | |||
| Unclassified Betacoronavirus | Pangolin coronavirus | Pangolin-CoV | NA_606875.1 | Pangolins | No | |||
| Gammacoronavirus | Cegacovirus | Beluga whale coronavirus SW1 | BWCoV-SW1 | NC_010646.1 | Whale | No | ||
| Igacovirus | Avian coronavirus | IBV | NC_001451.1 | Birds | No | |||
| Deltacoronavirus | Andecovirus | Wigeon coronavirus HKU20 | WiCoV-HKU20 | NC_016995.1 | Birds | No | ||
| Buldecovirus | Bulbul coronavirus HKU11 | BuCoV-HKU11 | NC_011547.1 | Birds | No | |||
| Coronavirus HKU15 | PoCoV-HKU15 | NC_039208.1 | Pigs | No | ||||
| Munia coronavirus HKU13 | MuCoV-HKU13 | NC_011550.1 | Birds | No | ||||
| White-eye coronavirus HKU16 | WECoV-HKU13 | NC_016991.1 | Birds | No | ||||
| Herdecovirus | Night heron coronavirus HKU19 | NHCoV-HKU19 | NC_016994.1 | Birds | No | |||
| Moordecovirus | Common moorhen coronavirus HKU21 | CMCoV-HKU21 | NC_016996.1 | Birds | No |
| Country | Number of Complete Genomes | Partial Sequences/Incomplete Genomes | Total |
|---|---|---|---|
| Algeria, Argentina, Czech Republic, Greece, Hungary, Saudi Arabia, Slovenia | 3 * | 0 | 3 * |
| Australia | 391 | 0 | 391 |
| Austria | 21 | 0 | 21 |
| Belarus, Columbia, Pakistan, Thailand, Turkey | 2 * | 0 | 2 * |
| Belgium | 322 | 0 | 322 |
| Brazil | 36 | 0 | 36 |
| Cambodia, Ecuador, Lithuania, Mexico, Nepal, Nigeria, Panama, Poland, South Africa, Sweden | 1 * | 0 | 1 * |
| Canada | 129 | 0 | 129 |
| Chile | 7 | 0 | 7 |
| China | 346 | 47 | 393 |
| Congo | 42 | 0 | 42 |
| Denmark, Mexico | 9 * | 0 | 9 * |
| Finland | 40 | 0 | 40 |
| France | 204 | 0 | 204 |
| Georgia | 13 | 0 | 13 |
| Germany | 64 | 0 | 64 |
| Ghana | 15 | 0 | 15 |
| Hong Kong | 64 | 26 | 90 |
| Iceland | 601 | 0 | 601 |
| India | 32 | 1 | 33 |
| Indonesia, Philippines | 0 | 4 * | 4 * |
| Iran | 1 | 23 | 24 |
| Ireland, South Korea | 13 * | 0 | 13 * |
| Italy | 39 | 5 | 44 |
| Japan | 102 | 1 | 103 |
| Kuwait, New Zealand, Vietnam | 8 * | 0 | 8 * |
| Latvia | 5 | 0 | 5 |
| Russia, Slovakia, Estonia | 4 * | 0 | 4 * |
| Luxembourg | 86 | 0 | 86 |
| Malaysia | 7 | 3 | 10 |
| Netherlands | 585 | 0 | 585 |
| Norway | 29 | 0 | 29 |
| Peru | 1 | 1 | 2 |
| Portugal | 100 | 0 | 100 |
| Senegal | 23 | 0 | 23 |
| Singapore | 37 | 0 | 37 |
| South Africa | 6 | 0 | 6 |
| Spain | 105 | 0 | 105 |
| Switzerland | 52 | 0 | 52 |
| Taiwan | 22 | 0 | 22 |
| United Kingdom | 2540 | 1 | 2541 |
| USA | 1467 | 2 | 1469 |
| Total | 7655 | 118 | 7773 |
| Gene | From | To | Gene Length (Nucleotide) | Protein | Protein Length (Amino Acid) |
|---|---|---|---|---|---|
| 5′ UTR | 1 | 265 | 265 | Untranslated | – |
| orf1ab | 266 | 21,555 | 21,290 | pp1ab | 7096 |
| pp1a | 4405 | ||||
| S | 21,563 | 25,384 | 3822 | S | 1273 |
| orf3a | 25,393 | 26,220 | 828 | orf3a | 275 |
| E | 26,245 | 26,472 | 228 | E | 75 |
| M | 26,523 | 27,191 | 669 | M | 222 |
| orf6 | 27,202 | 27,387 | 186 | orf6 | 61 |
| orf7a | 27,394 | 27,759 | 366 | orf7a | 121 |
| orf7b | 27,756 | 27,887 | 132 | orf7b | 43 |
| orf8 | 27,894 | 28,259 | 366 | orf8 | 121 |
| N | 28,274 | 29,533 | 1260 | N | 419 |
| orf10 | 29,558 | 29,674 | 117 | orf10 | 38 |
| 3′UTR | 29,675 | 29,903 | 229 | Untranslated | – |
| Date | Number of Infected Countries and Territories | Cumulative Confirmed Cases | Cumulative Number of Deaths | ||||
|---|---|---|---|---|---|---|---|
| Globally | China | Outside China | Globally | China | Outside China | ||
| 13 December–6 January | 1 | 44 | 44 | 0 | 0 | 0 | 0 |
| 13 January | 2 | 45 | 44 | 1 | 1 | 1 | 0 |
| 20 January | 4 | 282 | 279 | 3 | 6 | 6 | 0 |
| 27 January | 12 | 2798 | 2741 | 37 | 80 | 80 | 0 |
| 3 February | 24 | 17,391 | 17,238 | 153 | 362 | 361 | 1 |
| 10 February | 25 | 40,554 | 40,235 | 319 | 910 | 909 | 1 |
| 17 February | 26 | 71,429 | 70,635 | 794 | 1775 | 1772 | 3 |
| 24 February | 30 | 79,331 | 77,262 | 2069 | 2618 | 2595 | 23 |
| 2 March | 65 | 88,948 | 80,174 | 8774 | 3043 | 2915 | 128 |
| 9 March | 105 | 109,577 | 80,904 | 28,673 | 3809 | 3123 | 686 |
| 16 March | 152 | 167,511 | 81,077 | 86,434 | 6606 | 3218 | 3388 |
| 23 March | 195 | 332,930 | 81,601 | 251,329 | 14,509 | 3267 | 11,242 |
| 30 March | 204 | 693,282 | 82,447 | 610,835 | 33,106 | 3310 | 29,796 |
| 6 April | 210 | 1,210,956 | 83,005 | 1,127,951 | 67,594 | 3340 | 64,254 |
| 13 April | 213 | 1,773,084 | 83,597 | 1,689,487 | 111,652 | 3351 | 108,301 |
| Drug/Treatment | Mode of Action | Antiviral Activity against COVID-19-Related Viruses | References |
|---|---|---|---|
| Lopinavir/Ritonavir | Protease inhibitor | - Provided good results against SARS-CoV decreased the viral load significantly and provided good results in COVID-19 patients | [97,98,109] |
| Arbidol | Inhibits membrane fusion | - Used for the treatment of influenza viruses in Russia and China - Lopinavir/ritonavir plus arbidol combination improved significantly the conditions of patients suffering from COVID-19 pneumonia | [102,110] |
| Interferon therapy | Inhibits many stages of virus replication: viral entry, transcription, replication, translation, assembly | - Opinavir/ritonavir plus interferon combination was used for the treatment of HIV infection - Ribavirin and interferon combination was used for the treatment of patients infected with MERS-CoV | [99,100,101] |
| Favipiravir | Inhibits viral RNA polymerase and mRNA capping | - Demonstrated an inhibitory effect on all influenza subtypes including neuraminidase- and M2 inhibitor-resistant strains - Showed inhibitory effects against Arenaviruses, Bunyaviruses, and Filoviruses | [111,112,113] |
| Chloroquine | Increases pH in host cell lysosomes and negatively influences virus–receptor binding, as well as interferes with the glycosylation of cellular receptors of SARS-CoV | - Exhibited a promising antiviral effect against SARS-CoV-2 in vitro - Improved COVID-19-pneumonia patients and shortened the course of the disease | [114] |
| Remdesivir | A monophosphoramidate of adenosine prodrug that incorporates into nascent viral RNA chains causing pre-mature termination | - Used against a wide range of RNA viruses such as Filoviridae, Paramyxoviridae, Pneumoviridae, and Coronaviridae; used successfully in COVID-19 treatment in the United States and showed no adverse events | [115] |
| Darunavir and Cobicistat | Inhibit 3 C-like protease (3CLpro). | - Used for the treatment of MERS-CoV in experimental animals - Used for the treatment of HIV-1 patients | [116] |
| Oseltamivir | Neuraminidase inhibitor | - Anti-influenza drug - Combination of nitazoxanide and oseltamivir is more effective in the treatment of ferrets infected with influenza virus compared to oseltamivir monotherapy | [97,117] |
| Steroid treatment (Methylprednisolon) | Anti-inflammatory | - Commonly used for the treatment of SARS patients suffering from severe pneumonia. - Because these drugs are immunosuppressive, they may delay viral clearance if given before viral replication is controlled | [102,118] |
| Convalescent plasma | SARS-CoV-2-neutralizing antibodies | - Immunotherapy combined with antiviral drugs is efficient against COVID-19 | [119,120] |
| Mesenchymal Stem Cells | Anti-inflammatory and immunomodulatory | - Enhances recovery in COVID-19 patients. | [121] |
| Ivermectin | Anti-parasitic and antiviral | - Inhibits the in vitro replication of SARS-CoV-2 on Vero-hSLAM cells with 5000-fold reduction in viral RNA in 48 hours | [122] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helmy, Y.A.; Fawzy, M.; Elaswad, A.; Sobieh, A.; Kenney, S.P.; Shehata, A.A. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. J. Clin. Med. 2020, 9, 1225. https://doi.org/10.3390/jcm9041225
Helmy YA, Fawzy M, Elaswad A, Sobieh A, Kenney SP, Shehata AA. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. Journal of Clinical Medicine. 2020; 9(4):1225. https://doi.org/10.3390/jcm9041225
Chicago/Turabian StyleHelmy, Yosra A., Mohamed Fawzy, Ahmed Elaswad, Ahmed Sobieh, Scott P. Kenney, and Awad A. Shehata. 2020. "The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control" Journal of Clinical Medicine 9, no. 4: 1225. https://doi.org/10.3390/jcm9041225
APA StyleHelmy, Y. A., Fawzy, M., Elaswad, A., Sobieh, A., Kenney, S. P., & Shehata, A. A. (2020). The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. Journal of Clinical Medicine, 9(4), 1225. https://doi.org/10.3390/jcm9041225