The Imbalance among Oxidative Biomarkers and Antioxidant Defense Systems in Thromboangiitis Obliterans (Winiwarter-Buerger Disease)





Abstract
1. Introduction
2. Patients and Methods
3. Results
3.1. Serum Levels of Total Oxidative Stress (TOS) and Total Antioxidant Capacity (TAC)
3.2. Serum Levels of Myeloperoxidase (MPO), Glutathione Reductase (GR), and Superoxide Dismutase (SOD) Activity
3.3. Serum Levels of Coenzyme Q10 (CoQ10) and Cholesterol
3.4. Biomarkers of Lipid Peroxidation and Protein Oxidation
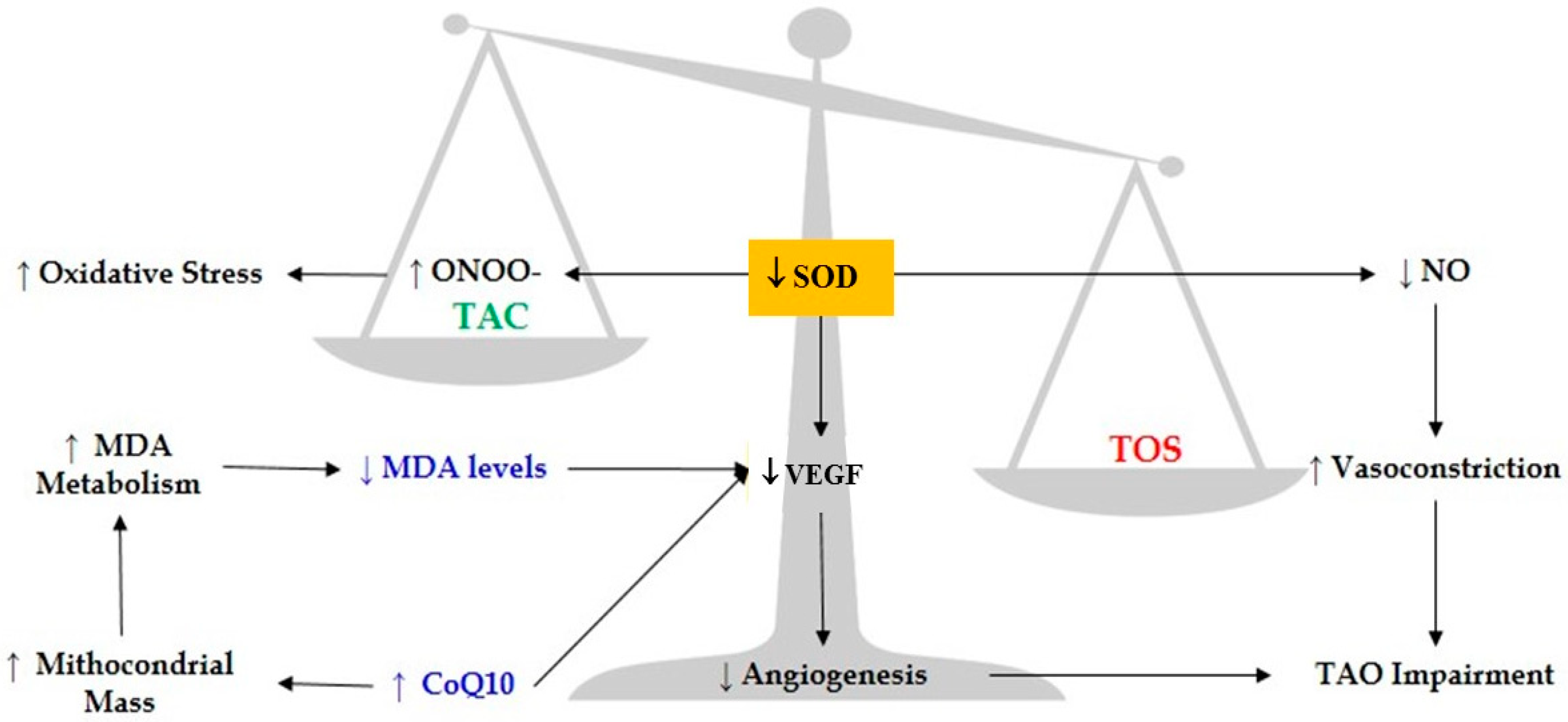
4. Discussions
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Von Winiwarter, F. Ueber eine eigenthumliche form von endoarteritis und endophlebitis mit gangran des fusses. Arch. Klin. Chir. 1879, 23, 202–226. [Google Scholar]
- Buerger, L. Thromboangiitis obliterans: A study of the vascular lesions leading to presenile spontaneous gangrene. Am. J. Med Sci. 1908, 136, 580–588. [Google Scholar] [CrossRef]
- Piazza, G.; Creager, M.A. Thromboangiitis obliterans. Circulation 2010, 121, 1858–1861. [Google Scholar] [CrossRef] [PubMed]
- Olin, J.W. Thromboangiitis obliterans: 110 years old and little progress made. J. Am. Heart Assoc. 2018, 7, e011214. [Google Scholar] [CrossRef]
- Desbois, A.C.; Cacoub, P. Cannabis-associated arterial disease. Ann. Vasc. Surg. 2013, 27, 996–1005. [Google Scholar] [CrossRef]
- Le Joncour, A.; Soudet, S.; Dupont, A.; Espitia, O.; Koskas, F.; Cluzel, P.; Hatron, P.Y.; Emmerich, J.; Cacoub, P.; Resche-Rigon, M.; et al. Long-term outcome and prognostic factors of complications in thromboangiitis obliterans (buerger’s disease): A multicenter study of 224 patients. J. Am. Heart Assoc. 2018, 7, e010677. [Google Scholar] [CrossRef] [PubMed]
- Cacione, D.G.; Baptista-Silva, J.C.; Macedo, C.R. Pharmacological treatment for buerger’s disease. Cochrane Database Syst. Rev. 2016, 2, CD011033. [Google Scholar] [PubMed]
- Fazeli, B.; Dadgar Moghadam, M.; Niroumand, S. How to treat a patient with thromboangiitis obliterans: A systematic review. Ann. Vasc. Surg. 2018, 49, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, B.M.; Karmota, A.G.; Abuhadema, A.M.; Shaker, A.A.; Abdelmawla, H.M.; Nasser, M.M.; Elmahdy, H.Y.; Mostafa, H.A.; Khairy, H.M. Management of buerger’s disease in endovascular era. Int. J. Angiol. Off. Publ. Int. Coll. Angiol. Inc. 2019, 28, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, B.; Rafatpanah, H.; Ravari, H.; Hosseini, R.F.; Rezaee, S.A. Investigation of the expression of mediators of neovascularization from mononuclear leukocytes in thromboangiitis obliterans. Vascular 2014, 22, 174–180. [Google Scholar] [CrossRef]
- Dellalibera-Joviliano, R.; Joviliano, E.E.; Silva, J.S.; Evora, P.R. Activation of cytokines corroborate with development of inflammation and autoimmunity in thromboangiitis obliterans patients. Clin. Exp. Immunol. 2012, 170, 28–35. [Google Scholar] [CrossRef]
- Keramat, S.; Sadeghian, M.H.; Keramati, M.R.; Fazeli, B. Assessment of t helper 17-associated cytokines in thromboangiitis obliterans. J. Inflamm. Res. 2019, 12, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, B.; Rafatpanah, H.; Ravari, H.; Farid Hosseini, R.; Tavakol Afshari, J.; Hamidi Alamdari, D.; Valizadeh, N.; Moheghi, N.; Rezaee, S.A. Sera of patients with thromboangiitis obliterans activated cultured human umbilical vein endothelial cells (huvecs) and changed their adhesive properties. Int. J. Rheum. Dis. 2014, 17, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Ketha, S.S.; Cooper, L.T. The role of autoimmunity in thromboangiitis obliterans (buerger’s disease). Ann. N. Y. Acad. Sci. 2013, 1285, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Halacheva, K.; Gulubova, M.V.; Manolova, I.; Petkov, D. Expression of icam-1, vcam-1, e-selectin and tnf-alpha on the endothelium of femoral and iliac arteries in thromboangiitis obliterans. Acta Histochem. 2002, 104, 177–184. [Google Scholar] [CrossRef]
- Iwai, T.; Inoue, Y.; Umeda, M. Buerger disease, smoking, and periodontitis. Ann. Vasc. Dis. 2008, 1, 80–84. [Google Scholar] [CrossRef]
- Chen, Y.W.; Umeda, M.; Nagasawa, T.; Takeuchi, Y.; Huang, Y.; Inoue, Y.; Iwai, T.; Izumi, Y.; Ishikawa, I. Periodontitis may increase the risk of peripheral arterial disease. Eur. J. Vasc. Endovasc. Surg. Off. J. Eur. Soc. Vasc. Surg. 2008, 35, 153–158. [Google Scholar] [CrossRef]
- Fazeli, B. Is rickettsia the key to solving the puzzle of buerger’s disease? Vascular 2014, 22, 393–394. [Google Scholar] [CrossRef]
- Fazeli, B.; Ravari, H.; Ghazvini, K. Rickettsia infection could be the missing piece of the buerger’s disease puzzle. Int. Angiol. J. Int. Union Angiol. 2017, 36, 410–416. [Google Scholar]
- Hewing, B.; Stangl, V.; Stangl, K.; Enke-Melzer, K.; Baumann, G.; Ludwig, A. Circulating angiogenic factors in patients with thromboangiitis obliterans. PLoS ONE 2012, 7, e34717. [Google Scholar] [CrossRef]
- Roncon-Albuquerque, R.; Serrao, P.; Vale-Pereira, R.; Costa-Lima, J.; Roncon-Albuquerque, R., Jr. Plasma catecholamines in buerger’s disease: Effects of cigarette smoking and surgical sympathectomy. Eur. J. Vasc. Endovasc. Surg. Off. J. Eur. Soc. Vasc. Surg. 2002, 24, 338–343. [Google Scholar] [CrossRef]
- Mousazadeh, B.; Sharebiani, H.; Taheri, H.; Valizedeh, N.; Fazeli, B. Unexpected inflammation in the sympathetic ganglia in thromboangiitis obliterans: More likely sterile or infectious induced inflammation? Clin. Mol. Allergy CMA 2019, 17, 10. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Cho, K.H.; Kim, K.L.; Kim, D.K.; Lee, T. Reduced circulating endothelial progenitor cells in thromboangiitis obliterans (buerger’s disease). Vasc. Med. 2013, 18, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Slavov, E.S.; Stanilova, S.A.; Petkov, D.P.; Dobreva, Z.G. Cytokine production in thromboangiitis obliterans patients: New evidence for an immune-mediated inflammatory disorder. Clin. Exp. Rheumatol. 2005, 23, 219–226. [Google Scholar] [PubMed]
- Matsushita, M.; Shionoya, S.; Matsumoto, T. Urinary cotinine measurement in patients with buerger’s disease-effects of active and passive smoking on the disease process. J. Vasc. Surg. 1991, 14, 53–58. [Google Scholar] [CrossRef]
- Chavoshan, A.; Sharebiani, H.; Taheri, H.; Fazeli, B. Antiphospholipid antibodies in buerger’s disease. Thromb. Res. 2019, 181, 64–66. [Google Scholar] [CrossRef]
- Akbarin, M.M.; Ravari, H.; Rajabnejad, A.; Valizadeh, N.; Fazeli, B. Investigation of the etiology of anemia in thromboangiitis obliterans. Int. J. Angiol. Off. Publ. Int. Coll. Angiol. Inc. 2016, 25, 153–158. [Google Scholar] [CrossRef]
- De Caridi, G.; Bitto, A.; Massara, M.; Pallio, G.; Pizzino, G.; Serra, R.; Altavilla, D.; Squadrito, F.; Spinelli, F. Increased serum hmgb-1, icam-1 and metalloproteinase-9 levels in buerger’s patients. Curr. Vasc. Pharmacol. 2016, 14, 382–387. [Google Scholar] [CrossRef]
- Li, M.D.; Wang, Y.F.; Yang, M.W.; Hong, F.F.; Yang, S.L. Risk factors, mechanisms and treatments of thromboangiitis obliterans: An overview of recent research. Curr. Med. Chem. 2019, 27. [Google Scholar] [CrossRef]
- Arslan, C.; Altan, H.; Besirli, K.; Aydemir, B.; Kiziler, A.R.; Denli, S. The role of oxidative stress and antioxidant defenses in buerger disease and atherosclerotic peripheral arterial occlusive disease. Ann. Vasc. Surg. 2010, 24, 455–460. [Google Scholar] [CrossRef]
- Alamdari, D.H.; Ravarit, H.; Tavallaie, S.; Fazeli, B. Oxidative and antioxidative pathways might contribute to thromboangiitis obliterans pathophysiology. Vascular 2014, 22, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Non-enzymatic antioxidant capacity assays: Limitations of use in biomedicine. Free Radic. Res. 2010, 44, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Anatoliotakis, N.; Deftereos, S.; Bouras, G.; Giannopoulos, G.; Tsounis, D.; Angelidis, C.; Kaoukis, A.; Stefanadis, C. Myeloperoxidase: Expressing inflammation and oxidative stress in cardiovascular disease. Curr. Top. Med. Chem. 2013, 13, 115–138. [Google Scholar] [CrossRef] [PubMed]
- Porto, B.N.; Stein, R.T. Neutrophil extracellular traps in pulmonary diseases: Too much of a good thing? Front. Immunol. 2016, 7, 311. [Google Scholar] [CrossRef]
- Kiyuna, L.A.; Albuquerque, R.P.E.; Chen, C.H.; Mochly-Rosen, D.; Ferreira, J.C.B. Targeting mitochondrial dysfunction and oxidative stress in heart failure: Challenges and opportunities. Free Radic. Biol. Med. 2018, 129, 155–168. [Google Scholar] [CrossRef]
- Cox, C.S.; McKay, S.E.; Holmbeck, M.A.; Christian, B.E.; Scortea, A.C.; Tsay, A.J.; Newman, L.E.; Shadel, G.S. Mitohormesis in mice via sustained basal activation of mitochondrial and antioxidant signaling. Cell Metab. 2018, 28, 776–786. [Google Scholar] [CrossRef]
- De Carvalho, F.O.; Felipe, F.A.; de Melo Costa, A.C.; Teixeira, L.G.; Silva, E.R.; Nunes, P.S.; Shanmugam, S.; de Lucca Junior, W.; Quintans, J.S.; de Souza Araujo, A.A. Inflammatory mediators and oxidative stress in animals subjected to smoke inhalation: A systematic review. Lung 2016, 194, 487–499. [Google Scholar] [CrossRef]
- Habashy, W.S.; Milfort, M.C.; Rekaya, R.; Aggrey, S.E. Cellular antioxidant enzyme activity and biomarkers for oxidative stress are affected by heat stress. Int. J. Biometeorol. 2019, 63, 1569–1584. [Google Scholar] [CrossRef]
- Shionoya, S. Diagnostic criteria of buerger’s disease. Int. J. Cardiol. 1998, 66, S243–S245. [Google Scholar] [CrossRef]
- Lazarides, M.K.; Georgiadis, G.S.; Papas, T.T.; Nikolopoulos, E.S. Diagnostic criteria and treatment of buerger’s disease: A review. Int. J. Low. Extrem. Wounds 2006, 5, 89–95. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Levine, R.L.; Wehr, N.; Williams, J.A.; Stadtman, E.R.; Shacter, E. Determination of carbonyl groups in oxidized proteins. Methods Mol. Biol. 2000, 99, 15–24. [Google Scholar] [PubMed]
- Sun, Y.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Ukeda, H.; Kawana, D.; Maeda, S.; Sawamura, M. Spectrophotometric assay for superoxide dismutase based on the reduction of highly water-soluble tetrazolium salts by xanthine-xanthine oxidase. Biosci. Biotechnol. Biochem. 1999, 63, 485–488. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Mills, J.L., Sr. Buerger’s disease in the 21st century: Diagnosis, clinical features, and therapy. Semin. Vasc. Surg. 2003, 16, 179–189. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 13. [Google Scholar] [CrossRef]
- Fazeli, B.; Rezaee, S.A. A review on thromboangiitis obliterans pathophysiology: Thrombosis and angiitis, which is to blame? Vascular 2011, 19, 141–153. [Google Scholar] [CrossRef]
- Mohareri, M.; Mirhosseini, A.; Mehraban, S.; Fazeli, B. Thromboangiitis obliterans episode: Autoimmune flare-up or reinfection? Vasc. Health Risk Manag. 2018, 14, 247–251. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Saeed Modaghegh, M.H.; Rahimi, H.R.; Ehteshamfar, S.M.; Tavakol Afshari, J. Molecular mechanisms regulating immune responses in thromboangiitis obliterans: A comprehensive review. Iran. J. Basic Med Sci. 2019, 22, 215–224. [Google Scholar] [PubMed]
- Fazeli, B.; Ravari, H. Mechanisms of thrombosis, available treatments and management challenges presented by thromboangiitis obliterans. Curr. Med. Chem. 2015, 22, 1992–2001. [Google Scholar] [CrossRef] [PubMed]
- Alukhanian, O.A.; Abramov Iu, G.; Korochanskaia, S.P. Indices of antioxidant system of blood in differential diagnosis of obliterating atherosclerosis and obliterating thromboangiitis of the lower limbs. Angiol. I Sosud. Khirurgiia Angiol. Vasc. Surg. 2007, 13, 25–30. [Google Scholar]
- Aliee, A.; Zahedi Avval, F.; Taheri, H.; Moghadam, S.M.; Soukhtanloo, M.; Hamidi Alamdari, D.; Fazeli, B. The status of nitric oxide and its backup, heme oxygenase 1, in thromboangiitis obliterans. Rep. Biochem. Mol. Biol. 2018, 6, 197–202. [Google Scholar]
- Lee, T.; Seo, J.W.; Sumpio, B.E.; Kim, S.J. Immunobiologic analysis of arterial tissue in buerger’s disease. Eur. J. Vasc. Endovasc. Surg. Off. J. Eur. Soc. Vasc. Surg. 2003, 25, 451–457. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadi, M.; Rahimi, H.R.; Esmaeili, H.; Mahmoudi, M.; Saeed Modaghegh, M.H.; Tavakol Afshari, J. The association of hla-a, b and drb1 with buerger’s disease. Rep. Biochem. Mol. Biol. 2019, 8, 153–160. [Google Scholar]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Glueck, C.J.; Haque, M.; Winarska, M.; Dharashivkar, S.; Fontaine, R.N.; Zhu, B.; Wang, P. Stromelysin-1 5a/6a and enos t-786c polymorphisms, mthfr c677t and a1298c mutations, and cigarette-cannabis smoking: A pilot, hypothesis-generating study of gene-environment pathophysiological associations with buerger’s disease. Clin. Appl. Thromb. Hemost. Off. J. Int. Acad. Clin. Appl. Thromb. Hemost. 2006, 12, 427–439. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- Ge, J.W.; Jiang, Y.P.; He, S.L. Changes in oxygen free radical and prostacyclin in thromboangiitis obliterans and its relationship with syndrome differentiation. Zhongguo Zhong Xi Yi Jie He Za Zhi Zhongguo Zhongxiyi Jiehe Zazhi Chin. J. Integr. Tradit. West. Med. 1993, 13, 652–654, 643. [Google Scholar]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vasc. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gegotek, A.; Skrzydlewska, E. Biological effect of protein modifications by lipid peroxidation products. Chem. Phys. Lipids 2019, 221, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 31. [Google Scholar] [CrossRef] [PubMed]
- Abdulhasan, M.K.; Li, Q.; Dai, J.; Abu-Soud, H.M.; Puscheck, E.E.; Rappolee, D.A. Coq10 increases mitochondrial mass and polarization, atp and oct4 potency levels, and bovine oocyte mii during ivm while decreasing ampk activity and oocyte death. J. Assist. Reprod. Genet. 2017, 34, 1595–1607. [Google Scholar] [CrossRef]
- El-Salahy, E.M.; Ahmed, M.I.; El-Gharieb, A.; Tawfik, H. New scope in angiogenesis: Role of vascular endothelial growth factor (vegf), no, lipid peroxidation, and vitamin e in the pathophysiology of pre-eclampsia among egyptian females. Clin. Biochem. 2001, 34, 323–329. [Google Scholar] [CrossRef]
- Fazeli, B.; Keramat, S.; Assadi, L.; Taheri, H. Angiogenesis induction in buerger’s disease: A disease management double-edged sword? Orphanet J. Rare Dis. 2019, 14, 189. [Google Scholar] [CrossRef]
- Olin, J.W.; Shih, A. Thromboangiitis obliterans (buerger’s disease). Curr. Opin. Rheumatol. 2006, 18, 18–24. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 294, 19683–19708. [Google Scholar] [CrossRef]
- Wilkie-Grantham, R.P.; Magon, N.J.; Harwood, D.T.; Kettle, A.J.; Vissers, M.C.; Winterbourn, C.C.; Hampton, M.B. Myeloperoxidase-dependent lipid peroxidation promotes the oxidative modification of cytosolic proteins in phagocytic neutrophils. J. Biol. Chem. 2015, 290, 9896–9905. [Google Scholar] [CrossRef]
- West, A.P. Mitochondrial dysfunction as a trigger of innate immune responses and inflammation. Toxicology 2017, 391, 54–63. [Google Scholar] [CrossRef]
- Pipinos, I.I.; Judge, A.R.; Zhu, Z.; Selsby, J.T.; Swanson, S.A.; Johanning, J.M.; Baxter, B.T.; Lynch, T.G.; Dodd, S.L. Mitochondrial defects and oxidative damage in patients with peripheral arterial disease. Free Radic. Biol. Med. 2006, 41, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Li, H.N.; Zimmerman, M.; Milledge, G.Z.; Hou, X.L.; Cheng, J.; Wang, Z.H.; Li, P.A. Water-soluble coenzyme q10 reduces rotenone-induced mitochondrial fission. Neurochem. Res. 2017, 42, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, H.; Ikeda, K.; Shinozaki, S.; Yasuhara, S.; Yu, Y.M.; Martyn, J.A.J.; Tompkins, R.G.; Yorozu, T.; Inoue, S.; Kaneki, M. Coenzyme q10 protects against burn-induced mitochondrial dysfunction and impaired insulin signaling in mouse skeletal muscle. FEBS Open Bio 2019, 9, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhu, H.; Wang, X.; Gao, Q.; Li, Z.; Huang, H. Coq10 ameliorates mitochondrial dysfunction in diabetic nephropathy through mitophagy. J. Endocrinol. 2019, 240, 445–465. [Google Scholar] [CrossRef]
- Yamamura, T.; Otani, H.; Nakao, Y.; Hattori, R.; Osako, M.; Imamura, H.; Das, D.K. Dual involvement of coenzyme q10 in redox signaling and inhibition of death signaling in the rat heart mitochondria. Antioxid. Redox Signal. 2001, 3, 103–112. [Google Scholar] [CrossRef]
- Widlansky, M.E.; Hill, R.B. Mitochondrial regulation of diabetic vascular disease: An emerging opportunity. Transl. Res. J. Lab. Clin. Med. 2018, 202, 83–98. [Google Scholar] [CrossRef]
- Choi, J.S.; Park, S.Y.; Yi, E.Y.; Kim, Y.J.; Jeong, J.W. Coenzyme q10 decreases basic fibroblast growth factor (bfgf)-induced angiogenesis by blocking erk activation. Oncol. Res. 2011, 19, 455–461. [Google Scholar] [CrossRef]
- Obradovic, M.; Essack, M.; Zafirovic, S.; Sudar-Milovanovic, E.; Bajic, V.P.; Van Neste, C.; Trpkovic, A.; Stanimirovic, J.; Bajic, V.B.; Isenovic, E.R. Redox control of vascular biology. Biofactors 2019, 37. [Google Scholar] [CrossRef]
- Kim, Y.W.; Byzova, T.V. Oxidative stress in angiogenesis and vascular disease. Blood 2014, 123, 625–631. [Google Scholar] [CrossRef]
- Oshikawa, J.; Urao, N.; Kim, H.W.; Kaplan, N.; Razvi, M.; McKinney, R.; Poole, L.B.; Fukai, T.; Ushio-Fukai, M. Extracellular sod-derived H2O2 promotes vegf signaling in caveolae/lipid rafts and post-ischemic angiogenesis in mice. PLoS ONE 2010, 5, e10189. [Google Scholar] [CrossRef]
- Mao, C.; Yuan, J.Q.; Lv, Y.B.; Gao, X.; Yin, Z.X.; Kraus, V.B.; Luo, J.S.; Chei, C.L.; Matchar, D.B.; Zeng, Y.; et al. Associations between superoxide dismutase, malondialdehyde and all-cause mortality in older adults: A community-based cohort study. BMC Geriatr. 2019, 19, 104. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Buerger’s Patients | Smoker Controls | p Value | |
|---|---|---|---|
| Total Oxidative Stress (TOS, μM) | 2.12 ± 0.5 | 0.06 ± 0.006 | 0.007 |
| Total Antioxidant Capacity (TAC, nM) | 0.32 ± 0.02 | 0.2 ± 0.05 | 0.04 |
| TOS-TAC Ratio | 6.4 ± 1.6 | 1.7 ± 1.2 | <0.001 |
| Myeloperoxidase (MPO, ng/mL) | 1.55 ± 0.61 | 1.42 ± 0.42 | 0.49 |
| Malondialdehyde (MDA, μM) | 5.3 ± 2.8 | 13 ± 6.2 | <0.001 |
| Superoxide Dismutase (SOD, U/L) | 52.1 ± 8.53 | 79.6 ± 31.8 | 0.002 |
| Glutathione Reductase (GR, U/L) | 43.5 ± 22.8 | 35.24 ± 25.1 | 0.22 |
| Protein Carbonyl (PC, ng/mL) | 22.4 ± 16.2 | 24.9 ± 16.2 | 0.83 |
| Coenzyme Q10 (CoQ10, pg/L) | 5.5 ± 1.2 | 3.7 ± 2.5 | <0.001 |
| Total Cholesterol (mg/dl) | 157.5 ± 35 | 193 ± 40 | 0.57 |
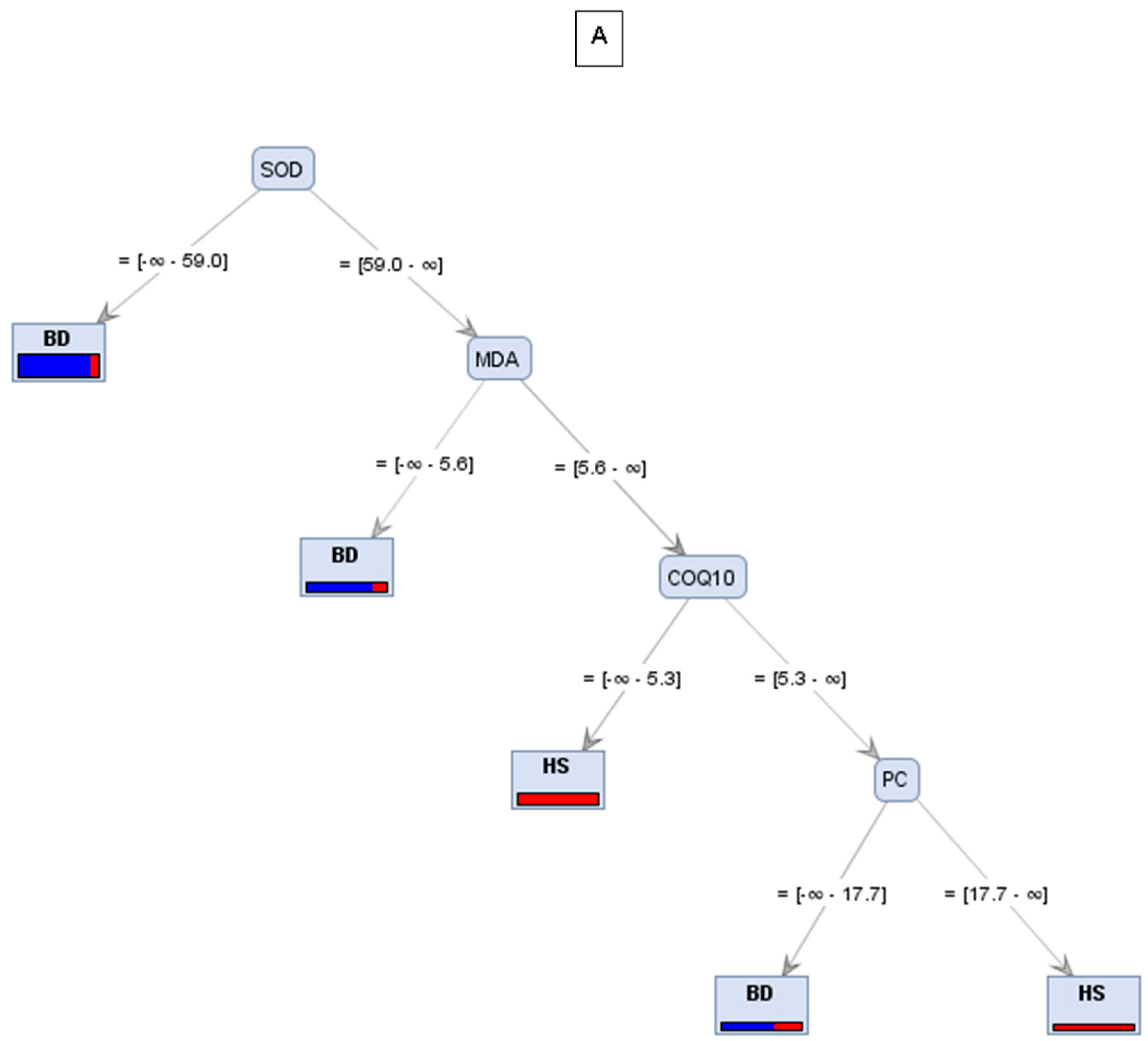
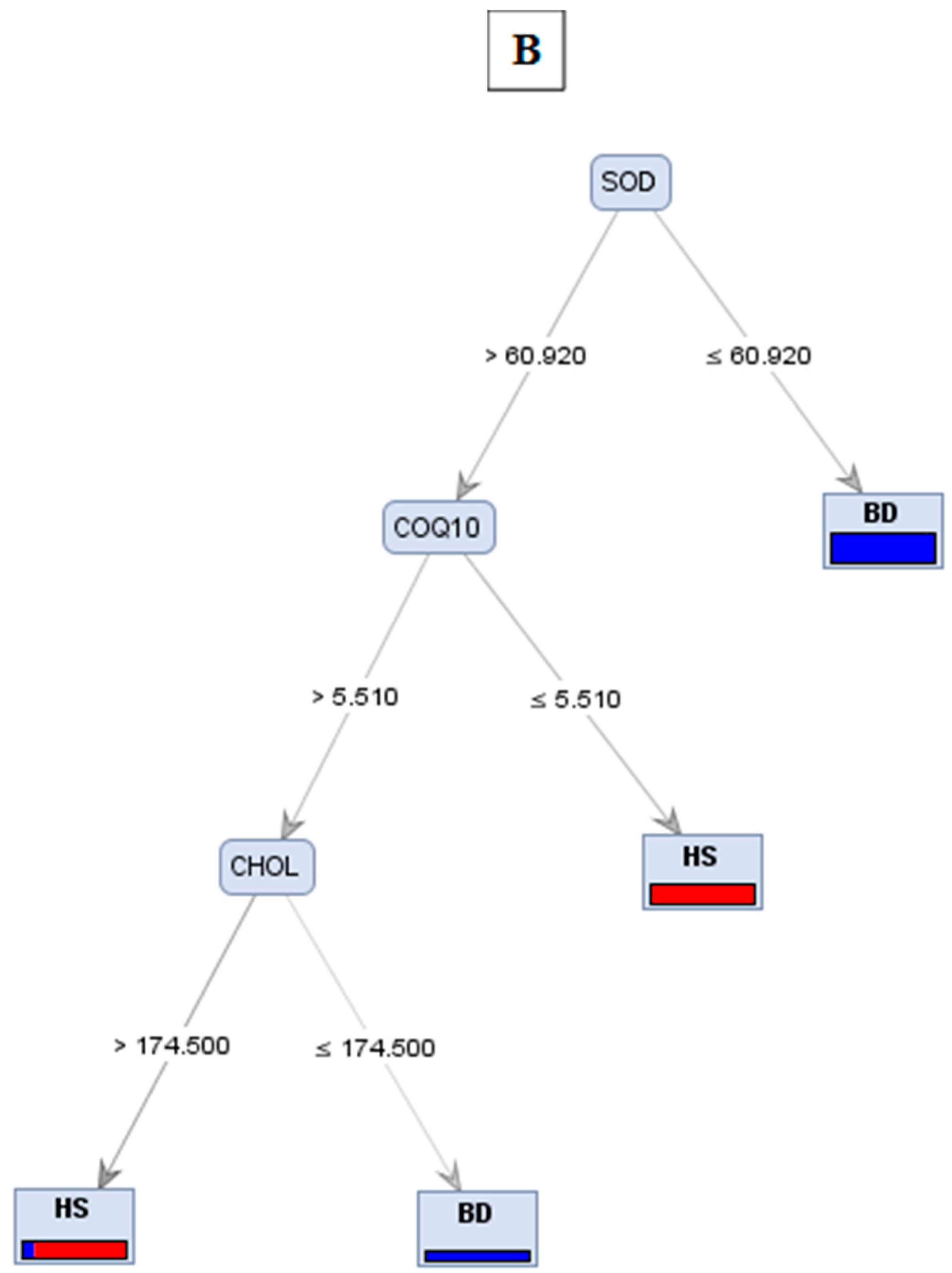
| Trees | Accuracy | Precision | Recall |
|---|---|---|---|
| A | 76.92% | 77.78% | 50% |
| B | 76.92% | 85.71% | 42.86% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharebiani, H.; Fazeli, B.; Maniscalco, R.; Ligi, D.; Mannello, F. The Imbalance among Oxidative Biomarkers and Antioxidant Defense Systems in Thromboangiitis Obliterans (Winiwarter-Buerger Disease). J. Clin. Med. 2020, 9, 1036. https://doi.org/10.3390/jcm9041036
Sharebiani H, Fazeli B, Maniscalco R, Ligi D, Mannello F. The Imbalance among Oxidative Biomarkers and Antioxidant Defense Systems in Thromboangiitis Obliterans (Winiwarter-Buerger Disease). Journal of Clinical Medicine. 2020; 9(4):1036. https://doi.org/10.3390/jcm9041036
Chicago/Turabian StyleSharebiani, Hiva, Bahare Fazeli, Rosanna Maniscalco, Daniela Ligi, and Ferdinando Mannello. 2020. "The Imbalance among Oxidative Biomarkers and Antioxidant Defense Systems in Thromboangiitis Obliterans (Winiwarter-Buerger Disease)" Journal of Clinical Medicine 9, no. 4: 1036. https://doi.org/10.3390/jcm9041036
APA StyleSharebiani, H., Fazeli, B., Maniscalco, R., Ligi, D., & Mannello, F. (2020). The Imbalance among Oxidative Biomarkers and Antioxidant Defense Systems in Thromboangiitis Obliterans (Winiwarter-Buerger Disease). Journal of Clinical Medicine, 9(4), 1036. https://doi.org/10.3390/jcm9041036