Radiation-Induced Salivary Gland Dysfunction: Mechanisms, Therapeutics and Future Directions

 ,
, 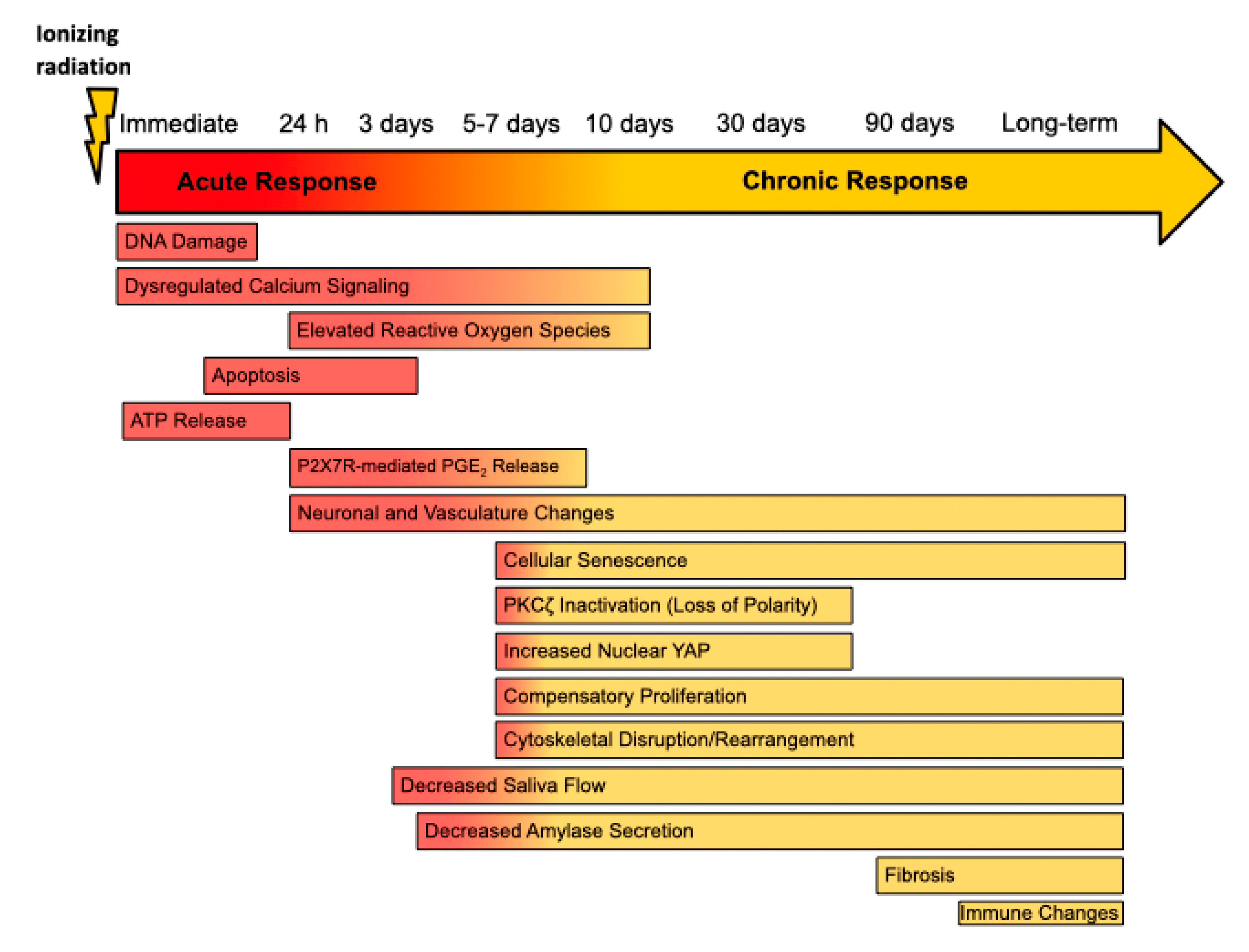{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Clinical Presentation
3. Animal Models Provide Mechanistic Insight into Radiation-Induced Salivary Gland Dysfunction
3.1. DNA Damage, Insufficient DNA Repair and Cell Cycle Arrest
3.2. Reactive Oxygen Species Generation
3.3. Dysregulated Calcium Signaling
3.4. Generation of Inflammatory Responses
3.5. Apoptosis, Autophagy and Cellular Senescence
3.6. Neuronal and Vascular Changes
3.7. Stem/Progenitor Cell Dysfunction
3.8. Compensatory Proliferation
3.9. Alterations in Cell Structure
3.10. Fibrosis
3.11. Immunomodulation
4. Bystander Effect: Potential Role for Purinergic Signaling
4.1. ATP Release
4.2. Dysregulated Calcium Signaling
4.3. MAPK Signaling
4.4. COX-2/Prostaglandin E2 Signaling
4.5. DNA Damage Response
4.6. Reactive Oxygen Species and TGF-β1
4.7. Immunomodulation
5. Therapeutics
5.1. Salivary Gland-Sparing Techniques
5.1.1. Dosing Strategies
5.1.2. Intensity Modulated Radiation Therapy (IMRT)
5.1.3. Volumetric Modulated Arc Therapy (VMAT)
5.1.4. Proton Beam Radiotherapy (PBRT)
5.1.5. Intraoral Stents and Temporary Submandibular Gland (SMG) Transplantation
5.2. Symptom Relief
5.2.1. Artificial Saliva
5.2.2. Sialagogues
5.3. Radioprotection
5.3.1. Amifostine
5.3.2. Promising Preclinical Studies
5.4. Regeneration
5.4.1. Gene Therapy
5.4.2. Stem Cell Therapies
5.4.3. Pharmacological Approaches
6. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Grundmann, O.; Mitchell, G.; Limesand, K. Sensitivity of Salivary Glands to Radiation: From Animal Models to Therapies. J. Dent. Res. 2009, 88, 894–903. [Google Scholar] [CrossRef]
- Konings, A.W.; Coppes, R.P.; Vissink, A. On the mechanism of salivary gland radiosensitivity. Int. J. Radiat. Oncol. 2005, 62, 1187–1194. [Google Scholar] [CrossRef]
- Arany, S.; Benoit, D.S.W.; Dewhurst, S.; Ovitt, C.E. Nanoparticle-mediated Gene Silencing Confers Radioprotection to Salivary Glands In Vivo. Mol. Ther. 2013, 21, 1182–1194. [Google Scholar] [CrossRef]
- Avila, J.L.; Grundmann, O.; Burd, R.; Limesand, K.H. Radiation-Induced Salivary Gland Dysfunction Results From p53-Dependent Apoptosis. Int. J. Radiat. Oncol. 2009, 73, 523–529. [Google Scholar] [CrossRef]
- Limesand, K.H.; Said, S.; Anderson, S.M. Suppression of Radiation-Induced Salivary Gland Dysfunction by IGF-1. PLoS ONE 2009, 4, e4663. [Google Scholar] [CrossRef]
- Limesand, K.H.; Schwertfeger, K.L.; Anderson, S.M. MDM2 Is Required for Suppression of Apoptosis by Activated Akt1 in Salivary Acinar Cells. Mol. Cell. Biol. 2006, 26, 8840–8856. [Google Scholar] [CrossRef]
- Stramandinoli-Zanicotti, R.T.; Sassi, L.M.; Schussel, J.L.; Torres, M.F.; Funchal, M.; Smaniotto, G.H.; Dissenha, J.L.; Carvalho, A.L. Effect of fractionated radiotherapy on the parotid gland: An experimental study in Brazilian minipigs. Int. Arch. Otorhinolaryngol. 2013, 17, 163–167. [Google Scholar]
- Coppes, R.P.; Meter, A.; Latumalea, S.P.; Roffel, A.F.; Kampinga, H.H. Defects in muscarinic receptor-coupled signal transduction in isolated parotid gland cells after in vivo irradiation: Evidence for a non-DNA target of radiation. Br. J. Cancer 2005, 92, 539–546. [Google Scholar] [CrossRef]
- Jensen, S.B.; Vissink, A.; Limesand, K.H.; Reyland, M. Salivary Gland Hypofunction and Xerostomia in Head and Neck Radiation Patients. J. Natl. Cancer Inst. Monogr. 2019, 2019. [Google Scholar] [CrossRef]
- Atkinson, J.C.; Grisius, M.; Massey, W. Salivary Hypofunction and Xerostomia: Diagnosis and Treatment. Dent. Clin. N. Am. 2005, 49, 309–326. [Google Scholar] [CrossRef]
- Khaw, A.; Logan, R.; Keefe, D.; Bartold, M. Radiation-induced oral mucositis and periodontitis-proposal for an inter-relationship. Oral Dis. 2014, 20, e7–e18. [Google Scholar] [CrossRef]
- Jensen, S.B.; Pedersen, A.M.L.; Vissink, A.; Andersen, E.; Brown, C.G.; Davies, A.N.; Dutilh, J.; Fulton, J.S.; Jankovic, L.; Lopes, N.N.F.; et al. A systematic review of salivary gland hypofunction and xerostomia induced by cancer therapies: Prevalence, severity and impact on quality of life. Support. Care Cancer 2010, 18, 1039–1060. [Google Scholar] [CrossRef]
- Wijers, O.B.; Levendag, P.C.; Braaksma, M.M.J.; Boonzaaijer, M.; Visch, L.L.; Schmitz, P.I.M. Patients with head and neck cancer cured by radiation therapy: A survey of the dry mouth syndrome in long-term survivors. Head Neck 2002, 24, 737–747. [Google Scholar] [CrossRef]
- Li, Y.; Taylor, J.M.G.; Haken, R.K.T.; Eisbruch, A. The impact of dose on parotid salivary recovery in head and neck cancer patients treated with radiation therapy. Int. J. Radiat. Oncol. 2007, 67, 660–669. [Google Scholar] [CrossRef]
- Fox, P.C. Salivary Enhancement Therapies. Caries Res. 2004, 38, 241–246. [Google Scholar] [CrossRef]
- Brizel, D.M.; Wasserman, T.H.; Henke, M.; Strnad, V.; Rudat, V.; Monnier, A.; Eschwege, F.; Zhang, J.; Russell, L.; Oster, W.; et al. Phase III randomized trial of amifostine as a radioprotector in head and neck cancer. J. Clin. Oncol. 2000, 18, 3339–3345. [Google Scholar] [CrossRef]
- King, M.; Joseph, S.; Albert, A.; Thomas, T.V.; Nittala, M.R.; Woods, W.C.; Vijayakumar, S.; Packianathan, S. Use of Amifostine for Cytoprotection during Radiation Therapy: A Review. Oncology 2019, 98, 61–80. [Google Scholar] [CrossRef]
- Cramer, J.D.; Burtness, B.; Le, Q.T.; Ferris, R.L. The changing therapeutic landscape of head and neck cancer. Nat. Rev. Clin. Oncol. 2019, 16, 669–683. [Google Scholar] [CrossRef]
- Dirix, P.; Nuyts, S.; Van den Bogaert, W. Radiation-induced xerostomia in patients with head and neck cancer: A literature review. Cancer 2006, 107, 2525–2534. [Google Scholar] [CrossRef]
- Dirix, P.; Nuyts, S.; Vander Poorten, V.; Delaere, P.; Van den Bogaert, W. The influence of xerostomia after radiotherapy on quality of life: Results of a questionnaire in head and neck cancer. Support Care Cancer 2008, 16, 171–179. [Google Scholar] [CrossRef]
- Pinna, R.; Campus, G.; Cumbo, E.; Mura, I.; Milia, E. Xerostomia induced by radiotherapy: An overview of the physiopathology, clinical evidence, and management of the oral damage. Ther. Clin. Risk Manag. 2015, 11, 171–188. [Google Scholar] [CrossRef]
- Meirovitz, A.; Murdoch-Kinch, C.A.; Schipper, M.; Pan, C.; Eisbruch, A. Grading xerostomia by physicians or by patients after intensity-modulated radiotherapy of head-and-neck cancer. Int. J. Radiat. Oncol. 2006, 66, 445–453. [Google Scholar] [CrossRef]
- Redman, R.S. On approaches to the functional restoration of salivary glands damaged by radiation therapy for head and neck cancer, with a review of related aspects of salivary gland morphology and development. Biotech. Histochem. 2008, 83, 103–130. [Google Scholar] [CrossRef]
- Emmerson, E.; May, A.J.; Berthoin, L.; Cruz-Pacheco, N.; Nathan, S.; Mattingly, A.J.; Chang, J.L.; Ryan, W.R.; Tward, A.D.; Knox, S.M. Salivary glands regenerate after radiation injury through SOX2-mediated secretory cell replacement. EMBO Mol. Med. 2018, 10, e8051. [Google Scholar] [CrossRef]
- Maria, O.M.; Eliopoulos, N.; Muanza, T. Radiation-Induced Oral Mucositis. Front. Oncol. 2017, 7, 89. [Google Scholar] [CrossRef]
- Trotti, A.; Bellm, L.; Epstein, J.B.; Frame, D.; Fuchs, H.J.; Gwede, C.K.; Komaroff, E.; Nalysnyk, L.; Zilberberg, M.D. Mucositis incidence, severity and associated outcomes in patients with head and neck cancer receiving radiotherapy with or without chemotherapy: A systematic literature review. Radiother. Oncol. 2003, 66, 253–262. [Google Scholar] [CrossRef]
- Funegard, U.; Franzen, L.; Ericson, T.; Henriksson, R. Parotid saliva composition during and after irradiation of head and neck cancer. Eur. J. Cancer B Oral Oncol. 1994, 30B, 230–233. [Google Scholar] [CrossRef]
- Wu, V.W.C.; Leung, K.Y. A Review on the Assessment of Radiation Induced Salivary Gland Damage after Radiotherapy. Front. Oncol. 2019, 9, 1090. [Google Scholar] [CrossRef]
- Makkonen, T.A.; Tenovuo, J.; Vilja, P.; Heimdahl, A. Changes in the protein composition of whole saliva during radiotherapy in patients with oral or pharyngeal cancer. Oral Surg. Oral Med. Oral Pathol. 1986, 62, 270–275. [Google Scholar] [CrossRef]
- Vesty, A.; Gear, K.; Boutell, S.; Taylor, M.W.; Douglas, R.G.; Biswas, K. Randomised, double-blind, placebo-controlled trial of oral probiotic Streptococcus salivarius M18 on head and neck cancer patients post-radiotherapy: A pilot study. Sci. Rep. 2020, 10, 13201. [Google Scholar] [CrossRef]
- Epstein, J.B.; Thariat, J.; Bensadoun, R.J.; Barasch, A.; Murphy, B.A.; Kolnick, L.; Popplewell, L.; Maghami, E. Oral complications of cancer and cancer therapy: From cancer treatment to survivorship. CA Cancer J. Clin. 2012, 62, 400–422. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.C.H.; Wu, V.W.C.; Kwong, D.L.W.; Ying, M.T.C. Assessment of post-radiotherapy salivary glands. Br. J. Radiol. 2011, 84, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Teymoortash, A.; Simolka, N.; Schrader, C.; Tiemann, M.; Werner, J. Lymphocyte subsets in irradiation-induced sialadenitis of the submandibular gland. Histopathology 2005, 47, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, J.O.; Sakuma, Y.; Seifert, G. Radiation-induced sialadenitis. Stage classification and immunohistology. Pathologist 1989, 10, 165–170. [Google Scholar]
- Sullivan, C.A.; Haddad, R.I.; Tishler, R.B.; Mahadevan, A.; Krane, J.F. Chemoradiation-Induced Cell Loss in Human Submandibular Glands. Laryngoscope 2005, 115, 958–964. [Google Scholar] [CrossRef]
- Luitje, M.E.; Israel, A.-K.; Cummings, M.A.; Giampoli, E.J.; Allen, P.D.; Newlands, S.D.; Ovitt, C.E. Long-Term Maintenance of Acinar Cells in Human Submandibular Glands After Radiation Therapy. Int. J. Radiat. Oncol. 2020, S0360–3016(20), 34483–34487. [Google Scholar] [CrossRef]
- Van Der Laan, H.P.; Bijl, H.P.; Steenbakkers, R.J.; Van Der Schaaf, A.; Chouvalova, O.; Hoek, J.G.V.-V.D.; Gawryszuk, A.; Van Der Laan, B.F.; Oosting, S.F.; Roodenburg, J.L.; et al. Acute symptoms during the course of head and neck radiotherapy or chemoradiation are strong predictors of late dysphagia. Radiother. Oncol. 2015, 115, 56–62. [Google Scholar] [CrossRef]
- Denham, J.W.; Peters, L.J.; Johansen, J.; Poulsen, M.; Lamb, D.S.; Hindley, A.; O’brien, P.C.; Spry, N.A.; Penniment, M.; Krawitz, H.; et al. Do acute mucosal reactions lead to consequential late reactions in patients with head and neck cancer? Radiother. Oncol. 1999, 52, 157–164. [Google Scholar] [CrossRef]
- Pal, M.; Gupta, N.; Rawat, S.; Grewal, M.S.; Garg, H.; Chauhan, D.; Ahlawat, P.; Tandon, S.; Khurana, R.; Pahuja, A.K.; et al. Radiation-induced dental caries, prevention and treatment–A systematic review. Natl. J. Maxillofac. Surg. 2015, 6, 160–166. [Google Scholar] [CrossRef]
- Sroussi, H.Y.; Epstein, J.B.; Bensadoun, R.-J.; Saunders, D.P.; Lalla, R.V.; Migliorati, C.A.; Heaivilin, N.; Zumsteg, Z.S. Common oral complications of head and neck cancer radiation therapy: Mucositis, infections, saliva change, fibrosis, sensory dysfunctions, dental caries, periodontal disease, and osteoradionecrosis. Cancer Med. 2017, 6, 2918–2931. [Google Scholar] [CrossRef]
- Roblegg, E.; Coughran, A.; Sirjani, D. Saliva: An all-rounder of our body. Eur. J. Pharm. Biopharm. 2019, 142, 133–141. [Google Scholar] [CrossRef]
- Spirk, C.; Hartl, S.; Pritz, E.; Gugatschka, M.; Kolb-Lenz, D.; Leitinger, G.; Roblegg, E. Comprehensive investigation of saliva replacement liquids for the treatment of xerostomia. Int. J. Pharm. 2019, 571, 118759. [Google Scholar] [CrossRef] [PubMed]
- Ambudkar, I.S. Calcium signaling defects underlying salivary gland dysfunction. Biochim. et Biophys. Acta (BBA) Bioenerg. 2018, 1865, 1771–1777. [Google Scholar] [CrossRef]
- Liu, X.; Ong, H.L.; Ambudkar, I.S. TRP Channel Involvement in Salivary Glands—Some Good, Some Bad. Cells 2018, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gong, B.; De Souza, L.B.; Ong, H.L.; Subedi, K.P.; Cheng, K.T.; Swaim, W.; Zheng, C.; Mori, Y.; Ambudkar, I.S. Radiation inhibits salivary gland function by promoting STIM1 cleavage by caspase-3 and loss of SOCE through a TRPM2-dependent pathway. Sci. Signal. 2017, 10, eaal4064. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cotrim, A.P.; Teos, L.Y.; Zheng, C.; Swaim, W.D.; Mitchell, J.B.; Mori, Y.; Ambudkar, I.S. Loss of TRPM2 function protects against irradiation-induced salivary gland dysfunction. Nat. Commun. 2013, 4, 1515. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.C.; Fillinger, J.L.; Sittadjody, S.; Avila, J.L.; Burd, R.; Limesand, K.H. IGF1 activates cell cycle arrest following irradiation by reducing binding of DeltaNp63 to the p21 promoter. Cell Death Dis. 2010, 1, e50. [Google Scholar] [CrossRef]
- Gilman, K.E.; Camden, J.M.; Klein, R.R.; Zhang, Q.; Weisman, G.A.; Limesand, K.H. P2X7 receptor deletion suppresses gamma-radiation-induced hyposalivation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, R687–R696. [Google Scholar] [CrossRef]
- Muhvic-Urek, M.; Bralic, M.; Curic, S.; Pezelj-Ribaric, S.; Borcic, J.; Tomac, J. Imbalance between apoptosis and proliferation causes late radiation damage of salivary gland in mouse. Physiol. Res. 2005, 55, 89–95. [Google Scholar]
- Wong, W.Y.; Pier, M.; Limesand, K.H. Persistent disruption of lateral junctional complexes and actin cytoskeleton in parotid salivary glands following radiation treatment. Am. J. Physiol. Integr. Comp. Physiol. 2018, 315, R656–R667. [Google Scholar] [CrossRef]
- Coppes, R.P.; Zeilstra, L.J.W.; Kampinga, H.H.; Konings, A.W.T. Early to late sparing of radiation damage to the parotid gland by adrenergic and muscarinic receptor agonists. Br. J. Cancer 2001, 85, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhu, Z.; Hai, B.; Chang, S.; Ma, L.; Xu, Y.; Li, X.; Feng, X.; Wu, X.; Zhao, Q.; et al. Intragland Shh gene delivery mitigated irradiation-induced hyposalivation in a miniature pig model. Theranostics 2018, 8, 4321–4331. [Google Scholar] [CrossRef] [PubMed]
- Knox, S.M.; Lombaert, I.M.A.; Haddox, C.L.; Abrams, S.R.; Cotrim, A.P.; Wilson, A.J.; Hoffman, M.P. Parasympathetic stimulation improves epithelial organ regeneration. Nat. Commun. 2013, 4, 1494. [Google Scholar] [CrossRef] [PubMed]
- Lombaert, I.M.; Patel, V.N.; Jones, C.E.; Villier, D.C.; Canada, A.E.; Moore, M.R.; Berenstein, E.; Zheng, C.; Goldsmith, C.M.; Chorini, J.A.; et al. CERE-120 Prevents Irradiation-Induced Hypofunction and Restores Immune Homeostasis in Porcine Salivary Glands. Mol. Ther. Methods Clin. Dev. 2020, 18, 839–855. [Google Scholar] [CrossRef]
- Chibly, A.M.; Wong, W.Y.; Pier, M.; Cheng, H.; Mu, Y.; Chen, J.; Ghosh, S.; Limesand, K.H. aPKCζ-dependent repression of Yap is necessary for functional restoration of irradiated salivary glands with IGF-1. Sci. Rep. 2018, 8, 6347. [Google Scholar] [CrossRef]
- Morgan-Bathke, M.; Hill, G.A.; Harris, Z.I.; Lin, H.H.; Chibly, A.M.; Klein, R.R.; Burd, R.S.; Ann, D.K.; Limesand, K.H. Autophagy Correlates with Maintenance of Salivary Gland Function Following Radiation. Sci. Rep. 2015, 4, 5206. [Google Scholar] [CrossRef] [PubMed]
- Radfar, L.; Sirois, D. Structural and functional injury in minipig salivary glands following fractionated exposure to 70 Gy of ionizing radiation: An animal model for human radiation-induced salivary gland injury. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2003, 96, 267–274. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, K.M.; Jung, M.H.; Jung, J.H.; Kang, K.M.; Jeong, B.K.; Park, J.J.; Woo, S.H.; Kim, J.P. Protective effects of alpha lipoic acid on radiation-induced salivary gland injury in rats. Oncotarget 2016, 7, 29143–29153. [Google Scholar] [CrossRef]
- Meyer, S.; Chibly, A.; Burd, R.; Limesand, K. Insulin-Like Growth Factor-1–Mediated DNA Repair in Irradiated Salivary Glands Is Sirtuin-1 Dependent. J. Dent. Res. 2016, 96, 225–232. [Google Scholar] [CrossRef]
- Hai, B.; Zhao, Q.; Deveau, M.A.; Liu, F. Delivery of Sonic Hedgehog Gene Repressed Irradiation-induced Cellular Senescence in Salivary Glands by Promoting DNA Repair and Reducing Oxidative Stress. Theranostics 2018, 8, 1159–1167. [Google Scholar]
- Akyuz, M.; Taysi, S.; Baysal, E.; Demir, E.; Alkis, H.; Akan, M.; Binici, H.; Karatas, Z.A. Radioprotective effect of thymoquinone on salivary gland of rats exposed to total cranial irradiation. Head Neck 2017, 39, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.Y.; Allie, S.; Limesand, K.H. PKCζ and JNK signaling regulate radiation-induced compensatory proliferation in parotid salivary glands. PLoS ONE 2019, 14, e0219572. [Google Scholar] [CrossRef] [PubMed]
- Humphries, M.J.; Limesand, K.H.; Schneider, J.C.; Nakayama, K.I.; Anderson, S.M.; Reyland, M.E. Suppression of apoptosis in the protein kinase c-δ null mouse in vivo. J. Biol. Chem. 2006, 281, 9728–9737. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.Y.; Gilman, K.; Limesand, K.H. Yap activation in irradiated parotid salivary glands is regulated by ROCK activity. PLoS ONE 2020, 15, e0232921. [Google Scholar] [CrossRef]
- Hill, G.; Headon, D.; Harris, Z.I.; Huttner, K.; Limesand, K.H. Pharmacological Activation of the EDA/EDAR Signaling Pathway Restores Salivary Gland Function following Radiation-Induced Damage. PLoS ONE 2014, 9, e112840. [Google Scholar] [CrossRef]
- Grundmann, O.; Fillinger, J.L.; Victory, K.R.; Burd, R.; Limesand, K.H. Restoration of radiation therapy-induced salivary gland dysfunction in mice by post therapy IGF-1 administration. BMC Cancer 2010, 10, 417. [Google Scholar] [CrossRef]
- Marmary, Y.; Adar, R.; Gaska, S.; Wygoda, A.; Maly, A.; Cohen, J.; Eliashar, R.; Mizrachi, L.; Orfaig-Geva, C.; Baum, B.J.; et al. Radiation-induced loss of salivary gland function is driven by cellular senescence and prevented by IL6 modulation. Cancer Res. 2016, 76, 1170–1180. [Google Scholar] [CrossRef]
- Peng, X.; Wu, Y.; Brouwer, U.; Van Vliet, T.; Wang, B.; DeMaria, M.; Barazzuol, L.; Coppes, R.P. Cellular senescence contributes to radiation-induced hyposalivation by affecting the stem/progenitor cell niche. Cell Death Dis. 2020, 11, 854. [Google Scholar] [CrossRef]
- Yokoyama, M.; Narita, T.; Sakurai, H.; Katsumata-Kato, O.; Sugiya, H.; Fujita-Yoshigaki, J. Maintenance of claudin-3 expression and the barrier functions of intercellular junctions in parotid acinar cells via the inhibition of Src signaling. Arch. Oral Biol. 2017, 81, 141–150. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeong, B.K.; Jang, S.J.; Yun, J.W.; Jung, M.H.; Kang, K.M.; Kim, T.; Woo, S.H. Alpha-Lipoic Acid Ameliorates Radiation-Induced Salivary Gland Injury by Preserving Parasympathetic Innervation in Rats. Int. J. Mol. Sci. 2020, 21, 2260. [Google Scholar] [CrossRef]
- De La Cal, C.; Fernández-Solari, J.; Mohn, C.; Prestifilippo, J.P.; Pugnaloni, A.; Medina, V.; Elverdin, J. Radiation Produces Irreversible Chronic Dysfunction in the Submandibular Glands of the Rat. Open Dent. J. 2012, 6, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.L.; Hill, G.A.; Klein, R.R.; Arnett, D.G.; Burd, R.; Limesand, K.H. Prevention of Radiation-Induced Salivary Gland Dysfunction Utilizing a CDK Inhibitor in a Mouse Model. PLoS ONE 2012, 7, e51363. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.-S.; Lee, S.; Kim, Y.M.; Lim, J. Hypoxia-Activated Adipose Mesenchymal Stem Cells Prevents Irradiation-Induced Salivary Hypofunction by Enhanced Paracrine Effect Through Fibroblast Growth Factor 10. Stem Cells 2018, 36, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Limesand, K.H.; Avila, J.L.; Victory, K.; Chang, H.H.; Shin, Y.J.; Grundmann, O.; Klein, R.R. IGF-1 preserves salivary gland function following fractionated radiation. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, 579–586. [Google Scholar] [CrossRef]
- Choi, J.-S.; Shin, H.-S.; An, H.-Y.; Kim, Y.-M.; Lim, J.-Y. Radioprotective effects of Keratinocyte Growth Factor-1 against irradiation-induced salivary gland hypofunction. Oncotarget 2017, 8, 13496–13508. [Google Scholar] [CrossRef]
- Lamas, D.J.M.; Carabajal, E.; Prestifilippo, J.P.; Rossi, L.; Elverdín, J.C.; Merani, S.; Bergoc, R.M.; Rivera, E.S.; Medina, V. Protection of Radiation-Induced Damage to the Hematopoietic System, Small Intestine and Salivary Glands in Rats by JNJ7777120 Compound, a Histamine H4 Ligand. PLoS ONE 2013, 8, e69106. [Google Scholar]
- Wie, S.M.; Adwan, T.S.; DeGregori, J.; Anderson, S.M.; Reyland, M.E. Inhibiting Tyrosine Phosphorylation of Protein Kinase Cδ (PKCδ) Protects the Salivary Gland from Radiation Damage. J. Biol. Chem. 2014, 289, 10900–10908. [Google Scholar] [CrossRef]
- Wie, S.M.; Wellberg, E.; Karam, S.D.; Reyland, M.E. Tyrosine Kinase Inhibitors Protect the Salivary Gland from Radiation Damage by Inhibiting Activation of Protein Kinase C-δ. Mol. Cancer Ther. 2017, 16, 1989–1998. [Google Scholar] [CrossRef]
- Heuckeroth, R.O.; Enomoto, H.; Grider, J.R.; Golden, J.P.; Hanke, J.; Jackman, A.; Molliver, D.C.; Bardgett, M.E.; Snider, W.D.; Johnson, E.M.; et al. Gene targeting reveals a critical role for neurturin in the development and maintenance of enteric, sensory, and parasympathetic neurons. Neuron 1999, 22, 253–263. [Google Scholar] [CrossRef]
- Rossi, J.; Luukko, K.; Poteryaev, D.; Laurikainen, A.; Sun, Y.F.; Laakso, T.; Eerikainen, S.; Tuominen, R.; Lakso, M.; Rauvala, H.; et al. Retarded growth and deficits in the enteric and parasympathetic nervous system in mice lacking GFR alpha2, a functional neurturin receptor. Neuron 1999, 22, 243–252. [Google Scholar] [CrossRef]
- Ferreira, J.N.; Zheng, C.; Lombaert, I.M.; Goldsmith, C.M.; Cotrim, A.P.; Symonds, J.M.; Patel, V.N.; Hoffman, M.P. Neurturin Gene Therapy Protects Parasympathetic Function to Prevent Irradiation-Induced Murine Salivary Gland Hypofunction. Mol. Ther. Methods Clin. Dev. 2018, 9, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Mizrachi, A.; Cotrim, A.P.; Katabi, N.; Mitchell, J.B.; Verheij, M.; Haimovitz-Friedman, A. Radiation-Induced Microvascular Injury as a Mechanism of Salivary Gland Hypofunction and Potential Target for Radioprotectors. Radiat. Res. 2016, 186, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Lombaert, I.M.A.; Brunsting, J.F.; Wierenga, P.K.; Kampinga, H.H.; De Haan, G.; Coppes, R.P. Cytokine Treatment Improves Parenchymal and Vascular Damage of Salivary Glands after Irradiation. Clin. Cancer Res. 2008, 14, 7741–7750. [Google Scholar] [CrossRef]
- Van Luijk, P.; Pringle, S.; Deasy, J.O.; Moiseenko, V.V.; Faber, H.; Hovan, A.; Baanstra, M.; Laan, H.P.; Kierkels, R.G.; Schaaf, A.; et al. Sparing the region of the salivary gland containing stem cells preserves saliva production after radiotherapy for head and neck cancer. Sci. Transl. Med. 2015, 7, 305ra147. [Google Scholar] [CrossRef]
- Aure, M.H.; Konieczny, S.F.; Ovitt, C.E. Salivary Gland Homeostasis Is Maintained through Acinar Cell Self-Duplication. Dev. Cell 2015, 33, 231–237. [Google Scholar] [CrossRef]
- Lombaert, I.M.A.; Brunsting, J.F.; Wierenga, P.K.; Faber, H.; Stokman, M.A.; Kok, T.; Visser, W.H.; Kampinga, H.H.; De Haan, G.; Coppes, R.P. Rescue of Salivary Gland Function after Stem Cell Transplantation in Irradiated Glands. PLoS ONE 2008, 3, e2063. [Google Scholar] [CrossRef]
- Weng, P.-L.; Aure, M.H.; Maruyama, T.; Ovitt, C.E. Limited Regeneration of Adult Salivary Glands after Severe Injury Involves Cellular Plasticity. Cell Rep. 2018, 24, 1464–1470.e3. [Google Scholar] [CrossRef] [PubMed]
- Morgan-Bathke, M.; Harris, Z.I.; Arnett, D.G.; Klein, R.R.; Burd, R.; Ann, D.K.; Limesand, K.H. The Rapalogue, CCI-779, Improves Salivary Gland Function following Radiation. PLoS ONE 2014, 9, e113183. [Google Scholar] [CrossRef]
- O’Keefe, K.; DeSantis, K.; Altrieth, A.; Nelson, D.; Taroc, E.; Stabell, A.; Pham, M.; Larsen, M. Regional Differences following Partial Salivary Gland Resection. J. Dent. Res. 2019, 99, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.S.; Fu, K.; Marks, J.; Silverman, S. Late effects of radiation therapy in the head and neck region. Int. J. Radiat. Oncol. 1995, 31, 1141–1164. [Google Scholar] [CrossRef]
- Gyorfi, A.H.; Matei, A.E.; Distler, J.H.W. Targeting TGF-beta signaling for the treatment of fibrosis. Matrix Biol. 2018, 68–69, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Ignotz, R.A.; Massague, J. Transforming growth factor-beta stimulates the expression of fibronectin and collagen and their incorporation into the extracellular matrix. J. Biol. Chem. 1986, 261, 4337–4345. [Google Scholar] [PubMed]
- Meng, X.M.; Huang, X.R.; Xiao, J.; Chen, H.Y.; Zhong, X.; Chung, A.C.; Lan, H.Y. Diverse roles of TGF-beta receptor II in renal fibrosis and inflammation in vivo and in vitro. J. Pathol. 2012, 227, 175–188. [Google Scholar] [CrossRef]
- Hakim, S.G.; Ribbat, J.; Berndt, A.; Richter, P.; Kosmehl, H.; Benedek, G.A.; Jacobsen, H.C.; Trenkle, T.; Sieg, P.; Rades, D.; et al. Expression of Wnt-1, TGF-beta and related cell-cell adhesion components following radiotherapy in salivary glands of patients with manifested radiogenic xerostomia. Radiother. Oncol. 2011, 101, 93–99. [Google Scholar] [CrossRef]
- Spiegelberg, L.; Swagemakers, S.M.; Van Ijcken, W.F.; Oole, E.; Wolvius, E.B.; Essers, J.; Braks, J.A.M. Gene expression analysis reveals inhibition of radiation-induced TGFbeta-signaling by hyperbaric oxygen therapy in mouse salivary glands. Mol. Med. 2014, 20, 257–269. [Google Scholar] [CrossRef]
- Woods, L.T.; Camden, J.M.; El-Sayed, F.G.; Khalafalla, M.G.; Petris, M.J.; Erb, L.; Weisman, G.A. Increased expression of TGF-beta signaling components in a mouse model of fibrosis induced by submandibular gland duct ligation. PLoS ONE 2015, 10, e0123641. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, J.Y.; Kim, J.M.; Yoo, B.R.; Han, S.Y.; Jung, Y.J.; Bae, H.; Cho, J. PM014 attenuates radiation-induced pulmonary fibrosis via regulating NF-kB and TGF-b1/NOX4 pathways. Sci. Rep. 2020, 10, 16112. [Google Scholar] [CrossRef]
- Flechsig, P.; Dadrich, M.; Bickelhaupt, S.; Jenne, J.; Hauser, K.; Timke, C.; Peschke, P.; Hahn, E.W.; Gröne, H.; Yingling, J.; et al. LY2109761 attenuates radiation-induced pulmonary murine fibrosis via reversal of TGF-beta and BMP-associated proinflammatory and proangiogenic signals. Clin. Cancer Res. 2012, 18, 3616–3627. [Google Scholar] [CrossRef]
- Liu, Y.; Kudo, K.; Abe, Y.; Hu, D.-L.; Kijima, H.; Nakane, A.; Ono, K. Inhibition of transforming growth factor-beta, hypoxia-inducible factor-1alpha and vascular endothelial growth factor reduced late rectal injury induced by irradiation. J. Radiat. Res. 2009, 50, 233–239. [Google Scholar] [CrossRef][Green Version]
- Campbell, A.M.; Decker, R.H. Harnessing the immunomodulatory effects of radiation therapy. Oncology 2018, 32, 370–374. [Google Scholar]
- Wirsdörfer, F.; Jendrossek, V. The Role of Lymphocytes in Radiotherapy-Induced Adverse Late Effects in the Lung. Front. Immunol. 2016, 7, 591. [Google Scholar]
- Daguenet, E.; Louati, S.; Wozny, A.-S.; Vial, N.; Gras, M.; Guy, J.-B.; Vallard, A.; Rodriguez-Lafrasse, C.; Magné, N. Radiation-induced bystander and abscopal effects: Important lessons from preclinical models. Br. J. Cancer 2020, 123, 339–348. [Google Scholar] [CrossRef]
- Tsuda, E.; Kawanishi, G.; Ueda, M.; Masuda, S.; Sasaki, R. The role of carbohydrate in recombinant human erythropoietin. JBIC J. Biol. Inorg. Chem. 1990, 188, 405–411. [Google Scholar] [CrossRef]
- Horsburgh, S.; Todryk, S.M.; Ramming, A.; Distler, J.H.; O’Reilly, S. Innate lymphoid cells and fibrotic regulation. Immunol. Lett. 2018, 195, 38–44. [Google Scholar] [CrossRef]
- Mikami, Y.; Takada, Y.; Hagihara, Y.; Kanai, T. Innate lymphoid cells in organ fibrosis. Cytokine Growth Factor Rev. 2018, 42, 27–36. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, J.; Tian, Z.; Van Velkinburgh, J.C.; Song, J.; Wu, Y.; Ni, B. Innate Lymphoid Cells: A Promising New Regulator in Fibrotic Diseases. Int. Rev. Immunol. 2015, 35, 399–414. [Google Scholar] [CrossRef]
- Cortez, V.S.; Cervantes-Barragan, L.; Robinette, M.L.; Bando, J.K.; Wang, Y.; Geiger, T.L.; Gilfillan, S.; Fuchs, A.; Vivier, E.; Sun, J.C.; et al. Transforming growth factor-beta signaling guides the differentiation of innate lymphoid cells in salivary glands. Immunity 2016, 44, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Cortez, V.S.; Fuchs, A.; Cella, M.; Gilfillan, S.; Colonna, M. Cutting edge: Salivary gland NK cells develop independently of Nfil3 in steady-state. J. Immunol. 2014, 192, 4487–4491. [Google Scholar] [CrossRef] [PubMed]
- Tsukimoto, M.; Homma, T.; Ohshima, Y.; Kojima, S. Involvement of purinergic signaling in cellular response to gamma radiation. Radiat. Res. 2010, 173, 298–309. [Google Scholar] [CrossRef]
- Hamada, N.; Maeda, M.; Otsuka, K.; Tomita, M. Signaling pathways underpinning the manifestations of ionizing radiation-induced bystander effects. Curr. Mol. Pharmacol. 2011, 4, 79–95. [Google Scholar] [CrossRef]
- Heeran, A.B.; Berrigan, H.P.; O’Sullivan, J.N. The Radiation-Induced Bystander Effect (RIBE) and its Connections with the Hallmarks of Cancer. Radiat. Res. 2019, 192, 668–679. [Google Scholar] [CrossRef]
- Christen, O.; Regad, C.; Neroni, M.; Thoenen, S.; Holz, J. [Substitute model of an in-vitro biological trials. I. Standardized method using human pulp cells]. J. Boil. Buccale 1989, 17, 275–284. [Google Scholar]
- Prise, K.M.; O’Sullivan, J.M. Radiation-induced bystander signalling in cancer therapy. Nat. Rev. Cancer 2009, 9, 351–360. [Google Scholar] [CrossRef]
- Kirolikar, S.; Prasannan, P.; Raghuram, G.V.; Pancholi, N.; Saha, T.; Tidke, P.; Chaudhari, P.; Shaikh, A.; Rane, B.; Pandey, R.; et al. Prevention of radiation-induced bystander effects by agents that inactivate cell-free chromatin released from irradiated dying cells. Cell Death Dis. 2018, 9, 1142. [Google Scholar] [CrossRef]
- Klammer, H.; Mladenov, E.; Li, F.; Iliakis, G. Bystander effects as manifestation of intercellular communication of DNA damage and of the cellular oxidative status. Cancer Lett. 2015, 356, 58–71. [Google Scholar] [CrossRef]
- Najafi, M.; Fardid, R.; Hadadi, G.; Fardid, M. The mechanisms of radiation-induced bystander effect. J. Biomed. Phys. Eng. 2014, 4, 163–172. [Google Scholar]
- Matsumoto, H.; Hayashi, S.; Hatashita, M.; Ohnishi, K.; Shioura, H.; Ohtsubo, T.; Kitai, R.; Ohnishi, T.; Kano, E. Induction of radioresistance by a nitric oxide-mediated bystander effect. Radiat. Res. 2001, 155, 387–396. [Google Scholar] [CrossRef]
- Azzam, E.I.; de Toledo, S.M.; Little, J.B. Direct evidence for the participation of gap junction-mediated intercellular communication in the transmission of damage signals from alpha -particle irradiated to nonirradiated cells. Proc. Natl. Acad. Sci. USA 2001, 98, 473–478. [Google Scholar] [CrossRef]
- Zhou, H.; Randers-Pehrson, G.; Waldren, C.A.; Vannais, D.; Hall, E.J.; Hei, T.K. Induction of a bystander mutagenic effect of alpha particles in mammalian cells. Proc. Natl. Acad. Sci. USA 2000, 97, 2099–2104. [Google Scholar] [CrossRef]
- Azzam, E.I.; de Toledo, S.M.; Gooding, T.; Little, J.B. Intercellular communication is involved in the bystander regulation of gene expression in human cells exposed to very low fluences of alpha particles. Radiat. Res. 1998, 150, 497–504. [Google Scholar] [CrossRef]
- Tsukimoto, M. Purinergic signaling is a novel mechanism of the cellular response to ionizing radiation. Biol. Pharm. Bull. 2015, 38, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, Y.; Tsukimoto, M.; Harada, H.; Kojima, S. Involvement of connexin43 hemichannel in ATP release after gamma-irradiation. J. Radiat. Res. 2012, 53, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, Y.; Tsukimoto, M.; Takenouchi, T.; Harada, H.; Suzuki, A.; Sato, M.; Kitani, H.; Kojima, S. Gamma-Irradiation induces P2X(7) receptor-dependent ATP release from B16 melanoma cells. Biochim. Biophys. Acta 2010, 1800, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.; Ohshima, Y.; Nakatsukasa, H.; Tsukimoto, M. Role of ATP as a key signaling molecule mediating radiation-induced biological effects. Dose Response 2017, 15, 1559325817690638. [Google Scholar] [CrossRef]
- Tanamachi, K.; Nishino, K.; Mori, N.; Suzuki, T.; Tanuma, S.I.; Abe, R.; Tsukimoti, M. Radiosensitizing effect of P2X7 receptor antagonist on melanoma in vitro and in vivo. Biol. Pharm. Bull. 2017, 40, 878–887. [Google Scholar] [CrossRef]
- Bill, M.A.; Srivastava, K.; Breen, C.; Butterworth, K.T.; McMahon, S.J.; Prise, K.M.; McCloskey, K. Dual effects of radiation bystander signaling in urothelial cancer: Purinergic-activation of apoptosis attenuates survival of urothelial cancer and normal urothelial cells. Oncotarget 2017, 8, 97331–97343. [Google Scholar] [CrossRef]
- Hoorelbeke, D.; Decrock, E.; De Smet, M.; De Bock, M.; Descamps, B.; Van Haver, V.; Delvaeye, T.; Krysko, D.; Vanhove, C.; Bultynck, G.; et al. Cx43 channels and signaling via IP3/Ca(2+), ATP, and ROS/NO propagate radiation-induced DNA damage to non-irradiated brain microvascular endothelial cells. Cell Death Dis. 2020, 11, 194. [Google Scholar] [CrossRef]
- Ohshima, Y.; Kitami, A.; Kawano, A.; Tsukimoto, M.; Kojima, S. Induction of extracellular ATP mediates increase in intracellular thioredoxin in RAW264.7 cells exposed to low-dose gamma-rays. Free Radic. Biol. Med. 2011, 51, 1240–1248. [Google Scholar] [CrossRef]
- Savio, L.E.B.; de Andrade Mello, P.; da Silva, C.G.; Coutinho-Silva, R. The P2X7 receptor in inflammatory diseases: Angel or demon? Front. Pharmacol. 2018, 9, 52. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 receptor in infection and inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Erb, L.; Woods, L.T.; Khalafalla, M.G.; Weisman, G.A. Purinergic signaling in Alzheimer’s disease. Brain Res. Bull. 2019, 151, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.S.; Camden, J.M.; Schrader, A.M.; Redman, R.S.; Turner, J.T. Reversible regulation of P2Y(2) nucleotide receptor expression in the duct-ligated rat submandibular gland. Am. J. Physiol. Cell Physiol. 2000, 279, C286–C294. [Google Scholar] [CrossRef] [PubMed]
- Schrader, A.M.; Camden, J.M.; Weisman, G.A. P2Y2 nucleotide receptor up-regulation in submandibular gland cells from the NOD.B10 mouse model of Sjogren’s syndrome. Arch. Oral Biol. 2005, 50, 533–540. [Google Scholar] [CrossRef]
- Tamaishi, N.; Tsukimoto, M.; Kitami, A.; Kojima, S. P2Y6 receptors and ADAM17 mediate low-dose gamma-ray-induced focus formation (activation) of EGF receptor. Radiat. Res. 2011, 175, 193–200. [Google Scholar] [CrossRef]
- Xu, P.; Xu, Y.; Hu, B.; Wang, J.; Pan, R.; Murugan, M.; Wu, L.; Tang, Y. Extracellular ATP enhances radiation-induced brain injury through microglial activation and paracrine signaling via P2X7 receptor. Brain Behav. Immun. 2015, 50, 87–100. [Google Scholar] [CrossRef]
- Ishibashi, K.; Okamura, K.; Yamazaki, J. Involvement of apical P2Y2 receptor-regulated CFTR activity in muscarinic stimulation of Cl(-) reabsorption in rat submandibular gland. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1729–R1736. [Google Scholar] [CrossRef]
- Nakamoto, T.; Brown, D.A.; Catalan, M.A.; Gonzalez-Begne, M.; Romanenko, V.G.; Melvin, J.E. Purinergic P2X7 receptors mediate ATP-induced saliva secretion by the mouse submandibular gland. J. Biol. Chem. 2009, 284, 4815–4822. [Google Scholar] [CrossRef]
- Khalafalla, M.G.; Woods, L.T.; Jasmer, K.J.; Forti, K.M.; Camden, J.M.; Jensen, J.L.; Limesand, K.H.; Galtung, H.K.; Weisman, G.A. P2 receptors as therapeutic targets in the salivary gland: From physiology to dysfunction. Front. Pharmacol. 2020, 11, 222. [Google Scholar] [CrossRef]
- Ramadan, R.; Vromans, E.; Anang, D.C.; Decrock, E.; Mysara, M.; Monsieurs, P.; Baatout, S.; Leybaert, L.; Aerts, A. Single and fractionated ionizing radiation induce alterations in endothelial connexin expression and channel function. Sci. Rep. 2019, 9, 4643. [Google Scholar]
- Zhang, Q.; Bai, X.; Liu, Y.; Wang, K.; Shen, B.; Sun, X. Current concepts and perspectives on connexin43: A Mini Review. Curr. Protein. Pept. Sci. 2018, 19, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, E.; Wang, N.; Decrock, E.; De Bock, M.; Vinken, M.; Van Moorhem, M.; Lai, C.; Culot, M.; Rogiers, V.; Cecchelli, R.; et al. Ca(2+) regulation of connexin 43 hemichannels in C6 glioma and glial cells. Cell Calcium. 2009, 46, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.; Lagos-Cabre, R.; Kong, M.; Cardenas, A.; Burgos-Bravo, F.; Schneider, P.; Quest, A.; Leyton, L. Integrin-mediated transactivation of P2X7R via hemichannel-dependent ATP release stimulates astrocyte migration. Biochim. Biophys. Acta 2016, 1863, 2175–2188. [Google Scholar] [CrossRef] [PubMed]
- Erb, L.; Weisman, G.A. Coupling of P2Y receptors to G proteins and other signaling pathways. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Decrock, E.; Hoorelbeke, D.; Ramadan, R.; Delvaeye, T.; De Bock, M.; Wang, N.; Krysko, D.; Baatout, S.; Bultynck, G.; Aerts, A.; et al. Calcium, oxidative stress and connexin channels, a harmonious orchestra directing the response to radiotherapy treatment? Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1099–1120. [Google Scholar] [CrossRef]
- Lyng, F.M.; Maguire, P.; McClean, B.; Seymour, C.; Mothersill, C. The involvement of calcium and MAP kinase signaling pathways in the production of radiation-induced bystander effects. Radiat. Res. 2006, 165, 400–409. [Google Scholar]
- Hei, T.K.; Zhou, H.; Chai, Y.; Ponnaiya, B.; Ivanov, V.N. Radiation induced non-targeted response: Mechanism and potential clinical implications. Curr. Mol. Pharmacol. 2011, 4, 96–105. [Google Scholar] [CrossRef]
- Shao, C.; Lyng, F.M.; Folkard, M.; Prise, K.M. Calcium fluxes modulate the radiation-induced bystander responses in targeted glioma and fibroblast cells. Radiat. Res. 2006, 166, 479–487. [Google Scholar] [CrossRef]
- Zhou, H.; Ivanov, V.N.; Lien, Y.C.; Davidson, M.; Hei, T.K. Mitochondrial function and nuclear factor-kappaB-mediated signaling in radiation-induced bystander effects. Cancer Res. 2008, 68, 2233–2240. [Google Scholar] [CrossRef]
- Zhou, H.; Ivanov, V.N.; Gillespie, J.; Geard, C.R.; Amundson, S.A.; Brenner, D.J.; Yu, Z.; Lieberman, H.B.; Hei, T.K. Mechanism of radiation-induced bystander effect: Role of the cyclooxygenase-2 signaling pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 14641–14646. [Google Scholar] [CrossRef]
- Hei, T.K. Cyclooxygenase-2 as a signaling molecule in radiation-induced bystander effect. Mol. Carcinog. 2006, 45, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Zhou, H.; Ivanov, V.N.; Hong, M.; Lieberman, H.B.; Brenner, D.J.; Amundson, S.A.; Geard, C.R. Mechanism of radiation-induced bystander effects: A unifying model. J. Pharm. Pharmacol. 2008, 60, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Asur, R.; Balasubramaniam, M.; Marples, B.; Thomas, R.A.; Tucker, J.D. Involvement of MAPK proteins in bystander effects induced by chemicals and ionizing radiation. Mutat. Res. 2010, 686, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; De Toledo, S.M.; Spitz, D.R.; Little, J.B. Oxidative metabolism modulates signal transduction and micronucleus formation in bystander cells from alpha-particle-irradiated normal human fibroblast cultures. Cancer Res. 2002, 62, 5436–5442. [Google Scholar]
- Ratchford, A.M.; Baker, O.J.; Camden, J.M.; Rikka, S.; Petris, M.J.; Seye, C.I.; Erb, L.; Weisman, G.A. P2Y2 nucleotide receptors mediate metalloprotease-dependent phosphorylation of epidermal growth factor receptor and ErbB3 in human salivary gland cells. J. Biol. Chem. 2010, 285, 7545–7555. [Google Scholar] [CrossRef] [PubMed]
- Hedden, L.; Benes, C.H.; Soltoff, S.P. P2X(7) receptor antagonists display agonist-like effects on cell signaling proteins. Biochim. Biophys. Acta 2011, 1810, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Calaf, G.M.; Zhou, H.; Ghandhi, S.A.; Elliston, C.D.; Wen, G.; Nohmi, T.; Amundson, S.A.; Hei, T.K. Radiation induced COX-2 expression and mutagenesis at non-targeted lung tissues of gpt delta transgenic mice. Br. J. Cancer 2013, 108, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Nishimaki, N.; Tsukimoto, M.; Kitami, A.; Kojima, S. Autocrine regulation of gamma-irradiation-induced DNA damage response via extracellular nucleotides-mediated activation of P2Y6 and P2Y12 receptors. DNA Repair 2012, 11, 657–665. [Google Scholar] [CrossRef]
- Han, W.; Wu, L.; Chen, S.; Bao, L.; Zhang, L.; Jiang, E.; Zhao, Y.; Xu, A.; Hei, T.K.; Yu, Z. Constitutive nitric oxide acting as a possible intercellular signaling molecule in the initiation of radiation-induced DNA double strand breaks in non-irradiated bystander cells. Oncogene 2007, 26, 2330–2339. [Google Scholar] [CrossRef]
- Shao, C.; Folkard, M.; Prise, K.M. Role of TGF-beta1 and nitric oxide in the bystander response of irradiated glioma cells. Oncogene 2008, 27, 434–440. [Google Scholar] [CrossRef]
- Khalafalla, M.G.; Woods, L.T.; Camden, J.M.; Khan, A.A.; Limesand, K.H.; Petris, M.J.; Erb, L.; Weisman, G.A. P2X7 receptor antagonism prevents IL-1beta release from salivary epithelial cells and reduces inflammation in a mouse model of autoimmune exocrinopathy. J. Biol. Chem. 2017, 292, 16626–16637. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.M.; Desai, L.P. Reciprocal regulation of TGF-beta and reactive oxygen species: A perverse cycle for fibrosis. Redox Biol. 2015, 6, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Murphy-Ullrich, J.E.; Poczatek, M. Activation of latent TGF-beta by thrombospondin-1: Mechanisms and physiology. Cytokine Growth Factor Rev. 2000, 11, 59–69. [Google Scholar] [CrossRef]
- Yu, Q.; Stamenkovic, I. Cell surface-localized matrix metalloproteinase-9 proteolytically activates TGF-beta and promotes tumor invasion and angiogenesis. Genes Dev. 2000, 14, 163–176. [Google Scholar] [PubMed]
- Munger, J.S.; Huang, X.; Kawakatsu, H.; Griffiths, M.J.; Dalton, S.L.; Wu, J.; Pittet, J.F.; Kaminski, N.; Garat, C.; Matthay, M.A.; et al. The integrin alpha v beta 6 binds and activates latent TGF beta 1: A mechanism for regulating pulmonary inflammation and fibrosis. Cell 1999, 96, 319–328. [Google Scholar] [CrossRef]
- Wipff, P.J.; Rifkin, D.B.; Meister, J.J.; Hinz, B. Myofibroblast contraction activates latent TGF-beta1 from the extracellular matrix. J. Cell. Biol. 2007, 179, 1311–1323. [Google Scholar] [CrossRef]
- Erb, L.; Liu, J.; Ockerhausen, J.; Kong, Q.; Garrad, R.C.; Griffin, K.; Neal, C.; Krugh, B.; Santiago-Pérez, L.I.; González, F.A.; et al. An RGD sequence in the P2Y(2) receptor interacts with alphaVbeta3 integrins and is required for G(o)-mediated signal transduction. J. Cell. Biol. 2001, 153, 491–501. [Google Scholar] [CrossRef]
- Camden, J.M.; Schrader, A.M.; Camden, R.E.; Gonzalez, F.A.; Erb, L.; Seye, C.I.; Weisman, G.A. P2Y2 nucleotide receptors enhance alpha-secretase-dependent amyloid precursor protein processing. J. Biol. Chem. 2005, 280, 18696–18702. [Google Scholar] [CrossRef]
- Dewan, M.Z.; Galloway, A.E.; Kawashima, N.; Dewyngaert, J.K.; Babb, J.S.; Formenti, S.C.; Demaria, S. Fractionated but not single-dose radiotherapy induces an immune-mediated abscopal effect when combined with anti-CTLA-4 antibody. Clin. Cancer Res. 2009, 15, 5379–5388. [Google Scholar] [CrossRef]
- Demaria, S.; Ng, B.; Devitt, M.L.; Babb, J.S.; Kawashima, N.; Liebes, L.; Formenti, S.C. Ionizing radiation inhibition of distant untreated tumors (abscopal effect) is immune mediated. Int. J. Radiat. Oncol. Biol. Phys. 2004, 58, 862–870. [Google Scholar] [CrossRef]
- Rodriguez-Ruiz, M.E.; Vanpouille-Box, C.; Melero, I.; Formenti, S.C.; Demaria, S. Immunological mechanisms responsible for radiation-induced abscopal effect. Trends Immunol. 2018, 39, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Coates, P.J.; Robinson, J.I.; Lorimore, S.A.; Wright, E.G. Ongoing activation of p53 pathway responses is a long-term consequence of radiation exposure in vivo and associates with altered macrophage activities. J. Pathol. 2008, 214, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Elliott, M.R.; Chekeni, F.B.; Trampont, P.C.; Lazarowski, E.R.; Kadl, A.; Walk, S.F.; Park, D.; Woodson, R.I.; Ostankovich, M.; Sharma, P.; et al. Nucleotides released by apoptotic cells act as a find-me signal to promote phagocytic clearance. Nature 2009, 461, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Eun, S.Y.; Park, S.W.; Lee, J.H.; Chang, K.C.; Kim, H.J. P2Y2R activation by nucleotides released from oxLDL-treated endothelial cells (ECs) mediates the interaction between ECs and immune cells through RAGE expression and reactive oxygen species production. Free Radic. Biol. Med. 2014, 69, 157–166. [Google Scholar] [CrossRef]
- Chen, Y.; Corriden, R.; Inoue, Y.; Yip, L.; Hashiguchi, N.; Zinkernagel, A.; Nizet, V.; Insel, P.; Junger, W. ATP release guides neutrophil chemotaxis via P2Y2 and A3 receptors. Science 2006, 314, 1792–1795. [Google Scholar] [CrossRef]
- Lacas, B.; Bourhis, J.; Overgaard, J.; Zhang, Q.; Grégoire, V.; Nankivell, M.; Zackrisson, B.; Szutkowski, Z.; Suwiński, R.; Poulsen, M.; et al. Role of radiotherapy fractionation in head and neck cancers (MARCH): An updated meta-analysis. Lancet Oncol. 2017, 18, 1221–1237. [Google Scholar] [CrossRef]
- Taylor, A.; Powell, M.E.B. Intensity-modulated radiotherapy—What is it? Cancer Imaging 2004, 4, 68–73. [Google Scholar] [CrossRef]
- Deasy, J.O.; Moiseenko, V.; Marks, L.; Chao, K.S.; Nam, J.; Eisbruch, A. Radiotherapy dose-volume effects on salivary gland function. Int. J. Radiat. Oncol. Biol. Phys. 2010, 76 (Suppl. 3), S58–S63. [Google Scholar] [CrossRef]
- Eisbruch, A.; Ten Haken, R.K.; Kim, H.M.; Marsh, L.H.; Ship, J.A. Dose, volume, and function relationships in parotid salivary glands following conformal and intensity-modulated irradiation of head and neck cancer. Int. J. Radiat. Oncol. Biol. Phys. 1999, 45, 577–587. [Google Scholar] [CrossRef]
- Murdoch-Kinch, C.A.; Kim, H.M.; Vineberg, K.A.; Ship, J.A.; Eisbruch, A. Dose-effect relationships for the submandibular salivary glands and implications for their sparing by intensity modulated radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2008, 72, 373–382. [Google Scholar] [CrossRef]
- Ghosh, G.; Gupta, G.; Malviya, A.; Saroj, D. Comparison three-dimensional conformal radiotherapy versus intensity modulated radiation therapy in local control of head and neck cancer. J. Cancer Res. Ther. 2018, 14, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Alterio, D.; Gugliandolo, S.G.; Augugliaro, M.; Marvaso, G.; Gandini, S.; Bellerba, F.; Russell-Edu, S.W.; Simone, I.D.; Cinquini, M.; Starzyńska, A.; et al. IMRT vs 2D/3D conformal RT in oropharyngeal cancer: A review of the literature and meta-analysis. Oral Dis. 2020. [Google Scholar] [CrossRef]
- Nutting, C.M.; Morden, J.P.; Harrington, K.J.; Urbano, T.G.; Bhide, S.A.; Clark, C.; Miles, E.A.; Miah, A.B.; Newbold, K.; Tanay, M.; et al. Parotid-sparing intensity modulated versus conventional radiotherapy in head and neck cancer (PARSPORT): A phase 3 multicentre randomised controlled trial. Lancet Oncol. 2011, 12, 127–136. [Google Scholar] [CrossRef]
- Lan, X.; Chan, J.Y.K.; Pu, J.J.; Qiao, W.; Pang, S.; Yang, W.F.; Wong, K.C.W.; Kwong, D.L.W.; Su, Y.X. Saliva electrolyte analysis and xerostomia-related quality of life in nasopharyngeal carcinoma patients following intensity-modulated radiation therapy. Radiother. Oncol. 2020, 150, 97–103. [Google Scholar] [CrossRef]
- Teoh, M.; Clark, C.H.; Wood, K.; Whitaker, S.; Nisbet, A. Volumetric modulated arc therapy: A review of current literature and clinical use in practice. Br. J. Radiol. 2011, 84, 967–996. [Google Scholar] [CrossRef] [PubMed]
- Mashhour, K.; Kamaleldin, M.; Hashem, W. RapidArc vs conventional IMRT for head and neck cancer irradiation: Is faster necessarily better? Asian Pac. J. Cancer Prev. 2018, 19, 207–211. [Google Scholar] [PubMed]
- Nagarajan, M.; Banu, R.; Sathya, B.; Sundaram, T.; Chellapandian, T.P. Dosimetric evaluation and comparison between volumetric modulated arc therapy (VMAT) and intensity modulated radiation therapy (IMRT) plan in head and neck cancers. Gulf J. Oncolog. 2020, 1, 45–50. [Google Scholar]
- Delaney, A.R.; Dahele, M.; Slotman, B.J.; Verbakel, W. Is accurate contouring of salivary and swallowing structures necessary to spare them in head and neck VMAT plans? Radiother. Oncol. 2018, 127, 190–196. [Google Scholar] [CrossRef]
- Ouyang, Z.; Liu Shen, Z.; Murray, E.; Kolar, M.; LaHurd, D.; Yu, N.; Joshi, N.; Koyfman, S.; Bzdusek, K.; Xia, P. Evaluation of auto-planning in IMRT and VMAT for head and neck cancer. J. Appl. Clin. Med. Phys. 2019, 20, 39–47. [Google Scholar] [CrossRef]
- Holliday, E.B.; Frank, S.J. Proton radiation therapy for head and neck cancer: A review of the clinical experience to date. Int. J. Radiat. Oncol. Biol. Phys. 2014, 89, 292–302. [Google Scholar] [CrossRef]
- Xiang, M.; Chang, D.T.; Pollom, E.L. Second cancer risk after primary cancer treatment with three-dimensional conformal, intensity-modulated, or proton beam radiation therapy. Cancer 2020, 126, 3560–3568. [Google Scholar] [CrossRef] [PubMed]
- Lukens, J.N.; Lin, A.; Hahn, S.M. Proton therapy for head and neck cancer. Curr. Opin. Oncol. 2015, 27, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Wang, P.; Gong, L.; Xu, L.; Zhang, J.; Zheng, J.; Zhang, D.; Tian, T.; Wang, P. Construction and clinical evaluation of a new customized bite block used in radiotherapy of head and neck cancer. Cancer Radiother. 2019, 23, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Stieb, S.; Perez-Martinez, I.; Mohamed, A.S.R.; Rock, S.; Bajaj, N.; Deshpande, T.S.; Zaid, M.; Garden, A.S.; Goepfert, R.P.; Cardoso, R.; et al. The impact of tongue-deviating and tongue-depressing oral stents on long-term radiation-associated symptoms in oropharyngeal cancer survivors. Clin. Transl. Radiat. Oncol. 2020, 24, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Verrone, J.R.; Alves, F.A.; Prado, J.D.; Marcicano, A.; de Assis Pellizzon, A.C.; Damascena, A.S.; Jaguar, G.C. Benefits of an intraoral stent in decreasing the irradiation dose to oral healthy tissue: Dosimetric and clinical features. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2014, 118, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Appendino, P.; Della Ferrera, F.; Nassisi, D.; Blandino, G.; Gino, E.; Solla, S.D.; Redda, M.G.R. Are intraoral customized stents still necessary in the era of highly conformal radiotherapy for head & neck cancer? Case series and literature review. Rep. Pract. Oncol. Radiother. 2019, 24, 491–498. [Google Scholar]
- Jha, N.; Seikaly, H.; Harris, J.; Williams, D.; Liu, R.; McGaw, T.; Hofmann, H.; Robinson, D.; Hanson, J.; Barnaby, P. Prevention of radiation induced xerostomia by surgical transfer of submandibular salivary gland into the submental space. Radiother. Oncol. 2003, 66, 283–289. [Google Scholar] [CrossRef]
- Pathak, K.A.; Bhalavat, R.L.; Mistry, R.C.; Deshpande, M.S.; Bhalla, V.; Desai, S.B.; Malpini, B.L. Upfront submandibular salivary gland transfer in pharyngeal cancers. Oral Oncol. 2004, 40, 960–963. [Google Scholar] [CrossRef]
- Valstar, M.H.; de Bakker, B.S.; Steenbakkers, R.; de Jong, K.H.; Smit, L.A.; Klein Nulent, T.J.W.; van Es, R.J.J.; Hofland, I.; de Keizer, B.; Jasperse, B.; et al. The tubarial salivary glands: A potential new organ at risk for radiotherapy. Radiother. Oncol. 2020. [Google Scholar] [CrossRef]
- Troyer, J.K.; Beckett, M.L.; Wright, G.L., Jr. Detection and characterization of the prostate-specific membrane antigen (PSMA) in tissue extracts and body fluids. Int. J. Cancer 1995, 62, 552–558. [Google Scholar]
- Harkema, J.R.C.S.; Wagner, J.G.; Dintzis, S.M.; Liggitt, D. Nose, Sinus, Pharynx, and Larynx. In Comparative Anatomy and Histology; Elsevier: London, UK, 2018. [Google Scholar]
- Leung, W.S.; Wu, V.W.C.; Liu, C.Y.W.; Cheng, A.C.K. A dosimetric comparison of the use of equally spaced beam (ESB), beam angle optimization (BAO), and volumetric modulated arc therapy (VMAT) in head and neck cancers treated by intensity modulated radiotherapy. J. Appl. Clin. Med. Phys. 2019, 20, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Roesink, J.M.; Moerland, M.A.; Hoekstra, A.; Van Rijk, P.P.; Terhaard, C.H. Scintigraphic assessment of early and late parotid gland function after radiotherapy for head-and-neck cancer: A prospective study of dose-volume response relationships. Int. J. Radiat. Oncol. Biol. Phys. 2004, 58, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Scarantino, C.; LeVeque, F.; Swann, R.S.; White, R.; Schulsinger, A.; Hodson, D.I.; Meredith, R.; Foote, R.; Brachman, D.; Lee, N. Effect of pilocarpine during radiation therapy: Results of RTOG 97-09, a phase III randomized study in head and neck cancer patients. J. Support Oncol. 2006, 4, 252–258. [Google Scholar] [PubMed]
- Rieke, J.W.; Hafermann, M.D.; Johnson, J.T.; LeVeque, F.G.; Iwamoto, R.; Steiger, B.W.; Muscoplat, C.; Gallagher, S.C. Oral pilocarpine for radiation-induced xerostomia: Integrated efficacy and safety results from two prospective randomized clinical trials. Int. J. Radiat. Oncol. Biol. Phys. 1995, 31, 661–669. [Google Scholar] [CrossRef]
- Zimmerman, R.P.; Mark, R.J.; Tran, L.M.; Juillard, G.F. Concomitant pilocarpine during head and neck irradiation is associated with decreased posttreatment xerostomia. Int. J. Radiat. Oncol. Biol. Phys. 1997, 37, 571–575. [Google Scholar] [CrossRef]
- Nyarady, Z.; Nemeth, A.; Ban, A.; Mukics, A.; Nyarady, J.; Ember, I.; Olasz, L. A randomized study to assess the effectiveness of orally administered pilocarpine during and after radiotherapy of head and neck cancer. Anticancer Res. 2006, 26, 1557–1562. [Google Scholar]
- Chambers, M.S.; Posner, M.; Jones, C.U.; Biel, M.A.; Hodge, K.M.; Vitti, R.; Armstrong, I.; Yen, C.; Weber, R.S. Cevimeline for the treatment of postirradiation xerostomia in patients with head and neck cancer. Int. J. Radiat. Oncol. Biol. Phys. 2007, 68, 1102–1109. [Google Scholar] [CrossRef]
- Chambers, M.S.; Jones, C.U.; Biel, M.A.; Weber, R.S.; Hodge, K.M.; Chen, Y.; Holland, J.M.; Ship, J.A.; Vitti, R.; Armstrong, I.; et al. Open-label, long-term safety study of cevimeline in the treatment of postirradiation xerostomia. Int. J. Radiat. Oncol. Biol. Phys. 2007, 69, 1369–1376. [Google Scholar] [CrossRef]
- Masunaga, H.; Ogawa, H.; Uematsu, Y.; Tomizuka, T.; Yasuda, H.; Takeshita, Y. Long-lasting salivation induced by a novel muscarinic receptor agonist SNI-2011 in rats and dogs. Eur. J. Pharmacol. 1997, 339, 1–9. [Google Scholar] [CrossRef]
- Epstein, J.B.; Burchell, J.L.; Emerton, S.; Le, N.D.; Silverman, S., Jr. A clinical trial of bethanechol in patients with xerostomia after radiation therapy. A pilot study. Oral Surg. Oral Med. Oral Pathol. 1994, 77, 610–614. [Google Scholar] [CrossRef]
- Gorsky, M.; Epstein, J.B.; Parry, J.; Epstein, M.S.; Le, N.D.; Silverman, S., Jr. The efficacy of pilocarpine and bethanechol upon saliva production in cancer patients with hyposalivation following radiation therapy. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2004, 97, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Jham, B.C.; Teixeira, I.V.; Aboud, C.G.; Carvalho, A.L.; Coelho Mde, M.; Freire, A.R. A randomized phase III prospective trial of bethanechol to prevent radiotherapy-induced salivary gland damage in patients with head and neck cancer. Oral Oncol. 2007, 43, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Jaguar, G.C.; Lima, E.N.; Kowalski, L.P.; Pellizzon, A.C.; Carvalho, A.L.; Boccaletti, K.W.; Alves, F.A. Double blind randomized prospective trial of bethanechol in the prevention of radiation-induced salivary gland dysfunction in head and neck cancer patients. Radiother. Oncol. 2015, 115, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Riley, P.; Glenny, A.M.; Hua, F.; Worthington, H.V. Pharmacological interventions for preventing dry mouth and salivary gland dysfunction following radiotherapy. Cochrane Database Syst. Rev. 2017, 7, CD012744. [Google Scholar] [PubMed]
- Kouvaris, J.R.; Kouloulias, V.E.; Vlahos, L.J. Amifostine: The first selective-target and broad-spectrum radioprotector. Oncologist 2007, 12, 738–747. [Google Scholar] [CrossRef]
- Wasserman, T.H.; Brizel, D.M.; Henke, M.; Monnier, A.; Eschwege, F.; Sauer, R.; Strnad, V. Influence of intravenous amifostine on xerostomia, tumor control, and survival after radiotherapy for head-and- neck cancer: 2-year follow-up of a prospective, randomized, phase III trial. Int. J. Radiat. Oncol. Biol. Phys. 2005, 63, 985–990. [Google Scholar] [CrossRef]
- Bourhis, J.; Blanchard, P.; Maillard, E.; Brizel, D.M.; Movsas, B.; Buentzel, J.; Langendijk, J.A.; Komaki, R.; Leong, S.S.; Levendag, P.; et al. Effect of amifostine on survival among patients treated with radiotherapy: A meta-analysis of individual patient data. J. Clin. Oncol. 2011, 29, 2590–2597. [Google Scholar] [CrossRef]
- Lee, M.G.; Freeman, A.R.; Roos, D.E.; Milner, A.D.; Borg, M.F. Randomized double-blind trial of amifostine versus placebo for radiation-induced xerostomia in patients with head and neck cancer. J. Med. Imaging Radiat. Oncol. 2019, 63, 142–150. [Google Scholar] [CrossRef]
- Rades, D.; Fehlauer, F.; Bajrovic, A.; Mahlmann, B.; Richter, E.; Alberti, W. Serious adverse effects of amifostine during radiotherapy in head and neck cancer patients. Radiother. Oncol. 2004, 70, 261–264. [Google Scholar] [CrossRef]
- Singh, V.K.; Seed, T.M. The efficacy and safety of amifostine for the acute radiation syndrome. Expert Opin. Drug Saf. 2019, 18, 1077–1090. [Google Scholar] [CrossRef]
- Grdina, D.J.; Kataoka, Y.; Murley, J.S. Amifostine: Mechanisms of action underlying cytoprotection and chemoprevention. Drug Metabol. Drug Interact. 2000, 16, 237–279. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Pang, B.; Iglesias-Bartolome, R.; Wu, X.; Hu, L.; Zhang, C.; Wang, J.; Gutkind, J.S.; Wang, S. Prevention of irradiation-induced salivary hypofunction by rapamycin in swine parotid glands. Oncotarget 2016, 7, 20271–20281. [Google Scholar] [CrossRef] [PubMed]
- Hai, B.; Qin, L.; Yang, Z.; Zhao, Q.; Shangguan, L.; Ti, X.; Zhao, Y.; Kim, S.; Rangaraj, D.; Liu, F. Transient activation of hedgehog pathway rescued irradiation-induced hyposalivation by preserving salivary stem/progenitor cells and parasympathetic innervation. Clin. Cancer Res. 2014, 20, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Lister, M.F.; Sharkey, J.; Sawatzky, D.A.; Hodgkiss, J.P.; Davidson, D.J.; Rossi, A.G.; Finlayson, K. The role of the purinergic P2X7 receptor in inflammation. J. Inflamm. 2007, 4, 5. [Google Scholar] [CrossRef]
- Di Virgilio, F. Liaisons dangereuses: P2X(7) and the inflammasome. Trends Pharmacol. Sci. 2007, 28, 465–472. [Google Scholar] [CrossRef]
- Woods, L.T.; Camden, J.M.; Batek, J.M.; Petris, M.J.; Erb, L.; Weisman, G.A. P2X7 receptor activation induces inflammatory responses in salivary gland epithelium. Am. J. Physiol. Cell Physiol. 2012, 303, C790–C801. [Google Scholar] [CrossRef]
- DeVries, T.A.; Neville, M.C.; Reyland, M.E. Nuclear import of PKCdelta is required for apoptosis: Identification of a novel nuclear import sequence. EMBO J. 2002, 21, 6050–6060. [Google Scholar] [CrossRef]
- Leitges, M.; Mayr, M.; Braun, U.; Mayr, U.; Li, C.; Pfister, G.; Ghaffari-Tabrizi, N.; Baier, G.; Hu, Y.; Xu, Q. Exacerbated vein graft arteriosclerosis in protein kinase Cdelta-null mice. J. Clin. Investig. 2001, 108, 1505–1512. [Google Scholar] [CrossRef]
- Allen-Petersen, B.L.; Miller, M.R.; Neville, M.C.; Anderson, S.M.; Nakayama, K.I.; Reyland, M.E. Loss of protein kinase C delta alters mammary gland development and apoptosis. Cell Death Dis. 2010, 1, e17. [Google Scholar] [CrossRef][Green Version]
- Li, J.; Shan, Z.; Ou, G.; Liu, X.; Zhang, C.; Baum, B.J.; Wang, S. Structural and functional characteristics of irradiation damage to parotid glands in the miniature pig. Int. J. Radiat. Oncol. Biol. Phys. 2005, 62, 1510–1516. [Google Scholar] [CrossRef]
- Lombaert, I.; Movahednia, M.M.; Adine, C.; Ferreira, J.N. Concise Review: Salivary gland regeneration: Therapeutic approaches from stem cells to tissue organoids. Stem Cells 2017, 35, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Baum, B.J.; Zheng, C.; Cotrim, A.P.; McCullagh, L.; Goldsmith, C.M.; Brahim, J.S.; Atkinson, J.C.; Turner, R.J.; Liu, S.; Nikolov, N.; et al. Aquaporin-1 gene transfer to correct radiation-induced salivary hypofunction. Handb. Exp. Pharmacol. 2009, 190, 403–418. [Google Scholar]
- Mobasheri, A.; Marples, D. Expression of the AQP-1 water channel in normal human tissues: A semiquantitative study using tissue microarray technology. Am. J. Physiol. Cell Physiol. 2004, 286, C529–C537. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hart, P.S.; Piesco, N.P.; Lu, X.; Gorry, M.C.; Hart, T.C. Aquaporin expression in developing human teeth and selected orofacial tissues. Calcif. Tissue Int. 2003, 72, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Gresz, V.; Kwon, T.H.; Hurley, P.T.; Varga, G.; Zelles, T.; Nielsen, S.; Case, R.M.; Steward, M.C. Identification and localization of aquaporin water channels in human salivary glands. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G247–G254. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Saga, T.; Watanabe, K.; Takahashi, N.; Tabira, Y.; Kusukawa, J.; Yamaki, K.I. An immunohistochemistry-based study on aquaporin (AQP)-1, 3, 4, 5 and 8 in the parotid glands, submandibular glands and sublingual glands of Sjogren’s syndrome mouse models chronically administered cevimeline. Kurume Med. J. 2013, 60, 7–19. [Google Scholar] [CrossRef]
- Akamatsu, T.; Parvin, M.N.; Murdiastuti, K.; Kosugi-Tanaka, C.; Yao, C.; Miki, O.; Kanamori, N.; Hosoi, K. Expression and localization of aquaporins, members of the water channel family, during development of the rat submandibular gland. Pflugers Arch. 2003, 446, 641–651. [Google Scholar] [CrossRef]
- Gao, R.; Yan, X.; Zheng, C.; Goldsmith, C.M.; Afione, S.; Hai, B.; Xu, J.; Zhou, J.; Chiorini, J.A.; Baum, B.J.; et al. AAV2-mediated transfer of the human aquaporin-1 cDNA restores fluid secretion from irradiated miniature pig parotid glands. Gene Ther. 2011, 18, 38–42. [Google Scholar] [CrossRef]
- Alevizos, I.; Zheng, C.; Cotrim, A.P.; Liu, S.; McCullagh, L.; Billings, M.E.; Goldsmith, C.M.; Tandon, M.; Helmerhorst, E.J.; Catalan, M.A.; et al. Late responses to adenoviral-mediated transfer of the aquaporin-1 gene for radiation-induced salivary hypofunction. Gene Ther. 2017, 24, 176–186. [Google Scholar] [CrossRef]
- Baum, B.J.; Alevizos, I.; Zheng, C.; Cotrim, A.P.; Liu, S.; McCullagh, L.; Goldsmith, C.M.; Burbelo, P.D.; Citrin, D.E.; Mitchell, J.B.; et al. Early responses to adenoviral-mediated transfer of the aquaporin-1 cDNA for radiation-induced salivary hypofunction. Proc. Natl. Acad. Sci. USA 2012, 109, 19403–19407. [Google Scholar] [CrossRef]
- Wang, Z.; Pradhan-Bhatt, S.; Farach-Carson, M.C.; Passineau, M.J. Artificial induction of native aquaporin-1 expression in human salivary cells. J. Dent. Res. 2017, 96, 444–449. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Wang, S.; Zhang, L.R.; Zhang, N.; Cheng, Z.; Liu, Q.; Shields, K.J.; Hu, B.; Passineau, M.J. CRISPR-Cas9 HDR system enhances AQP1 gene expression. Oncotarget 2017, 8, 111683–111696. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Lin, Y.; Cao, H.; Sirjani, D.; Giaccia, A.J.; Koong, A.C.; Kong, C.S.; Diehn, M.; Le, Q.T. Neurotrophic factor GDNF promotes survival of salivary stem cells. J. Clin. Investig. 2014, 124, 3364–3377. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, L.S.; Baanstra, M.; Faber, H.; Rocchi, C.; Zwart, E.; de Haan, G.; van Os, R.; Coppes, R.P. Purification and ex vivo expansion of fully functional salivary gland stem cells. Stem Cell Rep. 2014, 3, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; van der Zwaag, M.; Stokman, M.A.; van Os, R.; Coppes, R.P. Isolation and characterization of human salivary gland cells for stem cell transplantation to reduce radiation-induced hyposalivation. Radiother. Oncol. 2009, 92, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Pringle, S.; Maimets, M.; van der Zwaag, M.; Stokman, M.A.; van Gosliga, D.; Zwart, E.; Witjes, M.J.H.; de Haan, G.; van Os, R.; Coppes, R.P. Human salivary gland stem cells functionally restore radiation damaged salivary glands. Stem Cells 2016, 34, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, T.; Fowler, E.W.; Hao, Y.; Ravikrishnan, A.; Harrington, D.A.; Witt, R.L.; Farach-Carson, M.C.; Pradhan-Bhatt, S.; Jia, X. Biomaterials-based strategies for salivary gland tissue regeneration. Biomater Sci. 2016, 4, 592–604. [Google Scholar] [CrossRef]
- Nam, K.; Kim, K.; Dean, S.M.; Brown, C.T.; Davis, R.S.; Okano, T.; Baker, O.J. Using cell sheets to regenerate mouse submandibular glands. NPJ Regen. Med. 2019, 4, 16. [Google Scholar] [CrossRef]
- Haara, O.; Fujimori, S.; Schmidt-Ullrich, R.; Hartmann, C.; Thesleff, I.; Mikkola, M.L. Ectodysplasin and Wnt pathways are required for salivary gland branching morphogenesis. Development 2011, 138, 2681–2691. [Google Scholar] [CrossRef]
- Fiaschi, M.; Kolterud, A.; Nilsson, M.; Toftgard, R.; Rozell, B. Targeted expression of GLI1 in the salivary glands results in an altered differentiation program and hyperplasia. Am. J. Pathol. 2011, 179, 2569–2579. [Google Scholar] [CrossRef]
- Hai, B.; Yang, Z.; Millar, S.E.; Choi, Y.S.; Taketo, M.M.; Nagy, A.; Liu, F. Wnt/beta-catenin signaling regulates postnatal development and regeneration of the salivary gland. Stem Cells Dev. 2010, 19, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Gera, J.; Lichtenstein, A. The mammalian target of rapamycin pathway as a therapeutic target in multiple myeloma. Leuk. Lymphoma 2011, 52, 1857–1866. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasmer, K.J.; Gilman, K.E.; Muñoz Forti, K.; Weisman, G.A.; Limesand, K.H. Radiation-Induced Salivary Gland Dysfunction: Mechanisms, Therapeutics and Future Directions. J. Clin. Med. 2020, 9, 4095. https://doi.org/10.3390/jcm9124095
Jasmer KJ, Gilman KE, Muñoz Forti K, Weisman GA, Limesand KH. Radiation-Induced Salivary Gland Dysfunction: Mechanisms, Therapeutics and Future Directions. Journal of Clinical Medicine. 2020; 9(12):4095. https://doi.org/10.3390/jcm9124095
Chicago/Turabian StyleJasmer, Kimberly J., Kristy E. Gilman, Kevin Muñoz Forti, Gary A. Weisman, and Kirsten H. Limesand. 2020. "Radiation-Induced Salivary Gland Dysfunction: Mechanisms, Therapeutics and Future Directions" Journal of Clinical Medicine 9, no. 12: 4095. https://doi.org/10.3390/jcm9124095
APA StyleJasmer, K. J., Gilman, K. E., Muñoz Forti, K., Weisman, G. A., & Limesand, K. H. (2020). Radiation-Induced Salivary Gland Dysfunction: Mechanisms, Therapeutics and Future Directions. Journal of Clinical Medicine, 9(12), 4095. https://doi.org/10.3390/jcm9124095