Mineralocorticoid Receptor May Regulate Glucose Homeostasis through the Induction of Interleukin-6 and Glucagon-Like peptide-1 in Pancreatic Islets

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experimental Section
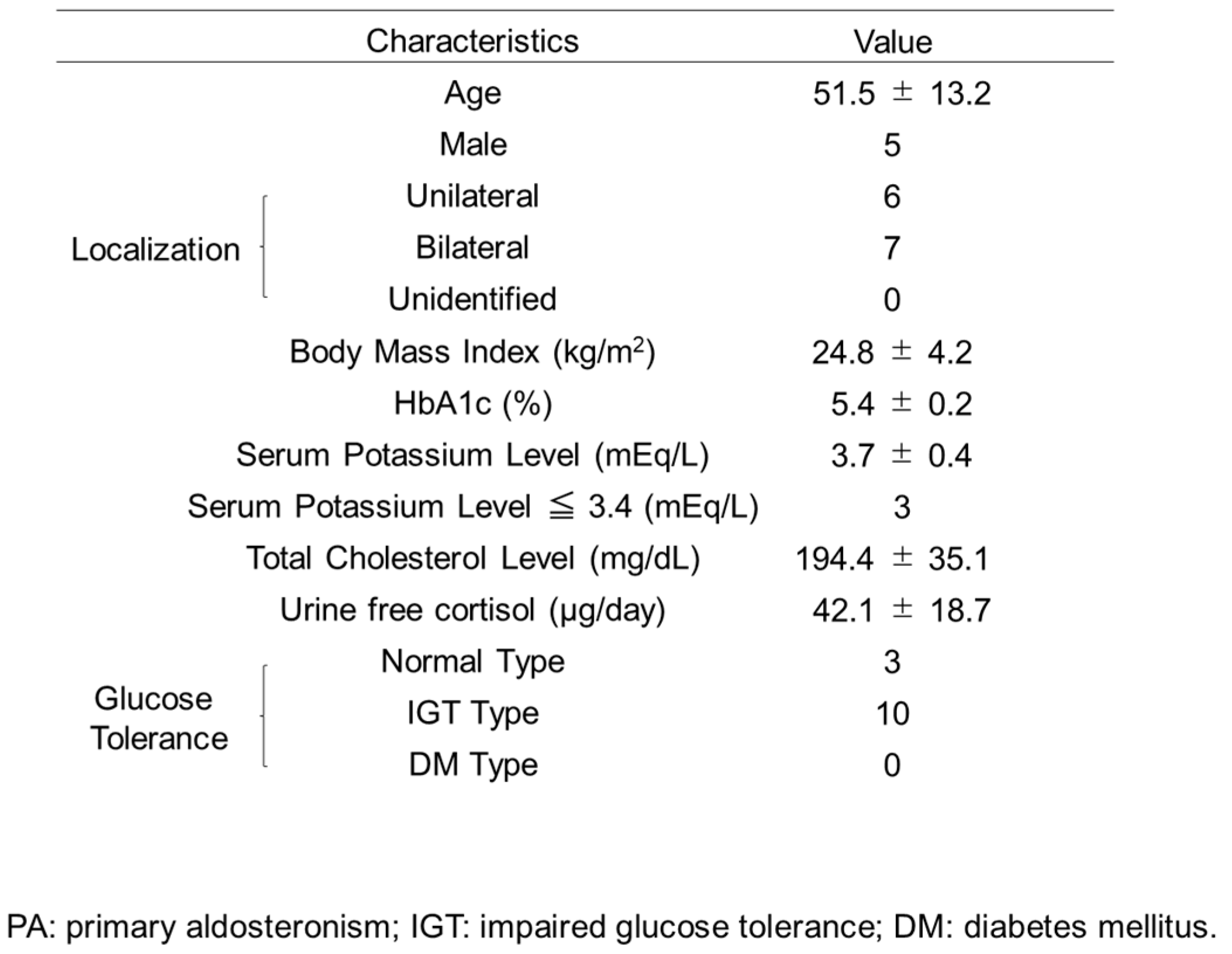
2.1. Materials Establishment of PA Model Rodents, Treatment Groups and Histological Analysis of Pancreatic Islets
2.2. Cell Lines and Treatments
2.3. Glucose Homeostasis of PA Patients before and after Treatment
2.4. ELISA Assays
2.5. RT-PCR
2.6. siRNA Transfection
2.7. Reporter Plasmids Construction and Luciferase Assay
2.8. Statistical Analysis
3. Results
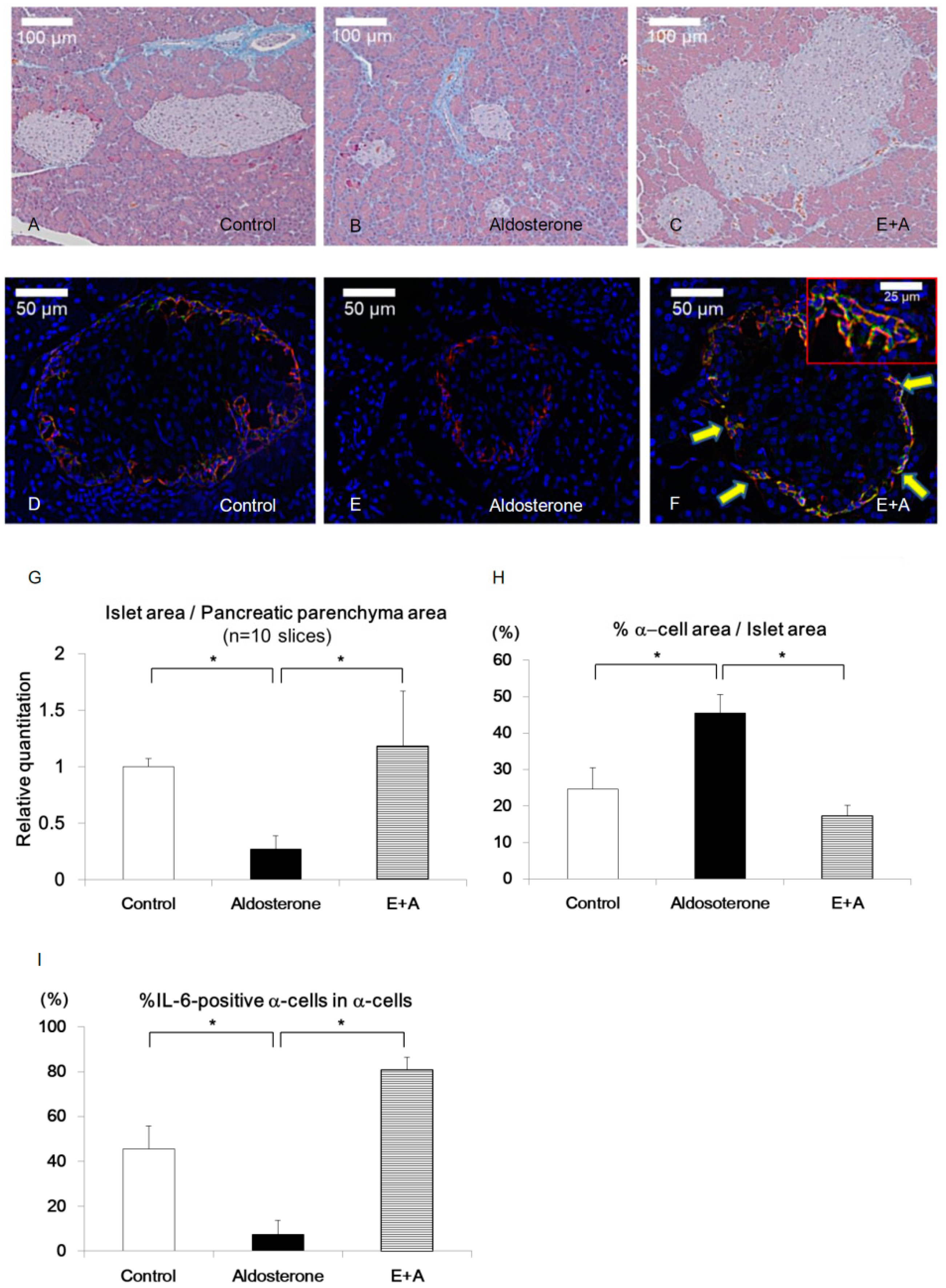
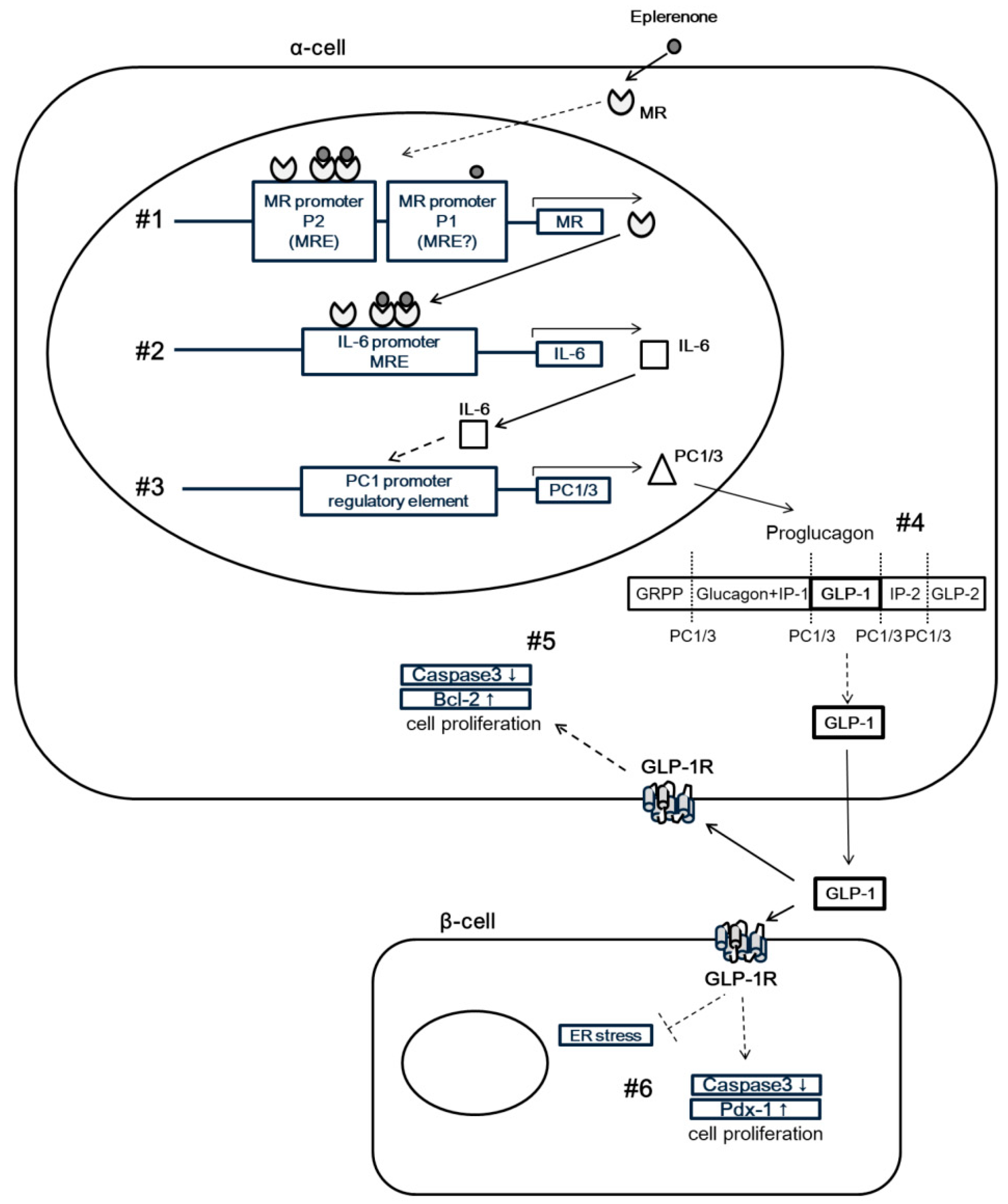
3.1. Eplerenone Treatment Protects Islet Cells from Cell Damage Caused by Aldosterone
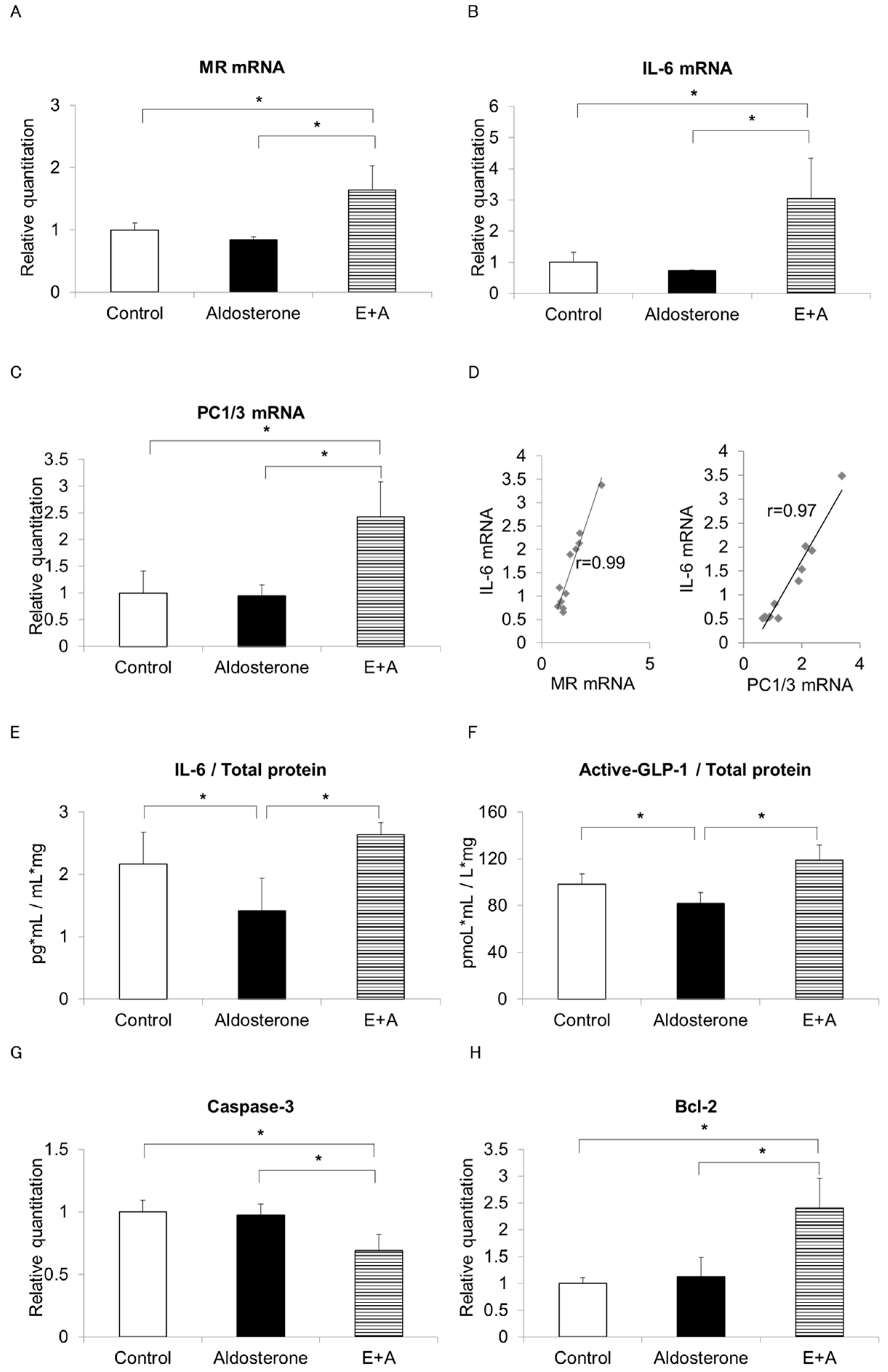
3.2. Eplerenone Treatment Increases MR, IL-6 and Active GLP-1 Secretion from α-Cells
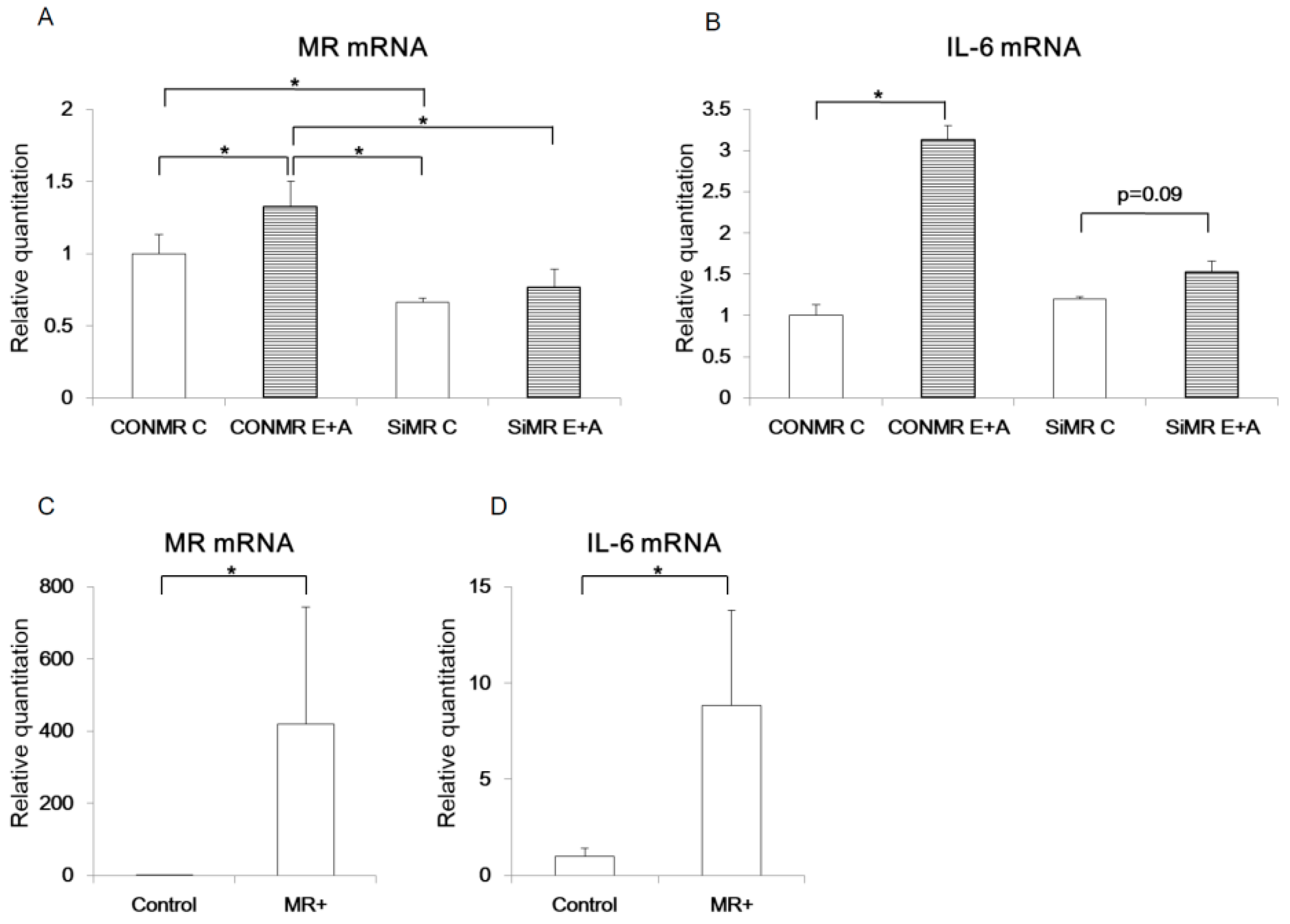
3.3. MR regulates IL-6 expression in α-cells
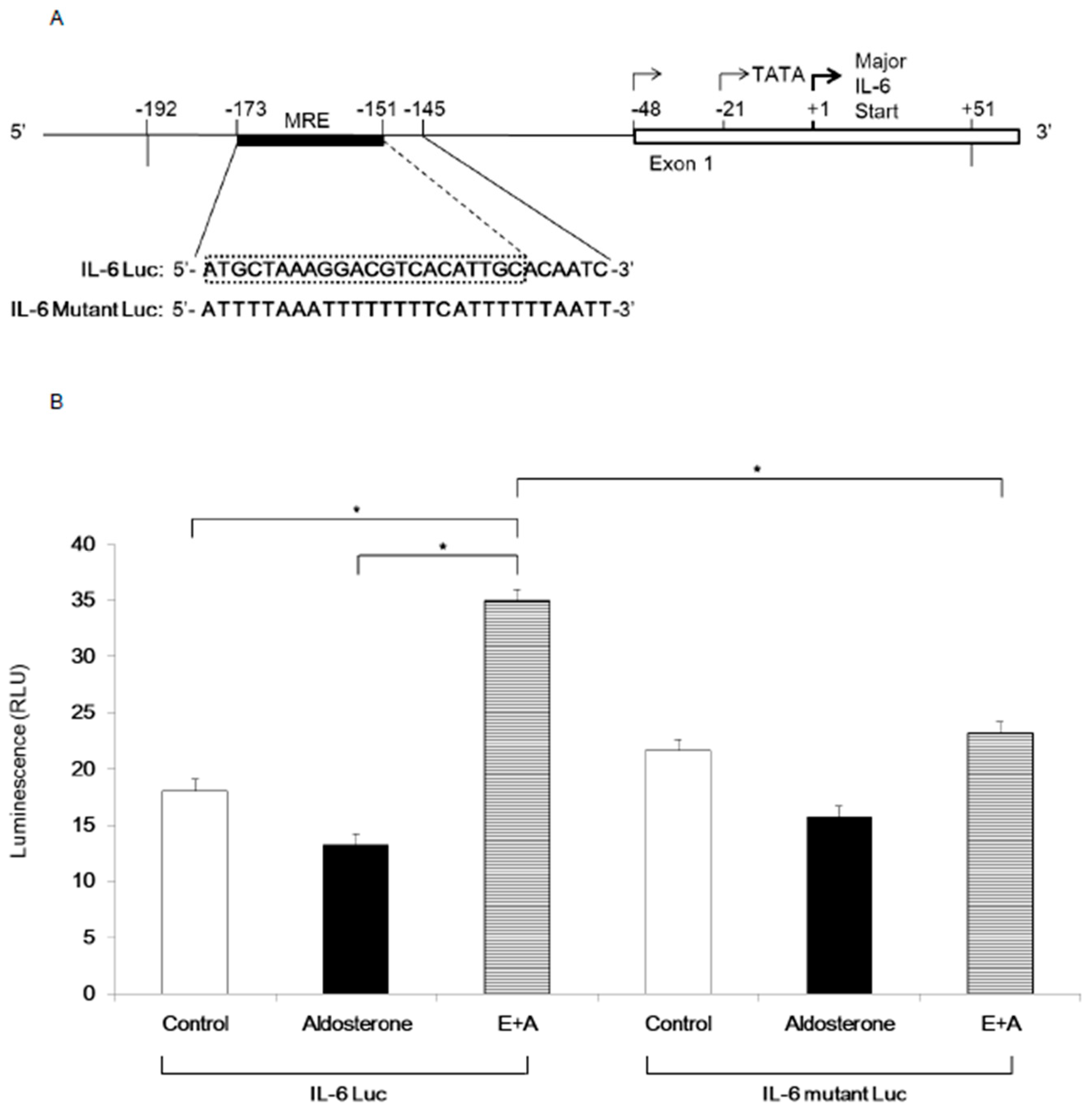
3.4. Induction of IL-6 Expression by Eplerenone Requires MR Binding Element in the IL-6 Promoter
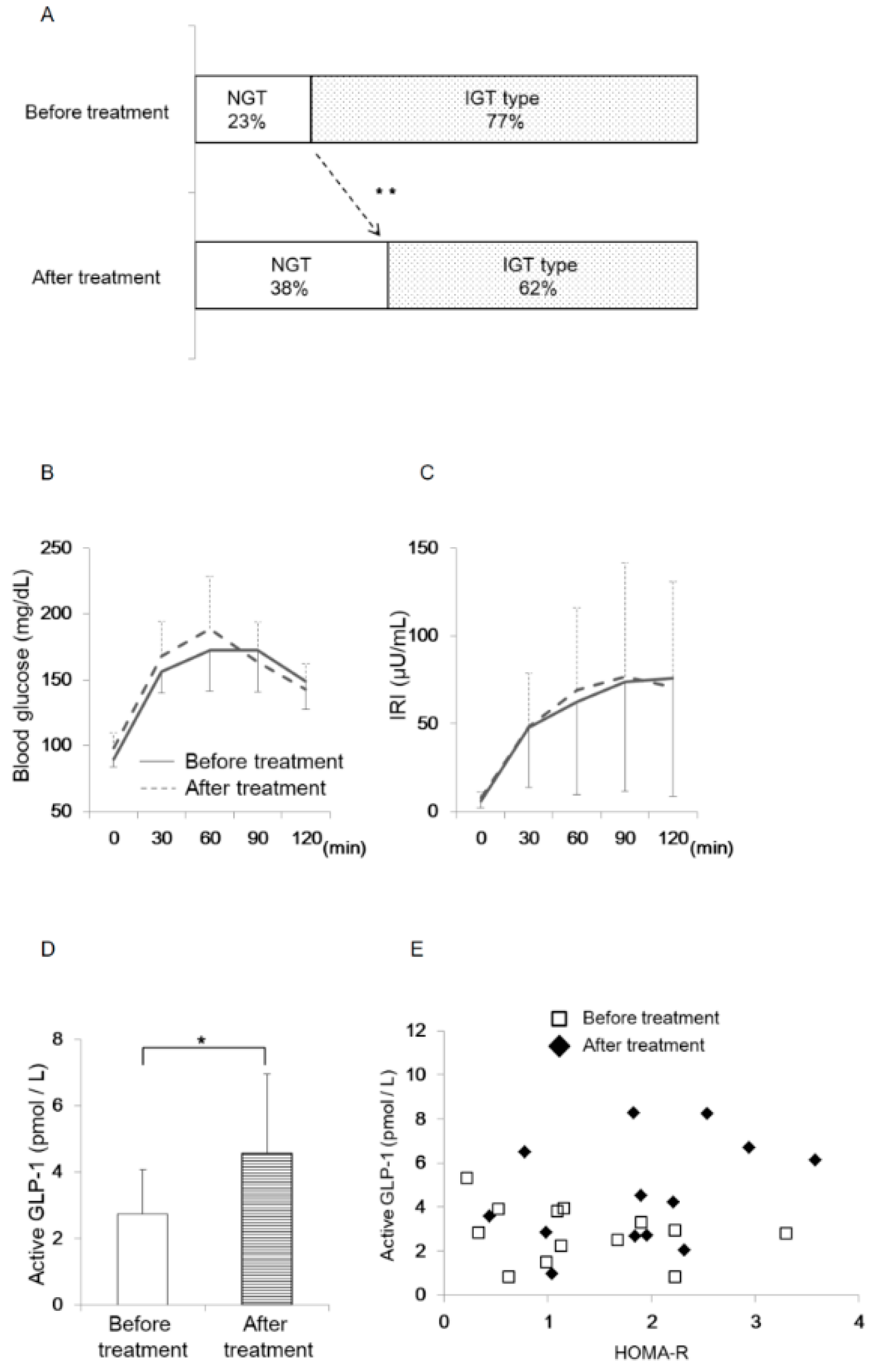
3.5. Oral Eplerenone Treatment Improves the Impairment of Glucose Homeostasis in Patients with PA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Elliottz, W.J.; Meyer, P.M. Incident diabetes in clinical trials of antihypertensive drugs: A network meta-analysis. Lancet 2007, 369, 201–207. [Google Scholar] [CrossRef]
- Luther, J.M.; Brown, N.J. The renin-angiotensin-aldosterone system and glucose homeostasis. Trends Pharmacol. Sci. 2011, 32, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.M. Effects of aldosterone on insulin sensitivity and secretion. Steroids 2014, 91, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.M.; Luo, P.; Kreger, M.T.; Brissova, M.; Dai, C.; Whitfield, T.T.; Kim, H.S.; Wasserman, D.H.; Powers, A.C.; Brown, N.J. Aldosterone decreases glucose-stimulated insulin secretion in vivo in mice and in murine islets. Diabetologia 2011, 54, 2152–2163. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, M.; Yada, T. Sub-chronic stimulation of glucocorticoid receptor impairs and mineralocorticoid receptor protects cytosolic ca2+ responses to glucose in pancreatic beta-cells. J. Endocrinol. 2008, 197, 221–229. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ellingsgaard, H.; Hauselmann, I.; Schuler, B.; Habib, A.M.; Baggio, L.L.; Meier, D.T.; Eppler, E.; Bouzakri, K.; Wueest, S.; Muller, Y.D.; et al. Interleukin-6 enhances insulin secretion by increasing glucagon-like peptide-1 secretion from l cells and alpha cells. Nat. Med. 2011, 17, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: Glp-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef]
- Fernandez-Millan, E.; de Toro-Martin, J.; Lizarraga-Mollinedo, E.; Escriva, F.; Alvarez, C. Role of endogenous il-6 in the neonatal expansion and functionality of wistar rat pancreatic alpha cells. Diabetologia 2013, 56, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Blasi, E.R.; Rocha, R.; Rudolph, A.E.; Blomme, E.A.; Polly, M.L.; McMahon, E.G. Aldosterone/salt induces renal inflammation and fibrosis in hypertensive rats. Kidney Int. 2003, 63, 1791–1800. [Google Scholar] [CrossRef]
- Kiyosue, A.; Nagata, D.; Myojo, M.; Sato, T.; Takahashi, M.; Satonaka, H.; Nagai, R.; Hirata, Y. Aldosterone-induced osteopontin gene transcription in vascular smooth muscle cells involves glucocorticoid response element. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2011, 34, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.A., 3rd; Tan, X.; Wilson, M.; Bhattacharyya, S.; Ludwig, S. Single plasmids expressing human steroid hormone receptors and a reporter gene for use in yeast signaling assays. Plasmid 2010, 63, 73–78. [Google Scholar] [CrossRef]
- Carey, K.L.; Richards, S.A.; Lounsbury, K.M.; Macara, I.G. Evidence using a green fluorescent protein-glucocorticoid receptor chimera that the ran/tc4 gtpase mediates an essential function independent of nuclear protein import. J. Cell Biol. 1996, 133, 985–996. [Google Scholar] [CrossRef]
- Ray, A.; LaForge, K.S.; Sehgal, P.B. On the mechanism for efficient repression of the interleukin-6 promoter by glucocorticoids: Enhancer, tata box and RNA start site (INR motif) occlusion. Mol. Cell. Biol. 1990, 10, 5736–5746. [Google Scholar] [CrossRef] [PubMed]
- Traub, S.; Meier, D.T.; Schulze, F.; Dror, E.; Nordmann, T.M.; Goetz, N.; Koch, N.; Dalmas, E.; Stawiski, M.; Makshana, V.; et al. Pancreatic alpha cell-derived glucagon-related peptides are required for beta cell adaptation and glucose homeostasis. Cell Rep. 2017, 18, 3192–3203. [Google Scholar] [CrossRef]
- Oyadomari, S.; Takeda, K.; Takiguchi, M.; Gotoh, T.; Matsumoto, M.; Wada, I.; Akira, S.; Araki, E.; Mori, M. Nitric oxide-induced apoptosis in pancreatic beta cells is mediated by the endoplasmic reticulum stress pathway. Proc. Natl. Acad. Sci. USA 2001, 98, 10845–10850. [Google Scholar] [CrossRef]
- Mihailidou, C.; Chatzistamou, I.; Papavassiliou, A.G.; Kiaris, H. Modulation of pancreatic islets’ function and survival during aging involves the differential regulation of endoplasmic reticulum stress by p21 and chop. Antioxid. Redox Signal. 2017, 27, 185–200. [Google Scholar] [CrossRef]
- Yusta, B.; Baggio, L.L.; Estall, J.L.; Koehler, J.A.; Holland, D.P.; Li, H.; Pipeleers, D.; Ling, Z.; Drucker, D.J. Glp-1 receptor activation improves beta cell function and survival following induction of endoplasmic reticulum stress. Cell Metab. 2006, 4, 391–406. [Google Scholar] [CrossRef]
- Zennaro, M.C.; Le Menuet, D.; Lombes, M. Characterization of the human mineralocorticoid receptor gene. Mol. Endocrinol. 1996, 10, 1549–1560. [Google Scholar] [PubMed]
- Munier, M.; Meduri, G.; Viengchareun, S.; Leclerc, P.; Le Menuet, D.; Lombes, M. Regulation of mineralocorticoid receptor expression during neuronal differentiation of murine embryonic stem cells. Endocrinology 2010, 151, 2244–2254. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, A.; Wilson, H.L.; Kiss-Toth, E.; Dower, S.K.; North, R.A.; Surprenant, A. Rapid secretion of interleukin-1beta by microvesicle shedding. Immunity 2001, 15, 825–835. [Google Scholar] [CrossRef]
- Samanta, A.; Dawn, B. Il-10 for cardiac autophagy modulation: New direction in the pursuit of perfection. J. Mol. Cell. Cardiol. 2016, 91, 204–206. [Google Scholar] [CrossRef]
- Sabat, R.; Grutz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginat, J. Biology of interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef]
- Hempel, L.; Korholz, D.; Bonig, H.; Schneider, M.; Klein-Vehne, A.; Packeisen, J.; Mauz-Korholz, C.; Burdach, S. Interleukin-10 directly inhibits the interleukin-6 production in t-cells. Scand. J. Immunol. 1995, 41, 462–466. [Google Scholar] [CrossRef]
- Le Menuet, D.; Munier, M.; Meduri, G.; Viengchareun, S.; Lombes, M. Mineralocorticoid receptor overexpression in embryonic stem cell-derived cardiomyocytes increases their beating frequency. Cardiovasc. Res. 2010, 87, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Inaba, H.; Pui, C.H. Glucocorticoid use in acute lymphoblastic leukaemia. Lancet Oncol. 2010, 11, 1096–1106. [Google Scholar] [CrossRef]
- Li, Q.L.; Jansen, E.; Friedman, T.C. Regulation of prohormone convertase 1 (pc1) by gp130-related cytokines. Mol. Cell. Endocrinol. 1999, 158, 143–152. [Google Scholar] [CrossRef]
- Habener, J.F.; Stanojevic, V. Alpha cells come of age. Trends Endocrinol. Metab. 2013, 24, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Chen, C.; Shi, J.; Ren, Z.; Hu, F.; van Goor, H.; Singhal, P.C.; Ding, G. Disparate effects of eplerenone, amlodipine and telmisartan on podocyte injury in aldosterone-infused rats. Nephrol. Dial. Transplant. 2011, 26, 789–799. [Google Scholar] [CrossRef]
- Ramirez, E.; Klett-Mingo, M.; Ares-Carrasco, S.; Picatoste, B.; Ferrarini, A.; Ruperez, F.J.; Caro-Vadillo, A.; Barbas, C.; Egido, J.; Tunon, J.; et al. Eplerenone attenuated cardiac steatosis, apoptosis and diastolic dysfunction in experimental type-ii diabetes. Cardiovasc. Diabetol. 2013, 12, 172. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Li, J.; Wang, M.; She, M.; Tang, Y.; Li, J.; Li, H.; Hui, H. Glp-1 treatment improves diabetic retinopathy by alleviating autophagy through glp-1r-erk1/2-hdac6 signaling pathway. Int. J. Med. Sci. 2017, 14, 1203–1212. [Google Scholar] [CrossRef]
- Piro, S.; Mascali, L.G.; Urbano, F.; Filippello, A.; Malaguarnera, R.; Calanna, S.; Rabuazzo, A.M.; Purrello, F. Chronic exposure to glp-1 increases glp-1 synthesis and release in a pancreatic alpha cell line (alpha-tc1): Evidence of a direct effect of glp-1 on pancreatic alpha cells. PLoS ONE 2014, 9, e90093. [Google Scholar] [CrossRef]
- Kodama, S.; Toyonaga, T.; Kondo, T.; Matsumoto, K.; Tsuruzoe, K.; Kawashima, J.; Goto, H.; Kume, K.; Kume, S.; Sakakida, M.; et al. Enhanced expression of pdx-1 and ngn3 by exendin-4 during beta cell regeneration in stz-treated mice. Biochem. Biophys. Res. Commun. 2005, 327, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Buteau, J. Glp-1 receptor signaling: Effects on pancreatic beta-cell proliferation and survival. Diabetes Metab. 2008, 34 (Suppl. 2), S73–S77. [Google Scholar] [CrossRef]
- Cornu, M.; Yang, J.Y.; Jaccard, E.; Poussin, C.; Widmann, C.; Thorens, B. Glucagon-like peptide-1 protects beta-cells against apoptosis by increasing the activity of an igf-2/igf-1 receptor autocrine loop. Diabetes 2009, 58, 1816–1825. [Google Scholar] [CrossRef]
- Liu, J.; Yin, F.; Xiao, H.; Guo, L.; Gao, X. Glucagon-like peptide 1 receptor plays an essential role in geniposide attenuating lipotoxicity-induced beta-cell apoptosis. Toxicol. In Vitro 2012, 26, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Pullen, T.J.; Huising, M.O.; Rutter, G.A. Analysis of purified pancreatic islet beta and alpha cell transcriptomes reveals 11beta-hydroxysteroid dehydrogenase (hsd11b1) as a novel disallowed gene. Front. Genet. 2017, 8, 41. [Google Scholar] [CrossRef]
- Turban, S.; Liu, X.; Ramage, L.; Webster, S.P.; Walker, B.R.; Dunbar, D.R.; Mullins, J.J.; Seckl, J.R.; Morton, N.M. Optimal elevation of beta-cell 11beta-hydroxysteroid dehydrogenase type 1 is a compensatory mechanism that prevents high-fat diet-induced beta-cell failure. Diabetes 2012, 61, 642–652. [Google Scholar] [CrossRef][Green Version]
- Shimamoto, K.; Shiiki, M.; Ise, T.; Miyazaki, Y.; Higashiura, K.; Fukuoka, M.; Hirata, A.; Masuda, A.; Nakagawa, M.; Iimura, O. Does insulin resistance participate in an impaired glucose tolerance in primary aldosteronism? J. Hum. Hypertens. 1994, 8, 755–759. [Google Scholar] [PubMed]
- Watanabe, D.; Yatabe, M.; Ichihara, A. Evaluation of insulin sensitivity and secretion in primary aldosteronism. Clin. Exp. Hypertens. 2016, 38, 613–617. [Google Scholar] [CrossRef]
- Nagai, E.; Katsuno, T.; Miyagawa, J.; Konishi, K.; Miuchi, M.; Ochi, F.; Kusunoki, Y.; Tokuda, M.; Murai, K.; Hamaguchi, T.; et al. Incretin responses to oral glucose load in japanese non-obese healthy subjects. Diabetes Ther. Res. Treat. Educ. Diabetes Relat. Disord. 2011, 2, 20–28. [Google Scholar] [CrossRef][Green Version]
- Muscelli, E.; Mari, A.; Casolaro, A.; Camastra, S.; Seghieri, G.; Gastaldelli, A.; Holst, J.J.; Ferrannini, E. Separate impact of obesity and glucose tolerance on the incretin effect in normal subjects and type 2 diabetic patients. Diabetes 2008, 57, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Lastya, A.; Saraswati, M.R.; Suastika, K. The low level of glucagon-like peptide-1 (glp-1) is a risk factor of type 2 diabetes mellitus. BMC Res. Notes 2014, 7, 849. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.L.; Chu, C.; Lv, Y.B.; Wang, Y.; Hu, J.W.; Ma, Q.; Yan, Y.; Cao, Y.M.; Dang, X.L.; Wang, K.K.; et al. Effect of salt intake on serum glucagon-like peptide-1 levels in normotensive salt-sensitive subjects. Kidney Blood Press. Res. 2017, 42, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Krug, A.W.; Stelzner, L.; Rao, A.D.; Lichtman, A.H.; Williams, G.H.; Adler, G.K. Effect of low dose mineralocorticoid receptor antagonist eplerenone on glucose and lipid metabolism in healthy adult males. Metab. Clin. Exp. 2013, 62, 386–391. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goto, R.; Kondo, T.; Ono, K.; Kitano, S.; Miyakawa, N.; Watanabe, T.; Sakaguchi, M.; Sato, M.; Igata, M.; Kawashima, J.; et al. Mineralocorticoid Receptor May Regulate Glucose Homeostasis through the Induction of Interleukin-6 and Glucagon-Like peptide-1 in Pancreatic Islets. J. Clin. Med. 2019, 8, 674. https://doi.org/10.3390/jcm8050674
Goto R, Kondo T, Ono K, Kitano S, Miyakawa N, Watanabe T, Sakaguchi M, Sato M, Igata M, Kawashima J, et al. Mineralocorticoid Receptor May Regulate Glucose Homeostasis through the Induction of Interleukin-6 and Glucagon-Like peptide-1 in Pancreatic Islets. Journal of Clinical Medicine. 2019; 8(5):674. https://doi.org/10.3390/jcm8050674
Chicago/Turabian StyleGoto, Rieko, Tatsuya Kondo, Kaoru Ono, Sayaka Kitano, Nobukazu Miyakawa, Takuro Watanabe, Masaji Sakaguchi, Miki Sato, Motoyuki Igata, Junji Kawashima, and et al. 2019. "Mineralocorticoid Receptor May Regulate Glucose Homeostasis through the Induction of Interleukin-6 and Glucagon-Like peptide-1 in Pancreatic Islets" Journal of Clinical Medicine 8, no. 5: 674. https://doi.org/10.3390/jcm8050674
APA StyleGoto, R., Kondo, T., Ono, K., Kitano, S., Miyakawa, N., Watanabe, T., Sakaguchi, M., Sato, M., Igata, M., Kawashima, J., Motoshima, H., Matsumura, T., Shimoda, S., & Araki, E. (2019). Mineralocorticoid Receptor May Regulate Glucose Homeostasis through the Induction of Interleukin-6 and Glucagon-Like peptide-1 in Pancreatic Islets. Journal of Clinical Medicine, 8(5), 674. https://doi.org/10.3390/jcm8050674