Increased Risk of Multiple Outpatient Surgeries in African-American Carriers of Transthyretin Val122Ile Mutation Is Modulated by Non-Coding Variants


Abstract
1. Introduction
2. Methods
2.1. Study Population
2.2. Genetic Data
2.3. Statistical Analysis
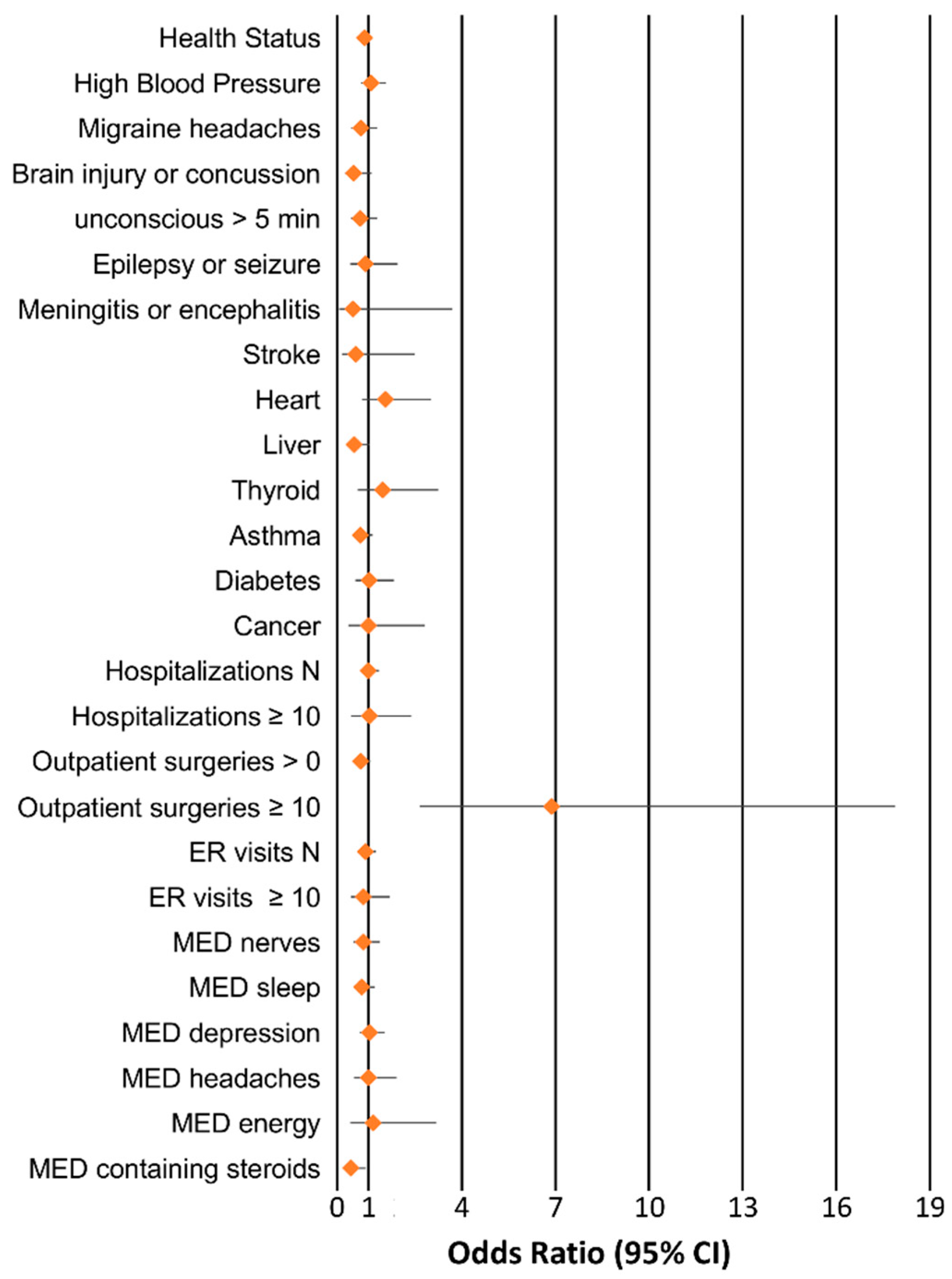
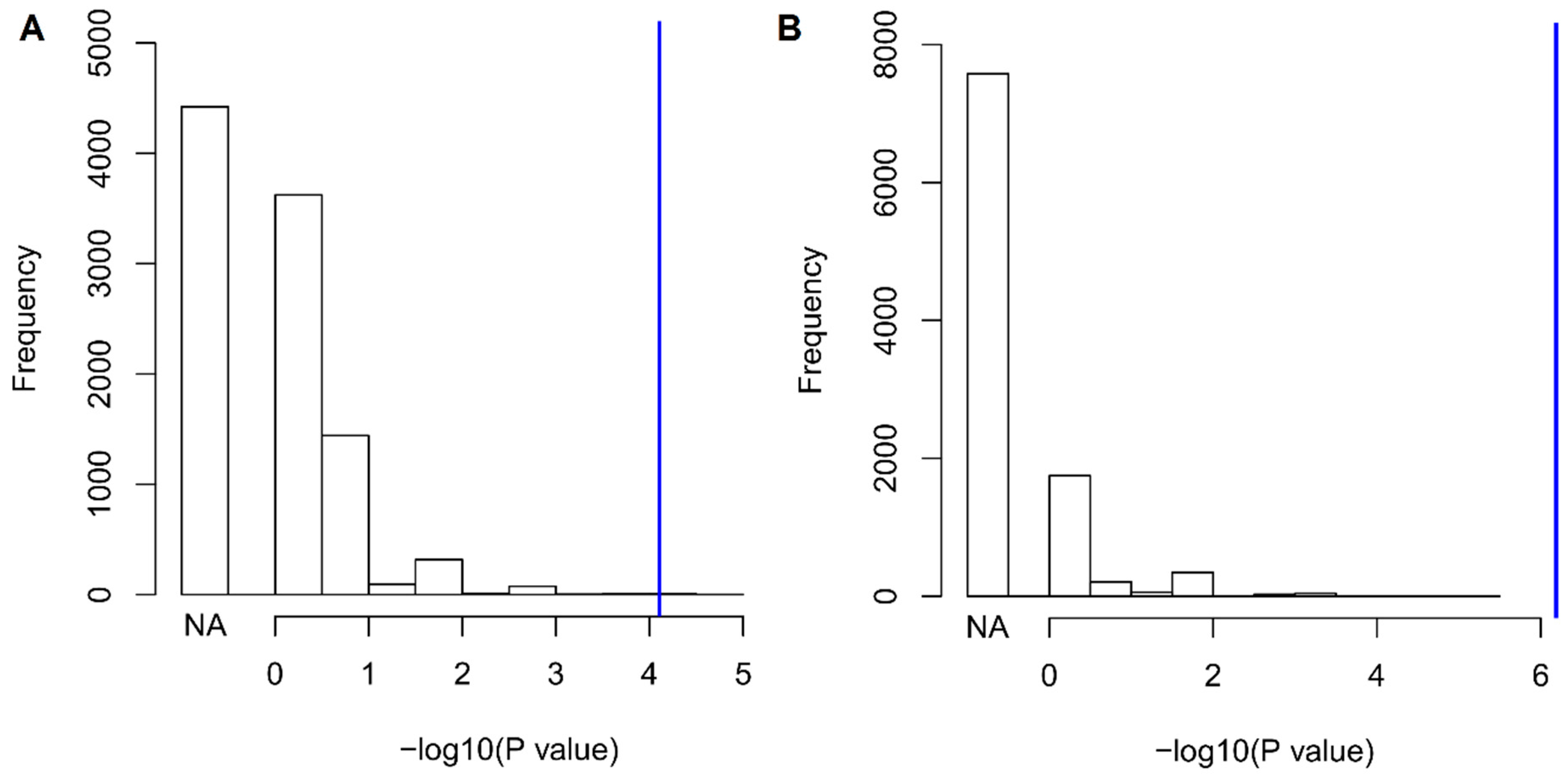
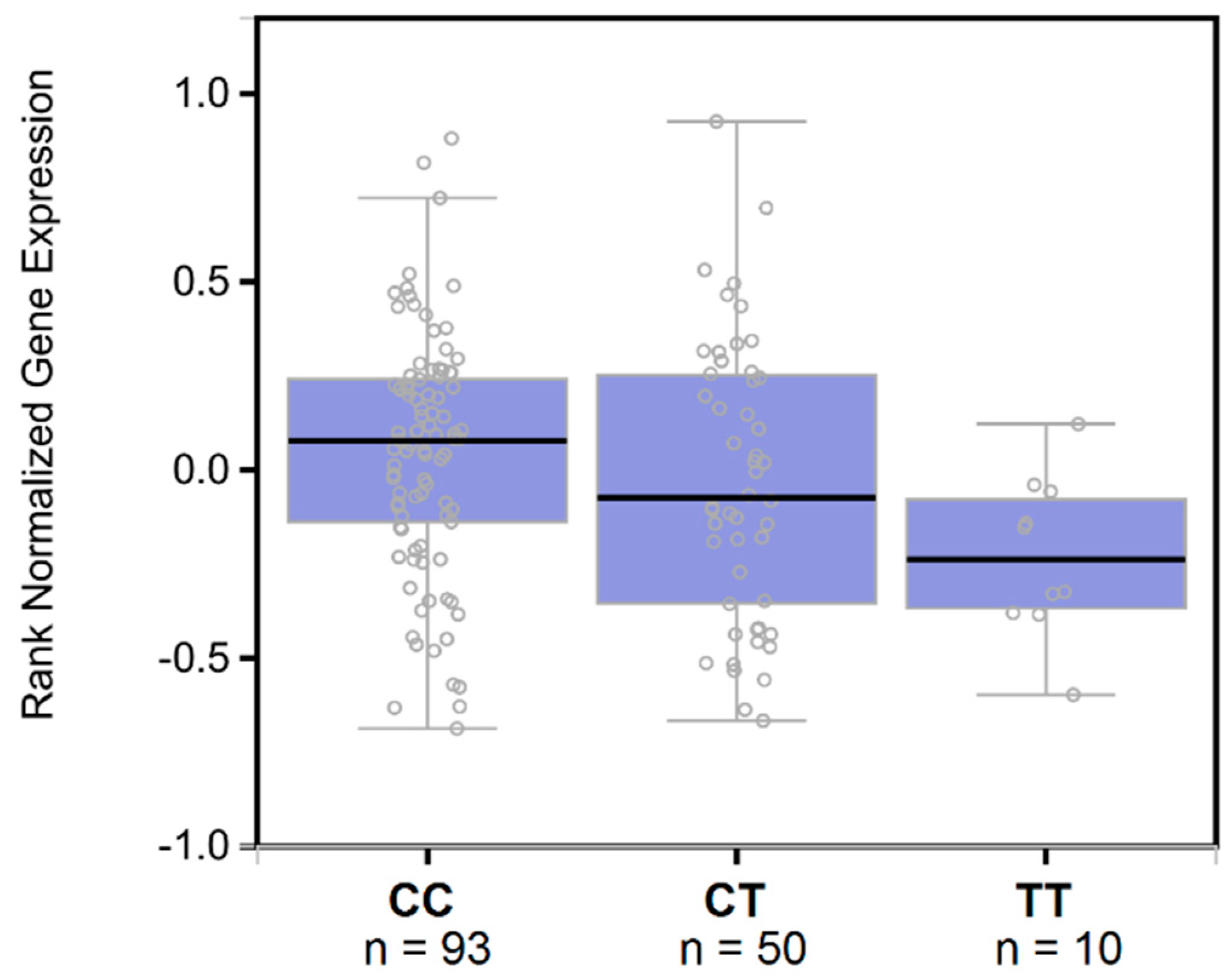
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Plante-Bordeneuve, V.; Said, G. Familial amyloid polyneuropathy. Lancet Neurol. 2011, 10, 1086–1097. [Google Scholar] [CrossRef]
- Ruberg, F.L.; Berk, J.L. Transthyretin (ttr) cardiac amyloidosis. Circulation 2012, 126, 1286–1300. [Google Scholar] [CrossRef] [PubMed]
- Parman, Y.; Adams, D.; Obici, L.; Galan, L.; Guergueltcheva, V.; Suhr, O.B.; Coelho, T. European Network for TTR-FAP (ATTReuNET). Sixty years of transthyretin familial amyloid polyneuropathy (ttr-fap) in europe: Where are we now? A european network approach to defining the epidemiology and management patterns for ttr-fap. Curr. Opin. Neurol. 2016, 29 (Suppl. 1), S3–S13. [Google Scholar] [CrossRef] [PubMed]
- Buxbaum, J.N.; Ruberg, F.L. Transthyretin v122i (pv142i)* cardiac amyloidosis: An age-dependent autosomal dominant cardiomyopathy too common to be overlooked as a cause of significant heart disease in elderly african americans. Genet. Med. 2017, 19, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Araki, S.; Ando, Y. Transthyretin-related familial amyloidotic polyneuropathy-progress in kumamoto, japan (1967-2010). Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, D.R.; Alexander, A.A.; Tagoe, C.; Buxbaum, J.N. Prevalence of the amyloidogenic transthyretin (ttr) v122i allele in 14 333 african-americans. Amyloid 2015, 22, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, D.R.; Alexander, A.A.; Tagoe, C.; Garvey, W.T.; Williams, S.M.; Tishkoff, S.; Modiano, D.; Sirima, S.B.; Kalidi, I.; Toure, A.; et al. The prevalence and distribution of the amyloidogenic transthyretin (ttr) v122i allele in africa. Mol. Genet. Genomic Med. 2016, 4, 548–556. [Google Scholar] [CrossRef]
- Reddi, H.V.; Jenkins, S.; Theis, J.; Thomas, B.C.; Connors, L.H.; Van Rhee, F.; Highsmith, W.E., Jr. Homozygosity for the v122i mutation in transthyretin is associated with earlier onset of cardiac amyloidosis in the african american population in the seventh decade of life. J. Mol. Diagn. 2014, 16, 68–74. [Google Scholar] [CrossRef]
- Alves-Ferreira, M.; Coelho, T.; Santos, D.; Sequeiros, J.; Alonso, I.; Sousa, A.; Lemos, C. A trans-acting factor may modify age at onset in familial amyloid polyneuropathy attrv30m in portugal. Mol. Neurobiol. 2018, 55, 3676–3683. [Google Scholar] [CrossRef]
- Polimanti, R.; Di Girolamo, M.; Manfellotto, D.; Fuciarelli, M. In silico analysis of ttr gene (coding and non-coding regions, and interactive network) and its implications in transthyretin-related amyloidosis. Amyloid 2014, 21, 154–162. [Google Scholar] [CrossRef]
- Iorio, A.; De Angelis, F.; Di Girolamo, M.; Luigetti, M.; Pradotto, L.; Mauro, A.; Manfellotto, D.; Fuciarelli, M.; Polimanti, R. Most recent common ancestor of ttr val30met mutation in italian population and its potential role in genotype-phenotype correlation. Amyloid 2015, 22, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Polimanti, R.; Di Girolamo, M.; Manfellotto, D.; Fuciarelli, M. Functional variation of the transthyretin gene among human populations and its correlation with amyloidosis phenotypes. Amyloid 2013, 20, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Iorio, A.; De Angelis, F.; Di Girolamo, M.; Luigetti, M.; Pradotto, L.G.; Mazzeo, A.; Frusconi, S.; My, F.; Manfellotto, D.; Fuciarelli, M.; et al. Population diversity of the genetically determined ttr expression in human tissues and its implications in ttr amyloidosis. BMC Genom. 2017, 18, 254. [Google Scholar] [CrossRef] [PubMed]
- Iorio, A.; De Lillo, A.; De Angelis, F.; Di Girolamo, M.; Luigetti, M.; Sabatelli, M.; Pradotto, L.; Mauro, A.; Mazzeo, A.; Stancanelli, C.; et al. Non-coding variants contribute to the clinical heterogeneity of ttr amyloidosis. Eur. J. Hum. Genet. 2017, 25, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Pierucci-Lagha, A.; Gelernter, J.; Feinn, R.; Cubells, J.F.; Pearson, D.; Pollastri, A.; Farrer, L.; Kranzler, H.R. Diagnostic reliability of the semi-structured assessment for drug dependence and alcoholism (ssadda). Drug Alcohol Depend. 2005, 80, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Pierucci-Lagha, A.; Gelernter, J.; Chan, G.; Arias, A.; Cubells, J.F.; Farrer, L.; Kranzler, H.R. Reliability of dsm-iv diagnostic criteria using the semi-structured assessment for drug dependence and alcoholism (ssadda). Drug Alcohol Depend. 2007, 91, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Polimanti, R.; Zhao, H.; Farrer, L.A.; Kranzler, H.R.; Gelernter, J. Ancestry-specific and sex-specific risk alleles identified in a genome-wide gene-by-alcohol dependence interaction study of risky sexual behaviors. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2017, 174, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Polimanti, R.; Kaufman, J.; Zhao, H.; Kranzler, H.R.; Ursano, R.J.; Kessler, R.C.; Gelernter, J.; Stein, M.B. A genome-wide gene-by-trauma interaction study of alcohol misuse in two independent cohorts identifies prkg1 as a risk locus. Mol. Psychiatry 2018, 23, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Gelernter, J.; Kranzler, H.R.; Sherva, R.; Koesterer, R.; Almasy, L.; Zhao, H.; Farrer, L.A. Genome-wide association study of opioid dependence: Multiple associations mapped to calcium and potassium pathways. Biol. Psychiatry 2014, 76, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Polimanti, R.; Zhang, H.; Smith, A.H.; Zhao, H.; Farrer, L.A.; Kranzler, H.R.; Gelernter, J. Genome-wide association study of body mass index in subjects with alcohol dependence. Addict. Biol. 2017, 22, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Forer, L.; Schonherr, S.; Sidore, C.; Locke, A.E.; Kwong, A.; Vrieze, S.I.; Chew, E.Y.; Levy, S.; McGue, M.; et al. Next-generation genotype imputation service and methods. Nat. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef] [PubMed]
- 1000 Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation plink: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C. Haploview: Visualization and analysis of snp genotype data. Cold Spring Harb. Protoc. 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.D.; Kellis, M. Haploreg: A resource for exploring chromatin states, conservation, and regulatory motif alterations within sets of genetically linked variants. Nucleic Acids Res. 2012, 40, D930–D934. [Google Scholar] [CrossRef] [PubMed]
- Boyle, A.P.; Hong, E.L.; Hariharan, M.; Cheng, Y.; Schaub, M.A.; Kasowski, M.; Karczewski, K.J.; Park, J.; Hitz, B.C.; Weng, S.; et al. Annotation of functional variation in personal genomes using regulomedb. Genome. Res. 2012, 22, 1790–1797. [Google Scholar] [CrossRef] [PubMed]
- GTEx Consortium. Human genomics. The genotype-tissue expression (gtex) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef]
- Jacobson, D.; Tagoe, C.; Schwartzbard, A.; Shah, A.; Koziol, J.; Buxbaum, J. Relation of clinical, echocardiographic and electrocardiographic features of cardiac amyloidosis to the presence of the transthyretin v122i allele in older african-american men. Am. J. Cardiol. 2011, 108, 440–444. [Google Scholar] [CrossRef]
- Sun, B.B.; Maranville, J.C.; Peters, J.E.; Stacey, D.; Staley, J.R.; Blackshaw, J.; Burgess, S.; Jiang, T.; Paige, E.; Surendran, P.; et al. Genomic atlas of the human plasma proteome. Nature 2018, 558, 73–79. [Google Scholar] [CrossRef]
- Akasaki, Y.; Reixach, N.; Matsuzaki, T.; Alvarez-Garcia, O.; Olmer, M.; Iwamoto, Y.; Buxbaum, J.N.; Lotz, M.K. Transthyretin deposition in articular cartilage: A novel mechanism in the pathogenesis of osteoarthritis. Arthritis Rheumatol. 2015, 67, 2097–2107. [Google Scholar] [CrossRef] [PubMed]
- Bert, J.M.; Hooper, J.; Moen, S. Outpatient total joint arthroplasty. Curr. Rev. Musculoskelet Med. 2017, 10, 567–574. [Google Scholar] [CrossRef] [PubMed]
- UK Biobank GWAS. Available online: https://github.com/Nealelab/UK_Biobank_GWAS (accessed on 2 February 2019).
- Cappelli, F.; Frusconi, S.; Bergesio, F.; Grifoni, E.; Fabbri, A.; Giuliani, C.; Falconi, S.; Bonifacio, S.; Perfetto, F. The val142ile transthyretin cardiac amyloidosis: Not only an afro-american pathogenic variant? A single-centre italian experience. J. Cardiovasc Med. (Hagerstown) 2016, 17, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Coelho, T.; Maurer, M.S.; Suhr, O.B. Thaos-the transthyretin amyloidosis outcomes survey: Initial report on clinical manifestations in patients with hereditary and wild-type transthyretin amyloidosis. Curr. Med. Res. Opin. 2013, 29, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Maurer, M.S.; Hanna, M.; Grogan, M.; Dispenzieri, A.; Witteles, R.; Drachman, B.; Judge, D.P.; Lenihan, D.J.; Gottlieb, S.S.; Shah, S.J.; et al. Genotype and phenotype of transthyretin cardiac amyloidosis: Thaos (transthyretin amyloid outcome survey). J. Am. Coll. Cardiol. 2016, 68, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Santos, M.J.; Alves-Ferreira, M.; Coelho, T.; Sequeiros, J.; Alonso, I.; Oliveira, P.; Sousa, A.; Lemos, C.; Grazina, M. Mtdna copy number associated with age of onset in familial amyloid polyneuropathy. J. Neurol. Neurosurg Psychiatry 2018, 89, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Panayiotou, E.; Papacharalambous, R.; Antoniou, A.; Christophides, G.; Papageorgiou, L.; Fella, E.; Malas, S.; Kyriakides, T. Genetic background modifies amyloidosis in a mouse model of attr neuropathy. Biochem. Biophys. Rep. 2016, 8, 48–54. [Google Scholar] [CrossRef]
- Chan, G.G.; Koch, C.M.; Connors, L.H. Serum proteomic variability associated with clinical phenotype in familial transthyretin amyloidosis (attrm). J. Proteome Res. 2017, 16, 4104–4112. [Google Scholar] [CrossRef]
- Moreira, L.; Beirao, J.M.; Beirao, I.; Pinho e Costa, P. Oligomeric ttr v30m aggregates compromise cell viability, erythropoietin gene expression and promoter activity in the human hepatoma cell line hep3b. Amyloid 2015, 22, 93–99. [Google Scholar] [CrossRef]
- Haga, S.B. Impact of limited population diversity of genome-wide association studies. Genet. Med. 2010, 12, 81–84. [Google Scholar] [CrossRef]
- Carr, A.S.; Pelayo-Negro, A.L.; Jaunmuktane, Z.; Scalco, R.S.; Hutt, D.; Evans, M.R.; Heally, E.; Brandner, S.; Holton, J.; Blake, J.; et al. Transthyretin v122i amyloidosis with clinical and histological evidence of amyloid neuropathy and myopathy. Neuromuscul. Disord. 2015, 25, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.; Alvarez, J.; Teruya, S.; Castano, A.; Lehman, R.A.; Weidenbaum, M.; Geller, J.A.; Helmke, S.; Maurer, M.S. Hip and knee arthroplasty are common among patients with transthyretin cardiac amyloidosis, occurring years before cardiac amyloid diagnosis: Can we identify affected patients earlier? Amyloid 2017, 24, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.; Suhr, O.B.; Hund, E.; Obici, L.; Tournev, I.; Campistol, J.M.; Slama, M.S.; Hazenberg, B.P.; Coelho, T. European Network for TTR-FAP (ATTReuNET). First european consensus for diagnosis, management, and treatment of transthyretin familial amyloid polyneuropathy. Curr. Opin. Neurol. 2016, 29 (Suppl. 1), S14–S26. [Google Scholar] [CrossRef] [PubMed]
- Quarta, C.C.; Buxbaum, J.N.; Shah, A.M.; Falk, R.H.; Claggett, B.; Kitzman, D.W.; Mosley, T.H.; Butler, K.R.; Boerwinkle, E.; Solomon, S.D. The amyloidogenic v122i transthyretin variant in elderly black americans. New Engl. J. Med. 2015, 372, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, G.; Ribeiro-Silva, C.; Ribeiro, R.; Gilberto, S.; Gomes, R.A.; Ferreira, A.; Mateus, E.; Barroso, E.; Coelho, A.V.; Freire, A.P.; et al. Transthyretin amyloidosis: Chaperone concentration changes and increased proteolysis in the pathway to disease. PLoS ONE 2015, 10, e0125392. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study Population (n = 4361) | |
| Age years, IQR | 35–48 |
| Sex, Women (%) | 1924 (44) |
| Val122Ile, Y (%) | 152 (3.5) |
| Health Status | |
| Excellent (%) | 675 (15) |
| Very Good (%) | 1135 (26) |
| Good (%) | 1420 (33) |
| Fair (%) | 940 (22) |
| Poor (%) | 191 (4) |
| Doctor’s diagnoses | |
| High blood pressure, Y (%) | 1325 (30) |
| Migraine headaches, Y (%) | 605 (14) |
| Brain injury or concussion, Y (%) | 388 (9) |
| Been unconscious for longer than 5 min, Y (%) | 554 (13) |
| Epilepsy or have had a seizure, Y (%) | 213 (5) |
| Meningitis or encephalitis, Y (%) | 57 (1) |
| Stroke, Y (%) | 88 (2) |
| Heart disease, Y (%) | 182 (4) |
| Liver disease, Y (%) | 481 (11) |
| Thyroid disease, Y (%) | 134 (3) |
| Asthma, Y (%) | 928 (21) |
| Diabetes, Y (%) | 374 (9) |
| Cancer, Y (%) | 108 (2) |
| Hospitalizations | |
| N, IQR | 0–3 |
| ≥10, Y (%) | 161 (4) |
| Outpatient surgeries | |
| >0, Y (%) | 1945 (45) |
| ≥10, Y (%) | 24 (0.6) |
| Emergency room visits | |
| N, IQR | 0–3 |
| ≥10, Y (%) | 300 (7) |
| Prescription medications | |
| for nerves, Y (%) | 610 (14) |
| for sleep, Y (%) | 924 (21) |
| for depression, Y (%) | 1012 (23) |
| for headaches, Y (%) | 292 (7) |
| for energy, Y (%) | 95 (2) |
| containing steroids, Y (%) | 451 (10) |
| LD Block | Location (Chr18) | TTR Region | Haplotype | Case-Control Frequencies | Chi Square | p Value |
|---|---|---|---|---|---|---|
| #12 | 28,387,576-28,391,602 | Upstream | GCACCGTATGGAGGGACCTC | 0.375, 0.030 | 24.395 | 7.85 × 10−7 |
| #14 | 28,456,564-28,481,528 | Upstream | TCTGTGTAGGGCC | 0.250, 0.007 | 35.494 | 2.56 × 10−9 |
| #15 | 28,481,625-28,484,316 | Upstream | GACAGA | 0.250, 0.007 | 35.494 | 2.56 × 10−9 |
| #16 | 28,509,237-28,525,062 | Upstream | TTTGTCCCTACAGGCCTTT | 0.500, 0.061 | 22.381 | 2.24 × 10−6 |
| #23 | 28,629,162-28,640,811 | Upstream | GAACTAATACCGA | 0.250, 0.007 | 35.344 | 2.76 × 10−9 |
| Block #16 Haplotype | V122I (G > A) | Case-Control Frequencies | Chi Square | p Value |
|---|---|---|---|---|
| CTTGTTCATACAGGCTATT | A | 0.286, 0.417 | 0.551 | 0.4581 |
| CTTGTTCATACAGGCTATT | G | 0.214, 0.300 | 0.275 | 0.5998 |
| TAAACCACCGTACATCTGC | G | 0.000, 0.075 | 0.646 | 0.4215 |
| TAAACCACCGTGCATCTGC | G | 0.000, 0.066 | 0.568 | 0.4509 |
| TTTGTCCCTACAGGCCTTT | A | 0.339, 0.048 | 12.482 | 4 × 10−4 |
| TAAACCACCGTACATCTG | A | 0.000, 0.033 | 0.276 | 0.5996 |
| TTTGTCCCTACAGGCCTTT | G | 0.161, 0.013 | 10.55 | 0.0012 |
| CTTGTTCATACAGGCCTTT | G | 0.000, 0.017 | 0.137 | 0.7117 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polimanti, R.; Nuñez, Y.Z.; Gelernter, J. Increased Risk of Multiple Outpatient Surgeries in African-American Carriers of Transthyretin Val122Ile Mutation Is Modulated by Non-Coding Variants. J. Clin. Med. 2019, 8, 269. https://doi.org/10.3390/jcm8020269
Polimanti R, Nuñez YZ, Gelernter J. Increased Risk of Multiple Outpatient Surgeries in African-American Carriers of Transthyretin Val122Ile Mutation Is Modulated by Non-Coding Variants. Journal of Clinical Medicine. 2019; 8(2):269. https://doi.org/10.3390/jcm8020269
Chicago/Turabian StylePolimanti, Renato, Yaira Z. Nuñez, and Joel Gelernter. 2019. "Increased Risk of Multiple Outpatient Surgeries in African-American Carriers of Transthyretin Val122Ile Mutation Is Modulated by Non-Coding Variants" Journal of Clinical Medicine 8, no. 2: 269. https://doi.org/10.3390/jcm8020269
APA StylePolimanti, R., Nuñez, Y. Z., & Gelernter, J. (2019). Increased Risk of Multiple Outpatient Surgeries in African-American Carriers of Transthyretin Val122Ile Mutation Is Modulated by Non-Coding Variants. Journal of Clinical Medicine, 8(2), 269. https://doi.org/10.3390/jcm8020269