1,25(OH)2D3 Differently Affects Immunomodulatory Activities of Mesenchymal Stem Cells Depending on the Presence of TNF-α, IL-1β and IFN-γ

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experimental Section
2.1. Ethics
2.2. Primary hPDLSC Isolation and Cultivation
2.3. Isolation of Primary CD4+ T Lymphocytes and CD14+ Monocytes
2.4. CD4+ T Lymphocyte Proliferation in Coculture with hPDLSCs
2.5. hPDLSCs and Macrophage Indirect Coculture
2.6. Immunomediator Expression in hPDLSCs
2.7. Statistical Analysis
3. Results
3.1. 1,25(OH)2D3 Differently Influences hPDLSC Mediated Suppression of CD4+ T Lymphocyte Proliferation Depending on the Presence of TNF-α, IL-1β and IFN-γ
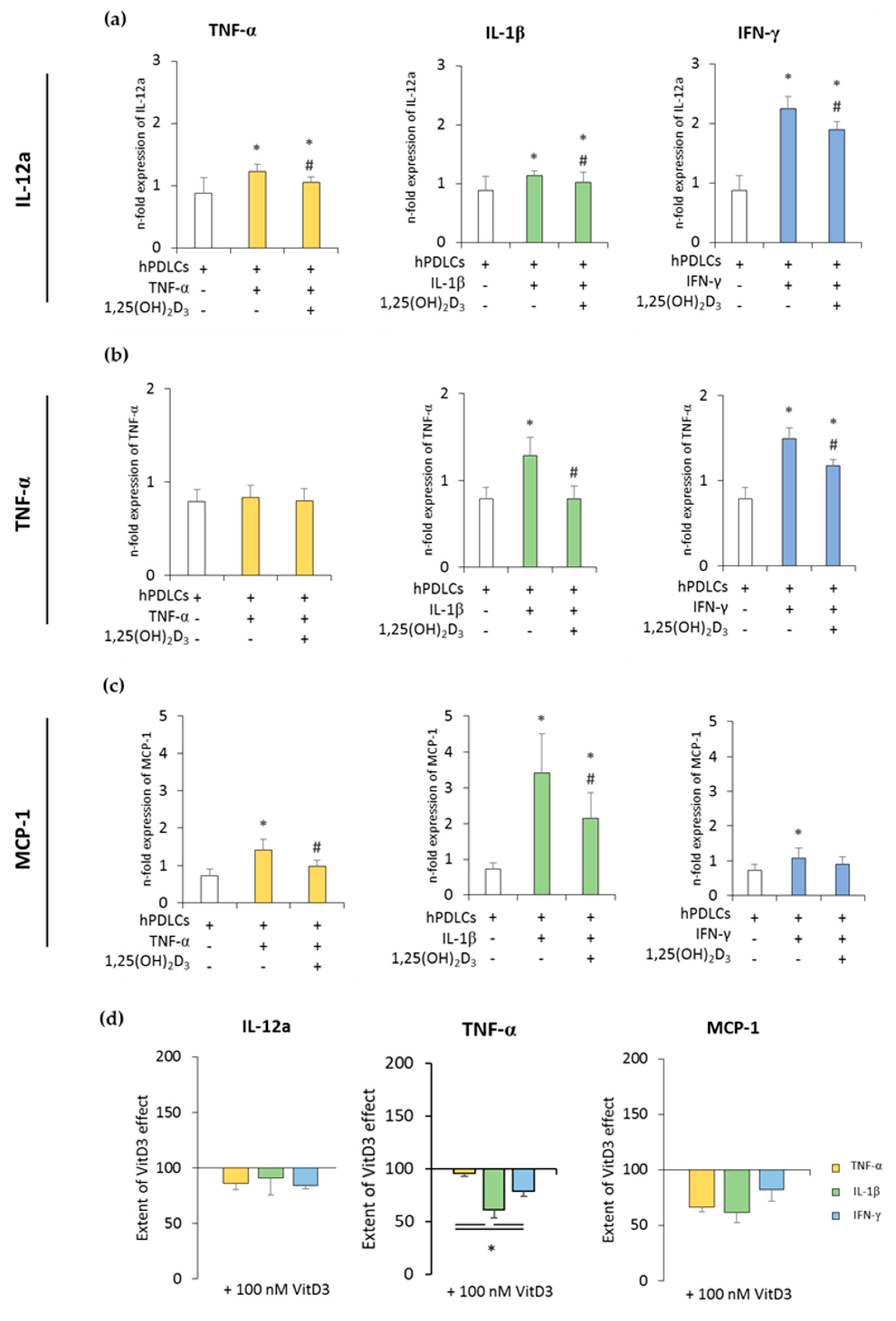
3.2. Expression of Pro- and Anti-Inflammatory Cytokines in CD68+ Macrophages is Differently Affected by hPDLSCs Primed with 1,25(OH)2D3 in the Presence of TNF-α or IL-1β or IFN-γ
3.3. 1,25(OH)2D3 Differently Influences IDO-1 Expression and Enzymatic Activity in hPDLSCs Depending on the Presence of TNF-α or IL-1β or IFN-γ
3.4. 1,25(OH)2D3 Differently Influences TSG-6 Expression in hPDLSCs, Depending on the Presence of TNF-α or IL-1β or IFN-γ
3.5. 1,25(OH)2D3 Differently Influences PTGS-2 Expression in hPDLSCs, Depending on the Presence of TNF-α or IL-1β or IFN-γ
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lips, P. Vitamin D physiology. Prog. Biophys. Mol. Biol. 2006, 92, 4–8. [Google Scholar] [CrossRef] [PubMed]
- White, J.H. Vitamin D signaling, infectious diseases, and regulation of innate immunity. Infect. Immun. 2008, 76, 3837–3843. [Google Scholar] [CrossRef] [PubMed]
- Jones, G. Pharmacokinetics of vitamin D toxicity. Am. J. Clin. Nutr. 2008, 88, 582S–586S. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, D. Extrarenal Expression of 25-Hydroxyvitamin d(3)-1 alpha-hydroxylase. J. Clin. Endocrinol. Metab. 2001, 86, 888–894. [Google Scholar] [PubMed]
- Wu, S.; Ren, S.; Nguyen, L.; Adams, J.S.; Hewison, M. Splice variants of the CYP27b1 gene and the regulation of 1,25-dihydroxyvitamin D3 production. Endocrinology 2007, 148, 3410–3418. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Meng, H.; Hou, J. Activity of 25-hydroxylase in human gingival fibroblasts and periodontal ligament cells. PLoS ONE 2012, 7, e52053. [Google Scholar] [CrossRef] [PubMed]
- Andrukhov, O.; Andrukhova, O.; Hulan, U.; Tang, Y.; Bantleon, H.-P.; Rausch-Fan, X. Both 25-hydroxyvitamin-D3 and 1,25-dihydroxyvitamin-D3 reduces inflammatory response in human periodontal ligament cells. PLoS ONE 2014, 9, e90301. [Google Scholar] [CrossRef]
- Beertsen, W.; McCulloch, C.A.G.; Sodek, J. The periodontal ligament: A unique, multifunctional connective tissue. Periodontology 2000 1997, 13, 20–40. [Google Scholar] [CrossRef]
- Seo, B.M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Najar, M.; Bouhtit, F.; Melki, R.; Afif, H.; Hamal, A.; Fahmi, H.; Merimi, M.; Lagneaux, L. Mesenchymal stromal cell-based therapy: New perspectives and challenges. J. Clin. Med. 2019, 8, 626. [Google Scholar] [CrossRef]
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, P.T.; Yianni, V. Perivascular-derived mesenchymal stem cells. Crit. Rev. Oral Biol. Med. 2019, 98, 1066–1072. [Google Scholar]
- Shi, S.; Gronthos, S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J. Bone Miner. Res. 2003, 18, 696–704. [Google Scholar] [CrossRef]
- Racz, G.Z.; Kadar, K.; Foldes, A.; Kallo, K.; Perczel-Kovach, K.; Keremi, B.; Nagy, A.; Varga, G. Immunomodulatory and potential therapeutic role of mesenchymal stem cells in periodontitis. J. Physiol. Pharmacol. 2014, 65, 327–339. [Google Scholar]
- Xiao, L.; Nasu, M. From regenerative dentistry to regenerative medicine: Progress, challenges, and potential applications of oral stem cells. Stem Cells Cloning Adv. Appl. 2014, 7, 89–99. [Google Scholar] [CrossRef]
- Tipnis, S.; Viswanathan, C.; Majumdar, A.S. Immunosuppressive properties of human umbilical cord-derived mesenchymal stem cells: Role of B7-H1 and IDO. Immunol. Cell Biol. 2010, 88, 795–806. [Google Scholar] [CrossRef]
- Andrukhov, O.; Behm, C.; Blufstein, A.; Rausch-Fan, X. Immunomodulatory properties of dental tissue-derived mesenchymal stem cells: Implication in disease and tissue regeneration. World J. Stem Cells 2019, 11, 604–617. [Google Scholar] [CrossRef]
- Wada, N.; Gronthos, S.; Bartold, P.M. Immunomodulatory effects of stem cells. Periodontology 2000 2013, 63, 198–216. [Google Scholar] [CrossRef]
- Krampera, M. Mesenchymal stromal cell licensing: A multistep process. Leukemia 2011, 25, 1408–1414. [Google Scholar] [CrossRef]
- Wada, N.; Menicanin, D.; Shi, S.; Bartold, P.M.; Gronthos, S. Immunomodulatory properties of human periodontal ligament stem cells. J. Cell. Physiol. 2009, 219, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Takegawa, D.; Nakanishi, T.; Hirao, K.; Yumoto, H.; Takahashi, K.; Matsuo, T. Modulatory roles of interferon-γ through indoleamine 2, 3-dioxygenase induction in innate immune response of dental pulp cells. J. Endod. 2014, 40, 1382–1387. [Google Scholar] [CrossRef] [PubMed]
- Nisapakultorn, K.; Makrudthong, J.; Sa-Ard-Iam, N.; Rerkyen, P.; Mahanonda, R.; Takikawa, O. Indoleamine 2,3-dioxygenase expression and regulation in chronic periodontitis. J. Periodontol. 2009, 80, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Calton, E.K.; Keane, K.N.; Newsholme, P.; Soares, M.J. The impact of Vitamin D levels on inflammatory status: A systematic review of immune cell studies. PLoS ONE 2015, 10, e0141770. [Google Scholar] [CrossRef]
- Pfeffer, P.E.; Lu, H.; Mann, E.H.; Chen, Y.H.; Ho, T.R.; Cousins, D.J.; Corrigan, C.; Kelly, F.J.; Mudway, I.S.; Hawrylowicz, C.M. Effects of Vitamin D on inflammatory and oxidative stress responses of human bronchial epithelial cells exposed to particulate matter. PLoS ONE 2018, 13, e0200040. [Google Scholar] [CrossRef]
- Hausmann, J.; Kubesch, A.; Amiri, M.; Filmann, N.; Blumenstein, I. Vitamin D deficiency is associated with increased disease activity in patients with inflammatory bowel disease. J. Clin. Med. 2019, 8, 1319. [Google Scholar] [CrossRef]
- Yin, K.; Agrawal, D.K. Vitamin D and inflammatory diseases. J. Inflamm. Res. 2014, 7, 69. [Google Scholar]
- Mousa, A.; Naderpoor, N.; Johnson, J.; Sourris, K.; De Courten, M.P.J.; Wilson, K.; Scragg, R.; Plebanski, M.; De Courten, B. Effect of Vitamin D supplementation on inflammation and nuclear factor kappa-B activity in overweight/obese adults: A randomized placebo-controlled trial. Sci. Rep. 2017, 7, 15154. [Google Scholar] [CrossRef]
- Chirumbolo, S.; Bjørklund, G.; Sboarina, A.; Vella, A. The role of Vitamin D in the immune system as a Pro-survival molecule. Clin. Ther. 2017, 39, 894–916. [Google Scholar] [CrossRef]
- Dietrich, T.; Nunn, M.; Dawson-Hughes, B.; Bischoff-Ferrari, H.A. Association between serum concentrations of 25-hydroxyvitamin D and gingival inflammation. Am. J. Clin. Nutr. 2005, 82, 575–580. [Google Scholar] [CrossRef]
- Dietrich, T.; Joshipura, K.J.; Dawson-Hughes, B.; Bischoff-Ferrari, H.A. Association between serum concentrations of 25-hydroxyvitamin D 3 and periodontal disease in the US population. Am. J. Clin. Nutr. 2004, 80, 108–113. [Google Scholar] [PubMed]
- Liu, K.; Meng, H.; Tang, X.; Xu, L.; Zhang, L.; Chen, Z.; Shi, D.; Feng, X.; Lu, R. Elevated plasma calcifediol is associated with aggressive periodontitis. J. Periodontol. 2009, 80, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Pan, Y.; Zhao, Y. Vitamin D inhibits the expression of interleukin-8 in human periodontal ligament cells stimulated with Porphyromonas gingivalis. Arch. Oral Biol. 2013, 58, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.; Hosokawa, I.; Shindo, S.; Ozaki, K.; Matsuo, T. Calcitriol Suppressed Inflammatory Reactions in IL-1β-Stimulated Human Periodontal Ligament Cells. Inflammation 2015, 38, 2252–2258. [Google Scholar] [CrossRef]
- Nebel, D.; Svensson, D.; Arosenius, K.; Larsson, E.; Jönsson, D.; Nilsson, B.O. 1α,25-dihydroxyvitamin D3 promotes osteogenic activity and downregulates proinflammatory cytokine expression in human periodontal ligament cells. J. Periodontal Res. 2015, 50, 666–673. [Google Scholar] [CrossRef]
- Andrukhov, O.; Andrukhova, O.; Özdemir, B.; Haririan, H.; Müller-Kern, M.; Moritz, A.; Rausch-Fan, X. Soluble CD14 enhances the response of periodontal ligament stem cells to P. gingivalis lipopolysaccharide. PLoS ONE 2016, 11, e0160848. [Google Scholar] [CrossRef]
- Behm, C.; Blufstein, A.; Gahn, J.; Noroozkhan, N.; Moritz, A.; Rausch-Fan, X.; Andrukhov, O. Soluble CD14 enhances the response of periodontal ligament stem cells to toll-like receptor 2 agonists. Mediat. Inflamm. 2019, 2019, 8127301. [Google Scholar] [CrossRef]
- Andrukhov, O.; Hong, J.S.-A.; Andrukhova, O.; Blufstein, A.; Moritz, A.; Rausch-Fan, X. Response of human periodontal ligament stem cells to IFN-gamma and TLR-agonists. Sci. Rep. 2017, 7, 12856. [Google Scholar] [CrossRef]
- Stein, S.H.; Livada, R.; Tipton, D.A. Re-evaluating the role of vitamin D in the periodontium. J. Periodontal Res. 2014, 49, 545–553. [Google Scholar] [CrossRef]
- Ceuppens, J.L.; Baroja, M.L.; Lorre, K.; Van Damme, J.; Billiau, A. Human T cell activation with phytohemagglutinin. The function of IL-6 as an accessory signal. J. Immunol. 1988, 141, 3868–3874. [Google Scholar]
- Usherwood, E.J.; Crowther, G.; Woodland, D.L. Apoptotic cells are generated at every division of in vitro cultured T cell lines. Cell. Immunol. 1999, 196, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, P.; Govindasamy, V.; Gnanasegaran, N.; Kunasekaran, W.; Vasanthan, P.; Musa, S.; Kasim, N.H.A. Expression patterns of immune genes in long-term cultured dental stem cells. Clin. Oral Investig. 2016, 20, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Engebretson, S.P.; Hey-Hadavi, J.; Ehrhardt, F.J.; Hsu, D.; Celenti, R.S.; Grbic, J.T.; Lamster, I.B. Gingival crevicular fluid levels of interleukin-1β and glycemic control in patients with chronic periodontitis and type 2 diabetes. J. Periodontol. 2004, 75, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Dutzan, N.; Vernal, R.; Hernandez, M.; Dezerega, A.; Rivera, O.; Silva, N.; Aguillon, J.C.; Puente, J.; Pozo, P.; Gamonal, J. Levels of interferon-gamma and transcription factor t-bet in progressive periodontal lesions in patients with chronic periodontitis. J. Periodontol. 2009, 80, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Daĝ, A.; Firat, E.T.; Kadiroĝlu, A.K.; Kale, E.; Yilmaz, M.E. Significance of elevated gingival crevicular fluid tumor necrosis factor-α and interleukin-8 levels in chronic hemodialysis patients with periodontal disease. J. Periodontal Res. 2010, 45, 445–450. [Google Scholar] [PubMed]
- Teng, Y.T.A. Protective and destructive immunity in the periodontium: Part 2—T-cell-mediated immunity in the periodontium. J. Dent. Res. 2006, 85, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Hasturk, H.; Kantarci, A. Activation and resolution of periodontal inflammation and its systemic impact. Periodontology 2000 2015, 69, 255–273. [Google Scholar] [CrossRef]
- Gao, Z.; Liu, K.; Meng, H. Preliminary investigation of the vitamin D pathway in periodontal connective tissue cells. J. Periodontol. 2018, 89, 294–302. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behm, C.; Blufstein, A.; Gahn, J.; Kubin, B.; Nemec, M.; Moritz, A.; Rausch-Fan, X.; Andrukhov, O. 1,25(OH)2D3 Differently Affects Immunomodulatory Activities of Mesenchymal Stem Cells Depending on the Presence of TNF-α, IL-1β and IFN-γ. J. Clin. Med. 2019, 8, 2211. https://doi.org/10.3390/jcm8122211
Behm C, Blufstein A, Gahn J, Kubin B, Nemec M, Moritz A, Rausch-Fan X, Andrukhov O. 1,25(OH)2D3 Differently Affects Immunomodulatory Activities of Mesenchymal Stem Cells Depending on the Presence of TNF-α, IL-1β and IFN-γ. Journal of Clinical Medicine. 2019; 8(12):2211. https://doi.org/10.3390/jcm8122211
Chicago/Turabian StyleBehm, Christian, Alice Blufstein, Johannes Gahn, Barbara Kubin, Michael Nemec, Andreas Moritz, Xiaohui Rausch-Fan, and Oleh Andrukhov. 2019. "1,25(OH)2D3 Differently Affects Immunomodulatory Activities of Mesenchymal Stem Cells Depending on the Presence of TNF-α, IL-1β and IFN-γ" Journal of Clinical Medicine 8, no. 12: 2211. https://doi.org/10.3390/jcm8122211
APA StyleBehm, C., Blufstein, A., Gahn, J., Kubin, B., Nemec, M., Moritz, A., Rausch-Fan, X., & Andrukhov, O. (2019). 1,25(OH)2D3 Differently Affects Immunomodulatory Activities of Mesenchymal Stem Cells Depending on the Presence of TNF-α, IL-1β and IFN-γ. Journal of Clinical Medicine, 8(12), 2211. https://doi.org/10.3390/jcm8122211