1. Introduction
Tuberculosis (TB), caused by
Mycobacterium tuberculosis (M. tb), continues to afflict millions of people worldwide. In 2017, approximately 10 million people suffered from active TB and 1.6 million died from this disease [
1]. Additionally, one third of the world’s population is latently infected with
M. tb. Individuals infected with
M. tb have a 5–15% lifetime risk of developing an active disease; however, immunocompromised patients, such as people living with diabetes, malnutrition, human immunodeficiency virus (HIV), or those who use tobacco, have a higher risk of developing active TB [
1]. Common symptoms of active pulmonary TB are coughs with a bloody sputum, chest pains, weakness, weight loss, fever, and night sweats, eventually resulting in death when untreated [
1].
M. tb infection occurs due to inhalation of infectious aerosolized droplets, and the bacteria become seeded in the lower respiratory tract where there is an enrichment of alveolar macrophages.
M. tb infection is initiated when the inhaled organisms are phagocytosed by these alveolar macrophages [
2]. In an immune-competent individual, the immune system is able to mount a formidable response against
M. tb, resulting in the formation of a solid and robust granuloma. Composed of a compact aggregate of immune cells [
3]. Mature macrophages in the granuloma can fuse into multinucleated giant cells or differentiate into foam cells and epithelioid cells [
3]. Alongside macrophages, other cells, such as neutrophils, dendritic cells, natural killer cells, fibroblasts, CD4
+ T cells, and cytotoxic CD8
+ T cells, are also recruited into the granuloma via cytokine mediation, leading to containment of the
M. tb infection [
3]. The effector responses inside the granulomas along with a lack of nutrients and oxygen causes
M. tb to become dormant and remain latent in a nonreplicating state. The contained
M. tb within a granuloma in the lungs is commonly referred to as latent tuberculosis (LTBI). A breakdown of immune responses designed to contain the infection in immunocompromised individuals can result in reactivation of
M. tb [
4]. This dysregulation promotes liquification of caseum and replication of
M. tb, thereby promoting cavity formation and the release of
M. tb to the exterior during coughing, ultimately spreading the infection to other parts of the lungs [
5]. Active
M. tb is able to deflect many host defense mechanisms via the cord factor, preventing phagosome-lysosome fusion and the degradation of the bacilli [
6].
In order to effectively contain
M. tb within the granuloma, proper cytokine-mediated signaling is essential to promote the necessary aggregation of cells [
7]. Cytokines, such as interferon gamma (IFN-γ) and tumor necrosis factor-α (TNF-α), play a critical role in both the innate and adaptive immune responses against
M. tb infection [
8]. TNF-α produced by macrophages induces the formation and maintenance of the granuloma. The T-helper 1 (Th1) subset of CD4
+ T cells releases IFN-γ to activate effector mechanisms in macrophages to not only kill
M. tb intracellularly but also enhance the effector functions of natural killer cells and cytotoxic T lymphocytes (CD8
+) T-cells [
9].
Once activated, CD8+ T cells and natural killer cells will then produce antimicrobial peptides, perforin and granulysin, to destroy intracellular M. tb and the host cells.
The programmed death receptor 1 (PD-1), a negative regulator of activated T cells, is markedly upregulated on the surface of pathogen-specific CD8
+ T cells in mice [
10]. Blockage of this pathway restores the CD8
+ T cell function and reduces the microbial load [
10]. PD-1 is also expressed on the surface of CD4
+ T cells, with a positive correlation in regard to the microbial load and an inverse correlation with the CD4
+ T cell count.
Although the immune system has a robust defense system in place to combat M. tb infection, it cannot always contain the infection. For this reason, host directed therapy is often required.
The Center for Disease Control (CDC) recommends four anti-TB agents to form the core of the treatment regimen for patients with active TB [
11]. These drugs include isoniazid (INH), rifampicin (RIF), ethambutol (EMB), and pyrazinamide (PZA). This extensive drug regimen often leads to noncompliance to the TB treatment, leading to multidrug-resistant (MDR)-TB, which is typically resistant to both INH and RIF [
12].
Although a myriad of antibiotics can be used to treat this mycobacterial infection, there is only one vaccine available to prime the immune system against
M. tb, and that is the
Mycobacterium bovis bacille Calmette–Guérin (BCG) vaccine [
13]. BCG used in this vaccine is an attenuated strain of
M. bovis [
13]. The WHO recommends that infants in countries with a high risk of
M. tb infection be immunized with the BCG vaccine soon after birth [
13]. Because the incidence of TB is low in the United States, it is not recommended for infants to be administered this vaccination. Additionally, estimates of the protective efficacy of the BCG vaccine against adult pulmonary TB very widely, ranging from 0 to 80% [
13].
Glutathione (GSH), a tripeptide antioxidant composed of glutamine, cysteine, and glycine, is found ubiquitously amongst all cell types. GSH prevents cellular damage by detoxifying reactive oxygen species (ROS) [
14]. GSH exists in both a reduced state (rGSH) and an oxidized form (GSSG) [
14]. rGSH contains the antioxidant properties, while GSSG is a simple byproduct of the oxidation of GSH and has no antioxidant effects [
14]. When exposed to ROS, two molecules of rGSH are converted to GSSG and water [
14]. Mycobacteria possess an alternative thiol, mycothiol, rather than GSH, to regulate their redox homeostasis [
15]. Due to this property, the presence of millimolar concentrations of GSH (physiological concentrations) inside infected macrophages can lead to inhibition in the growth of
M. tb [
15]. Our laboratory has previously demonstrated that GSH-enhancement by N-acetyl cysteine (NAC) supplementation resulted in a significant reduction of
M. tb burden among both healthy and diabetic individuals [
15]. Additionally, enhancement of GSH by means of NAC has the potential implications of not only reducing the toxicity of anti-TB medications through GSH’s redox potential but may possibly permit lower antibiotic dosage to promote enhanced patient compliance [
15]. For this reason, our lab tested the effects of liposomal glutathione (L-GSH) in the presence and absence of sub-optimal concentration of INH and PZA in improving the ability of immune cells isolated from BCG-vaccinated and non-vaccinated individuals to control
M. tb infection.
In this study, we determined the additive effects of BCG vaccination and in vitro GSH-enhancement in improving the ability of immune cells to control M. tb infection by measuring the differences in the immune responses between vaccinated and non-vaccinated individuals. Fluorescent staining and other antibody assays were also performed to determine the underlying mechanistic differences in the ability of immune cells from vaccinated and unvaccinated groups to respond to L-GSH, cytokine production, the surface expression of CD4, CD8, PD-1, and their ability to induce autophagy.
2. Materials and Methods
2.1. Peripheral Blood Mononuclear Cell Isolation
Peripheral blood mononuclear cells (PBMCs) were isolated from the whole blood of both BCG-vaccinated and non-BCG-vaccinated participants. Whole blood was layered in a 1:1 ratio onto ficoll histopaque (Sigma, St. Louis, MO, USA), a high density-pH neutral polysaccharide solution, for density gradient centrifugation (1800 rpm for 30 min) [
15]. The PBMCs at the interface were aspirated, washed twice with sterile 1X PBS (Sigma, St. Louis, MO, USA), and resuspended in Roswell Park Memorial Institute (RPMI) (Sigma, St Louis, MO, USA) with 5% human AB serum (Sigma, St. Louis, MO, USA). PBMC counts were determined by trypan blue exclusion staining.
2.2. Generation of In Vitro Granulomas
Our laboratory had successfully established an
in vitro human granuloma model using PBMCs, isolated from healthy subjects and individuals with type 2 diabetes [
15,
16,
17,
18]. These granulomas [
15,
16,
17,
18] exhibit a physically well-demarcated aggregation of mononuclear cells with a denser central core descending towards the periphery, which can be seen in the in vitro granulomas. Multi-nucleated giant cells (MNGs hallmark of granulomas), T cells, and activated macrophages were also seen. These features are reminiscent of early stage, cellular lung granulomas in experimental animal models of TB, including rabbits and non-human primates. Such granulomas are also noted in the lungs of mice during chronic
M. tb infection. Using our previously published protocol, we developed in vitro granulomas for the current study. Isolated PBMCs from the two study groups resuspended in RPMI were infected with the Erdman strain of
M. tb at a multiplicity of infection (MOI) of 0.1:1 cell ratio. 500 µL of the cell suspension containing PBMCs and
M. tb were added to the 24-well plates. To ensure proper adhesion of isolated immune cells, 24-well plates (Corning, Corning, NY, USA) were coated with 0.001% poly-lysine (Sigma, St. Louis, MO, USA) overnight [
15,
16]. PBMCs (6 × 10
5 cells/well) were distributed into the poly-lysine coated 24-well plates [
15,
16]. PBMCs in the wells were either sham-treated or treated with the minimum inhibitory concentration (MIC), a 1.10 dilution, and a 1.100 dilution of two first-line antibiotics with and without 120 µM of liposomal glutathione (L-GSH (Your Energy Systems)). This comprised INH (0.125 micrograms/mL) standalone, 120 µM of L-GSH, 1/10 INH (0.0125 micrograms/mL) standalone, 120 µM of L-GSH, 1/100 INH (0.00125 micrograms/mL) standalone, 120 µM of L-GSH, PZA (50 micrograms/mL), 120 µM of L-GSH, 1/10 PZA (5 micrograms/mL) standalone, or 120 µM of L-GSH and 1/100 PZA (0.5 micrograms/mL) standalone or 120 µM of L-GSH. All tissue culture plates with infected PBMCs were maintained at 37 °C with 5% CO
2 until they were terminated at 8 days post infection.
2.3. Termination of Granulomas
Following 8 days post infection, the minimum time needed for granuloma formation,
in vitro granulomas were terminated to determine the intracellular survival of
M. tb [
15,
16]. To terminate, the supernatants of each category were aspirated and collected into eppendorf tubes separated by a treatment group, and 250 µL of ice cold, sterile 1× PBS was replaced in lieu of the supernatants followed by gentle scraping of the wells [
15,
16]. Scraping was done to ensure maximum recovery of granuloma lysates from the wells.
2.4. Colony Forming Units
Collected supernatants and lysates from termination were plated on 7H11 agar media (Hi Media, Santa Maria, CA, USA) enriched with Albumin Dextrose Complex (ADC) (GEMINI, Calabasas, CA, US). They were incubated for a minimum of 3 weeks and 3 days to evaluate the mycobacterial survival under the different treatment conditions by counting the colony forming units (CFUs).
2.5. Cytokine Measurements
To measure cytokine levels, the sandwich enzyme-linked immunosorbent assay (ELISA) technique was used. The assay was performed via the manufacturer’s protocol (Invitrogen, Carlsbad, CA, USA). The cytokines measured were IFN-γ and TNF- in the supernatants at 8 days post-infection to determine the effects of the antibiotics with and without L-GSH treatments on cytokine levels in BCG-vaccinated and non-vaccinated individuals.
2.6. Glutathione Measurements
Levels of GSH from the granulomas of non-vaccinated and BCG-vaccinated subjects were measured by the colorimetric method using an assay kit from Arbor Assay (K006-H1). Granuloma lysates were mixed in a 1:1 ratio with cold 5% sulfosalicylic acid (SSA), incubated for 10 min at 4 °C, followed by centrifugation at 14,000 rpm for 10 min. The GSH was measured in the lysates following the manufacturer’s instructions. The reduced GSH (rGSH) was calculated by subtracting the oxidized glutathione (GSSG) from the total GSH.
2.7. Staining and Imaging Techniques
Each trial contained designated wells for fluorescent and light microscopic studies. Cover glasses were allotted into 24-well plates for granuloma formation observation. The cover glasses were fixed with 4% paraformaldehyde (PFA) for 1 h at room temperature and washed three times with 1× PBS for 5 min to remove cell debris. Fixed granulomas were then stained with Hematoxylin and Eosin (H&E) (Poly Scientific, Bay Shore, NY, USA) for 2 min at room temperature and destained with deionized water. The granuloma-stained cover glasses were mounted onto glass slides with HistoChoice mounting media. Fixed granulomas on cover glasses were also permeabilized with Triton X for 2 min and stained overnight with antibodies conjugated with fluorescent markers (CD4-PE, CD8-PE, and LC3B-PE). Cover glasses were washed with phosphate buffer saline (PBS) and mounted on clean glass slides with mounting media containing 4’,6-diamidino-2-phenylindole DAPI For PD1 staining, fixed granulomas on cover glasses were permeabilized with Triton X for 2 min and incubated overnight with anti-PD1 (Pro-Sci), followed by incubation for another 2 h with c-Myc. Cover glasses were then incubated overnight with secondary antibodies (mouse anti-human) and conjugated with fluorescein isothiocyanate (FITC). Cover glasses were mounted using a mounting media containing DAPI. Slides were observed under the fluorescent microscope. Fluorescent images were captured, and the fluorescent intensity was quantified using the ImageJ software (version 8, GraphPad, San Diego, CA, USA).
2.8. Statistical Analysis
Statistical data analysis was performed using GraphPad Prism Software 8 using the unpaired t-test with Welch correction for two sampled graphs. A one-way ANOVA (analysis of variance) was performed for samples with greater than two categories with Tukey corrections. Reported values are the means with each respective category. A p < 0.05 was considered significant. The p value style consisted of 0.1234 as not significant, 0.0332 with one asterisk (*), 0.0021 with two asterisks (**), 0.0002 with three asterisks (***), and less than 0.0001 with four asterisks (****). A hash mark (#) indicates categories compared to control, and an asterisk indicates categories compared to the previous category directly before it.
4. Discussion
Using in vitro granulomas derived from PBMCs, this study compares the differences in the immune responses against M. tb in both BCG-vaccinated and non-vaccinated subjects. The effects of GSH enhancement and the first line antibiotics (INH and PZA) in further enhancing the ability of immune cells to control M. tb infection in both the groups was also evaluated. This study also focuses on the mechanistic actions of PBMC-derived in vitro granuloma-like structures infected with the Erdman strain of M. tb from non-vaccinated subjects and BCG-vaccinated subjects to illustrate their immune response in a regulated environment. Our in vitro granulomas constituted cell types, such as macrophages, monocytes, and CD4 and CD8 T cells, all of which contribute to the immune responses necessary for granuloma formation.
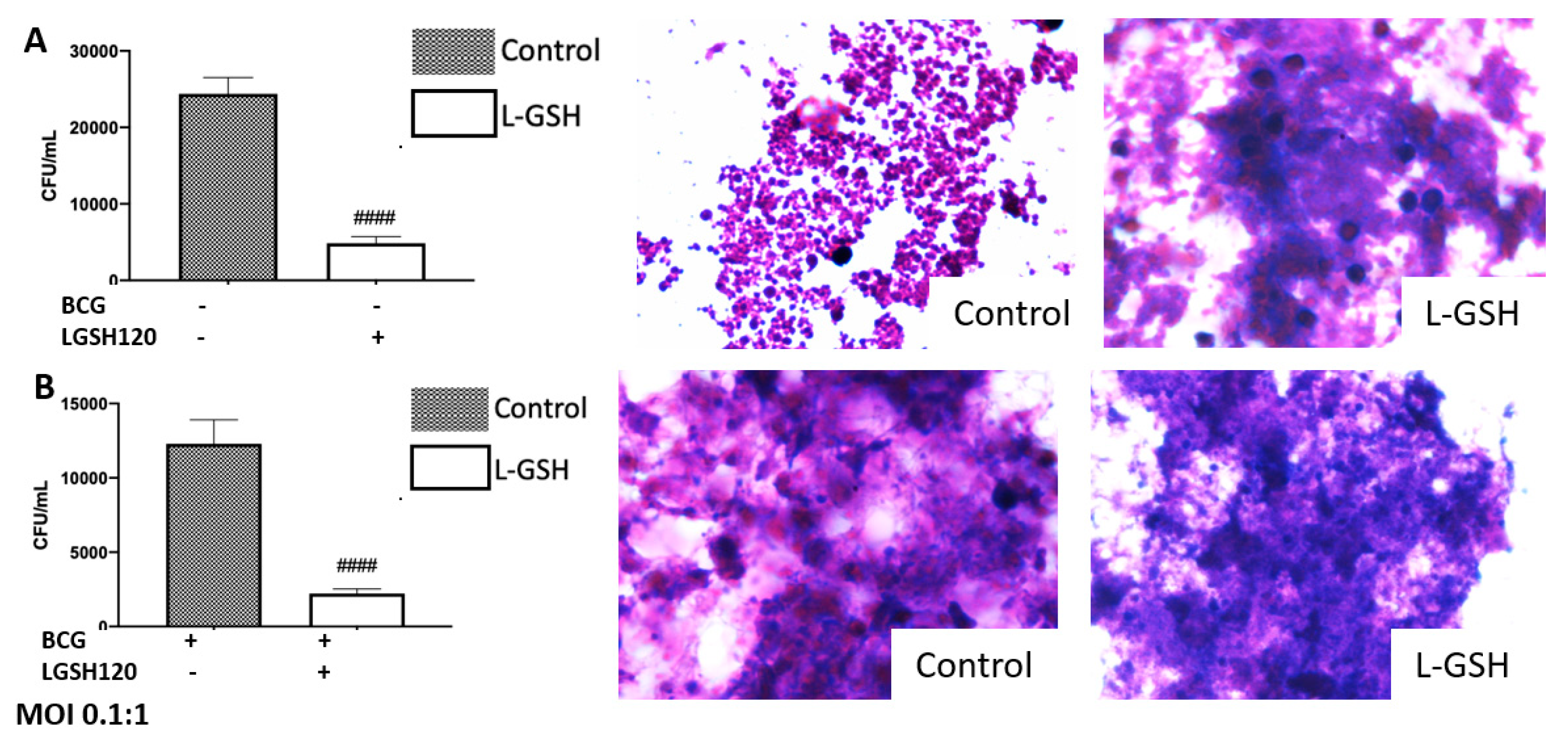
We observed an active replication of
M. tb to in the sham-treated granulomas, from the non-vaccinated group (
Figure 1A). Furthermore, the addition of L-GSH led to a significant reduction in the viability of
M. tb (
Figure 1A). This confirms previous findings that indicate that L-GSH possesses antimycobacterial and immune-enhancing abilities. Furthermore, H&E staining was performed on these granuloma-like structures to visualize the morphology of these structures. The absence of L-GSH in the sham-treated category led to a formation of a sparse array of a granuloma, while L-GSH addition to this sample formed a larger, more robust granuloma-like structure (
Figure 1A). This demonstrates the
M. tb containment ability of L-GSH (
Figure 1A). A similar trend was found in the BCG-vaccinated subjects. Notably, the mycobacterial count in the sham-treated control of the BCG-vaccinated group was 50% lower than the non-vaccinated subjects, and the addition of L-GSH lead to a further significant reduction in the bacterial load (
Figure 1B). H&E staining demonstrated that the sham-treated and L-GSH-treated categories resulted in dense, robust granuloma-like structures, indicating the ability of BCG-vaccinated individuals to mount a robust immune response to hinder
M. tb growth.
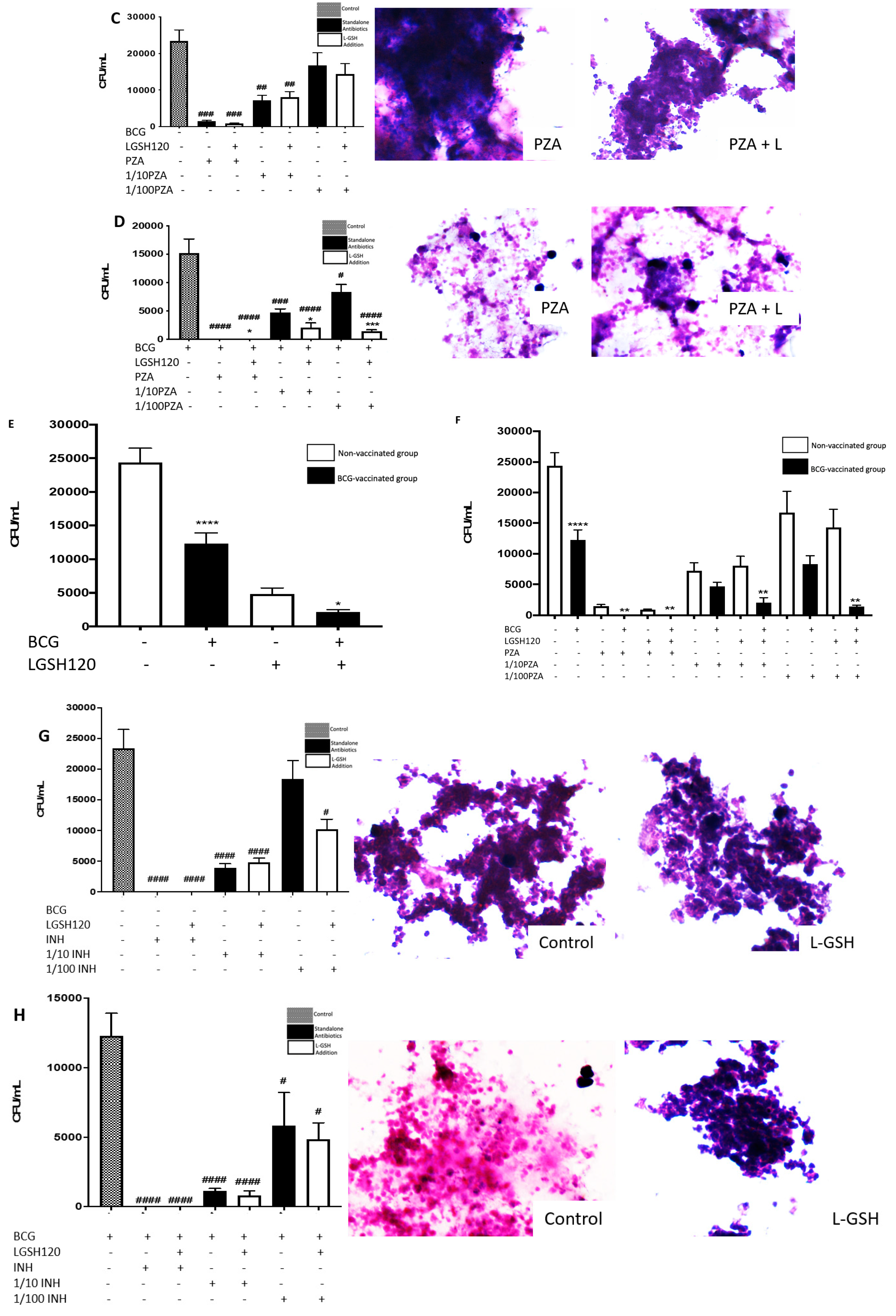
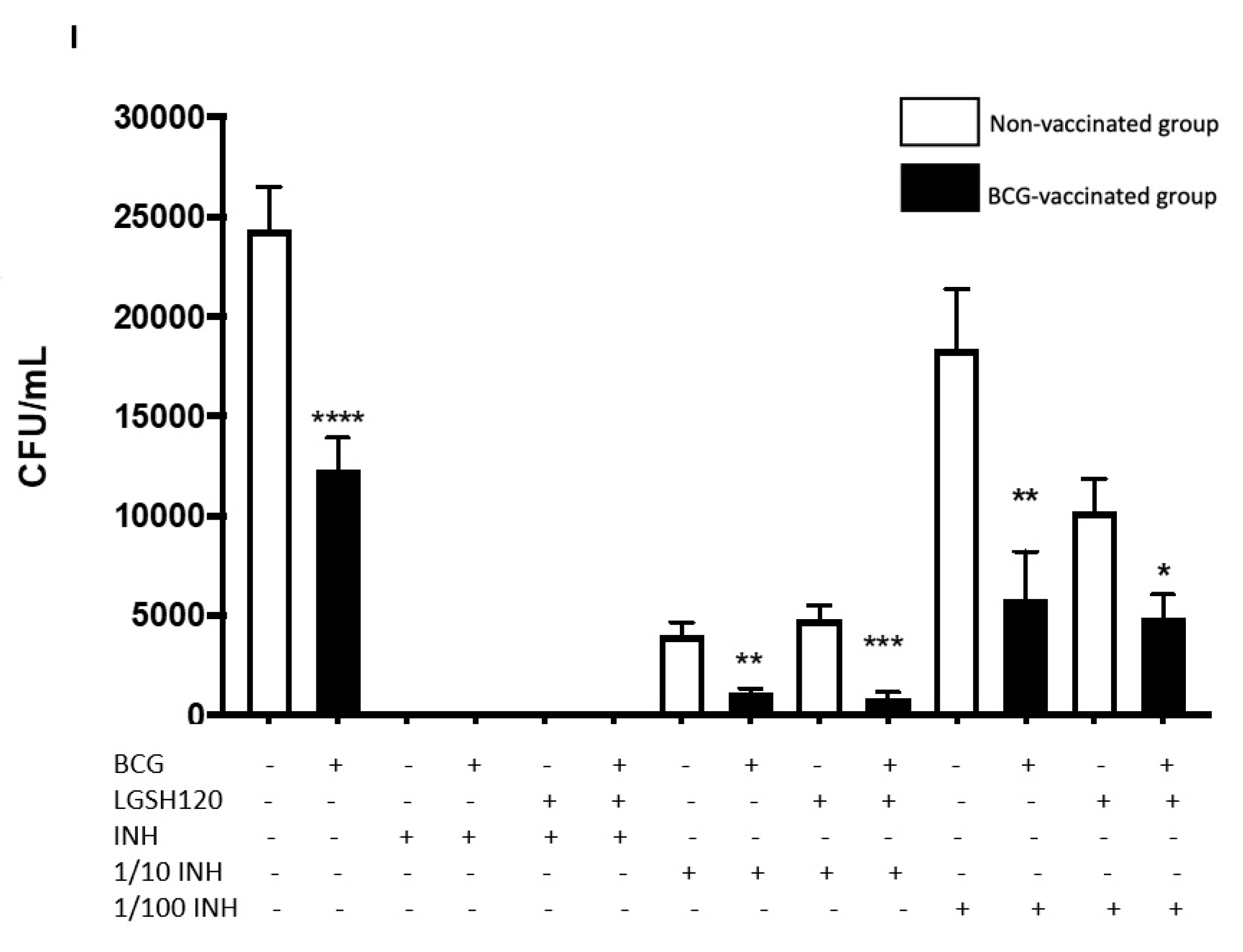
Findings from our study indicate that treatment of
in vitro granulomas with PZA + L-GSH and INH+L-GSH resulted in a significant reduction in the bacterial load in the BCG-vaccinated samples compared to the non-vaccinated samples (
Figure 1F,I). This evidence suggests that BCG vaccination enhances the ability of immune cells to effectively respond to GSH, PZA, and INH treatments, leading to containment and killing of
M. tb (
Figure 1E,F,I).
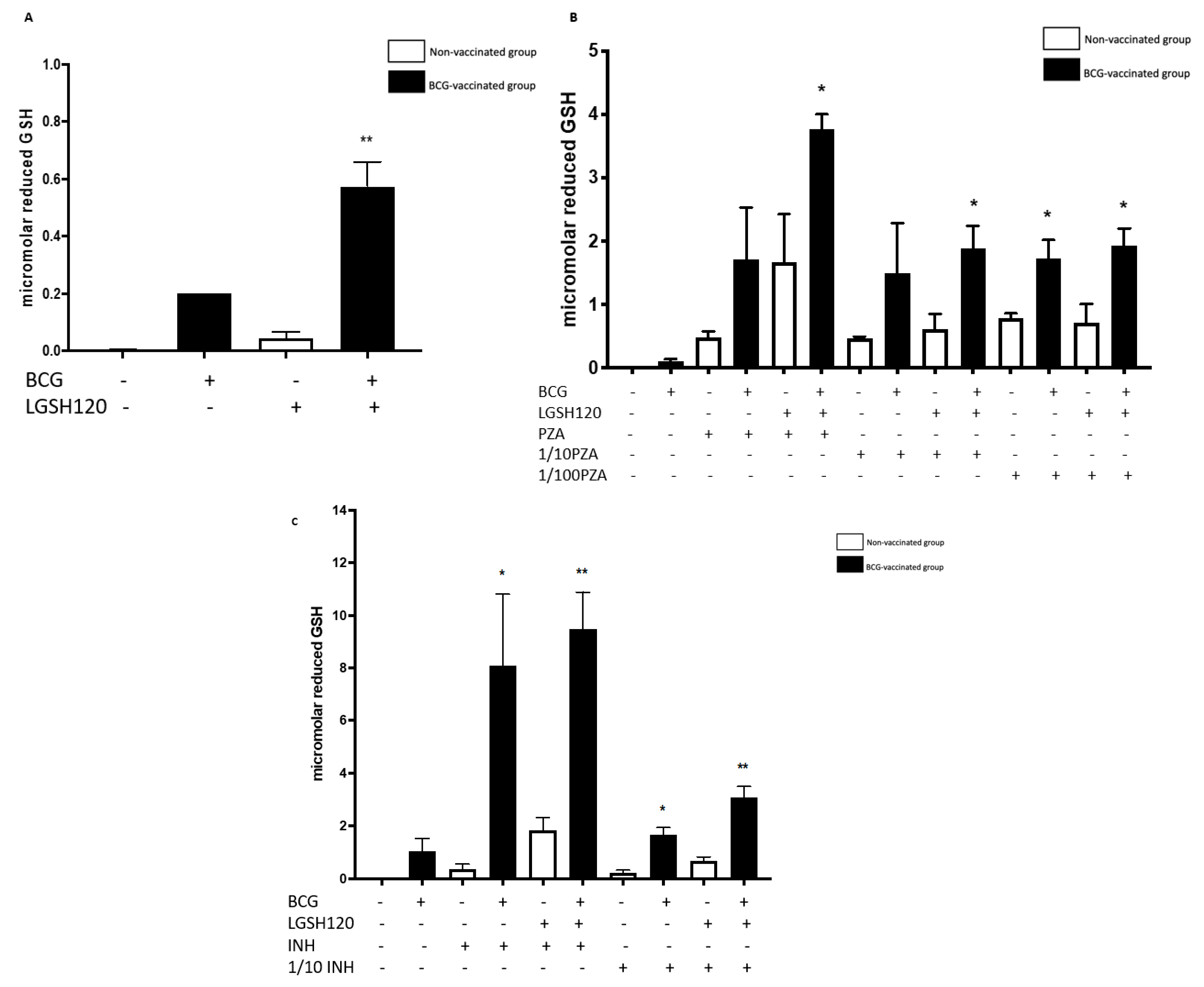
Our results also indicate that BCG-vaccinated individuals innately had higher levels of GSH when compared to the non-vaccinated group. Furthermore, L-GSH treatment resulted in a significant three log increase in the levels of GSH in BCG-vaccinated subjects when compared to non-vaccinated subjects (
Figure 2A). PZA or INH treatments resulted in a significant increase in the levels of GSH in the granulomas from the BCG-vaccinated group when compared to the non-vaccinated group (
Figure 2). Increased levels of GSH in the
in vitro granulomas of BCG-vaccinated subjects further illustrate GSH’s antimycobacterial activity, as indicated by low
M. tb numbers in the CFU assays in the BCG-vaccinated group in comparison to the non-vaccinated group.
Macrophages, CD4 and CD8 T cells play an important role in the control of
M. tb infection. Cytokines, such as TNF-α, interleukin-2 (IL-2), and IFN-γ, produced by macrophages and T cells can enhance the effector responses against
M. tb infection. In order to understand the immune effector mechanisms, by which BCG-vaccinated versus non-vaccinated subjects contain
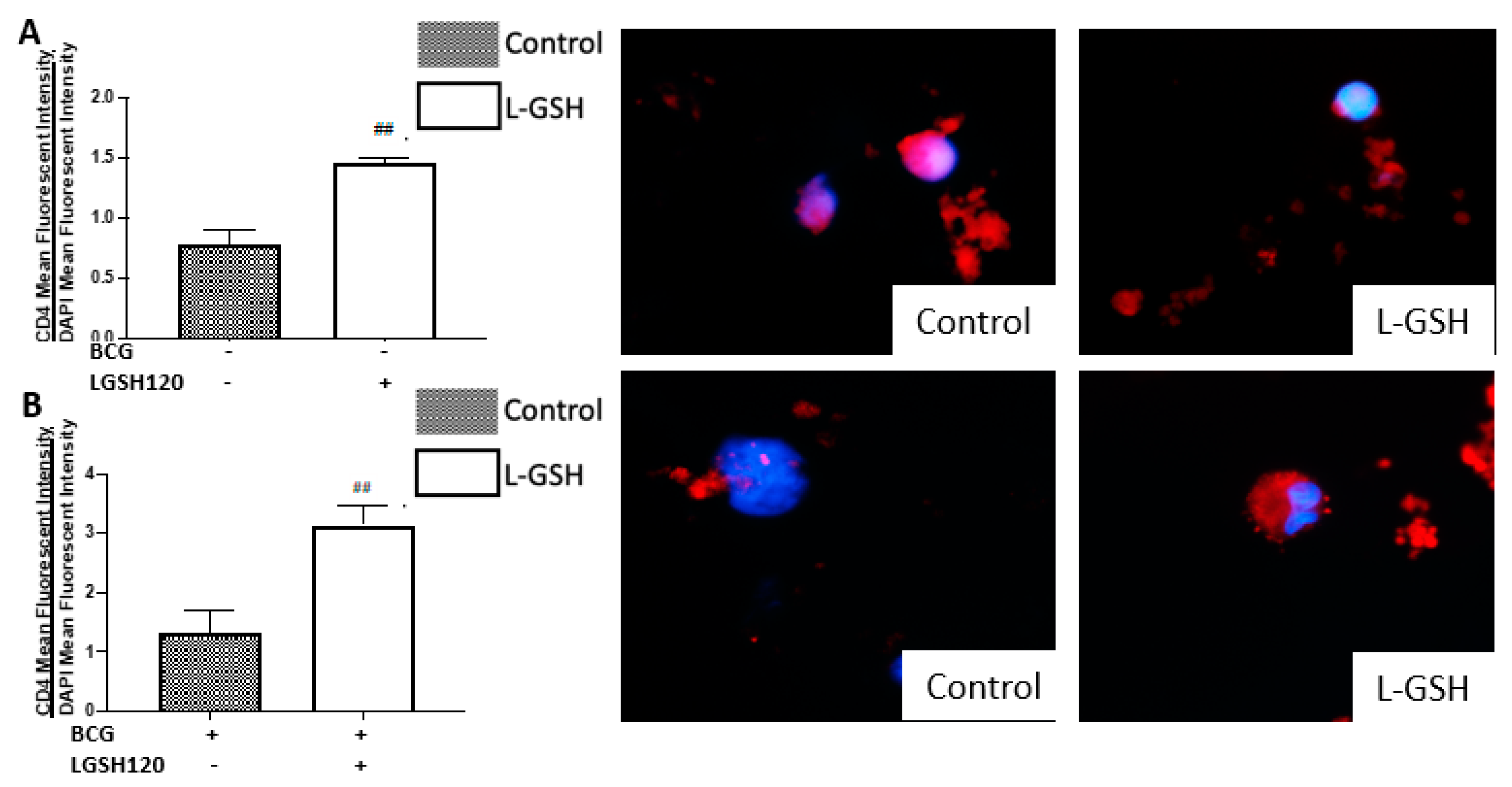
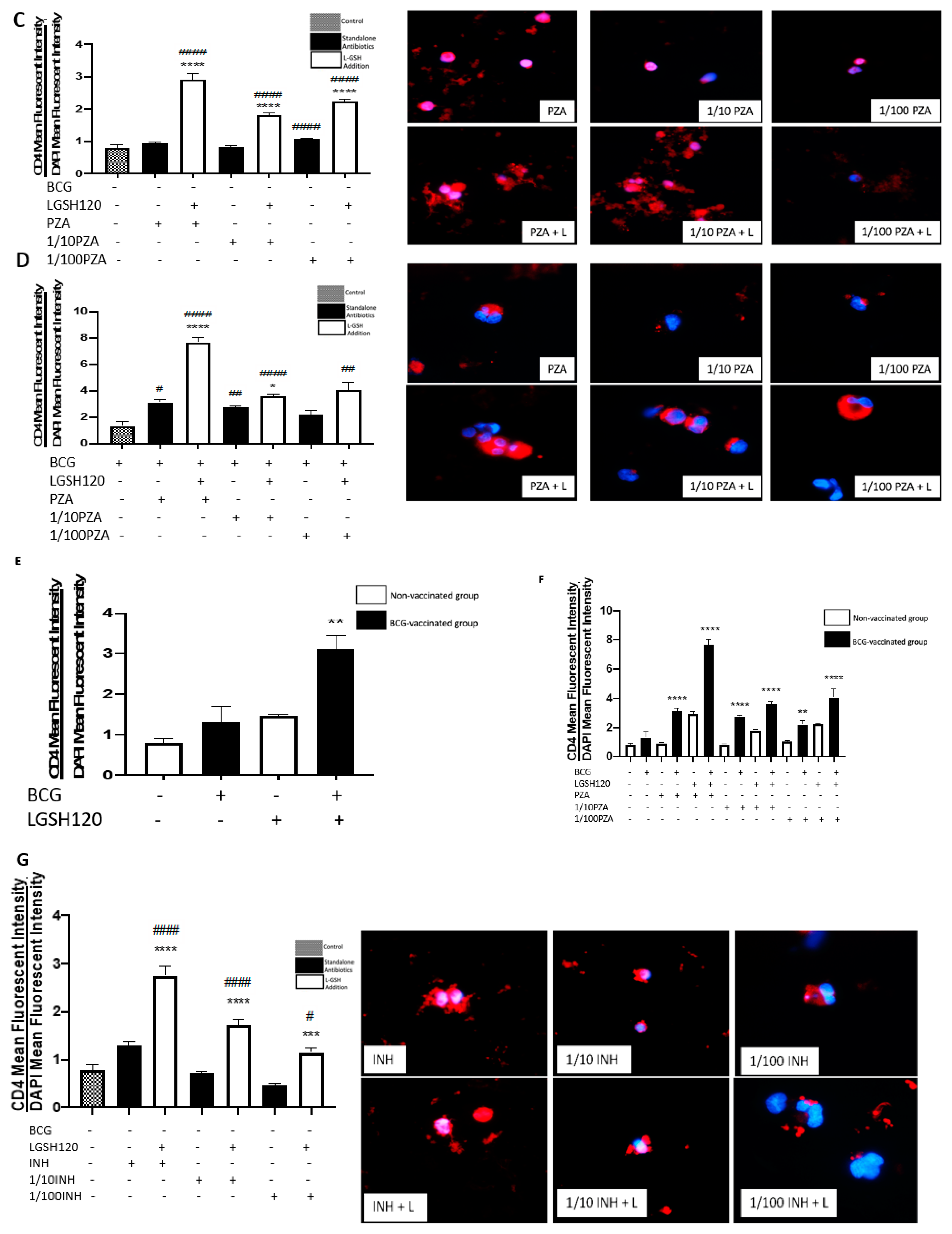
M. tb infection, we first measured the expression levels of CD4 in the granulomas. We initially observed the mean fluorescence of CD4 in the granulomas cultured in the presence and/or absence of L-GSH, PZA and PZA. We observed a dramatic, significant increase in the expression of CD4 in the granulomas from the BCG-vaccinated group with L-GSH addition (
Figure 3E), PZA addition at every concentration of the antibiotic in the presence and absence of L-GSH, and INH + L-GSH treatment at every concentration of the antibiotic (
Figure 3F,I), indicating the efficacy of L-GSH, PZA, and INH to maintain the viability CD4 T cells (
Figure 3). This evidence also demonstrates that there is less CD4 T cell exhaustion in BCG-vaccinated subjects.
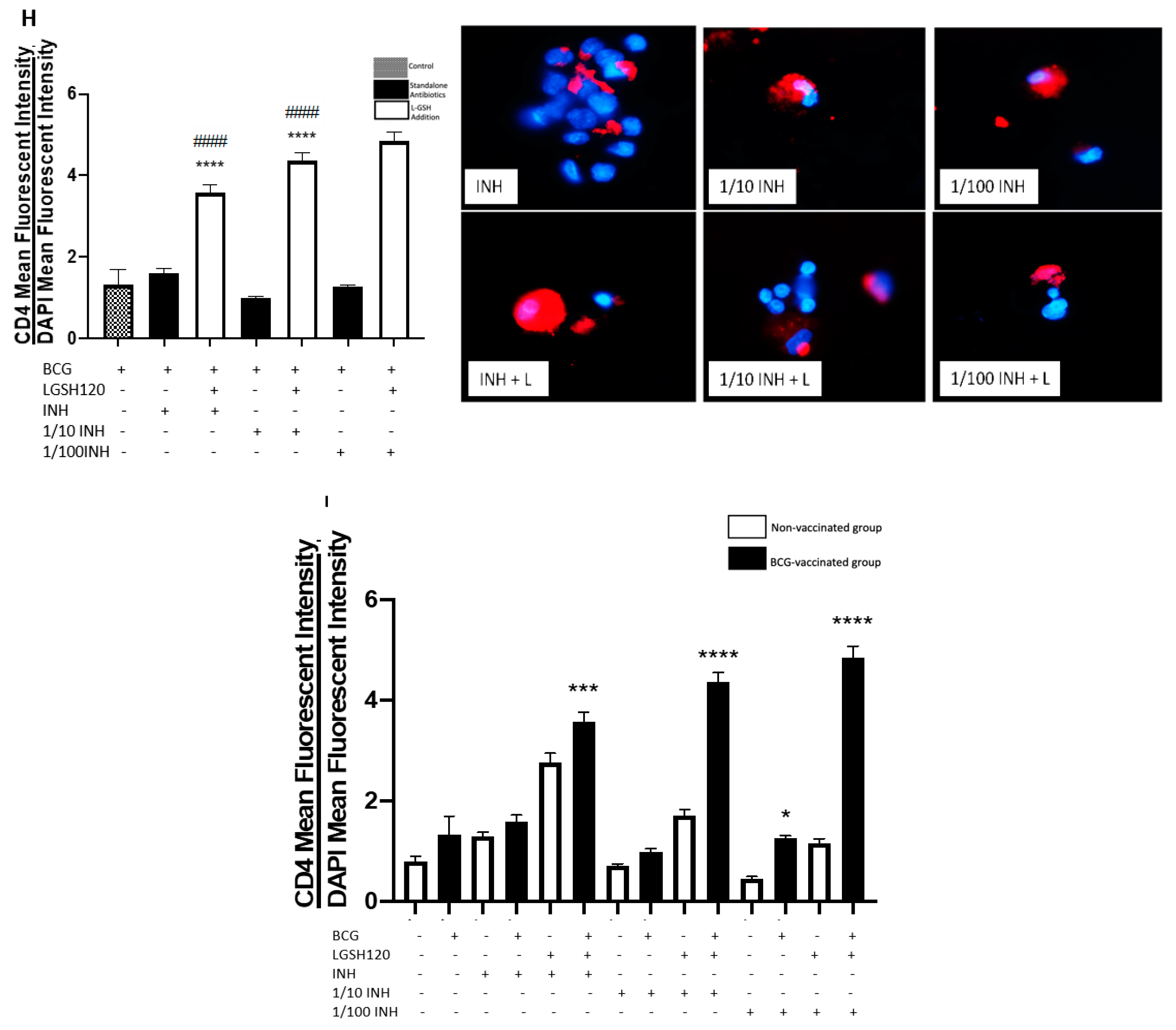
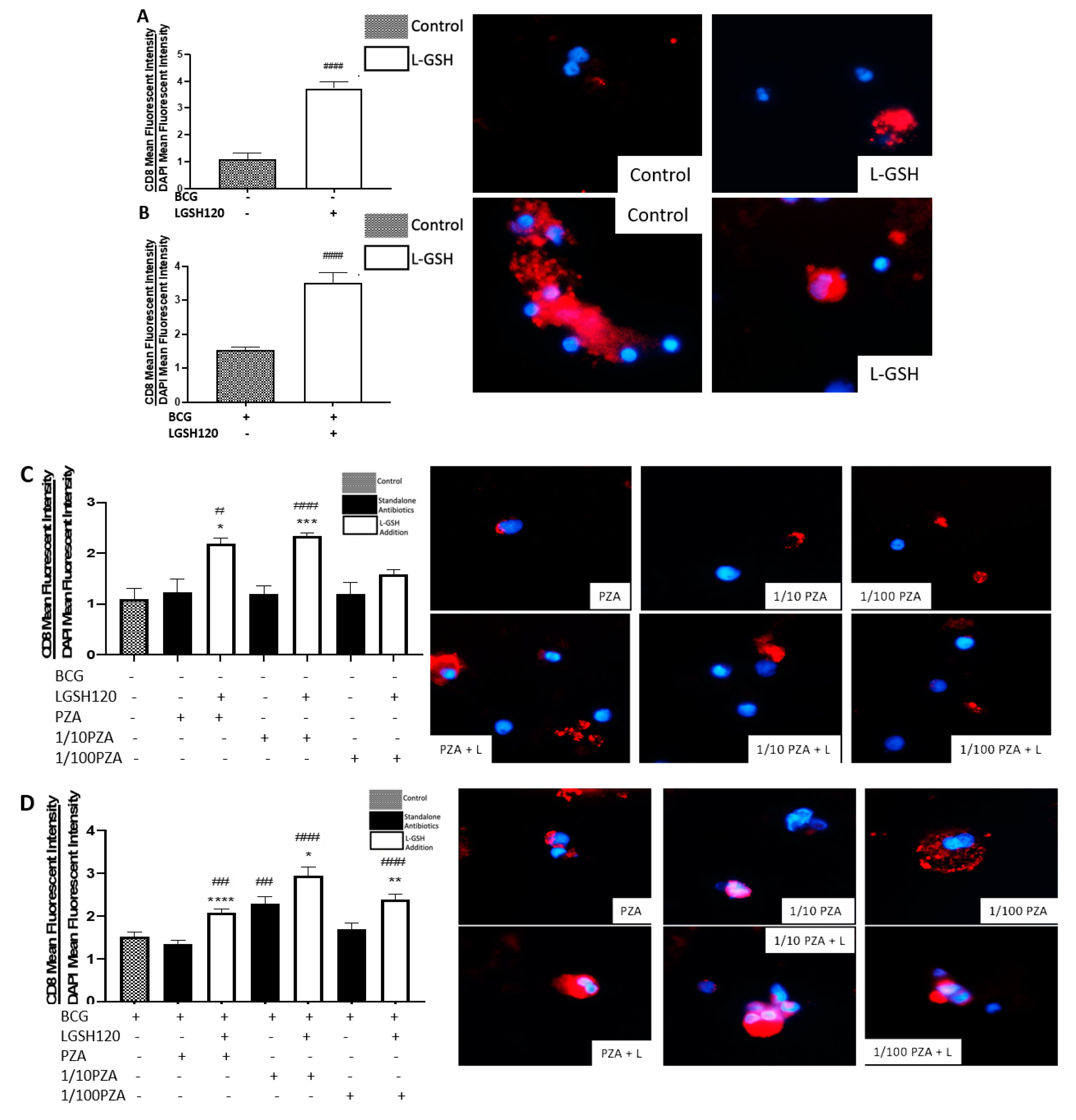
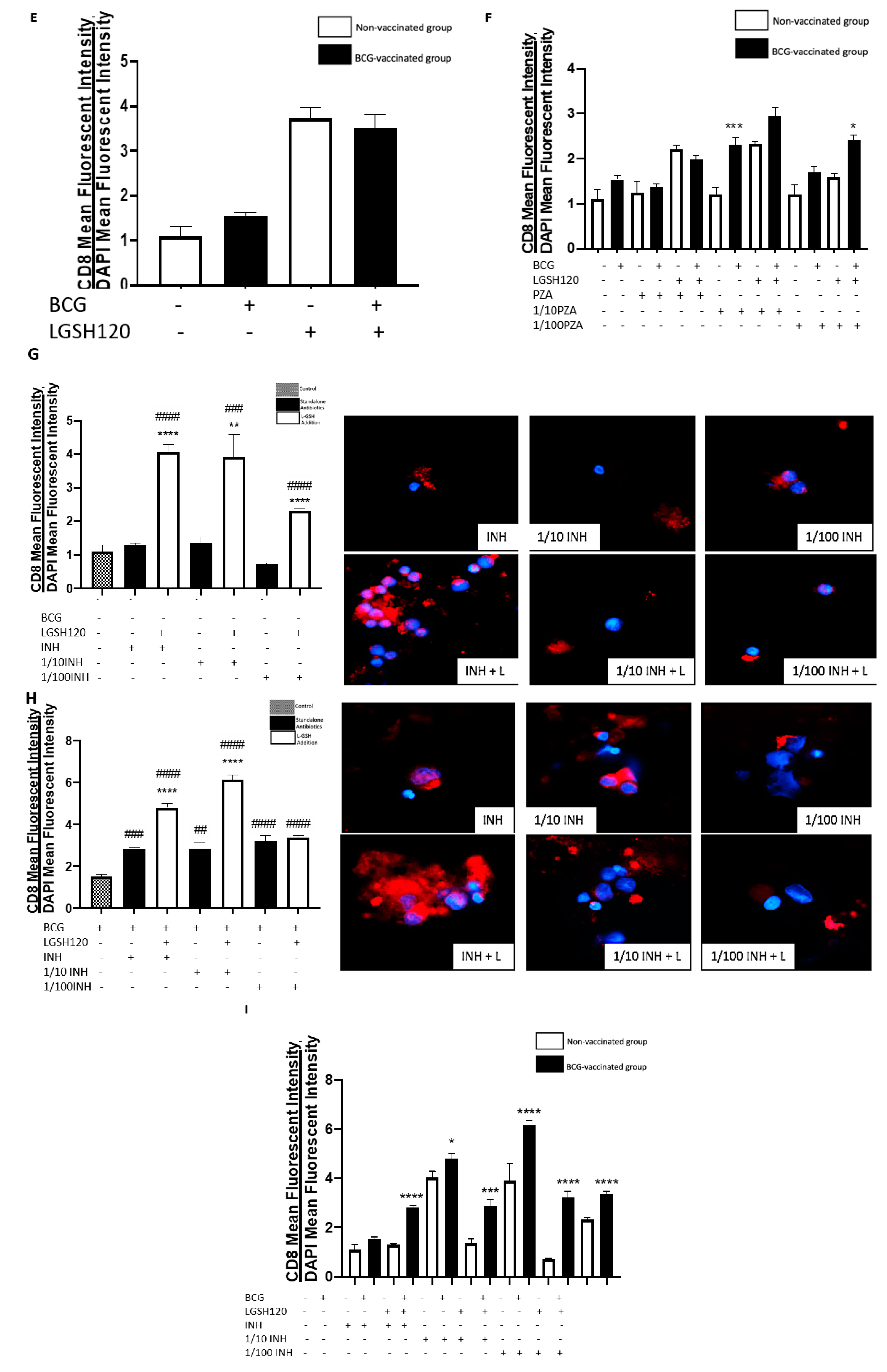
CD8
+ T cells have also been shown to produce IFN-γ that can augment effector functions to control
M. tb infection [
20,
21]. Importantly, CD8
+ T cells possess cytolytic and cytotoxic functions to kill
M. tb infected cells via a granule-mediated function (using perforin, granzymes, and granulysin). CD8 expression was determined in the granulomas of non-vaccinated and BCG-vaccinated subjects. Higher CD8 expression was observed following L-GSH treatment in the granulomas from both non-vaccinated and BCG-vaccinated subjects (
Figure 4E). The addition of the PZA and INH in the presence or absence of L-GSH resulted in an increase in the expression of CD8 in the BCG-vaccinated category when compared to the non-vaccinated group (
Figure 4F,I). Based on this general upward trend in the expression of CD8 in BCG-vaccinated subjects, we conclude that individuals who have been vaccinated with BCG have not only higher CD4 T cells in the presence of L-GSH, but also elevated levels of CD8 T cells.
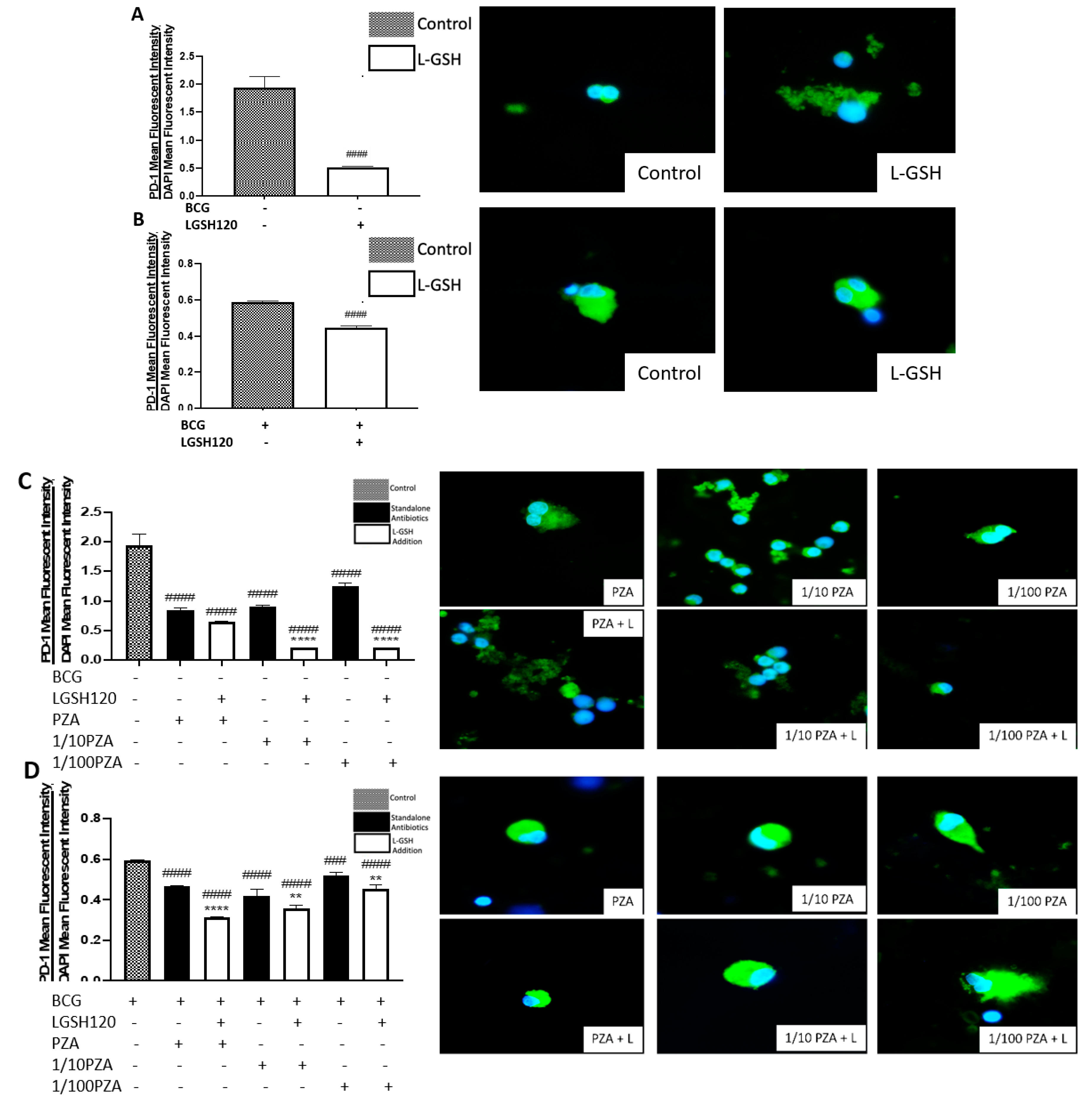
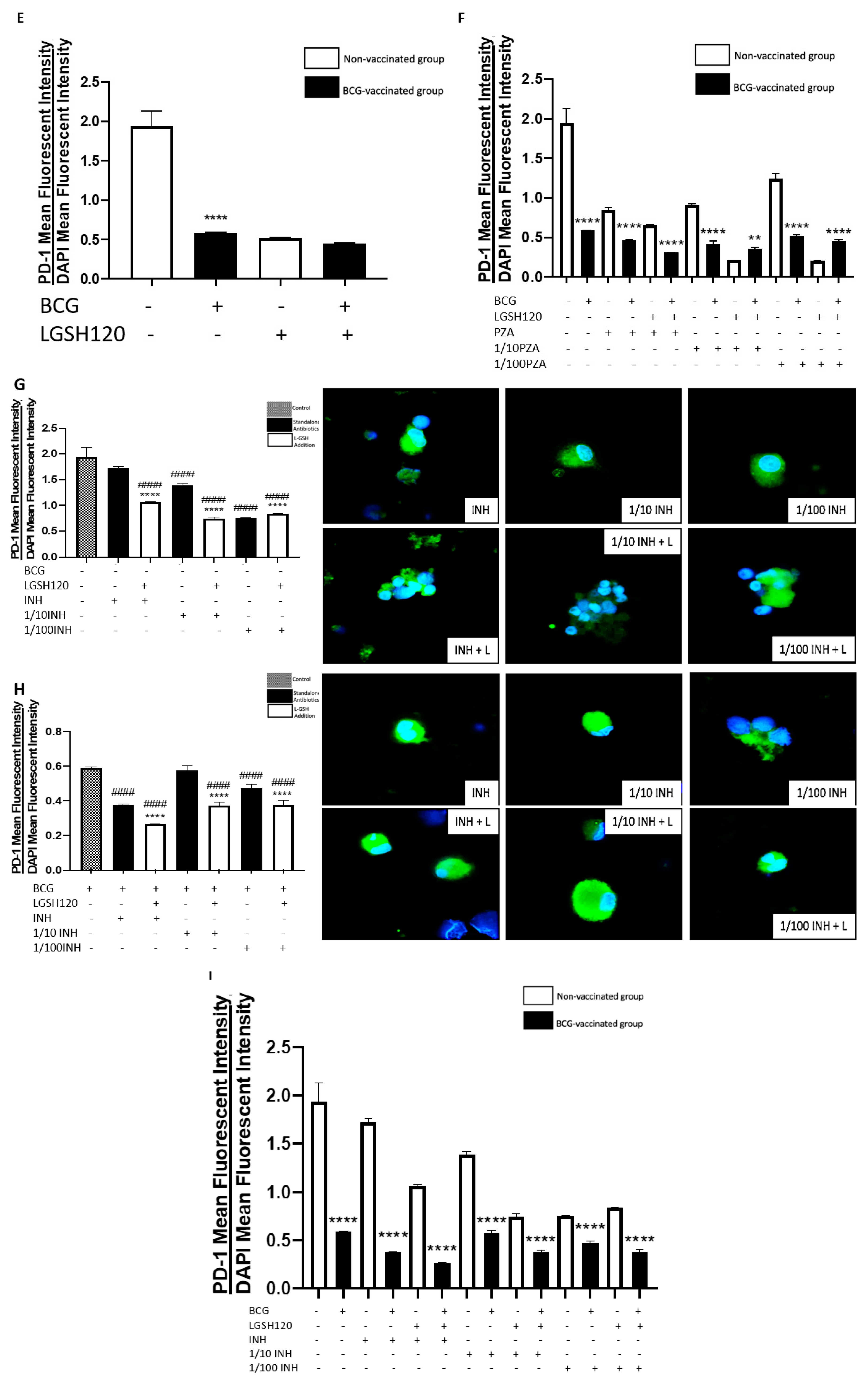
A programmed death receptor 1 (PD-1) is a type I transmembrane protein expressed in immune cells, such as T, B, and NK cells [
22,
23]. When it binds to its receptor, PD-L1, PD-1 strongly interferes with T cell receptor (TCR) signal transduction [
22,
23]. PD-1 is a negative regulator of activated T cells, and blockage of this path restores T cell functions [
10,
24]. For this reason, our lab quantified the levels of PD-1 in non-vaccinated and BCG-vaccinated subjects. We observed a significant decrease in the expression of PD1 in the sham control, PZA-treated, and INH-treated granulomas from BCG-vaccinated individuals when compared to the same categories in the non-vaccinated group (
Figure 5F,I). The addition of L-GSH significantly reduced PD-1 expression in both non-vaccinated and BCG-vaccinated groups (
Figure 5A,B). These results indicate that low PD-1 expression in BCG-vaccinated individuals leads to less T cell exhaustion during
M. tb infection. Decreased PD-1 expression in BCG-vaccinated subjects, along with increased CD4 and CD8 expressions, resulted in improved killing and containment of
M. tb.
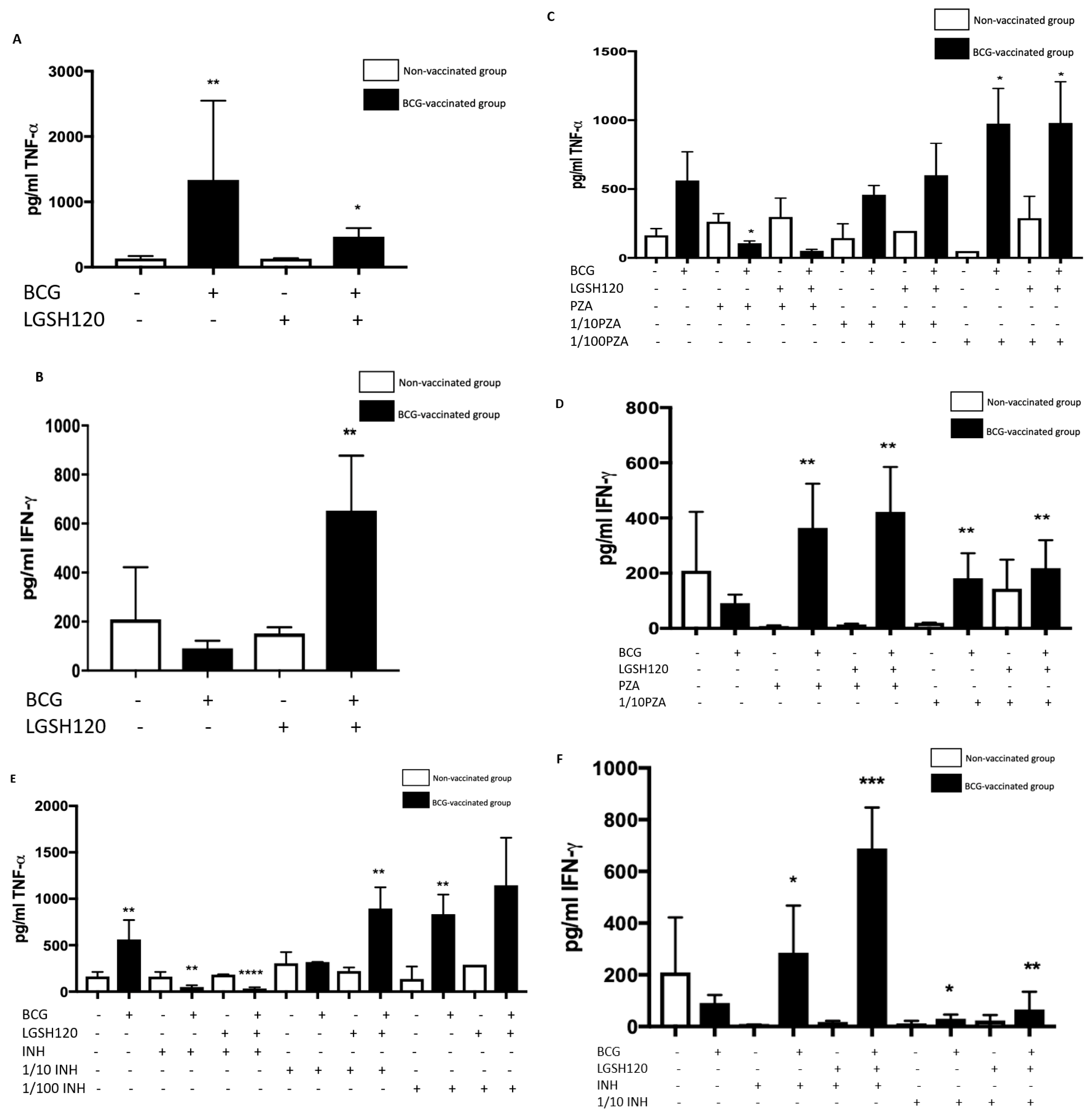
Cytokine production is a key determinate in the containment of a mycobacterial infection. TNF-α released during initiation of host immune response against
M. tb infection causes recruitment of immune cells to form a granuloma while IFN-γ enhances effector functions of macrophages to control an intracellular
M. tb infection. While TNF-α is a vital cytokine in containing
M. tb, overexpression of TNF-α has been connected to multiple autoimmune inflammatory diseases [
24,
25,
26]. Hence modulation of this cytokine is necessary. BCG-vaccinated individuals had higher levels of TNF-α in the sham-treated category (
Figure 6A). L-GSH has a modulatory effect on the production of TNF-α in the BCG-vaccinated subjects (
Figure 6A). Treatment of granulomas from BCG-vaccinated subjects with lower concentrations of PZA and INH in the presence or absence of L-GSH led to higher TNF-α production (
Figure 6C,E). In the MIC concentration for both PZA and INH, TNF-α levels were diminished in the BCG-vaccinated categories. This is because at MIC, the CFUs were undetectable and, hence, there is restoration of homeostasis in the immune responses. L-GSH, PZA, and INH treatments also significantly increased the production of IFN-γ in granulomas from BCG-vaccinated subjects when compared to the non-vaccinated subjects (
Figure 6D,F). This increased production of IFN-γ may be the mechanism by which BCG-vaccinated subjects can control
M. tb infection.
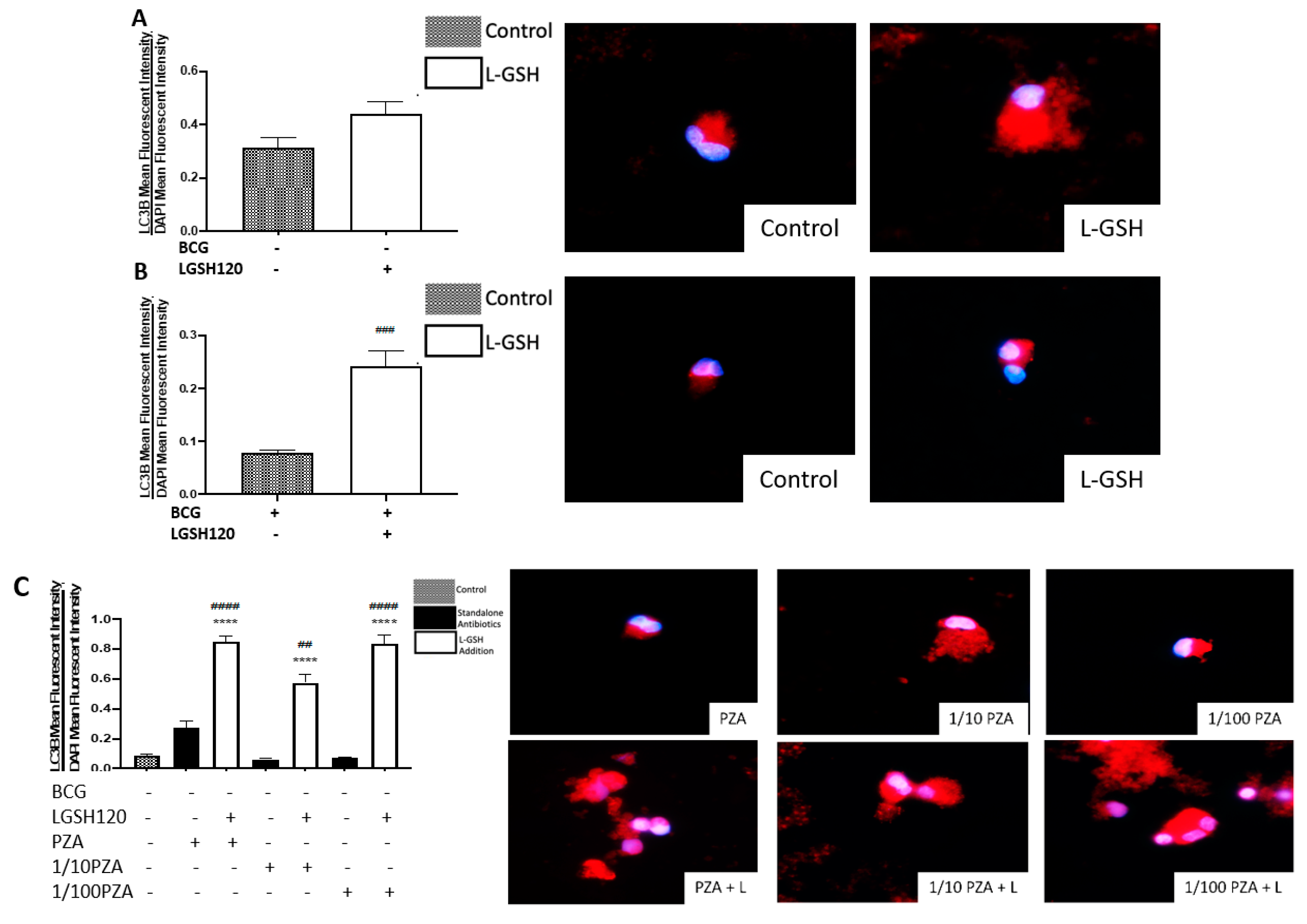
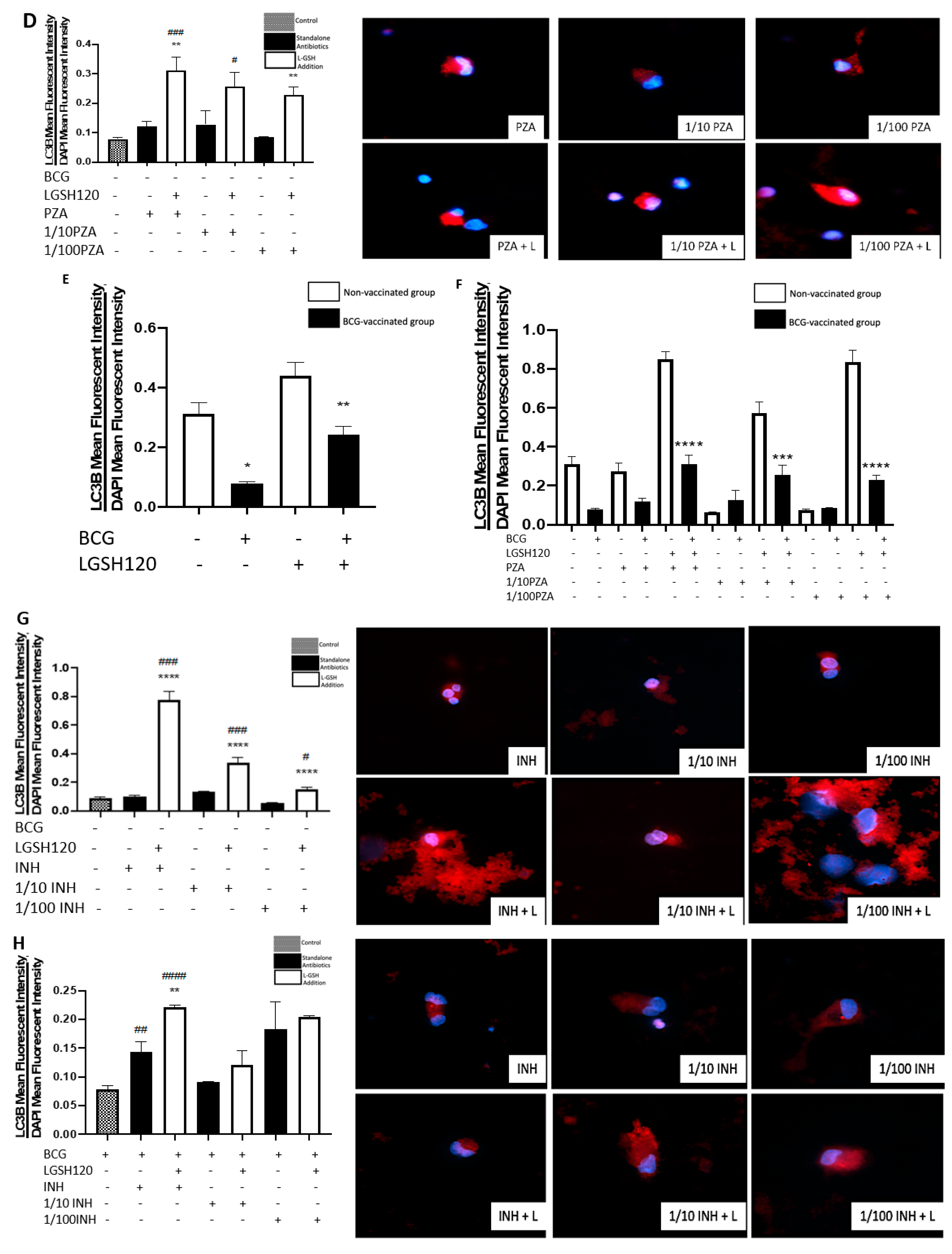
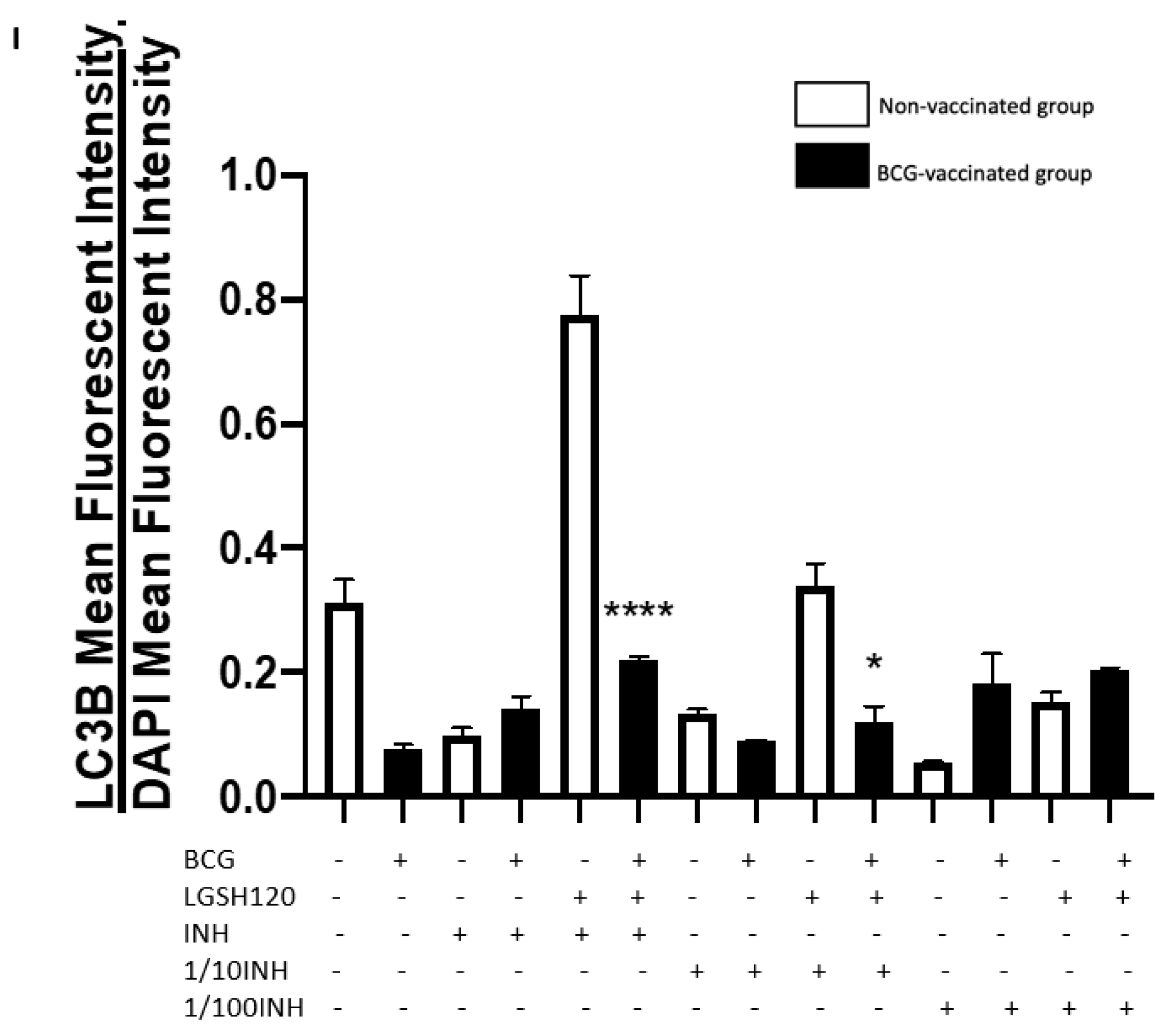
Recently, autophagy has also been recognized as an immune effector mechanism against intracellular pathogens [
27,
28]. We observed that BCG-vaccinated individuals have a significant decrease in the expression of LC3B when compared to non-vaccinated individuals in the presence and absence of L-GSH, PZA and INH (
Figure 7F,I). Our results indicate that autophagy could be a compensatory mechanism by which non-vaccinated individuals combat an
M. tb infection.
Our study findings illustrate that the addition of L-GSH to the antibiotic treatment elicited a significant improvement in the granulomatous responses against M. tb infection. Our results indicate that in non-vaccinated individuals there was increased survival of M. tb, low CD8 T cell counts, and higher expression of PD-1. The addition of L-GSH to granulomas from non-vaccinated individuals restored the viability of CD8 and CD4 T cells and promoted autophagy. In contrast to the non-vaccinated subjects, BCG-vaccinated subjects innately able to kill M. tb more effectively, and this was accompanied by increased levels of GSH, higher CD8 counts, and decreased expression of PD-1. The addition of L-GSH to granulomas from BCG-vaccinated subjects lead to increased CD4 T cell viability, increased TNF-α and IFN-γ production, decreased PD-1 expression, and diminished T cell exhaustion. Therefore, we believe that enhancing GSH by means of L-GSH supplementation, along with anti-TB treatment, would not only reduce toxicity and treatment duration, but can also enhance the host immune responses to combat an active infection to promote enhanced treatment compliance.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}














