Metformin Promotes Osteogenic Differentiation of Adipose-Derived Stromal Cells and Exerts Pro-Osteogenic Effect Stimulating Bone Regeneration

Abstract
1. Introduction
2. Experimental Section
2.1. Isolation and Culture of Rat Adipose-Derived Multipotent Mesenchymal Stromal Cells (rASC)
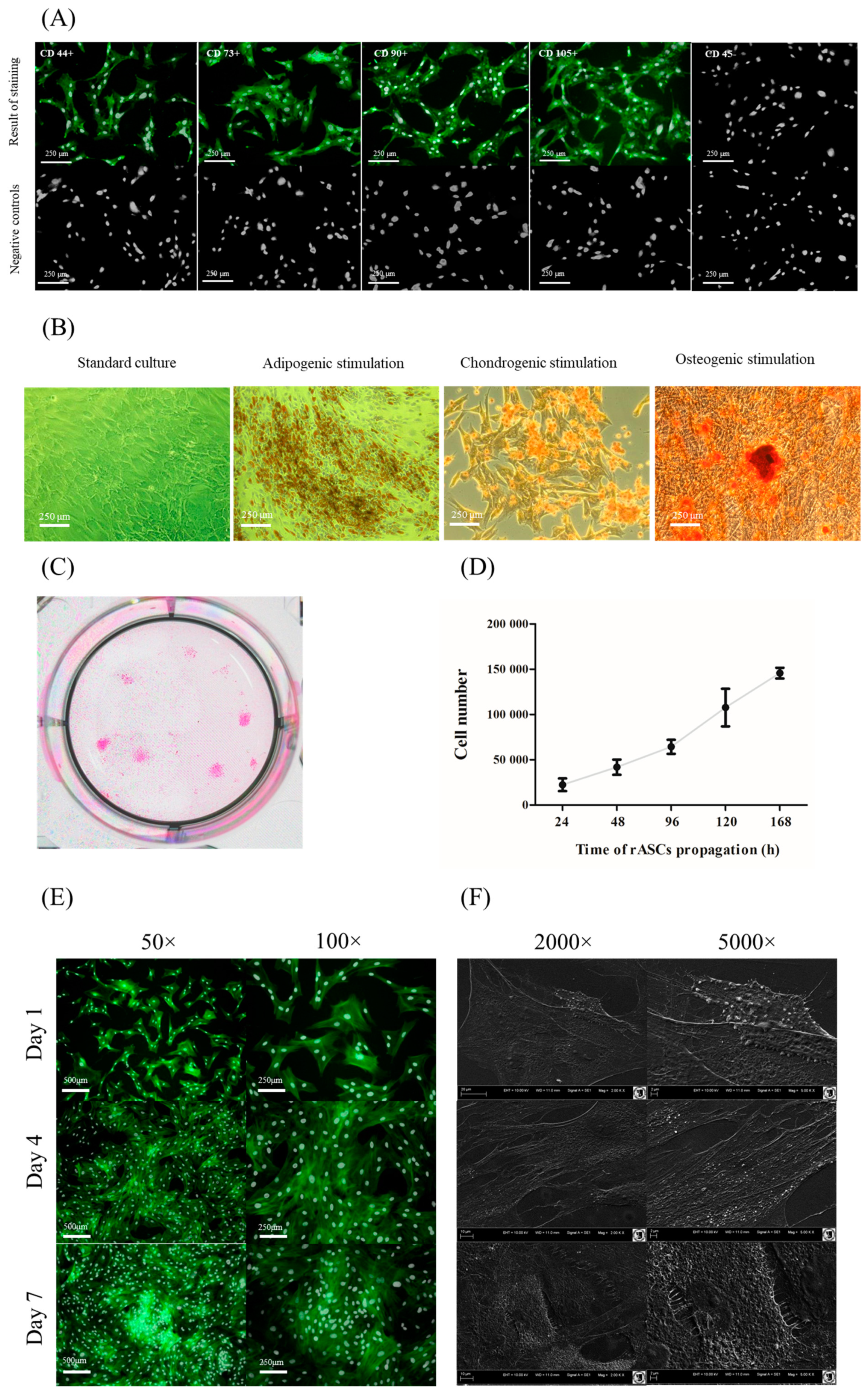
2.2. Characterization of rASCs
2.2.1. Immunophentotype
2.2.2. Multipotency Assay
2.2.3. The Proliferative Activity
2.2.4. Morphology and Growth Pattern
2.3. Effect of Metformin on rASCs Osteogenic Cultures-In Vitro Model
2.3.1. Experimental Cultures with Metformin
2.3.2. Efficiency of Osteogenesis in rASCcultures with Metformin
2.3.3. Mechanisms of Metformin Action
2.4. The Effect of Metformin on Bone Mass and Fracture Healing in Rats (In Vivo Model)
3. Results
3.1. Characteristics of rASC Used in the Experiment
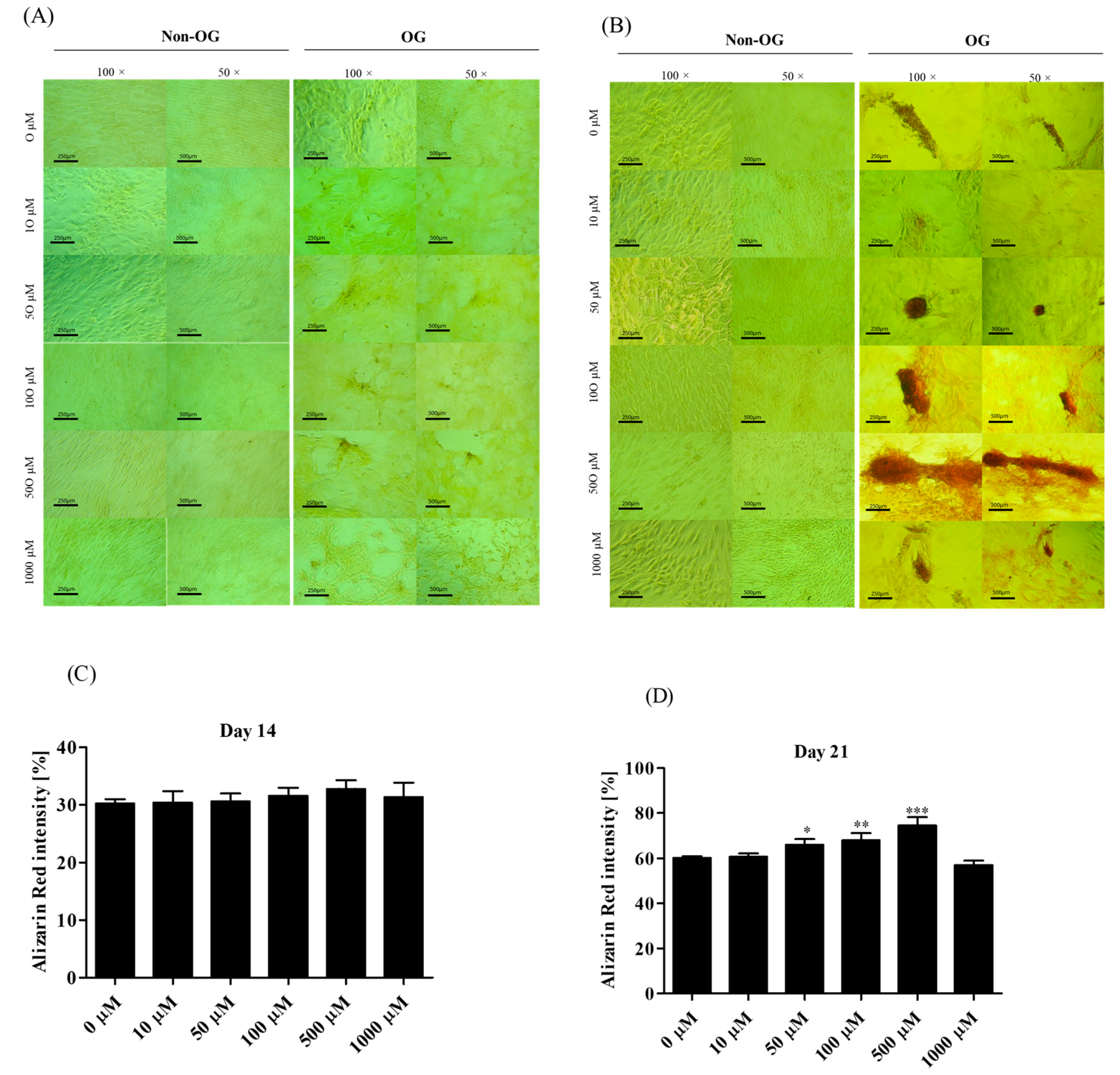
3.2. Osteogenic Cultures with Metformin
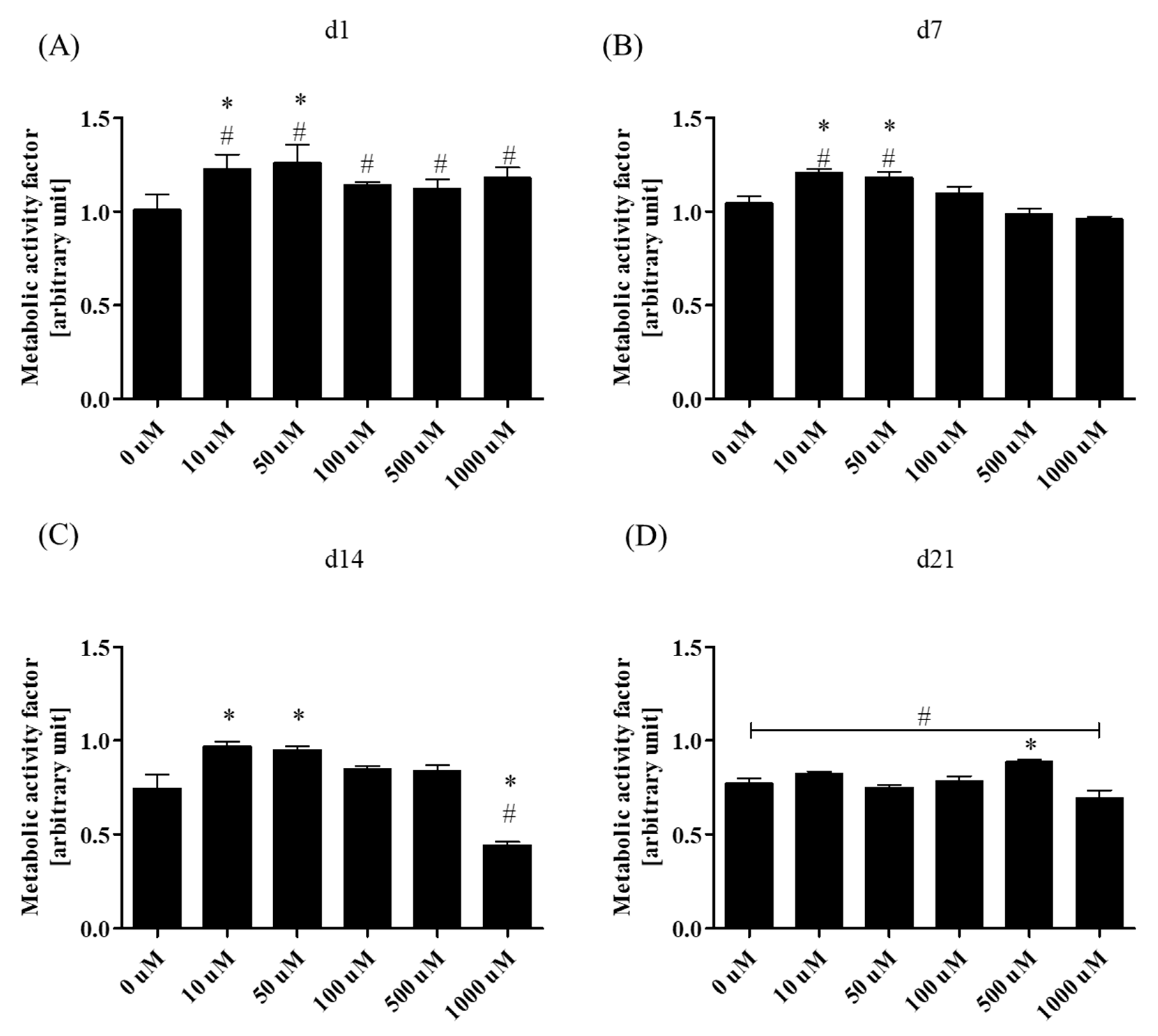
3.2.1. Metabolic Activity
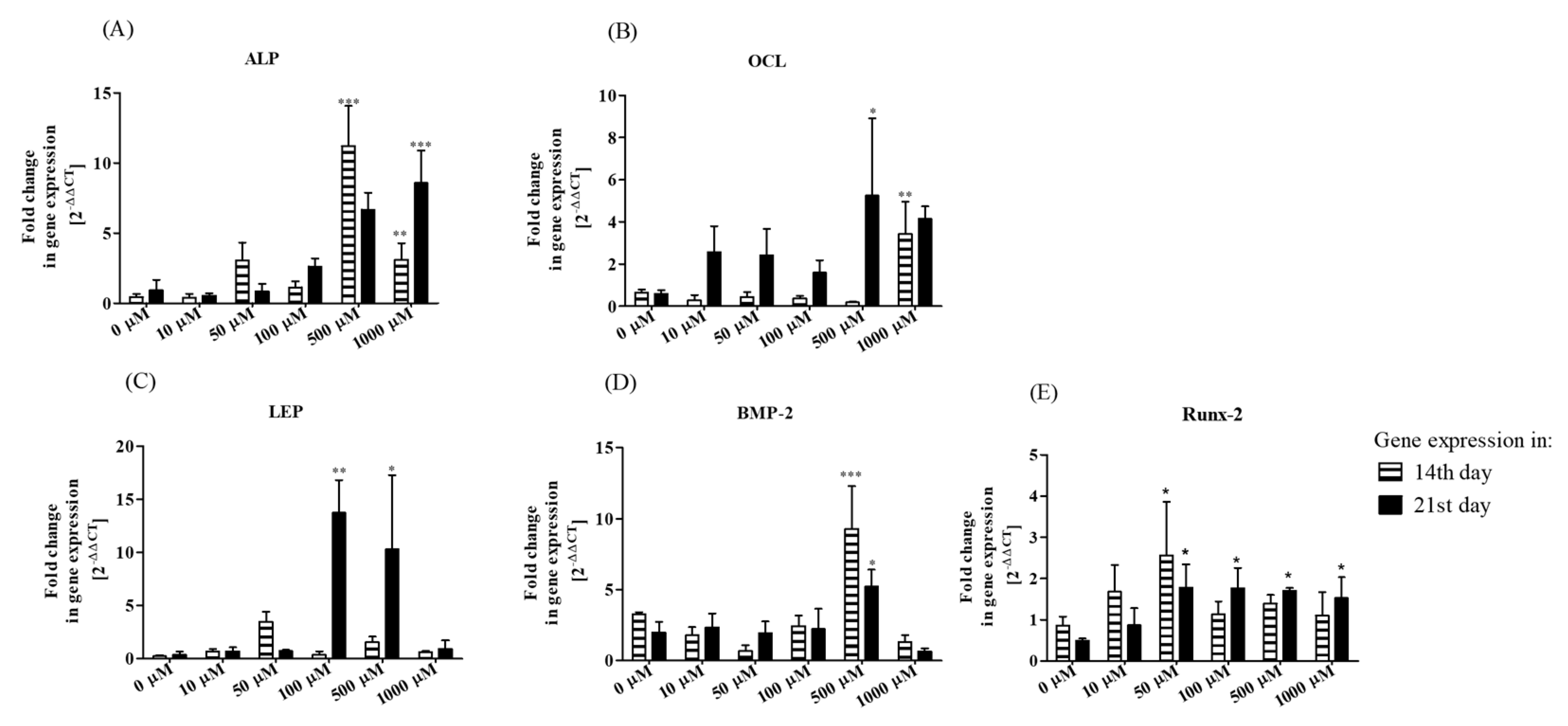
3.2.2. Bone-Associated Markers’ Gene Expression Analysis
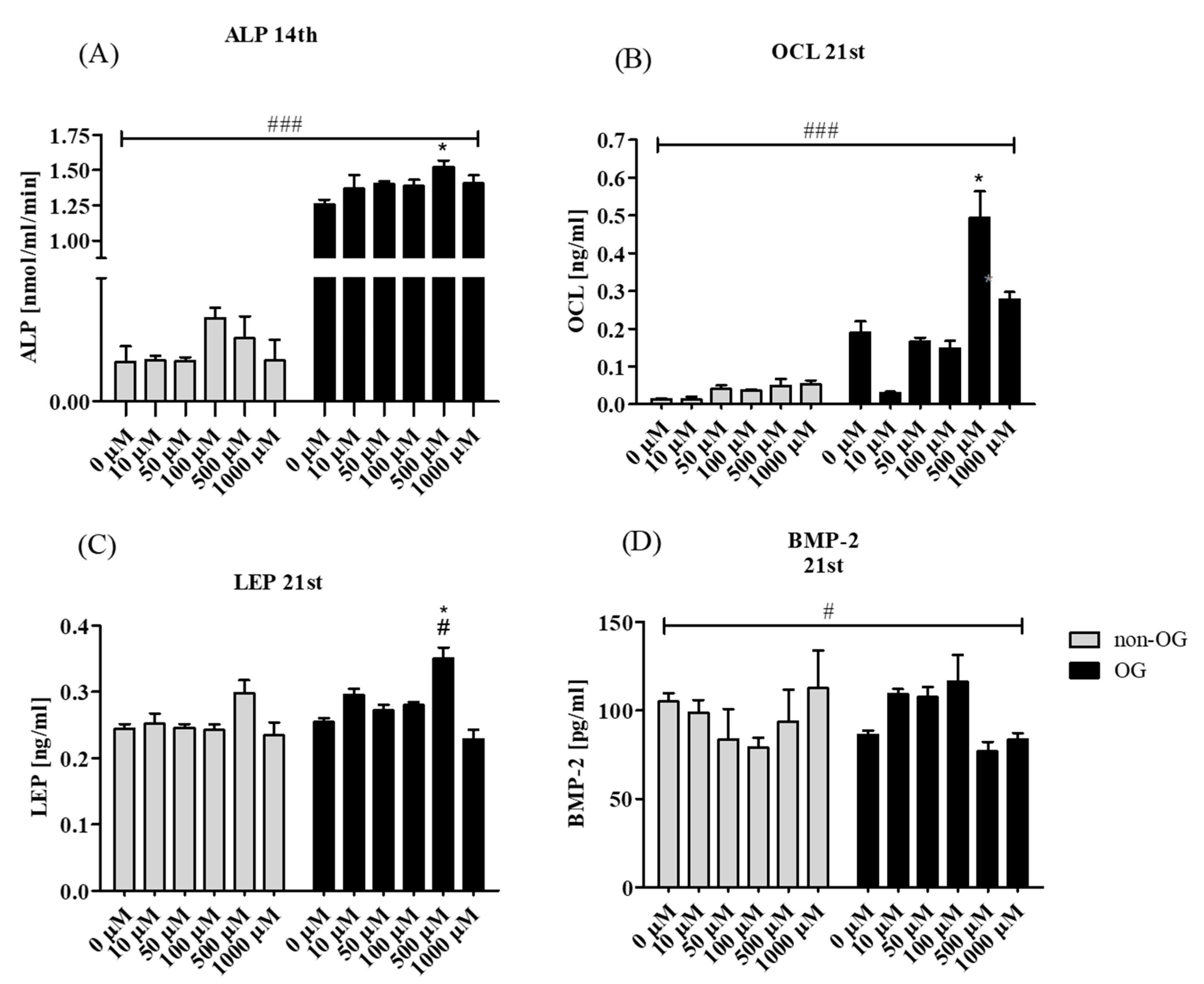
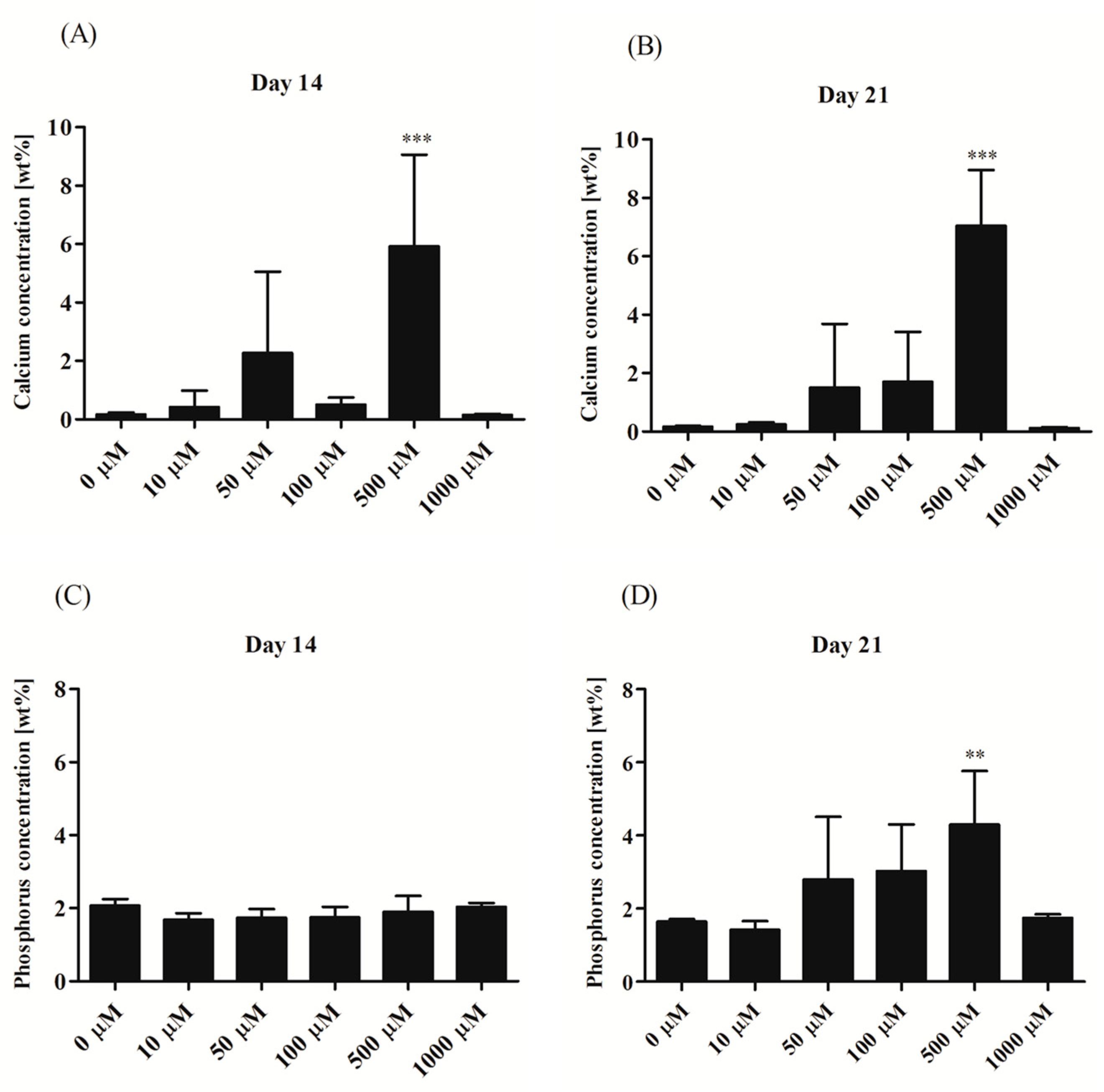
3.2.3. Secretory Activity
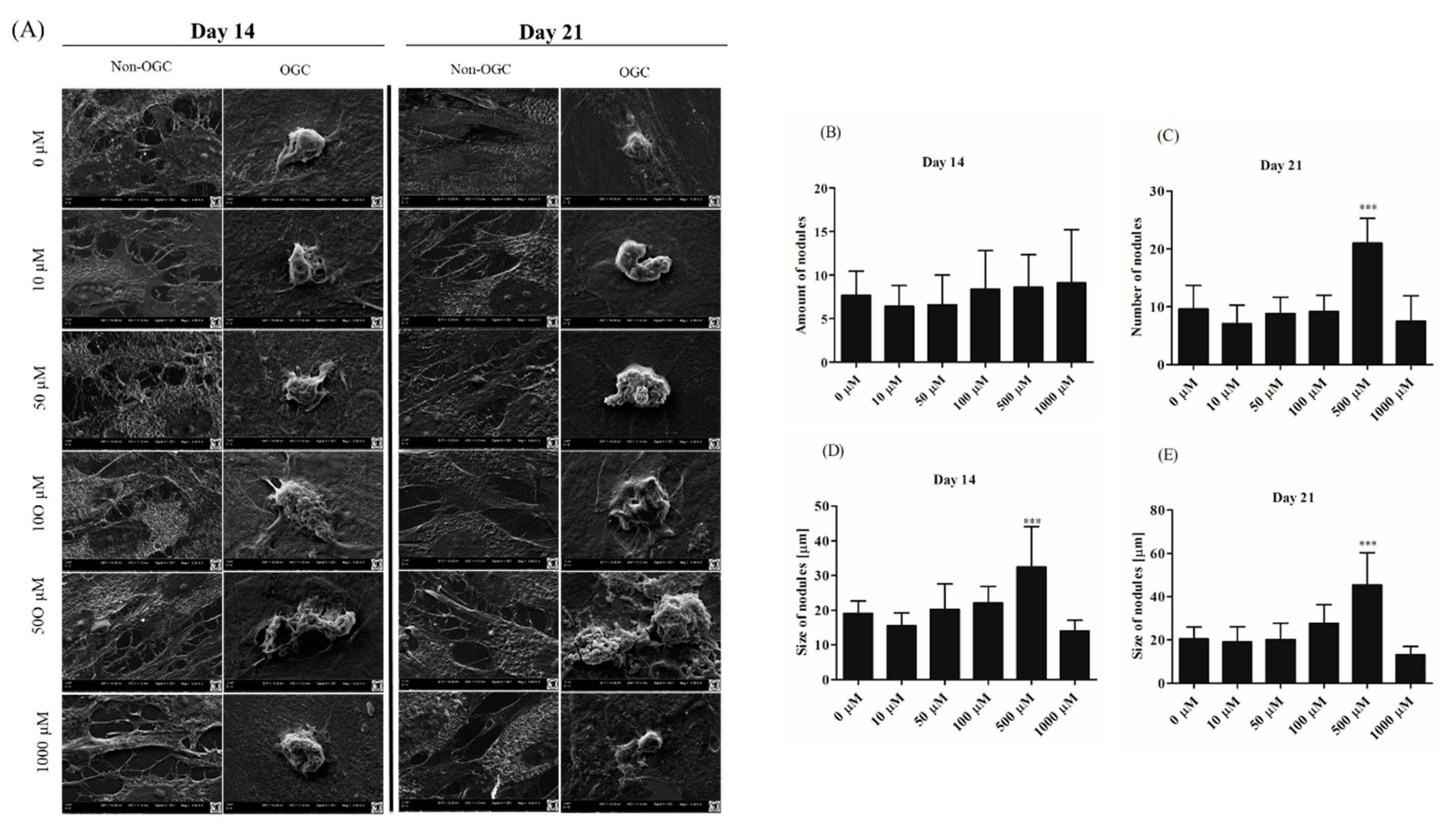
3.2.4. Morphological Alternations of rASCs Cultures in the Course of Osteogenic Differentiation
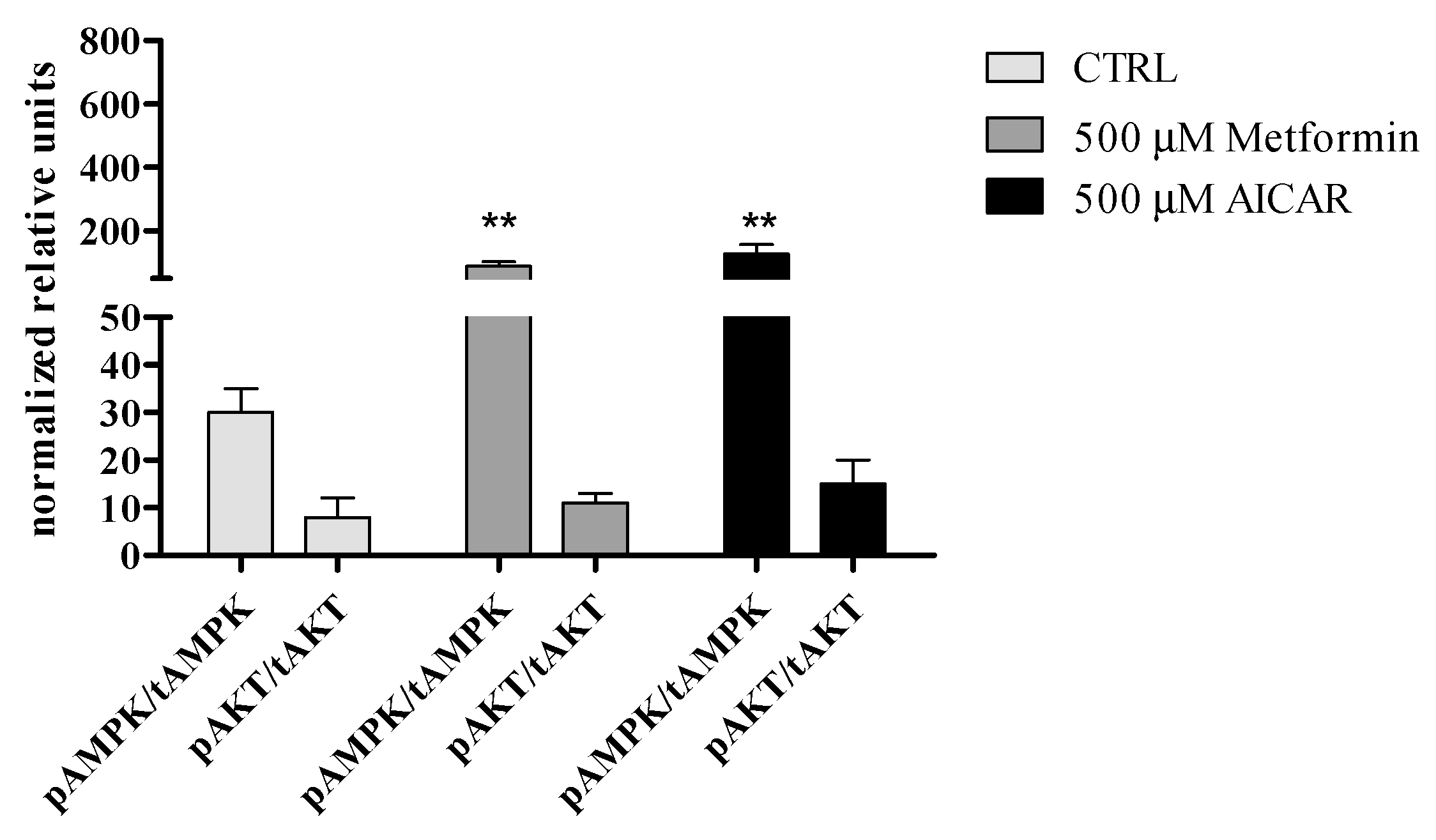
3.2.5. The Effect of Metformin Action on AMPK and AKT Signaling Pathways in rASCs
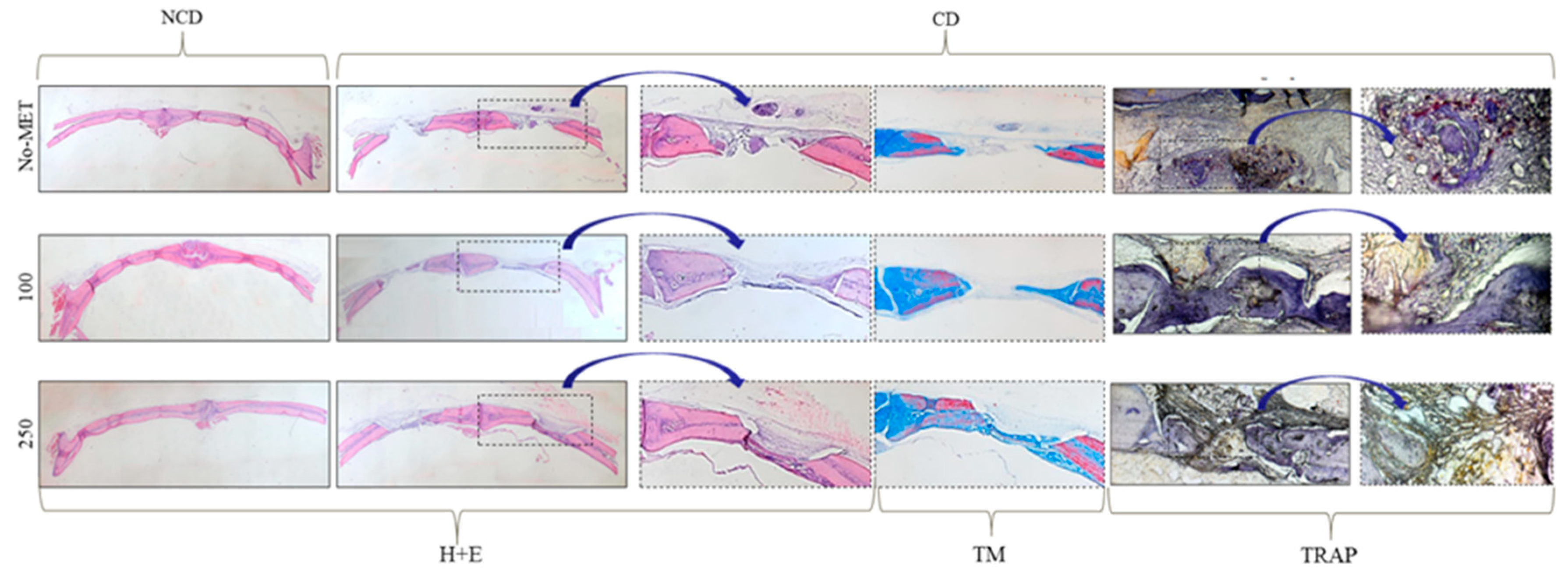
3.3. Osteogenic Effect of Metformin-In Vivo
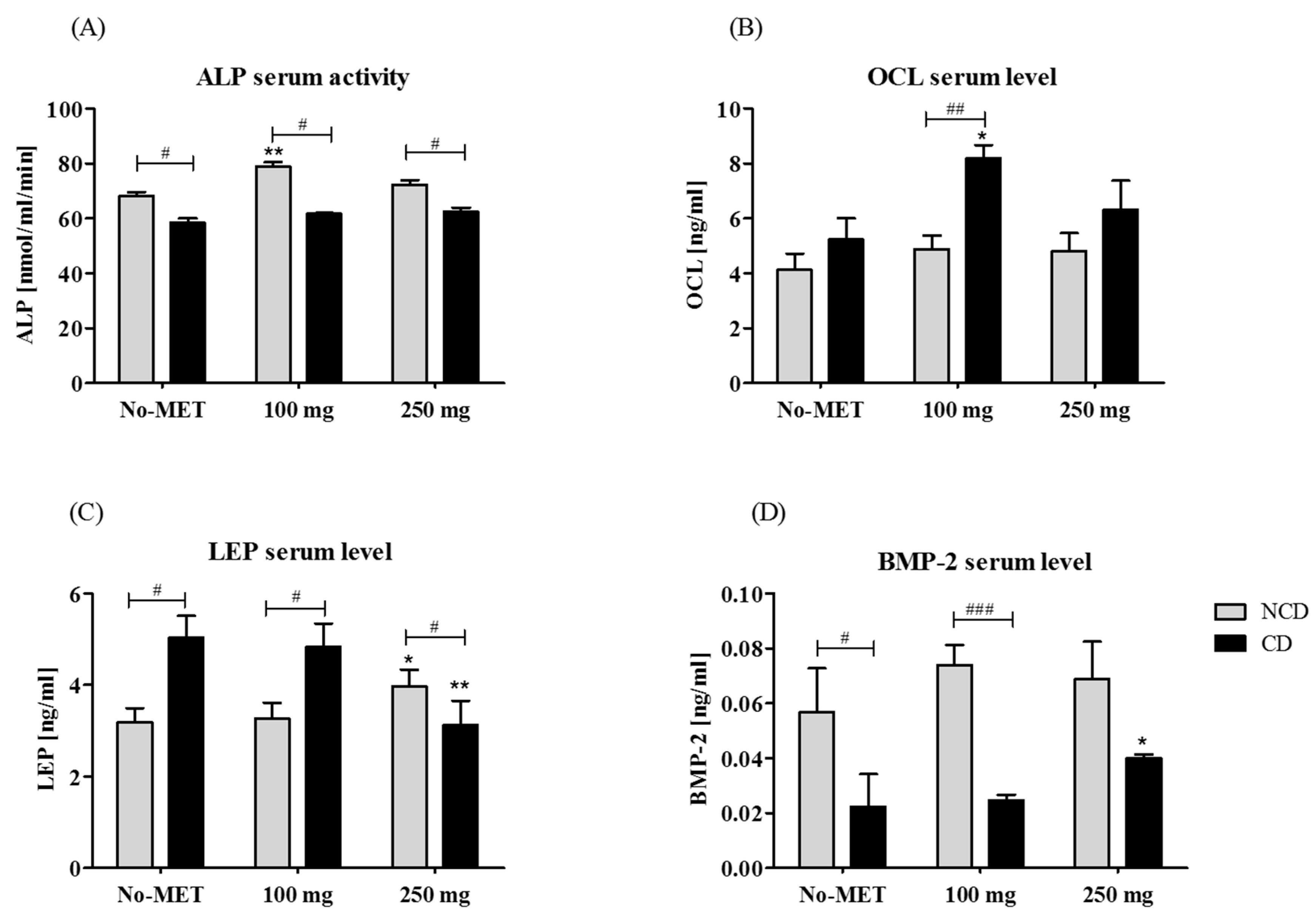
3.3.1. The Circulating Concentration of Osteogenic Proteins and Leptin
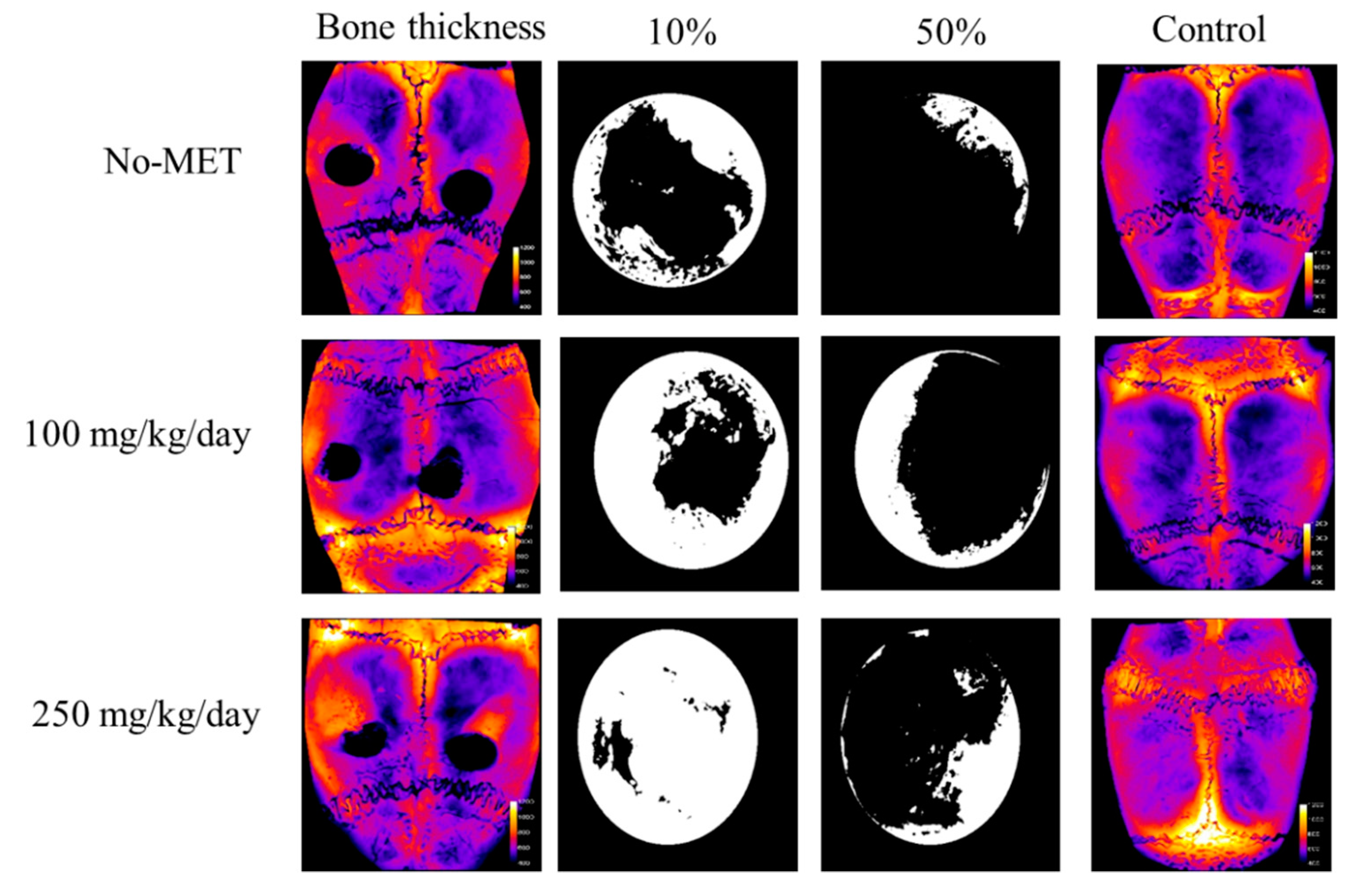
3.3.2. Microcomputed Tomography Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Odén, A.; McCloskey, E.V.; Kanis, J.A.; Harvey, N.C.; Johansson, H. Burden of high fracture probability worldwide: secular increases 2010–2040. Osteoporos. Int. 2015, 26, 2243–2248. [Google Scholar] [CrossRef]
- Borgström, F.; Lekander, I.; Ivergård, M.; Ström, O.; Svedbom, A.; Alekna, V.; Bianchi, M.L.; Clark, P.; Curiel, M.D.; Dimai, H.P.; et al. The International Costs and Utilities Related to Osteoporotic Fractures Study (ICUROS)-quality of life during the first 4 months after fracture. Osteoporos. Int. 2013, 24, 811–823. [Google Scholar] [CrossRef]
- van Houdt, C.I.A.; Tim, C.R.; Crovace, M.C.; Zanotto, E.D.; Peitl, O.; Ulrich, D.J.; Jansen, J.A.; Parizotto, N.A.; Renno, A.C.; van den Beucken, J.J. Bone regeneration and gene expression in bone defects under healthy and osteoporotic bone conditions using two commercially available bone graft substitutes. Biomed. Mater. 2015, 10, 035003. [Google Scholar] [CrossRef]
- Shao, J.; Zhang, W.; Yang, T. Using mesenchymal stem cells as a therapy for bone regeneration and repairing. Biol. Res. 2015, 48, 62. [Google Scholar] [CrossRef]
- Niemeyer, P.; Fechner, K.; Milz, S.; Richter, W.; Suedkamp, N.P.; Mehlhorn, A.T.; Pearce, S.; Kasten, P. Comparison of mesenchymal stem cells from bone marrow and adipose tissue for bone regeneration in a critical size defect of the sheep tibia and the influence of platelet-rich plasma. Biomaterials 2010, 31, 3572–3579. [Google Scholar] [CrossRef]
- Li, C.; Wu, X.Y.; Tong, J.B.; Yang, X.X.; Zhao, J.L.; Zheng, Q.F.; Zhao, G.B.; Ma, Z.J. Comparative analysis of human mesenchymal stem cells from bone marrow and adipose tissue under xeno-free conditions for cell therapy. Stem Cell Res. Ther. 2015, 6, 55. [Google Scholar] [CrossRef]
- Han, D.; Han, N.; Zhang, P.; Jiang, B. Local transplantation of osteogenic pre-differentiated autologous adipose-derived mesenchymal stem cells may accelerate non-union fracture healing with limited pro-metastatic potency. Int. J. Clin. Exp. Med. 2015, 8, 1406–1410. [Google Scholar]
- Śmieszek, A.; Szydlarska, J.; Mucha, A.; Chrapiec, M.; Marycz, K. Enhanced cytocompatibility and osteoinductive properties of sol-gel-derived silica/zirconium dioxide coatings by metformin functionalization. J. Biomater. Appl. 2017, 32, 570–586. [Google Scholar] [CrossRef]
- Gao, Y.; Xue, J.; Li, X.; Jia, Y.; Hu, J. Metformin regulates osteoblast and adipocyte differentiation of rat mesenchymal stem cells. J. Pharm. Pharmacol. 2008, 60, 1695–1700. [Google Scholar] [CrossRef]
- Gao, Y.; Li, Y.; Xue, J.; Jia, Y.; Hu, J. Effect of the anti-diabetic drug metformin on bone mass in ovariectomized rats. Eur. J. Pharmacol. 2010, 635, 231–236. [Google Scholar] [CrossRef]
- Molinuevo, M.S.; Schurman, L.; McCarthy, A.D.; Cortizo, A.M.; Tolosa, M.J.; Gangoiti, M.V.; Arnol, V.; Sedlinsky, C. Effect of metformin on bone marrow progenitor cell differentiation: In vivo and in vitro studies. J. Bone Miner. Res. 2010, 25, 211–221. [Google Scholar] [CrossRef]
- Sedlinsky, C.; Molinuevo, M.S.; Cortizo, A.M.; Tolosa, M.J.; Felice, J.I.; Sbaraglini, M.L.; Schurman, L.; McCarthy, A.D. Metformin prevents anti-osteogenic in vivo and ex vivo effects of rosiglitazone in rats. Eur. J. Pharmacol. 2011, 668, 477–485. [Google Scholar] [CrossRef]
- Bromage, D.I.; Yellon, D.M. The pleiotropic effects of metformin: time for prospective studies. Cardiovasc. Diabetol. 2015, 14, 109. [Google Scholar] [CrossRef]
- Hattori, Y.; Hattori, K.; Hayashi, T. Pleiotropic benefits of metformin: Macrophage targeting its anti-inflammatory mechanisms. Diabetes 2015, 64, 1907–1909. [Google Scholar] [CrossRef]
- Cifarelli, V.; Lashinger, L.M.; Devlin, K.L.; Dunlap, S.M.; Huang, J.; Kaaks, R.; Pollak, M.N.; Hursting, S.D. Metformin and rapamycin reduce pancreatic cancer growth in obese prediabetic mice by distinct microrna-regulated mechanisms. Diabetes 2015, 64, 1632–1642. [Google Scholar] [CrossRef]
- Lalau, J.D.; Arnouts, P.; Sharif, A.; De Broe, M.E. Metformin and other antidiabetic agents in renal failure patients. Kidney Int. 2015, 87, 308–322. [Google Scholar] [CrossRef]
- Vestergaard, P.; Rejnmark, L.; Mosekilde, L. Relative fracture risk in patients with diabetes mellitus, and the impact of insulin and oral antidiabetic medication on relative fracture risk. Diabetologia 2005, 48, 1292–1299. [Google Scholar] [CrossRef]
- Cortizo, A.M.; Sedlinsky, C.; McCarthy, A.D.; Blanco, A.; Schurman, L. Osteogenic actions of the anti-diabetic drug metformin on osteoblasts in culture. Eur. J. Pharmacol. 2006, 536, 38–46. [Google Scholar] [CrossRef]
- Kanazawa, I.; Yamaguchi, T.; Yano, S.; Yamauchi, M.; Sugimoto, T. Metformin enhances the differentiation and mineralization of osteoblastic MC3T3-E1 cells via AMP kinase activation as well as eNOS and BMP-2 expression. Biochem. Biophys. Res. Commun. 2008, 375, 414–419. [Google Scholar] [CrossRef]
- Kovacic, S.; Soltys, C.-L.M.; Barr, A.J.; Shiojima, I.; Walsh, K.; Dyck, J.R. Akt activity negatively regulates phosphorylation of AMP-activated protein kinase in the heart. J. Biol. Chem. 2003, 278, 39422–39427. [Google Scholar] [CrossRef]
- Pantovic, A.; Krstic, A.; Janjetovic, K.; Kocic, J.; Harhaji-Trajkovic, L.; Bugarski, D.; Trajkovic, V. Coordinated time-dependent modulation of AMPK/Akt/mTOR signaling and autophagy controls osteogenic differentiation of human mesenchymal stem cells. Bone 2013, 52, 524–531. [Google Scholar] [CrossRef]
- Luo, T.; Nocon, A.; Fry, J.; Sherban, A.; Rui, X.; Jiang, B.; Xu, X.J.; Han, J.; Yan, Y.; Yang, Q. AMPK Activation by Metformin Suppresses Abnormal Extracellular Matrix Remodeling in Adipose Tissue and Ameliorates Insulin Resistance in Obesity. Diabetes 2016, 65, 2295–2310. [Google Scholar] [CrossRef]
- Śmieszek, A.; Basińska, K.; Chrząstek, K.; Marycz, K. In vitro and in vivo effects of metformin on osteopontin expression in mice adipose-derived multipotent stromal cells and adipose tissue. J. Diabetes Res. 2015. [Google Scholar] [CrossRef]
- Marycz, K.; Tomaszewski, K.A.; Kornicka, K.; Henry, B.M.; Wroński, S.; Tarasiuk, J.; Maredziak, M. Metformin decreases reactive oxygen species, enhances osteogenic properties of adipose-derived multipotent mesenchymal stem cells in vitro, and increases bone density in vivo. Oxid. Med. Cell Longev. 2016, 2016, 9785890. [Google Scholar] [CrossRef]
- Jeyabalan, J.; Viollet, B.; Smitham, P.; Ellis, S.A.; Zaman, G.; Bardin, C.; Goodship, A.; Roux, J.; Pierre, M.; Chenu, C. The anti-diabetic drug metformin does not affect bone mass in vivo or fracture healing. Osteoporos. Int. 2013, 24, 2659–2670. [Google Scholar] [CrossRef]
- Hegazy, S.K. Evaluation of the anti-osteoporotic effects of metformin and sitagliptin in postmenopausal diabetic women. J. Bone Miner. Metab. 2015, 33, 207–212. [Google Scholar] [CrossRef]
- Marycz, K.; Śmieszek, A.; Jeleń, M.; Chrząstek, K.; Grzesiak, J.; Meissner, J. The effect of the bioactive sphingolipids S1P and C1P on multipotent stromal cells-new opportunities in regenerative medicine. Cell Mol. Biol. Lett. 2015, 20, 510–533. [Google Scholar] [CrossRef]
- Roth, V. Doubling Time Computing. 2006. Available online: http://www.doubling-time.com/compute.php (accessed on 11 October 2018).
- Zimoch-Korzycka, A.; Śmieszek, A.; Jarmoluk, A.; Nowak, U.; Marycz, K. Potential Biomedical Application of Enzymatically Treated Alginate/Chitosan Hydrosols in Sponges—Biocompatible Scaffolds Inducing Chondrogenic Differentiation of Human Adipose Derived Multipotent Stromal Cells. Polymers 2016, 8, 320. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Grzesiak, J.; Śmieszek, A.; Marycz, K. Ultrastructural changes during osteogenic differentiation in mesenchymal stromal cells cultured in alginate hydrogel. Cell Biosci. 2017, 7, 2. [Google Scholar] [CrossRef]
- Śmieszek, A.; Stręk, Z.; Kornicka, K.; Grzesiak, J.; Weiss, C.; Marycz, K. Antioxidant and Anti-Senescence Effect of Metformin on Mouse Olfactory Ensheathing Cells (mOECs) May Be Associated with Increased Brain-Derived Neurotrophic Factor Levels-An Ex Vivo Study. Int. J. Mol. Sci. 2017, 18, 872. [Google Scholar] [CrossRef]
- Santana, R.B.; Xu, L.; Chase, H.B.; Amar, S.; Graves, D.T.; Trackman, P.C. A role for advanced glycation end products in diminished bone healing in type 1 diabetes. Diabetes 2003, 52, 1502–1510. [Google Scholar] [CrossRef]
- Parasuraman, S.; Raveendran, R.; Kesavan, R. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacother. 2010, 1, 87–93. [Google Scholar] [CrossRef]
- OSA|Practical Cone-Beam Algorithm. Available online: https://www.osapublishing.org/josaa/abstract.cfm?URI=josaa-1-6-612 (accessed on 12 November 2018).
- VGSTUDIO MAX. High-End Software for CT Data. Available online: https://www.volumegraphics.com/en/products/vgstudio-max.html (accessed on 12 November 2018).
- Fiji is just ImageJ. Available online: https://fiji.sc/#cite (accessed on 12 November 2018).
- Doube, M.; Kłosowski, M.M.; Arganda-Carreras, I.; Cordelières, F.P.; Dougherty, R.P.; Jackson, J.S.; Schmid, B.; Hutchinson, J.R.; Shefelbine, S.J. BoneJ: Free and extensible bone image analysis in ImageJ. Bone 2010, 47, 1076–1079. [Google Scholar] [CrossRef]
- Voxler®|3D Geologic and Scientific Modeling Software. Available online: https://www.goldensoftware.com/products/voxler (accessed on 12 November 2018).
- Metformin Rescues the MG63 Osteoblasts against the Effect of High Glucose on Proliferation. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24812633 (accessed on 12 November 2018).
- Casteilla, L.; Planat-Benard, V.; Laharrague, P.; Cousin, B. Adipose-derived stromal cells: Their identity and uses in clinical trials, an update. World J. Stem Cells 2011, 3, 25–33. [Google Scholar] [CrossRef]
- Dubey, N.K.; Mishra, V.K.; Dubey, R.; Deng, Y.H.; Tsai, F.C.; Deng, W.P. Revisiting the advances in isolation, characterization and secretome of adipose-derived stromal/stem cells. Int. J. Mol. Sci. 2018, 19, 2200. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Ets Transcription Factors and Targets in Osteogenesis. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11175361 (accessed on 12 November 2018).
- Transcription Adaptation during In Vitro Adipogenesis and Osteogenesis of Porcine Mesenchymal Stem Cells: Dynamics of Pathways, Biological Processes, Up-Stream Regulators, and Gene Networks. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0137644 (accessed on 12 November 2018).
- Wang, P.; Ma, T.; Guo, D.; Hu, K.; Shu, Y.; Xu, H.H.K.; Schneider, A. Metformin induces osteoblastic differentiation of human induced pluripotent stem cell-derived mesenchymal stem cells. J. Tissue Eng. Regen. Med. 2018, 12, 437–446. [Google Scholar] [CrossRef]
- Beederman, M.; Lamplot, J.D.; Nan, G.; Wang, J.; Liu, X.; Yin, L.; Li, R.; Shui, W.; Zhang, H.; Kim, S.H. BMP signaling in mesenchymal stem cell differentiation and bone formation. J. Biomed. Sci. Eng. 2013, 6, 32–52. [Google Scholar] [CrossRef]
- Guo, Y.; Xiao, P.; Lei, S.; Xiao, G.G.; Liu, Y.; Chen, X.; Li, L.; Wu, S.; Chen, Y.; Jiang, H.; et al. How is mRNA expression predictive for protein expression? A correlation study on human circulating monocytes. Acta Biochim. Biophys. Sin. 2008, 40, 426–436. [Google Scholar] [CrossRef]
- Włodarski, K.; Włodarski, P. Leptin as a modulator of osteogenesis. Ortop. Traumatol. Rehabil. 2009, 11, 1–6. [Google Scholar]
- Coen, G. Leptin and bone metabolism. J. Nephrol. 2004, 17, 187–189. [Google Scholar]
- Zheng, B.; Jiang, J.; Luo, K.; Liu, L.; Lin, M.; Chen, Y.; Yan, F. Increased osteogenesis in osteoporotic bone marrow stromal cells by overexpression of leptin. Cell Tissue Res. 2015, 361, 845–856. [Google Scholar] [CrossRef]
- Baruah, M.P.; Kalra, S.; Ranabir, S. Metformin; A character actor in the leptin story! Indian J. Endocrinol. Metab. 2012, 16, S532–S533. [Google Scholar]
- Güler, S.; Cakir, B.; Gürsoy, G.; Serter, R.; Aral, Y. Leptin Concentrations are Related to Glycaemic Control, But Do Not Change with Short-Term Oral Antidiabetic Therapy in Female Patients with Type 2 Diabetes Mellitus. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1463-1326.2000.00073.x (accessed on 12 November 2018).
- Kim, Y.W.; Kim, J.Y.; Park, Y.H.; Park, S.Y.; Won, K.C.; Choi, K.H.; Huh, J.Y.; Moon, K.H. Metformin restores leptin sensitivity in high-fat-fed obese rats with leptin resistance. Diabetes 2006, 55, 716–724. [Google Scholar] [CrossRef]
- AMP-Activated Protein Kinase (AMPK) Activation Regulates in Vitro Bone Formation And Bone Mass. Available online: https://www.ncbi.nlm.nih.gov/pubmed/20399918 (accessed on 12 November 2018).
- Son, H.J.; Lee, J.; Lee, S.Y.; Kim, E.K.; Park, M.J.; Kim, K.W.; Park, S.H.; Cho, M.L. Metformin attenuates experimental autoimmune arthritis through reciprocal regulation of Th17/Treg balance and osteoclastogenesis. Mediat. Inflamm. 2014, 2014, 973986. [Google Scholar] [CrossRef]
- Watanabe, S.; Takeuchi, Y.; Fukumoto, S.; Fujita, H.; Nakano, T.; Fujita, T. Decrease in serum leptin by troglitazone is associated with preventing bone loss in type 2 diabetic patients. J. Bone Miner. Metab. 2003, 21, 166–171. [Google Scholar] [CrossRef]
- Borges, J.L.C.; Bilezikian, J.P.; Jones-Leone, A.R.; Acusta, A.P.; Ambery, P.D.; Nino, A.J.; Grosse, M.; Fitzpatrick, L.A.; Cobitz, A.R. A randomized, parallel group, double-blind, multicentre study comparing the efficacy and safety of Avandamet (rosiglitazone/metformin) and metformin on long-term glycaemic control and bone mineral density after 80 weeks of treatment in drug-naïve type 2 diabetes mellitus patients. Diabetes Obes. Metab. 2011, 13, 1036–1046. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight of Animal (g) | |
|---|---|---|
| No-MET | NCD | 395 ± 5 |
| 100 | 361 ± 12 *# | |
| 250 | 304 ± 29 *# | |
| No-MET | CD | 364 ± 5 |
| 100 | 345 ± 3 * | |
| 250 | 345 ± 6 * | |
| Characteristic | CD | ||
|---|---|---|---|
| No-MET | 100 mg/kg/day | 250 mg/kg/day | |
| Newly formed bone area/Total defect area (%) | 3 ± 1.61 | 28.9 ± 7.80 ** | 32.9 ± 9.63 ** |
| Newly formed bone area (mm2) | 0.02 ± 0.01 | 0.23 ± 0.06 ** | 0.26 ± 0.08 ** |
| TRAP area/Reossification area (%) | 14.59 ± 1.03 | 9.95 ± 2.47 **# | 5.58 ± 2.48 **# |
| Group | V (mm3) | r (mm) | S10/S0 (%) | S50/S0 (%) |
|---|---|---|---|---|
| No-MET_L | 0.59 | 1.13 | 43.3 | 11.0 |
| No-MET_R | 0.41 | 1.12 | 41.2 | 9.5 |
| 100_L | 0.68 | 1.18 | 68.3 | 32.6 |
| 100_R | 0.59 | 1.1 | 53.1 | 18.9 |
| 250_L | 1.02 | 1.14 | 91.9 | 25.0 |
| 250_R | 0.71 | 1.23 | 69.6 | 15.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Śmieszek, A.; Tomaszewski, K.A.; Kornicka, K.; Marycz, K. Metformin Promotes Osteogenic Differentiation of Adipose-Derived Stromal Cells and Exerts Pro-Osteogenic Effect Stimulating Bone Regeneration. J. Clin. Med. 2018, 7, 482. https://doi.org/10.3390/jcm7120482
Śmieszek A, Tomaszewski KA, Kornicka K, Marycz K. Metformin Promotes Osteogenic Differentiation of Adipose-Derived Stromal Cells and Exerts Pro-Osteogenic Effect Stimulating Bone Regeneration. Journal of Clinical Medicine. 2018; 7(12):482. https://doi.org/10.3390/jcm7120482
Chicago/Turabian StyleŚmieszek, Agnieszka, Krzysztof A. Tomaszewski, Katarzyna Kornicka, and Krzysztof Marycz. 2018. "Metformin Promotes Osteogenic Differentiation of Adipose-Derived Stromal Cells and Exerts Pro-Osteogenic Effect Stimulating Bone Regeneration" Journal of Clinical Medicine 7, no. 12: 482. https://doi.org/10.3390/jcm7120482
APA StyleŚmieszek, A., Tomaszewski, K. A., Kornicka, K., & Marycz, K. (2018). Metformin Promotes Osteogenic Differentiation of Adipose-Derived Stromal Cells and Exerts Pro-Osteogenic Effect Stimulating Bone Regeneration. Journal of Clinical Medicine, 7(12), 482. https://doi.org/10.3390/jcm7120482