
TGF-β Signaling in Gastrointestinal Cancers: Progress in Basic and Clinical Research



Abstract
:1. Introduction
2. Role of TGF-β in Cancer
2.1. Significance of TGF-β Expression in Gastrointestinal Cancers
2.2. Role of TGFBI in Cancer
2.3. TGFBI in Gastrointestinal Cancers
2.3.1. Esophageal Cancer
2.3.2. Gastric Cancer
2.3.3. Pancreatic and Biliary Cancer
2.3.4. Colorectal Cancer
3. TGF-β and TGFBI as Molecular Targets
4. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Heldin, C.H.; Vanlandewijck, M.; Moustakas, A. Regulation of EMT by TGFβ in cancer. FEBS Lett. 2012, 586, 1959–1970. [Google Scholar] [CrossRef] [PubMed]
- Massague, J. TGFβ signalling in context, Nature reviews. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar]
- Nana, A.W.; Yang, P.M.; Lin, H.Y. Overview of Transforming Growth Factor beta Superfamily Involvement in Glioblastoma Initiation and Progression. Asian Pac. J. Cancer Prev. 2015, 16, 6813–6823. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shi, Y.; Massague, J. Mechanisms of TGF-β signaling from cell membrane to the nucleus. Cell 2003, 113, 685–700. [Google Scholar] [CrossRef]
- Ikushima, H.; Miyazono, K. Cellular context-dependent “colors” of transforming growth factor-β signaling. Cancer Sci. 2010, 101, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Iwatsuki, M.; Mimori, K.; Yokobori, T.; Ishi, H.; Beppu, T.; Nakamori, S.; Baba, H.; Mori, M. Epithelial-mesenchymal transition in cancer development and its clinical significance. Cancer Sci. 2010, 101, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Padua, D.; Massague, J. Roles of TGFβ in metastasis. Cell Res. 2009, 19, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, I.; Fernando, J.; Mainez, J.; Sancho, P. TGF-β signaling in cancer treatment. Curr. Pharm. Des. 2014, 20, 2934–2947. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J. TGFβ in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Ikushima, H.; Miyazono, K. TGF-beta signal transduction spreading to a wider field: A broad variety of mechanisms for context-dependent effects of TGF-β. Cell Tissue Res. 2012, 347, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Levy, L.; Hill, C.S. Alterations in components of the TGF-β superfamily signaling pathways in human cancer. Cytokine Growth Factor Rev. 2006, 17, 41–58. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, K.; Shia, A.; Cavicchioli, F.; Haley, V.; Comino, A.; Merlano, M.; Mauri, F.; Walter, K.; Lackner, M.; Wischnewsky, M.B.; et al. Identification of Endoglin as an epigenetically regulated tumour-suppressor gene in lung cancer. Br. J. Cancer 2015, 113, 970–978. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kuo, K.K.; Jian, S.F.; Li, Y.J.; Wan, S.W.; Weng, C.C.; Fang, K.; Wu, D.C.; Cheng, K.H. Epigenetic inactivation of transforming growth factor-beta1 target gene HEYL, a novel tumor suppressor, is involved in the P53-induced apoptotic pathway in hepatocellular carcinoma. Hepatol. Res. 2015, 45, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kadota, M.; Tang, B.; Yang, H.H.; Yang, Y.A.; Shan, M.; Weng, J.; Welsh, M.A.; Flanders, K.C.; Nagano, Y.; et al. An integrated genomic approach identifies persistent tumor suppressive effects of transforming growth factor-β in human breast cancer. Breast Cancer Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, R.; Tas, F.; Yasasever, C.T.; Aksit, E.; Karabulut, S.; Sen, F.; Keskin, S.; Kilic, L.; Yildiz, I.; Bozbey, H.U.; et al. High serum transforming growth factor β 1 (TGFB1) level predicts better survival in breast cancer. Tumour Biol. 2014, 35, 6941–6948. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H.; Sohn, M.; Lim, H.J.; Yoon, S.; Oh, H.; Shin, S.; Shin, J.H.; Oh, S.H.; Kim, J.; Lee, D.K.; et al. Ahnak functions as a tumor suppressor via modulation of TGFβ/Smad signaling pathway. Oncogene 2014, 33, 4675–4684. [Google Scholar] [CrossRef] [PubMed]
- Polyak, K.; Weinberg, R.A. Transitions between epithelial and mesenchymal states: Acquisition of malignant and stem cell traits. Nat. Rev. Cancer 2009, 9, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, K. Transforming growth factor-beta signaling in epithelial-mesenchymal transition and progression of cancer. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 314–323. [Google Scholar] [CrossRef] [PubMed]
- De Craene, B.; Berx, G. Regulatory networks defining EMT during cancer initiation and progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.F.; Miao, Q.; Zeng, X.Q.; Luo, T.C.; Ma, L.L.; Liu, Y.M.; Lian, J.J.; Gao, H.; Chen, S.Y. Transforming growth factor-β1 and -β2 in gastric precancer and cancer and roles in tumor-cell interactions with peripheral blood mononuclear cells in vitro. PLoS ONE 2013, 8, e54249. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Kasuya, K.; Tsuchida, A.; Mimuro, A.; Inoue, K.; Aoki, T.; Aoki, T.; Koyanagi, Y. Immunohistochemical analysis of transforming growth factor β in gallbladder cancer. Oncol. Rep. 2003, 10, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Fukai, Y.; Fukuchi, M.; Masuda, N.; Osawa, H.; Kato, H.; Nakajima, T.; Kuwano, H. Reduced expression of transforming growth factor-β receptors is an unfavorable prognostic factor in human esophageal squamous cell carcinoma. Int. J. Cancer. 2003, 104, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shady, M.; Baer, H.U.; Friess, H.; Berberat, P.; Zimmermann, A.; Graber, H.; Gold, L.I.; Korc, M.; Buchler, M.W. Transforming growth factor betas and their signaling receptors in human hepatocellular carcinoma. Am. J. Surg. 1999, 177, 209–215. [Google Scholar] [CrossRef]
- Tsushima, H.; Kawata, S.; Tamura, S.; Ito, N.; Shirai, Y.; Kiso, S.; Imai, Y.; Shimomukai, H.; Nomura, Y.; Matsuda, Y.; et al. High levels of transforming growth factor beta 1 in patients with colorectal cancer: Association with disease progression. Gastroenterology 1996, 110, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Friess, H.; Yamanaka, Y.; Buchler, M.; Ebert, M.; Beger, H.G.; Gold, L.I.; Korc, M. Enhanced expression of transforming growth factor beta isoforms in pancreatic cancer correlates with decreased survival. Gastroenterology 1993, 105, 1846–1856. [Google Scholar] [CrossRef]
- Voon, D.C.; Wang, H.; Koo, J.K.; Chai, J.H.; Hor, Y.T.; Tan, T.Z.; Chu, Y.S.; Mori, S.; Ito, Y. EMT-induced stemness and tumorigenicity are fueled by the EGFR/Ras pathway. PLoS ONE 2013, 8, e70427. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Q.; Li, K.; Chen, L.; Li, W.; Hou, M.; Liu, T.; Yang, J.; Lindvall, C.; Bjorkholm, M.; et al. Telomerase reverse transcriptase promotes epithelial-mesenchymal transition and stem cell-like traits in cancer cells. Oncogene 2013, 32, 4203–4213. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Harada, K.; Itatsu, K.; Ikeda, H.; Kakuda, Y.; Shimomura, S.; Ren, X.S.; Yoneda, N.; Sasaki, M.; Nakanuma, Y. Epithelial-mesenchymal transition induced by transforming growth factor-β1/Snail activation aggravates invasive growth of cholangiocarcinoma. Am. J. Pathol. 2010, 177, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.J.; Yun, B.R.; Kwon, J.H.; Ahn, H.S.; Seol, M.A.; Lee, M.J.; Yu, G.R.; Yu, H.C.; Hong, B.; Choi, K.; et al. Genetic and expression alterations in association with the sarcomatous change of cholangiocarcinoma cells. Exp. Mol. Med. 2009, 41, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Ween, M.P.; Oehler, M.K.; Ricciardelli, C. Transforming Growth Factor-β-Induced Protein (TGFBI)/(βig-H3): A Matrix Protein with Dual Functions in Ovarian Cancer. Int. J. Mol. Sci. 2012, 13, 10461–10477. [Google Scholar] [CrossRef] [PubMed]
- Skonier, J.; Neubauer, M.; Madisen, L.; Bennett, K.; Plowman, G.D.; Purchio, A.F. cDNA cloning and sequence analysis of βig-h3, a novel gene induced in a human adenocarcinoma cell line after treatment with transforming growth factor-β. DNA Cell Biol. 1992, 11, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Han, K.E.; Choi, S.I.; Kim, T.I.; Maeng, Y.S.; Stulting, R.D.; Ji, Y.W.; Kim, E.K. Pathogenesis and treatments of TGFBI corneal dystrophies. Prog. Retin. Eye Res. 2016, 50, 67–88. [Google Scholar] [CrossRef] [PubMed]
- Cha, D.R.; Kim, I.S.; Kang, Y.S.; Han, S.Y.; Han, K.H.; Shin, C.; Ji, Y.H.; Kim, N.H. Urinary concentration of transforming growth factor-β-inducible gene-h3(βig-h3) in patients with Type 2 diabetes mellitus. Diabet. Med. 2005, 22, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lim, S.W.; Choi, B.S.; Lee, S.H.; Cha, J.H.; Kim, I.S.; Kim, J.; Yang, C.W. Inhibitory effect of pravastatin on transforming growth factor β1-inducible gene h3 expression in a rat model of chronic cyclosporine nephropathy. Am. J. Nephrol. 2005, 25, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.J.; Kim, M.O.; Kim, S.O.; Park, J.; Kwon, Y.K.; Kim, I.S.; Lee, E.H. Induction of TGF-β-inducible gene-h3 (βig-h3) by TGF-β1 in astrocytes: Implications for astrocyte response to brain injury. Brain Res. Mol. Brain Res. 2002, 107, 57–64. [Google Scholar] [CrossRef]
- O’Brien, E.R.; Bennett, K.L.; Garvin, M.R.; Zderic, T.W.; Hinohara, T.; Simpson, J.B.; Kimura, T.; Nobuyoshi, M.; Mizgala, H.; Purchio, A.; et al. βig-h3, a transforming growth factor-β-inducible gene, is overexpressed in atherosclerotic and restenotic human vascular lesions. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, D.; Yokobori, T.; Sohda, M.; Sakai, M.; Hara, K.; Honjo, H.; Kato, H.; Miyazaki, T.; Kuwano, H. TGFBI Expression in Cancer Stromal Cells is Associated with Poor Prognosis and Hematogenous Recurrence in Esophageal Squamous Cell Carcinoma. Ann. Surg. Oncol. 2016, 23, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.I.; Maeng, Y.S.; Kim, K.S.; Kim, T.I.; Kim, E.K. Autophagy is induced by raptor degradation via the ubiquitin/proteasome system in granular corneal dystrophy type 2. Biochem. Biophys. Res. Commun. 2014, 450, 1505–1511. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.I.; Kim, E.K. Autophagy in granular corneal dystrophy type 2. Exp. Eye Res. 2016, 144, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.I.; Jin, J.Y.; Maeng, Y.S.; Kim, T.I.; Kim, E.K. TGF-β regulates TGFBIp expression in corneal fibroblasts via miR-21, miR-181a, and Smad signaling. Biochem. Biophys. Res. Commun. 2016, 472, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Dokmanovic, M.; Chang, B.D.; Fang, J.; Roninson, I.B. Retinoid-induced growth arrest of breast carcinoma cells involves co-activation of multiple growth-inhibitory genes. Cancer Biol. Ther. 2002, 1, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Nam, E.J.; Sa, K.H.; You, D.W.; Cho, J.H.; Seo, J.S.; Han, S.W.; Park, J.Y.; Kim, S.I.; Kyung, H.S.; Kim, I.S.; et al. Up-regulated transforming growth factor β-inducible gene h3 in rheumatoid arthritis mediates adhesion and migration of synoviocytes through αvβ3 integrin: Regulation by cytokines. Arthritis Rheum. 2006, 54, 2734–2744. [Google Scholar] [CrossRef] [PubMed]
- Gratchev, A.; Guillot, P.; Hakiy, N.; Politz, O.; Orfanos, C.E.; Schledzewski, K.; Goerdt, S. Alternatively activated macrophages differentially express fibronectin and its splice variants and the extracellular matrix protein βIG-H3. Scand. J. Immunol. 2001, 53, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Tumbarello, D.A.; Andrews, M.R.; Brenton, J.D. SPARC Regulates Transforming Growth Factor Beta Induced (TGFBI) Extracellular Matrix Deposition and Paclitaxel Response in Ovarian Cancer Cells. PLoS ONE 2016, 11, e0162698. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.I.; Lee, E.; Jeong, J.B.; Akuzum, B.; Maeng, Y.S.; Kim, T.I.; Kim, E.K. 4-Phenylbutyric acid reduces mutant-TGFBIp levels and ER stress through activation of ERAD pathway in corneal fibroblasts of granular corneal dystrophy type 2. Biochem. Biophys. Res. Commun. 2016, 477, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Hung, M.S.; Chen, I.C.; You, L.; Jablons, D.M.; Li, Y.C.; Mao, J.H.; Xu, Z.; Lung, J.H.; Yang, C.T.; Liu, S.T. Knockdown of cullin 4A inhibits growth and increases chemosensitivity in lung cancer cells. J. Cell. Mol. Med. 2016, 20, 1295–1306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wen, G.; Shao, G.; Wang, C.; Lin, C.; Fang, H.; Balajee, A.S.; Bhagat, G.; Hei, T.K.; Zhao, Y. TGFBI deficiency predisposes mice to spontaneous tumor development. Cancer Res. 2009, 69, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Cai, H.; Chen, Y.; Hu, B.; Luo, H.; Wu, Y.; Wu, J. The role of TGFBI (βig-H3) in gastrointestinal tract tumorigenesis. Mol. Cancer 2015. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Semba, S.; Aoyagi, K.; Sasaki, H.; Yokozaki, H. Mesenchymal stem cells provide an advantageous tumor microenvironment for the restoration of cancer stem cells. Pathobiology 2012, 79, 290–306. [Google Scholar] [CrossRef] [PubMed]
- Turtoi, A.; Musmeci, D.; Wang, Y.; Dumont, B.; Somja, J.; Bevilacqua, G.; de Pauw, E.; Delvenne, P.; Castronovo, V. Identification of novel accessible proteins bearing diagnostic and therapeutic potential in human pancreatic ductal adenocarcinoma. J. Proteome Res. 2011, 10, 4302–4313. [Google Scholar] [CrossRef] [PubMed]
- Patry, M.; Teinturier, R.; Goehrig, D.; Zetu, C.; Ripoche, D.; Kim, I.S.; Bertolino, P.; Hennino, A. βig-h3 Represses T-Cell Activation in Type 1 Diabetes. Diabetes 2015, 64, 4212–4219. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Nie, S.; Wu, J.; Lubman, D.M. Target proteomic profiling of frozen pancreatic CD24+ adenocarcinoma tissues by immuno-laser capture microdissection and nano-LC-MS/MS. J. Proteome Res. 2013, 12, 2791–2804. [Google Scholar] [CrossRef] [PubMed]
- Jinawath, N.; Chamgramol, Y.; Furukawa, Y.; Obama, K.; Tsunoda, T.; Sripa, B.; Pairojkul, C.; Nakamura, Y. Comparison of gene expression profiles between Opisthorchis viverrini and non-Opisthorchis viverrini associated human intrahepatic cholangiocarcinoma. Hepatology 2006, 44, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Buckhaults, P.; Rago, C.; Croix, B.S.; Romans, K.E.; Saha, S.; Zhang, L.; Vogelstein, B.; Kinzler, K.W. Secreted and cell surface genes expressed in benign and malignant colorectal tumors. Cancer Res. 2001, 61, 6996–7001. [Google Scholar] [PubMed]
- Turtoi, A.; Blomme, A.; Debois, D.; Somja, J.; Delvaux, D.; Patsos, G.; di Valentin, E.; Peulen, O.; Mutijima, E.N.; de Pauw, E.; et al. Organized proteomic heterogeneity in colorectal cancer liver metastases and implications for therapies. Hepatology 2014, 59, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Rong, Y.; Radiloff, D.R.; Datto, M.B.; Centeno, B.; Bao, S.; Cheng, A.W.; Lin, F.; Jiang, S.; Yeatman, T.J.; et al. Extracellular matrix protein βig-h3/TGFBI promotes metastasis of colon cancer by enhancing cell extravasation. Genes Dev. 2008, 22, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Fakhrai, H.; Mantil, J.C.; Liu, L.; Nicholson, G.L.; Murphy-Satter, C.S.; Ruppert, J.; Shawler, D.L. Phase I clinical trial of a TGF-β antisense-modified tumor cell vaccine in patients with advanced glioma. Cancer Gene Ther. 2006, 13, 1052–1060. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Melisi, D.; Ishiyama, S.; Sclabas, G.M.; Fleming, J.B.; Xia, Q.; Tortora, G.; Abbruzzese, J.L.; Chiao, P.J. LY2109761, a novel transforming growth factor β receptor type I and type II dual inhibitor, as a therapeutic approach to suppressing pancreatic cancer metastasis. Mol. Cancer Ther. 2008, 7, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Nemunaitis, J.; Dillman, R.O.; Schwarzenberger, P.O.; Senzer, N.; Cunningham, C.; Cutler, J.; Tong, A.; Kumar, P.; Pappen, B.; Hamilton, C.; et al. Phase II study of belagenpumatucel-L, a transforming growth factor beta-2 antisense gene-modified allogeneic tumor cell vaccine in non-small-cell lung cancer. J. Clin. Oncol. 2006, 24, 4721–4730. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, G.; Mikulits, W.; Dooley, S.; Fabregat, I.; Moustakas, A.; Dijke, P.t.; Portincasa, P.; Winter, P.; Janssen, R.; Leporatti, S.; et al. The rationale for targeting TGF-β in chronic liver diseases. Eur. J. Clin. Investig. 2016, 46, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Giaccone, G.; Bazhenova, L.A.; Nemunaitis, J.; Tan, M.; Juhasz, E.; Ramlau, R.; van den Heuvel, M.M.; Lal, R.; Kloecker, G.H.; Eaton, K.D.; et al. A phase III study of belagenpumatucel-L, an allogeneic tumour cell vaccine, as maintenance therapy for non-small cell lung cancer. Eur. J. Cancer 2015, 51, 2321–2329. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.C.; Tan, A.R.; Olencki, T.E.; Shapiro, G.I.; Dezube, B.J.; Reiss, M.; Hsu, F.J.; Berzofsky, J.A.; Lawrence, D.P. Phase I study of GC1008 (fresolimumab): A human anti-transforming growth factor-beta (TGFβ) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PLoS ONE 2014, 9, e90353. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.; Mills, A.D.; Ibrahim, A.E.; Temple, J.; Blenkiron, C.; Vias, M.; Massie, C.E.; Iyer, N.G.; McGeoch, A.; Crawford, R.; et al. The extracellular matrix protein TGFBI induces microtubule stabilization and sensitizes ovarian cancers to paclitaxel. Cancer Cell 2007, 12, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Irigoyen, M.; Pajares, M.J.; Agorreta, J.; Ponz-Sarvise, M.; Salvo, E.; Lozano, M.D.; Pio, R.; Gil-Bazo, I.; Rouzaut, A. TGFBI expression is associated with a better response to chemotherapy in NSCLC. Mol. Cancer 2010. [Google Scholar] [CrossRef] [PubMed]
- Nummela, P.; Lammi, J.; Soikkeli, J.; Saksela, O.; Laakkonen, P.; Holtta, E. Transforming growth factor beta-induced (TGFBI) is an anti-adhesive protein regulating the invasive growth of melanoma cells. Am. J. Pathol. 2012, 180, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.; Partridge, M.A.; Li, B.; Hong, M.; Liao, W.; Cheng, S.K.; Zhao, Y.; Calaf, G.M.; Liu, T.; Zhou, J.; et al. TGFBI expression reduces in vitro and in vivo metastatic potential of lung and breast tumor cells. Cancer Lett. 2011, 308, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wen, G.; Zhao, Y.; Tong, J.; Hei, T.K. The role of TGFBI in mesothelioma and breast cancer: Association with tumor suppression. BMC Cancer 2012, 12, 239. [Google Scholar] [CrossRef] [PubMed]
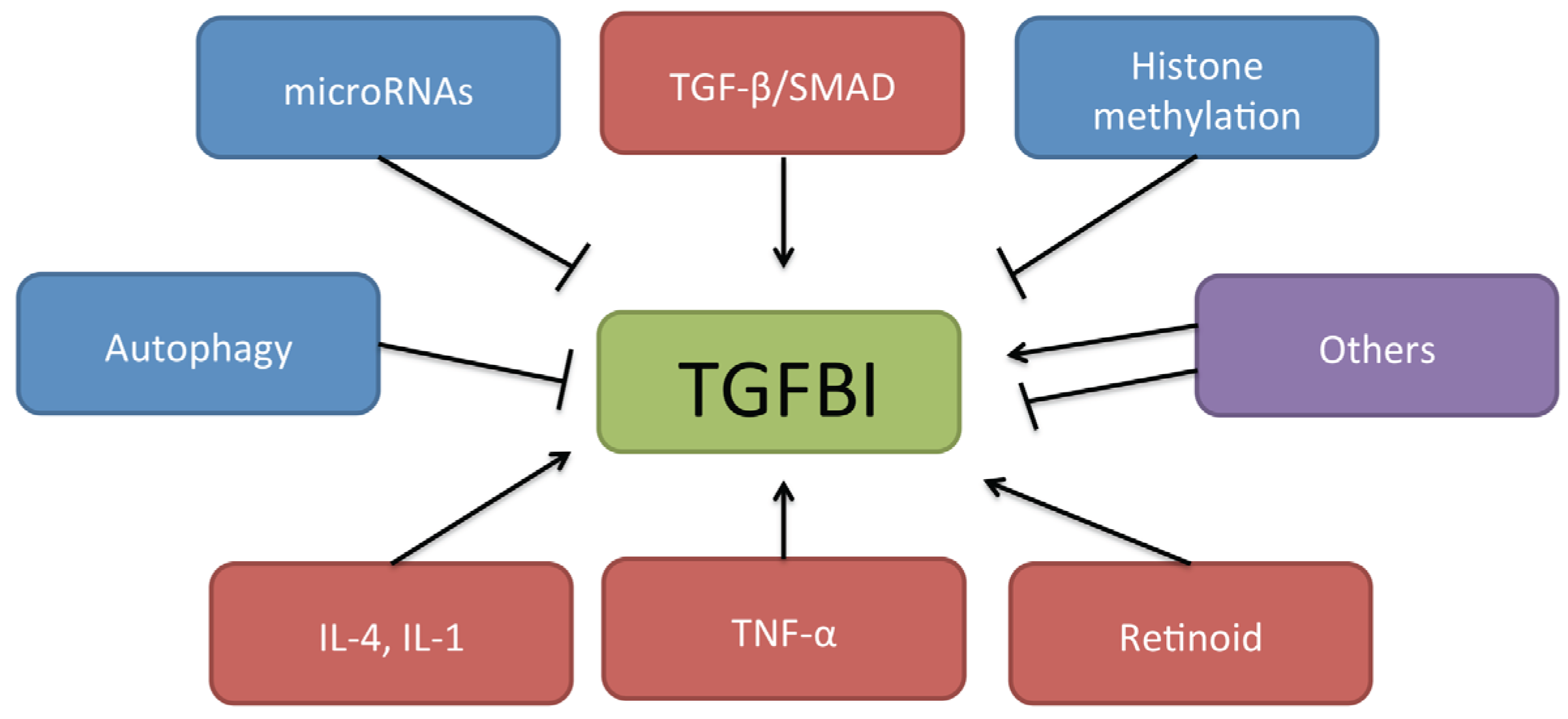

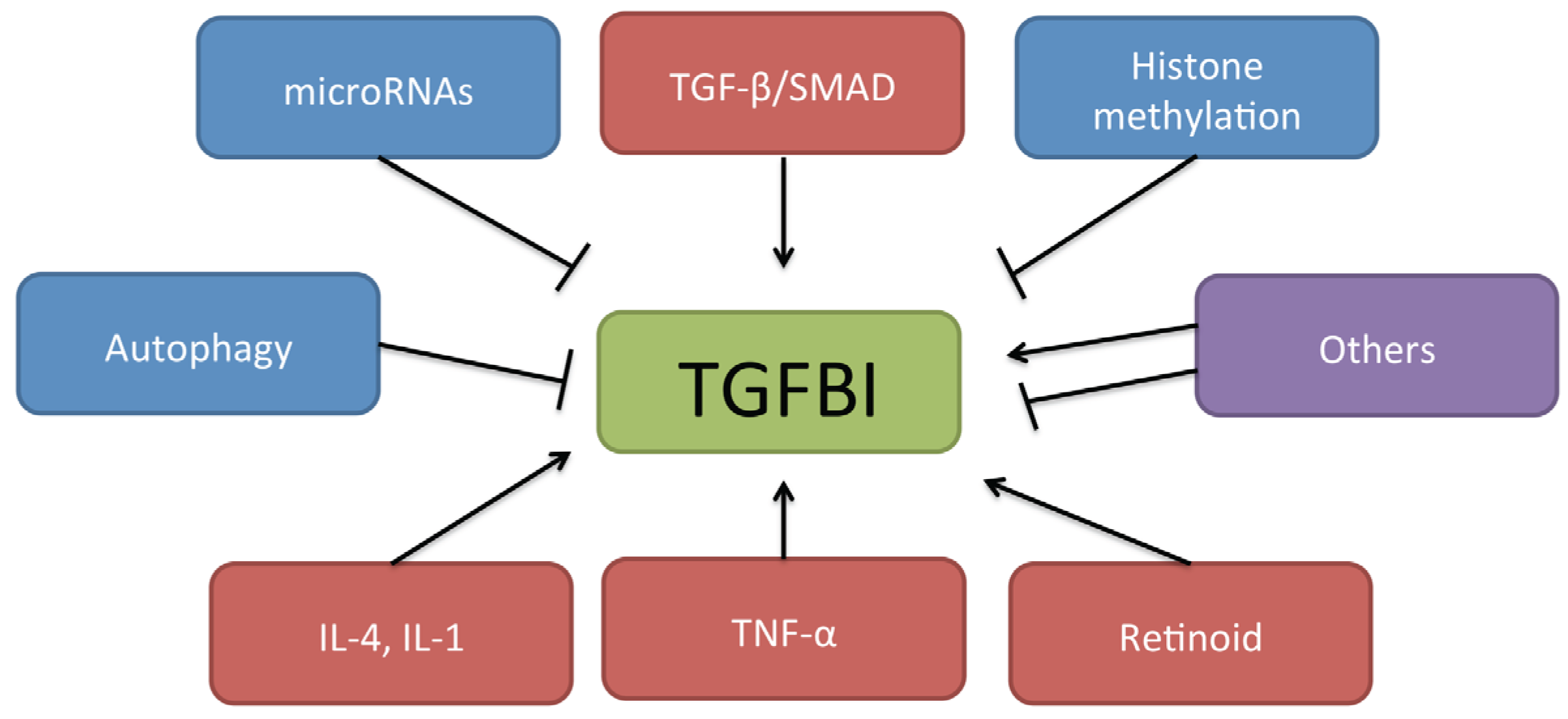{kind=link}
| Tumor Type | Significance | PMID Reference |
|---|---|---|
| Esophageal tumor | TGFBI expression was higher in the extracellular matrix (ECM) of tumors as compared to normal tissues. | 19082484 |
| High TGFBI was associated with tumor progression and poor prognosis in esophageal cancer. | 25448803 | |
| Gastric tumor | TGFBI overexpression in stomach tissue of TGFBI knock-in mice caused gastric tumors. Serum TGFBI level was higher in gastric cancer than in non-cancer patients. | 25889002 |
| Bone-marrow-derived mesenchymal stem cells induced TGFBI expression in co-cultured gastric cancer cells. | 22688186 | |
| Pancreatic and biliary tumor | Serum TGFBI expression was higher in pancreatic and biliary carcinoma patients than in non-cancer patients. | 25889002 |
| TGFBI expression in pancreas inhibited T-cell activation and production of cytotoxic molecules including granzyme B and interferon-γ via suppression of Lck tyrosine kinase. | 26470788 | |
| TGFBI was overexpressed in pancreatic cancer stem cells. | 23679566 | |
| Proteomic analysis revealed that TGFBI was upregulated in pancreatic cancer as compared to non-cancer tissues. | 21755970 | |
| TGF-β induced TGFBI mRNA expression in pancreatic cancer cells as compared to normal control tissue. | 12379307 | |
| TGFBI was overexpressed in non-Opisthorchis viverrini-related intrahepatic cholangiocarcinoma. | 17006947 | |
| TGFBI upregulation was associated with sarcomatous changes such as EMT induction in cholangiocarcinoma. | 19287191 | |
| Colorectal tumor | TGFBI mRNA level was higher in colon cancer than in adenomas and non-cancerous tissue. | 11585723 |
| TGFBI secretion in colon cancer cells was found to be related to cancer aggressiveness and extravasation. | 18245446 | |
| Stromal TGFBI expression was higher in metastatic as compared to normal liver tissue. TGFBI expression was localized in the center part of the metastatic area. | 23832580 |
© 2017 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yokobori, T.; Nishiyama, M. TGF-β Signaling in Gastrointestinal Cancers: Progress in Basic and Clinical Research. J. Clin. Med. 2017, 6, 11. https://doi.org/10.3390/jcm6010011
Yokobori T, Nishiyama M. TGF-β Signaling in Gastrointestinal Cancers: Progress in Basic and Clinical Research. Journal of Clinical Medicine. 2017; 6(1):11. https://doi.org/10.3390/jcm6010011
Chicago/Turabian StyleYokobori, Takehiko, and Masahiko Nishiyama. 2017. "TGF-β Signaling in Gastrointestinal Cancers: Progress in Basic and Clinical Research" Journal of Clinical Medicine 6, no. 1: 11. https://doi.org/10.3390/jcm6010011
APA StyleYokobori, T., & Nishiyama, M. (2017). TGF-β Signaling in Gastrointestinal Cancers: Progress in Basic and Clinical Research. Journal of Clinical Medicine, 6(1), 11. https://doi.org/10.3390/jcm6010011