
Unexpectedly High and Difficult-to-Explain Regenerative Capacity in an 82-Year-Old Patient with Insulin-Requiring Type 2 Diabetes and End-Stage Renal Disease


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. The Case
1.2. General Research Plan
- -
- the genetic study of the intestinal microbiota, in order to identify possible microbial alterations and their impact on the immunological behavior and underlying pathology of the patients;
- -
- the complex study of innate immunity (inflammation) and acquired immunity (RIC);
- -
- the study of tissue regeneration capacity.
2. Methods
2.1. The Genetic Study of the Gut Microbiota
Statistical Analysis
2.2. The Immunological Study
2.3. The Study of Tissue Regeneration Capacity
Statistical Analysis
3. Results
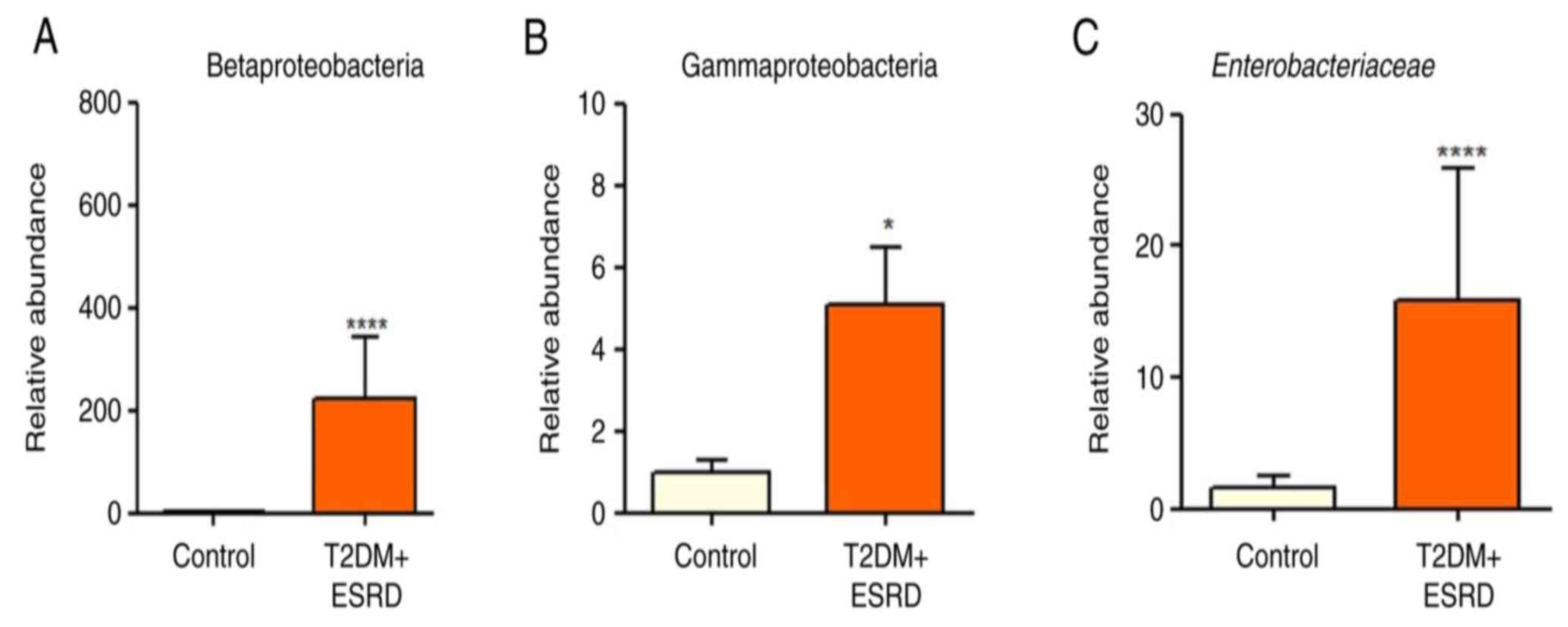
3.1. The Genetic Study of the Intestinal Microbiota Results
3.2. The Results of the Immunological Study
3.3. The Results of Tissue Regeneration Capacity Study
4. Discussion
4.1. The Genetic Analysis of the Gut Microbiota
4.2. The Study of Immunological Pattern
4.2.1. The Study of Innate Immunity
- a.
- IL-6 is the prototypical cytokine of a large family of signal molecules, which also include IL-11, LIF (leukemia inhibitory factor), OSM (oncostatin M), CNTF (ciliary inhibitory factor), CT-1 (cardiotrophin 1), NNT-1 (cardiotrophin-like related cytokine and stimulating neurotrophin1/B-cell stimulating factor 3), NPN (neuropoietin), IL-27, and IL-31 [11].
- b.
- The serum level of IL-1β also offered surprises. Thus, we obtained serum levels that were approximately 6-fold higher than the mean of the test group (T2DM) and 26 times higher than the mean value of the control group (ESRD).
- -
- -
- -
- -
- stimulates the synthesis of arachidonic acid metabolites and the release of chondrocytes and other cell types from fibroblasts [31];
- -
- -
- -
- -
- c.
- TNFα was also a major surprise. The patient’s TNFα serum level was 5.2 times lower than the test group’s mean value and 5.5 times lower than the control group’s. We must note that the mean value of the test group (T2DM+ESRD) was close to that of the control group (ESRD). The initial, intermediary conclusion is that the presence of T2DM does not amplify TNFα’s pro-inflammatory effect.
- d.
- The serum level of IL10, the prototypical anti-inflammatory cytokine, was 9.4 times higher than the test group’s mean and 10.88 times higher than the average of the control group.
- -
- Inhibition of gene reading for pro-inflammatory proteins in monocytes, macrophages, and dendritic cells, with the consequent inhibition of IL-1β, IL-1α, TNFα, IL-6, IL-12, and IL-18 secretion, as well as of chemokines CCL2, CCL5, CCL12, IL-8, CXCL2, and CXCL10 [58];
- -
- Stimulates the synthesis of the IL-1R antagonist (IL-1Ra), which blocks IL-1β signaling [82];
- -
- -
- Inhibits IL-2 and IFNγ release from TH1 cells [84];
- -
- Suppresses the ability of APCs to present MHCII-epitope complexes to TH1 and TH2 cells [85];
- -
- Inhibits the expression of the intercellular adhesion molecule ICAM-1, the ligand CD80 (B7), and the CD 56 receptor on the APC membrane, thus inhibiting the formation of the immune synapse between the APC and T cells (TH and cytotoxic T cells, respectively) and the complete activation of T cells [86,87,88];
- -
- -
- -
- -
- It strongly stimulates B lymphocytes to pass in the intermediate stage of activation, i.e., to become B lymphoblasts (BLb);
- -
- -
- -
- Stimulates the proliferation of thymic cells [99];
- -
- Stimulates CD8+T cells (cytotoxic T lymphocytes) by increasing their expression of MHCI molecules and the secretion of IFNγ and granzymes [100];
- -
- -
- Stimulates mast cells [105].
- -
- Reduces neuronal injury during infections, inflammation, ischemia, and trauma;
- -
- Increases the lifespan of neurons and axonal regeneration;
- -
- Modulates neurogenesis in adults.
4.2.2. The Study of Acquired Immunity
- -
- stimulates the development of lymphoid organs. It is experimentally proven that TNFβ stimulates the formation of intestinal Payer’s plaques through TNFR1 [117];
- -
- Stimulation of the expression of intercellular adhesion molecules (ICAMs) and E-selectin on endothelial cell membranes [129]. This aspect has been clearly demonstrated for the endothelial cells of the inflamed pancreas and kidney [130] and is of major importance for our study addressing the associated pathologies of these two organs. Furthermore, this exposure of adhesion molecules was shown to be independent of cytokines secreted by T and B cells;
- LTα induces the expression of the chemokines RANTES (regulated upon activation normal T cell expressed and secreted) and MCP-1 (monocyte chemoattractant protein 1) on murine endothelial cells [131];
- Stimulates the synthesis of pro-inflammatory cytokines from macrophages at the same level as TNFα [132];
- Stimulates inflammatory lymphangiogenesis [133].
4.3. Tissue Regeneration Capacity in Patients with T2DM and ESRD
- a.
- Overall, results showed decreased values of serum NT-3 and VEGFβ levels in both groups. However, the mean serum level of NT-3 was approx. eight times higher, and that of VEGFβ was about seventeen times higher in the test group compared to the control group [4]. This proves the necessity of increased tissue regeneration in diabetes patients.
- b.
- Because both chronic hemodialysis and diabetic angiopathy predispose to the occurrence of vascular and tissue damage, we also monitored the regeneration capacity of these structures in the hemodialysis patients included in this study.
- (1)
- some that have direct angiogenic actions, such as vascular endothelial growth factor (VEGF) and fibroblastic growth factor (acidic (aFGF/FGF1) and basic (bFGF/FGF2));
- (2)
- others that have indirect angiogenic actions, such as TNFα and transforming growth factor β (TGF-β).
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- New Global Kidney Health Report Sheds Light on Current Capacity Around the World to Deliver Kidney Care. Available online: https://www.theisn.org/blog/2023/03/30/new-global-kidney-health-report-sheds-light-on-current-capacity-around-the-world-to-deliver-kidney-care/ (accessed on 28 March 2025).
- Monardo, P.; Lacquaniti, A.; Campo, S.; Bucca, M.; Casuscelli di Tocco, T.; Rovito, S.; Ragusa, A.; Santoro, A. Updates on hemodialysis techniques with a common denominator: The personalization of the dialytic therapy. Semin. Dial. 2021, 34, 183–195. [Google Scholar] [PubMed]
- Wang, Y.; Gao, L. Inflammation and Cardiovascular Disease Associated With Hemodialysis for End-Stage Renal Disease. Front. Pharmacol. 2022, 13, 800950. [Google Scholar] [CrossRef] [PubMed]
- Trandafir, M.F.; Savu, O.; Pasarica, D.; Bleotu, C.; Gheorghiu, M. Interleukin-6 as a Director of Immunological Events and Tissue Regenerative Capacity in Hemodialyzed Diabetes Patients. Med. Sci. 2024, 12, 31. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Trandafir, M.; Pircalabioru, G.G.; Savu, O. Microbiota analysis in individuals with type two diabetes mellitus and end-stage renal disease: A pilot study. Exp. Ther. Med. 2024, 27, 211. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A common factor in human diseases. Biomed. Res. Int. 2017, 2017, 9351507. [Google Scholar]
- Dang, H.; Chen, R.; Wang, L.; Shao, S.; Dai, L.; Ye, Y.; Guo, L.; Huang, G.; Klotz, M.G. Molecular characterization of putative biocorroding microbiota with a novel niche detection of Epsilon- and Zetaproteobacteria in Pacific Ocean coastal seawaters. Environ. Microbiol. 2011, 13, 3059–3074. [Google Scholar]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Grebenciucova, E.; VanHaerents, S. Interleukin 6: At the interface of human health and disease. Front. Immunol. 2023, 14, 1255533. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bello, A.; Kawar, B.; El Kossi, M.; El Nahas, M. Chapter 75—Epidemiology and Pathophysiology of Chronic Kidney Disease. In Comprehensive Clinical Nephrology, 4th ed.; Johnson, R.J., Feehally, J., Floege, J., Eds.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2010; pp. 907–918. ISBN 9780323058766. [Google Scholar] [CrossRef]
- Avraham, S.; Korin, B.; Chung, J.J.; Oxburgh, L.; Shaw, A.S. The Mesangial cell—The glomerular stromal cell. Nat. Rev. Nephrol. 2021, 17, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Hibi, M.; Nakagawa, N.; Nakagawa, T.; Yasukawa, K.; Yamanishi, K.; Taga, T.; Kishimoto, T. IL-6-induced homodimerization of gp130 and associated activation of a tyrosine kinase. Science 1993, 260, 1808–1810. [Google Scholar] [CrossRef]
- Kishimoto, T.; Taga, T.; Akira, S. Cytokine signal transduction. Cell 1994, 76, 253–262. [Google Scholar] [CrossRef]
- Lust, J.A.; Donovan, K.A.; Kline, M.P.; Greipp, P.R.; Kyle, R.A.; Maihle, N.J. Isolation of an mRNA encoding a soluble form of the human interleukin-6 receptor. Cytokine 1992, 4, 96–100. [Google Scholar] [CrossRef]
- Schumertl, T.; Lokau, J.; Rose-John, S.; Garbers, C. Function and proteolytic generation of the soluble interleukin-6 receptor in health and disease. Biochim. Biophys. Acta BBA—Mol. Cell Res. 2022, 1869, 119143. [Google Scholar] [CrossRef]
- Heink, S.; Yogev, N.; Garbers, C.; Herwerth, M.; Aly, L.; Gasperi, C.; Husterer, V.; Croxford, A.L.; Möller-Hackbarth, K.; Bartsch, H.S.; et al. Transpresentation of IL-6 by dendritic cells is required for the priming of pathogenic TH17 cells. Nat. Immunol. 2017, 18, 74–85. [Google Scholar] [CrossRef]
- Di Santo, E.; Alonzi, T.; Poli, V.; Fattori, E.; Toniatti, C.; Sironi, M.; Ricciardi-Castagnoli, P.; Ghezzi, P. Differential effects of il-6 on systemic and central production of tnf: A study with il-6-deficient mice. Cytokine 1997, 9, 300–306. [Google Scholar] [CrossRef]
- Schindler, R.; Mancilla, J.; Endres, S.; Ghorbani, R.; Clark, C.S.; Dinarello, A.D. Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumour necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and, TNF. Blood 1990, 75, 40–47. [Google Scholar] [CrossRef]
- Bennett, I.L., Jr.; Beeson, P.B. Studies on the Pathogenesis of Fever: I. The Effect of the Injection of Extracts and Suspensions of Uninfected Rabbit Tissues Upon the Body Temperature of Normal Rabbits. J. Exp. Med. 1953, 98, 477–492. [Google Scholar] [CrossRef]
- Bodel, P. Studies on the Mechanism of Endogenous Pyrogen Production: III. Human Blood Monocytes. J. Exp. Med. 1974, 140, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Atkins, E. Fever: The Old and the New. J. Infect. Dis. 1984, 149, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Infection, Fever, and Exogenous and Endogenous Pyrogens: Some Concepts Have Changed. J. Endotoxin Res. 2004, 10, 201–222. [Google Scholar] [CrossRef]
- Cybulsky, M.I.; Colditz, I.G.; Movat, H.Z. The Role of Interleukin-1 in Neutrophil Leukocyte Emigration Induced by Endotoxin. Am. J. Pathol. 1986, 124, 367–372. [Google Scholar]
- Dinarello, C.A. Interleukin-1 and the Pathogenesis of the Acute-Phase Response. N. Engl. J. Med. 1984, 311, 1413–1418. [Google Scholar] [CrossRef]
- Pike, B.L.; Nossal, G.J.V. Interleukin 1 can Act as a B-Cell Growth and Differentiation Factor. Proc. Natl. Acad. Sci. USA 1985, 82, 8153–8157. [Google Scholar] [CrossRef]
- Lowenthal, J.W.; Cerottini, J.-C.; MacDonald, H.R. Interleukin 1-Dependent Induction of Both Interleukin 2 Secretion and Interleukin 2 Receptor Expression by Thymoma Cells. J. Immunol. 1986, 137, 1226–1231. [Google Scholar] [CrossRef]
- Lichtman, A.H.; Chin, J.; Schmidt, J.A.; Abbas, A.K. Role of Interleukin 1 in the Activation of T Lymphocytes. Proc. Natl. Acad. Sci. USA 1988, 85, 9699–9703. [Google Scholar] [CrossRef]
- Saklatvala, J.; Pilsworth, L.M.; Sarsfield, S.J.; Gavrilovic, J.; Heath, J.K. Pig Catabolin is a Form of Interleukin 1. Cartilage and Bone Resorb, Fibroblasts Make Prostaglandin and Collagenase, and Thymocyte Proliferation is Augmented in Response to One Protein. Biochem. J. 1984, 224, 461–466. [Google Scholar] [CrossRef]
- Besedovsky, H.; del Rey, A.; Sorkin, E.; Dinarello, C.A. Immunoregulatory Feedback Between Interleukin-1 and Glucocorticoid Hormones. Science 1986, 233, 652–654. [Google Scholar] [CrossRef]
- del Rey, A.; Besedovsky, H. Interleukin 1 Affects Glucose Homeostasis. Am. J. Physiol. 1987, 22, R794–R798. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Villa, L.; Volpini, G.; Bossù, P.; Censini, S.; Ghiara, P.; Scapigliat, G.; Nencioni, L.; Bartalini, M.; Matteucci, G.; et al. Differential Activity of Interleukin 1a and Interleukin 1b in the Stimulation of the Immune Response In Vivo. Eur. J. Immunol. 1990, 20, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Ichinohe, T.; Lee, H.K.; Ogura, Y.; Flavell, R.; Iwasaki, A. Inflammasome Recognition of Influenza Virus is Essential for Adaptive Immune Responses. J. Exp. Med. 2009, 206, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.C.; Scull, M.A.; Moore, C.B.; Holl, E.K.; McElvania-TeKippe, E.; Taxman, D.J.; Guthrie, E.H.; Pickles, R.J.; Ting, J.P. The NLRP3 Inflammasome Mediates In Vivo Innate Immunity to Influenza A Virus Through Recognition of Viral, RNA. Immunity 2009, 30, 556–565. [Google Scholar] [CrossRef]
- Dinarello, C.A. The IL-1 Cytokine and Receptor Family. In Seminars in Immunology; John Wiley & Sons Ltd.: Oxford, UK, 2018; 247p. [Google Scholar]
- Dinarello, C.A. Overview of the IL-1 Family in Innate Inflammation and Acquired Immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef]
- Rivers-Auty, J.; Daniels, M.J.D.; Colliver, I.; Robertson, D.L.; Brough, D. Redefining the Ancestral Origins of the Interleukin-1 Superfamily. Nat. Commun. 2018, 9, 1156. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Weil, S.; Martin, M.U. The Family of Interleukin-1 Receptors. Immunol. Rev. 2018, 281, 197–232. [Google Scholar] [CrossRef]
- Touzani, O.; Boutin, H.; LeFeuvre, R.; Parker, L.; Miller, A.; Luheshi, G.; Rothwell, N. Interleukin-1 Influences Ischemic Brain Damage in the Mouse Independently of the Interleukin-1 Type I Receptor. J. Neurosci. 2002, 22, 38–43. [Google Scholar] [CrossRef]
- Ramsden, L.; Rider, C.C. Selective and Differential Binding of Interleukin (IL)-1a, IL-1b, IL-2 and IL-6 to Glycosaminoglycans. Eur. J. Immunol. 1992, 22, 3027–3031. [Google Scholar] [CrossRef]
- Cebo, C.; Dambrouck, T.; Maes, E.; Laden, C.; Strecker, G.; Michalski, J.-C.; Zanetta, J.P. Recombinant Human Interleukins IL-1a, IL-1b, IL-4, IL-6, and IL-7 Show Different and Specific Calcium-Independent Carbohydrate-Binding Properties. J. Biol. Chem. 2001, 8, 5685–5691. [Google Scholar] [CrossRef]
- Fukushima, K.; Hara-Kuge, S.; Ohkura, T.; Seko, A.; Ideo, H.; Inazu, T.; Yamashita, K. Lectin-Like Characteristics of Recombinant Human Interleukin-1beta Recognizing Glycans of the Glycosylphosphatidylinositol Anchor. J. Biol. Chem. 1997, 272, 10579–10584. [Google Scholar] [CrossRef] [PubMed]
- Tandai-Hiruma, M.; Endo, T.; Kobata, A. Detection of Novel Carbohydrate Binding Activity of Interleukin-1. J. Biol. Chem. 1999, 274, 4459–4466. [Google Scholar] [CrossRef] [PubMed]
- Vergoten, G.; Zanetta, J.P. Structural Differences Between the Putative Carbohydrate-Recognition Domains of Human IL-1 Alpha, IL-1 Beta and IL-1 Receptor Antagonist Obtained by in Silico Modelling. Glycoconj. J. 2007, 24, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Sandstedt, J.; Sandstedt, M.; Lundqvist, A.; Jansson, M.; Sopasakis, V.R.; Jeppsson, A.; Hultén, L.M. Human cardiac fibroblasts isolated from patients with severe heart failure are immune-competent cells mediating an inflammatory response. Cytokine 2019, 113, 319–325. [Google Scholar] [CrossRef]
- Lonnemann, G.; Shapiro, L.; Engler-Blum, G.; Müller, G.A.; Koch, K.M.; Dinarello, C.A. Cytokines in human renal interstitial fibrosis. I. Interleukin-1 is a paracrine growth factor for cultured fibrosis-derived kidney fibroblasts. Kidney Int. 1995, 47, 837–844. [Google Scholar]
- Vincenti, M.P.; Coon, C.I.; Lee, O.; Brinckerhoff, C.E. Regulation of collagenase gene expression by IL-1 beta requires transcriptional and post-transcriptional mechanisms. Nucleic Acids Res. 1994, 22, 4818–4827. [Google Scholar]
- Siamwala, J.H.; Pagano, F.S.; Dubielecka, P.M.; Ivey, M.J.; Guirao-Abad, J.P.; Zhao, A.; Chen, S.; Granston, H.; Jeong, J.Y.; Rounds, S.; et al. IL-1β-mediated adaptive reprogramming of endogenous human cardiac fibroblasts to cells with immune features during fibrotic remodeling. Commun. Biol. 2023, 6, 1200. [Google Scholar] [CrossRef]
- Holbrook, J.; Lara-Reyna, S.; Jarosz-Griffiths, H.; McDermott, M. Tumour necrosis factor signalling in health and disease. F1000Research 2019, 8, 111. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Faustman, D.; Davis, M. TNF receptor 2 pathway: Drug target for autoimmune diseases. Nat. Rev. Drug Discov. 2010, 9, 482–493. [Google Scholar] [CrossRef]
- Carpentier, I.; Coornaert, B.; Beyaert, R. Function and regulation of tumor necrosis factor receptor type 2. Curr. Med. Chem. 2004, 11, 2205–2212. [Google Scholar] [CrossRef]
- Nagashima, H.; Ishii, N.; So, T. Regulation of Interleukin-6 Receptor Signaling by TNF Receptor-Associated Factor 2 and 5 During Differentiation of Inflammatory CD4+ T Cells. Front. Immunol. 2018, 9, 1986. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; O’Garra, A. Il-10 family cytokines il-10 and il-22: From basic science to clinical translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef] [PubMed]
- Lobo-Silva, D.; Carriche, G.M.; Castro, A.G.; Roque, S.; Saraiva, M. Balancing the immune response in the brain: Il-10 and its regulation. J. Neuroinflamm. 2016, 13, 297. [Google Scholar] [CrossRef]
- Hulsmans, M.; Sager, H.B.; Roh, J.D.; Valero-Munoz, M.; Houstis, N.E.; Iwamoto, Y.; Sun, Y.; Wilson, R.M.; Wojtkiewicz, G.; Tricot, B.; et al. Cardiac macrophages promote diastolic dysfunction. J. Exp. Med. 2018, 215, 423–440. [Google Scholar] [CrossRef]
- Howes, A.; Gabryšová, L.; O’Garra, A. Role of IL-10 and the IL-10 Receptor in Immune Responses, Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Vuillefroy de Silly, R.; Ducimetiere, L.; Yacoub Maroun, C.; Dietrich, P.Y.; Derouazi, M.; Walker, P.R. Phenotypic switch of Cd8(+) T cells reactivated under hypoxia toward Il-10 secreting, poorly proliferative effector cells. Eur. J. Immunol. 2015, 45, 2263–2275. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Aljamaei, H.M.; Stadnyk, A.W. The production and function of endogenous interleukin-10 in intestinal epithelial cells and gut homeostasis. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1343–1352. [Google Scholar] [CrossRef]
- Mestrallet, G.; Rouas-Freiss, N.; LeMaoult, J.; Fortunel, N.O.; Martin, M.T. Skin immunity and tolerance: Focus on epidermal keratinocytes expressing hla-G. Front. Immunol. 2021, 12, 772516. [Google Scholar] [CrossRef]
- Piipponen, M.; Li, D.; Landen, N.X. The immune functions of keratinocytes in skin wound healing. Int. J. Mol. Sci. 2020, 21, 8790. [Google Scholar] [CrossRef]
- Ciazynska, M.; Olejniczak-Staruch, I.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibinska, M.; Lesiak, A. Ultraviolet radiation and chronic inflammation-molecules and mechanisms involved in skin carcinogenesis: A narrative review. Life 2021, 11, 326. [Google Scholar] [CrossRef]
- Itakura, E.; Huang, R.R.; Wen, D.R.; Paul, E.; Wunsch, P.H.; Cochran, A.J. Il-10 expression by primary tumor cells correlates with melanoma progression from radial to vertical growth phase and development of metastatic competence. Mod. Pathol. 2011, 24, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.; Wu, C.; Chen, J.; Wang, W. Relationship between il-10 and pd-L1 in esophageal carcinoma tissues and il-10 down-regulates pd-L1 expression Via met signaling pathway. J. Gastrointest. Oncol. 2020, 11, 337–355. [Google Scholar] [CrossRef] [PubMed]
- Labani-Motlagh, A.; Ashja-Mahdavi, M.; Loskog, A. The tumor microenvironment: A milieu hindering and obstructing antitumor immune responses. Front. Immunol. 2020, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- Herbeuval, J.P.; Lelievre, E.; Lambert, C.; Dy, M.; Genin, C. Recruitment of Stat3 for production of il-10 by colon carcinoma cells induced by macrophage-derived il-6. J. Immunol. 2004, 172, 4630–4636. [Google Scholar] [CrossRef]
- Davey, M.S.; Tamassia, N.; Rossato, M.; Bazzoni, F.; Calzetti, F.; Bruderek, K.; Sironi, M.; Zimmer, L.; Bottazzi, B.; Mantovani, A.; et al. Failure to detect production of il-10 by activated human neutrophils. Nat. Immunol. 2011, 12, 1017–1018. [Google Scholar] [CrossRef]
- Minshawi, F.; Lanvermann, S.; McKenzie, E.; Jeffery, R.; Couper, K.; Papoutsopoulou, S.; Roers, A.; Muller, W. The generation of an engineered interleukin-10 protein with improved stability and biological function. Front. Immunol. 2020, 11, 1794. [Google Scholar] [CrossRef]
- Sharifinejad, N.; Zaki-Dizaji, M.; Sepahvandi, R.; Fayyaz, F.; Dos Santos Vilela, M.M.; El Ghazali, G.; Abolhassani, H.; Ochs, H.D.; Azizi, G. The clinical, molecular, and therapeutic features of patients with Il10/Il10r deficiency: A systematic review. Clin. Exp. Immunol. 2022, 208, 281–291. [Google Scholar] [CrossRef]
- Zhu, L.; Shi, T.; Zhong, C.; Wang, Y.; Chang, M.; Liu, X. Il-10 and il-10 receptor mutations in very early onset inflammatory bowel disease. Gastroenterol. Res. 2017, 10, 65–69. [Google Scholar] [CrossRef]
- Krawiec, P.; Pawlowska-Kamieniak, A.; Pac-Kozuchowska, E. Interleukin 10 and interleukin 10 receptor in paediatric inflammatory bowel disease: From bench to bedside lesson. J. Inflamm. 2021, 18, 13. [Google Scholar] [CrossRef]
- Lopiz, D.; Ruiz, M.; Infante, S.; Villanueva, L.; Silva, L.; Hervas-Stubbs, S.; Alignani, D.; Guruceaga, E.; Lasarte, J.J.; Sarobe, P. Il-10 expression defines an immunosuppressive dendritic cell population induced by antitumor therapeutic vaccination. Oncotarget 2017, 8, 2659–2671. [Google Scholar] [CrossRef]
- Shouval, D.S.; Ouahed, J.; Biswas, A.; Goettel, J.A.; Horwitz, B.H.; Klein, C.; Muise, A.M.; Snapper, S.B. Interleukin 10 receptor signaling: Master regulator of intestinal mucosal homeostasis in mice and humans. Adv. Immunol. 2014, 122, 177–210. [Google Scholar] [CrossRef] [PubMed]
- Carlini, V.; Noonan, D.M.; Abdalalem, E.; Goletti, D.; Sansone, C.; Calabrone, L.; Albini, A. The multifaceted nature of IL-10: Regulation, role in immunological homeostasis and its relevance to cancer, COVID-19 and post-COVID conditions. Front. Immunol. 2023, 14, 1161067. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Briukhovetska, D.; Dorr, J.; Endres, S.; Libby, P.; Dinarello, C.A.; Kobold, S. Interleukins in cancer: From biology to therapy. Nat. Rev. Cancer 2021, 21, 481–499. [Google Scholar] [CrossRef]
- Wilke, C.M.; Wei, S.; Wang, L.; Kryczek, I.; Kao, J.; Zou, W. Dual biological effects of the cytokines interleukin-10 and interferon-gamma. Cancer Immunol. Immunother. 2011, 60, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Penaloza, H.F.; Schultz, B.M.; Nieto, P.A.; Salazar, G.A.; Suazo, I.; Gonzalez, P.A.; Riedel, C.A.; Alvarez-Lobos, M.M.; Kalergis, A.M.; Bueno, S.M. Opposing roles of il-10 in acute bacterial infection. Cytokine Growth Factor Rev. 2016, 32, 17–30. [Google Scholar] [CrossRef]
- Cyktor, J.C.; Turner, J. Interleukin-10 and immunity against prokaryotic and eukaryotic intracellular pathogens. Infect. Immun. 2011, 79, 2964–2973. [Google Scholar] [CrossRef]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef]
- Verma, R.; Balakrishnan, L.; Sharma, K.; Khan, A.A.; Advani, J.; Gowda, H.; Tripathy, S.P.; Suar, M.; Pandey, A.; Gandotra, S.; et al. A network map of interleukin-10 signaling pathway. J. Cell Commun. Signal. 2016, 10, 61–67. [Google Scholar] [CrossRef]
- Schulke, S. Induction of interleukin-10 producing dendritic cells as a tool to suppress allergen-specific T helper 2 responses. Front. Immunol. 2018, 9, 455. [Google Scholar] [CrossRef]
- Fiorentino, D.F.; Bond, M.W.; Mosmann, T.R. Two types of mouse T helper cell. iv.Th2 clones secrete a factor that inhibits cytokine production by Th1 clones. J. Exp. Med. 1989, 170, 2081–2095. [Google Scholar] [CrossRef]
- Mittal, S.K.; Roche, P.A. Suppression of antigen presentation by il-10. Curr. Opin. Immunol. 2015, 34, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Mittal, S.K.; Cho, K.J.; Ishido, S.; Roche, P.A. Interleukin 10 (Il-10)-Mediated immunosuppression: March-I induction regulates antigen presentation by macrophages but not dendritic cells. J. Biol. Chem. 2015, 290, 27158–27167. [Google Scholar] [CrossRef] [PubMed]
- Bandola-Simon, J.; Roche, P.A. Dysfunction of antigen processing and presentation by dendritic cells in cancer. Mol. Immunol. 2019, 113, 31–37. [Google Scholar] [CrossRef]
- Wilson, E.B.; Brooks, D.G. The role of il-10 in regulating immunity to persistent viral infections. Curr. Top. Microbiol. Immunol. 2011, 350, 39–65. [Google Scholar] [CrossRef]
- Kumar, R.; Ng, S.; Engwerda, C. The role of il-10 in malaria: A double edged sword. Front. Immunol. 2019, 10, 229. [Google Scholar] [CrossRef]
- Goletti, D.; Weissman, D.; Jackson, R.W.; Collins, F.; Kinter, A.; Fauci, A.S. The in vitro induction of human immunodeficiency virus (Hiv) replication in purified protein derivative-positive hiv-infected persons by recall antigen response to mycobacterium tuberculosis is the result of a balance of the effects of endogenous interleukin-2 and proinflammatory and antiinflammatory cytokines. J. Infect. Dis. 1998, 177, 1332–1338. [Google Scholar] [CrossRef][Green Version]
- Taylor, A.; Akdis, M.; Joss, A.; Akkoc, T.; Wenig, R.; Colonna, M.; Daigle, I.; Flory, E.; Blaser, K.; Akdis, C.A. Il-10 inhibits Cd28 and icos costimulations of T cells Via src homology 2 domain-containing protein tyrosine phosphatase 1. J. Allergy Clin. Immunol. 2007, 120, 76–83. [Google Scholar] [CrossRef]
- Eiwegger, T.; Gruber, S.; Szepfalusi, Z.; Akdis, C.A. Novel developments in the mechanisms of immune tolerance to allergens. Hum. Vaccin. Immunother. 2012, 8, 1485–1491. [Google Scholar] [CrossRef][Green Version]
- Heine, G.; Drozdenko, G.; Grun, J.R.; Chang, H.D.; Radbruch, A.; Worm, M. Autocrine il-10 promotes human b-cell differentiation into igm- or igg-secreting plasmablasts. Eur. J. Immunol. 2014, 44, 1615–1621. [Google Scholar] [CrossRef]
- Laidlaw, B.J.; Lu, Y.; Amezquita, R.A.; Weinstein, J.S.; Vander Heiden, J.A.; Gupta, N.T.; Kleinstein, S.H.; Kaech, S.M.; Craft, J. Interleukin-10 from Cd4(+) follicular regulatory T cells promotes the germinal center response. Sci. Immunol. 2017, 2, eaan4767. [Google Scholar] [CrossRef]
- Rubtsov, Y.P.; Rasmussen, J.P.; Chi, E.Y.; Fontenot, J.; Castelli, L.; Ye, X.; Treuting, P.; Siewe, L.; Roers, A.; Henderson, W.R., Jr.; et al. Regulatory T cell-derived interleukin-10 limits inflammation at environmental interfaces. Immunity 2008, 28, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Santner-Nanan, B.; Hu, M.; Skarratt, K.; Lee, C.H.; Stormon, M.; Wong, M.; Fuller, S.J.; Nanan, R. Il-10 potentiates differentiation of human induced regulatory T cells Via Stat3 and Foxo1. J. Immunol. 2015, 195, 3665–3674. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Ma, R.; Yang, S.; Fan, T.; Cao, J.; Wang, Y.; Ma, W.; Yang, W.; Wang, F.; et al. Il-10 enhances T cell survival and is associated with faster relapse in patients with inactive ulcerative colitis. Mol. Immunol. 2020, 121, 92–98. [Google Scholar] [CrossRef]
- Yogev, N.; Bedke, T.; Kobayashi, Y.; Brockmann, L.; Lukas, D.; Regen, T.; Croxford, A.L.; Nikolav, A.; Hövelmeyer, N.; von Stebut, E.; et al. Cd4(+) T-Cell-Derived il-10 promotes cns inflammation in mice by sustaining effector T cell survival. Cell Rep. 2022, 38, 110565. [Google Scholar] [CrossRef] [PubMed]
- de Gruijter, N.M.; Jebson, B.; Rosser, E.C. Cytokine production by human b cells: Role in health and autoimmune disease. Clin. Exp. Immunol. 2022, 210, 253–262. [Google Scholar] [CrossRef]
- Oft, M. Immune regulation and cytotoxic T cell activation of il-10 agonists—Preclinical and clinical experience. Semin. Immunol. 2019, 44, 101325. [Google Scholar] [CrossRef]
- Mocellin, S.; Panelli, M.; Wang, E.; Rossi, C.R.; Pilati, P.; Nitti, D.; Lise, M.; Marincola, F.M. Il-10 stimulatory effects on human nk cells explored by gene profile analysis. Genes. Immun. 2004, 5, 621–630. [Google Scholar] [CrossRef]
- Park, J.Y.; Lee, S.H.; Yoon, S.R.; Park, Y.J.; Jung, H.; Kim, T.D.; Choi, I. Il-15-Induced il-10 increases the cytolytic activity of human natural killer cells. Mol. Cells 2011, 32, 265–272. [Google Scholar] [CrossRef]
- Mannino, M.H.; Zhu, Z.; Xiao, H.; Bai, Q.; Wakefield, M.R.; Fang, Y. The paradoxical role of il-10 in immunity and cancer. Cancer Lett. 2015, 367, 103–107. [Google Scholar] [CrossRef]
- Wang, Z.; Guan, D.; Huo, J.; Biswas, S.K.; Huang, Y.; Yang, Y.; Xu, S.; Lam, K.P. Il-10 enhances human natural killer cell effector functions Via metabolic reprogramming regulated by Mtorc1 signaling. Front. Immunol. 2021, 12, 619195. [Google Scholar] [CrossRef]
- Polukort, S.H.; Rovatti, J.; Carlson, L.; Thompson, C.; Ser-Dolansky, J.; Kinney, S.R.; Schneider, S.S.; Mathias, C.B. Il-10 enhances ige-mediated mast cell responses and is essential for the development of experimental food allergy in il-10-Deficient mice. J. Immunol. 2016, 196, 4865–4876. [Google Scholar] [CrossRef] [PubMed]
- Porro, C.; Cianciulli, A.; Panaro, M.A. The regulatory role of il-10 in neurodegenerative diseases. Biomolecules 2020, 10, 1017. [Google Scholar] [CrossRef] [PubMed]
- Short, W.D.; Steen, E.; Kaul, A.; Wang, X.; Olutoye, O.O., 2nd; Vangapandu, H.V.; Templeman, N.; Blum, A.J.; Moles, C.M.; Narmoneva, D.A.; et al. Il-10 promotes endothelial progenitor cell infiltration and wound healing Via Stat3. FASEB J. 2022, 36, e22298. [Google Scholar] [CrossRef]
- Pennica, D.; Nedwin, G.E.; Hayflick, J.S.; Seeburg, P.H.; Derynck, R.; Palladino, M.A.; Kohr, W.J.; Aggarwal, B.B.; Goeddel, D.V. Human tumour necrosis factor: Precursor structure, expression and homology to lymphotoxin. Nature 1984, 312 (Suppl. 5996), 724–729. [Google Scholar]
- Williams, T.W.; Granger, G.A. Lymphocyte in vitro cytotoxicity: Lymphotoxins of several mammalian species. Nature 1968, 219 (Suppl. 5158), 1076–1077. [Google Scholar]
- Aggarwal, B.B.; Moffat, B.; Harkins, R.N. Human lymphotoxin. Production by a lymphoblastoid cell line, purifi cation, and initial characterization. J. Biol. Chem. 1984, 259 (Suppl. 1), 686–691. [Google Scholar]
- Aggarwal, B.B.; Henzel, W.J.; Moffat, B.; Kohr, W.J.; Harkins, R.N. Primary structure of human lymphotoxin derived from 1788 lymphoblastoid cell line. J. Biol. Chem. 1985, 260 (Suppl. 4), 2334–2344. [Google Scholar]
- Ware, C.F. Network communications: Lymphotoxins, LIGHT, and, TNF. Annu. Rev. Immunol. 2005, 23, 787–819. [Google Scholar]
- Lindhout, I.A.; Murray, T.E.; Richards, C.M.; Klegeris, A. Potential neurotoxic activity of diverse molecules released by microglia. Neurochem. Int. 2021, 148, 105117. [Google Scholar] [CrossRef]
- Das, T.; Hoarau, J.J.; Jaffar Bandjee, M.C.; Maquart, M.; Gasque, P. Multifaceted innate immune responses engaged by astrocytes, microglia and resident dendritic cells against Chikungunya neuroinfection. J. Gen. Virol. 2015, 96, 294–310. [Google Scholar] [CrossRef]
- Medvedev, A.E.; Espevik, T.; Ranges, G.; Sundan, A. Distinct roles of the two tumor necrosis factor (TNF) receptors in modulating TNF and lymphotoxin alpha eff ects. J. Biol. Chem. 1996, 271 (Suppl. 16), 9778–9784. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.L.; Dougas, I.; Ngam-ek, A.; Bourdon, P.R.; Ehrenfels, B.N.; Miatkowski, K.; Zafari, M.; Yampaglia, A.M.; Lawton, P.; Meier, W.; et al. Characterization of surface lymphotoxin forms. Use of specific monoclonal antibodies and soluble receptors. J. Immunol. 1995, 154 (Suppl. 1), 33–46. [Google Scholar] [CrossRef]
- Fu, Y.X.; Chaplin, D.D. Development and maturation of secondary lymphoid tissues. Annu. Rev. Immunol. 1999, 17, 399–433. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, K.; Chaplin, D.D.; Wang, Y.; Wu, Q.; Pegg, L.E.; Yokoyama, W.M.; Fu, Y.X. Requirement for membrane lymphotoxin in natural killer cell development. Proc. Natl. Acad. Sci. USA 1999, 96 (Suppl. 11), 6336–6340. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, Y.; Wang, J.; Lin, X.; Wang, Y.; Pegg, L.E.; Futterer, A.; Pfeffer, K.; Fu, Y.X. Signal via lymphotoxin-beta R on bone marrow stromal cells is required for an early checkpoint of NK cell development. J. Immunol. 2001, 166 (Suppl. 3), 1684–1689. [Google Scholar] [CrossRef]
- Smyth, M.J.; Johnstone, R.W.; Cretney, E.; Haynes, N.M.; Sedgwick, J.D.; Korner, H.; Poulton, L.D.; Baxter, A.G. Multiple defi ciencies underlie NK cell inactivity in lymphotoxin-alpha gene-targeted mice. J. Immunol. 1999, 163 (Suppl. 3), 1350–1353. [Google Scholar] [CrossRef]
- James Bates, R.E.; Browne, E.; Schalks, R.; Jacobs, H.; Tan, L.; Parekh, P.; Magliozzi, R.; Calabrese, M.; Mazarakis, N.D.; Reynolds, R. Lymphotoxin-alpha expression in the meninges causes lymphoid tissue formation and neurodegeneration. Brain 2022, 145, 4287–4307. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Powell, M.B.; Mitchell, D.; Lederman, J.; Buckmeier, J.; Zamvil, S.S.; Graham, M.; Ruddle, N.H.; Steinman, L. Lymphotoxin and tumor necrosis factor-alpha production by myelin basic protein-specifi c T cell clones correlates with encephalitogenicity. Int. Immunol. 1990, 2 (Suppl. 6), 539–544. [Google Scholar] [CrossRef]
- Hultgren, O.; Eugster, H.P.; Sedgwick, J.D.; Korner, H.; Tarkowski, A. TNF/lymphotoxin-alpha double-mutant mice resist septic arthritis but display increased mortality in response to Staphylococcus aureus. J. Immunol. 1998, 161 (Suppl. 11), 5937–5942. [Google Scholar] [CrossRef]
- Roach, D.R.; Briscoe, H.; Saunders, B.; France, M.P.; Riminton, S.; Britton, W.J. Secreted lymphotoxin-alpha is essential for the control of an intracellular bacterial infection. J. Exp. Med. 2001, 193 (Suppl. 2), 239–246. [Google Scholar] [CrossRef]
- Ehlers, S.; Holscher, C.; Scheu, S.; Tertilt, C.; Hehlgans, T.; Suwinski, J.; Endres, R.; Pfeffer, K. The lymphotoxin beta receptor is critically involved in controlling infections with the intracellular pathogens Mycobacterium tuberculosis and Listeria monocytogenes. J. Immunol. 2003, 170 (Suppl. 10), 5210–5218. [Google Scholar] [CrossRef] [PubMed]
- Engwerda, C.R.; Mynott, T.L.; Sawhney, S.; De Souza, J.B.; Bickle, Q.D.; Kaye, P.M. Locally up-regulated lymphotoxin alpha, not systemic tumor necrosis factor alpha, is the principle mediator of murine cerebral malaria. J. Exp. Med. 2002, 195 (Suppl. 10), 1371–1377. [Google Scholar] [PubMed]
- Engwerda, C.R.; Ato, M.; Stager, S.; Alexander, C.E.; Stanley, A.C.; Kaye, P.M. Distinct roles for lymphotoxin-alpha and tumor necrosis factor in the control of Leishmania donovani infection. Am. J. Pathol. 2004, 165 (Suppl. 6), 2123–2133. [Google Scholar] [CrossRef]
- Sacca, R.; Cuff, C.A.; Lesslauer, W.; Ruddle, N.H. Differential activities of secreted lymphotoxin-alpha3 and membrane lymphotoxin-alpha1beta2 in lymphotoxin-induced infl ammation: Critical role of TNF receptor 1 signaling. J. Immunol. 1998, 160 (Suppl. 1), 485–491. [Google Scholar] [CrossRef]
- Pober, J.S.; Lapierre, L.A.; Stolpen, A.H.; Brock, T.A.; Springer, T.A.; Fiers, W.; Bevilacqua, M.P.; Mendrick, D.L.; Gimbrone, M.A., Jr. Activation of cultured human endothelial cells by recombinant lymphotoxin: Comparison with tumor necrosis factor and interleukin 1 species. J. Immunol. 1987, 138 (Suppl. 10), 3319–3324. [Google Scholar] [CrossRef]
- Kratz, A.; Campos-Neto, A.; Hanson, M.S.; Ruddle, N.H. Chronic inflammation caused by lymphotoxin is lymphoid neogenesis. J. Exp. Med. 1996, 183 (Suppl. 4), 1461–1472. [Google Scholar] [CrossRef]
- Cuff, C.A.; Schwartz, J.; Bergman, C.M.; Russell, K.S.; Bender, J.R.; Ruddle, N.H. Lymphotoxin alpha3 induces chemokines and adhesion molecules: Insight into the role of LT alpha in inflammation and lymphoid organ development. J. Immunol. 1998, 161 (Suppl. 12), 6853–6860. [Google Scholar]
- Etemadi, N.; Holien, J.K.; Chau, D.; Dewson, G.; Murphy, J.M.; Alexander, W.S.; Parker, M.W.; Silke, J.; Nachbur, U. Lymphotoxin α induces apoptosis, necroptosis and inflammatory signals with the same potency as tumour necrosis factor. FEBS J. 2013, 280, 5283–5297. [Google Scholar] [CrossRef] [PubMed]
- Mounzer, R.H.; Svendsen, O.S.; Baluk, P.; Bergman, C.M.; Padera, T.P.; Wiig, H.; Jain, R.K.; McDonald, D.M.; Ruddle, N.H. Lymphotoxin-alpha contributes to lymphangiogenesis. Blood 2010, 116 (Suppl. 12), 2173–2182. [Google Scholar] [CrossRef]
- Kakizawa, S. Subchapter 41C—Neurotrophin-3. In Handbook of Hormones, 2nd ed.; Ando, H., Ukena, K., Nagata, S., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 483–485. ISBN 9780128206492. [Google Scholar] [CrossRef]
- Wan, T.; Zhang, F.-S.; Qin, M.-Y.; Jiang, H.-R.; Zhang, M.; Qu, Y.; Wang, Y.-L.; Zhang, P.-X. Growth factors: Bioactive macromolecular drugs for peripheral nerve injury treatment—Molecular mechanisms and delivery platforms. Biomed. Pharmacother. 2024, 170, 116024. [Google Scholar] [CrossRef]
- Manoukian, O.S.; Rudraiah, S.; Arul, M.R.; Bartley, J.M.; Baker, J.T.; Yu, X.; Kumbar, S.G. Biopolymer-nanotube nerve guidance conduit drug delivery for peripheral nerve regeneration: In vivo structural and functional assessment. Bioact. Mater. 2021, 6, 2881–2893. [Google Scholar] [PubMed]
- Glaser, T.; Oliveira, Á.; Sardà-Arroyo, L.; Henning, U. Chapter 7—Growth and Neurotrophic Factor Receptors in Neural Differentiation and Phenotype Specification; Pruszak, J., Antigens, N.S., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 77–90. ISBN 9780128007815. [Google Scholar] [CrossRef]
- Li, G.; Zhang, B.; Sun, J.H.; Shi, L.Y.; Huang, M.Y.; Huang, L.J.; Lin, Z.J.; Lin, Q.Y.; Lai, B.Q.; Ma, Y.H.; et al. An NT-3-releasing bioscaffold supports the formation of -modified neural stem cell-derived neural network tissue with efficacy in repairing spinal cord injury. Bioact. Mater. 2021, 6, 3766–3781. [Google Scholar] [PubMed]
- Müller, M.L.; Peglau, L.; Moon, D.F.L.; Groß, S.; Schulze, J.; Ruhnau, J.; Vogelgesang, A. Neurotrophin-3 attenuates human peripheral blood T cell and monocyte activation status and cytokine production post stroke. Exp. Neurol. 2022, 347, 113901. [Google Scholar]
- Boato, F.; Hechler, D.; Rosenberger, K.; Lüdecke, D.; Peters, E.M.; Nitsch, R.; Hendrix, S. Interleukin-1 beta and neurotrophin-3 synergistically promote neurite growth in vitro. J. Neuroinflamm. 2011, 8, 183. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, S.C.; Dickson, D.W.; Brosnan, C.F. Interleukin-1, nitric oxide and reactive astrocytes. Brain Behav. Immun. 1995, 9, 345–354. [Google Scholar]
- Feder, L.S.; Laskin, D.L. Regulation of hepatic endothelial cell and macrophage proliferation and nitric oxide production by GM-CSF, MCSF, and IL-1 beta following acute endotoxemia. J. Leukoc. Biol. 1994, 55, 507–513. [Google Scholar]
- Giulian, D.; Lachman, L.B. Interleukin-1 stimulation of astroglial proliferation after brain injury. Science 1985, 228, 497–499. [Google Scholar]
- Giulian, D.; Woodward, J.; Young, D.G.; Krebs, J.F.; Lachman, L.B. Interleukin-1 injected into mammalian brain stimulates astrogliosis and neovascularization. J. Neurosci. 1988, 8, 2485–2490. [Google Scholar]
- Marschinke, F.; Stromberg, I. Dual effects of TNFalpha on nerve fiber formation from ventral mesencephalic organotypic tissue cultures. Brain Res. 2008, 1215, 30–39. [Google Scholar]
- Hakkoum, D.; Stoppini, L.; Muller, D. Interleukin-6 promotes sprouting and functional recovery in lesioned organotypic hippocampal slice cultures. J. Neurochem. 2007, 100, 747–757. [Google Scholar]
- Lindholm, D.; Heumann, R.; Meyer, M.; Thoenen, H. Interleukin-1 regulates synthesis of nerve growth factor in non-neuronal cells of rat sciatic nerve. Nature 1987, 330, 658–659. [Google Scholar] [PubMed]
- Friedman, W.J.; Larkfors, L.; Ayer-LeLievre, C.; Ebendal, T.; Olson, L.; Persson, H. Regulation of beta-nerve growth factor expression by inflammatory mediators in hippocampal cultures. J. Neurosci. Res. 1990, 27, 374–382. [Google Scholar] [PubMed]
- Gadient, R.A.; Cron, K.C.; Otten, U. Interleukin-1 beta and tumor necrosis factor-alpha synergistically stimulate nerve growth factor (NGF) release from cultured rat astrocytes. Neurosci. Lett. 1990, 117, 335–340. [Google Scholar] [PubMed]
- Quarta, S.; Baeumer, B.E.; Scherbakov, N.; Andratsch, M.; Rose-John, S.; Dechant, G.; Bandtlow, C.E.; Kress, M. Peripheral nerve regeneration and NGF-dependent neurite outgrowth of adult sensory neurons converge on STAT3 phosphorylation downstream of neuropoietic cytokine receptor gp130. J. Neurosci. 2014, 34, 13222–13233. [Google Scholar]
- Zafar, M.I.; Zheng, J.; Kong, W.; Ye, X.; Gou, L.; Regmi, A.; Chen, L.L. The role of vascular endothelial growth factor-B in metabolic homoeostasis: Current evidence. Biosci. Rep. 2017, 37, BSR20171089. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, X.; Lee, C.; Tang, Z.; Zhang, F.; Arjunan, P.; Li, Y.; Hou, X.; Kumar, A.; Dong, L. VEGF-B: A survival, or an angiogenic factor? Cell. Adh. Migr. 2009, 3, 322–327. [Google Scholar]
- Li, Y.; Zhang, F.; Nagai, N.; Tang, Z.; Zhang, S.; Scotney, P.; Lennartsson, J.; Zhu, C.; Qu, Y.; Fang, C.; et al. VEGF-B inhibits apoptosis via VEGFR-1-mediated suppression of the expression of BH3-only protein genes in mice and rats. J. Clin. Investig. 2008, 118, 913–923. [Google Scholar]
- Robciuc, M.R.; Kivelä, R.; Williams, I.M.; de Boer, J.F.; van Dijk, T.H.; Elamaa, H.; Tigistu-Sahle, F.; Molotkov, D.; Leppänen, V.M.; Käkelä, R.; et al. VEGFB/VEGFR1-induced expansion of adipose vasculature counteracts obesity and related metabolic complications. Cell Metab. 2016, 23, 712–724. [Google Scholar]
- Zhang, F.; Tang, Z.; Hou, X.; Lennartsson, J.; Li, Y.; Koch, A.W.; Scotney, P.; Lee, C.; Arjunan, P.; Dong, L.; et al. VEGF-B is dispensable for blood vessel growth but critical for their survival, and VEGF-B targeting inhibits pathological angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 6152–6157. [Google Scholar]
- Malik, A.K.; Baldwin, M.E.; Peale, F.; Fuh, G.; Liang, W.C.; Lowman, H.; Meng, G.; Ferrara, N.; Gerber, H.P. Redundant roles of VEGF-B and PlGF during selective VEGF-A blockade in mice. Blood 2006, 107, 550–557. [Google Scholar]
- Olofsson, B.; Korpelainen, E.; Pepper, M.S.; Mandriota, S.J.; Aase, K.; Kumar, V.; Gunji, Y.; Jeltsch, M.M.; Shibuya, M.; Alitalo, K.; et al. Vascular endothelial growth factor B (VEGF-B) binds to VEGF receptor-1 and regulates plasminogen activator activity in endothelial cells. Proc. Natl. Acad. Sci. USA 1998, 95, 11709–11714. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M. Vascular endothelial growth factor receptor-1 (VEGFR-1/Flt-1), a dual regulator for angiogenesis. Angiogenesis 2006, 9, 225–230. [Google Scholar] [PubMed]
- Bielenberg, D.R.; Pettaway, C.A.; Takashima, S.; Klagsbrun, M. Neuropilins in neoplasms: Expression, regulation and function. Exp. Cell Res. 2006, 312, 584–593. [Google Scholar]
- Sun, Y.; Jin, K.; Childs, J.T.; Xie, L.; Mao, X.O.; Greenberg, D.A. Increased severity of cerebral ischemic injury in vascular endothelial growth factor-B-deficient mice. J. Cereb. Blood Flow. Metab. 2004, 24, 1146–1152. [Google Scholar] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gheorghiu, M.; Trandafir, M.-F.; Savu, O.; Pasarica, D.; Bleotu, C. Unexpectedly High and Difficult-to-Explain Regenerative Capacity in an 82-Year-Old Patient with Insulin-Requiring Type 2 Diabetes and End-Stage Renal Disease. J. Clin. Med. 2025, 14, 2556. https://doi.org/10.3390/jcm14082556
Gheorghiu M, Trandafir M-F, Savu O, Pasarica D, Bleotu C. Unexpectedly High and Difficult-to-Explain Regenerative Capacity in an 82-Year-Old Patient with Insulin-Requiring Type 2 Diabetes and End-Stage Renal Disease. Journal of Clinical Medicine. 2025; 14(8):2556. https://doi.org/10.3390/jcm14082556
Chicago/Turabian StyleGheorghiu, Mihaela, Maria-Florina Trandafir, Octavian Savu, Daniela Pasarica, and Coralia Bleotu. 2025. "Unexpectedly High and Difficult-to-Explain Regenerative Capacity in an 82-Year-Old Patient with Insulin-Requiring Type 2 Diabetes and End-Stage Renal Disease" Journal of Clinical Medicine 14, no. 8: 2556. https://doi.org/10.3390/jcm14082556
APA StyleGheorghiu, M., Trandafir, M.-F., Savu, O., Pasarica, D., & Bleotu, C. (2025). Unexpectedly High and Difficult-to-Explain Regenerative Capacity in an 82-Year-Old Patient with Insulin-Requiring Type 2 Diabetes and End-Stage Renal Disease. Journal of Clinical Medicine, 14(8), 2556. https://doi.org/10.3390/jcm14082556