
The Respiratory Microbiome in Paediatric Chronic Wet Cough: What Is Known and Future Directions

 , , , ,
, , , , 
Abstract
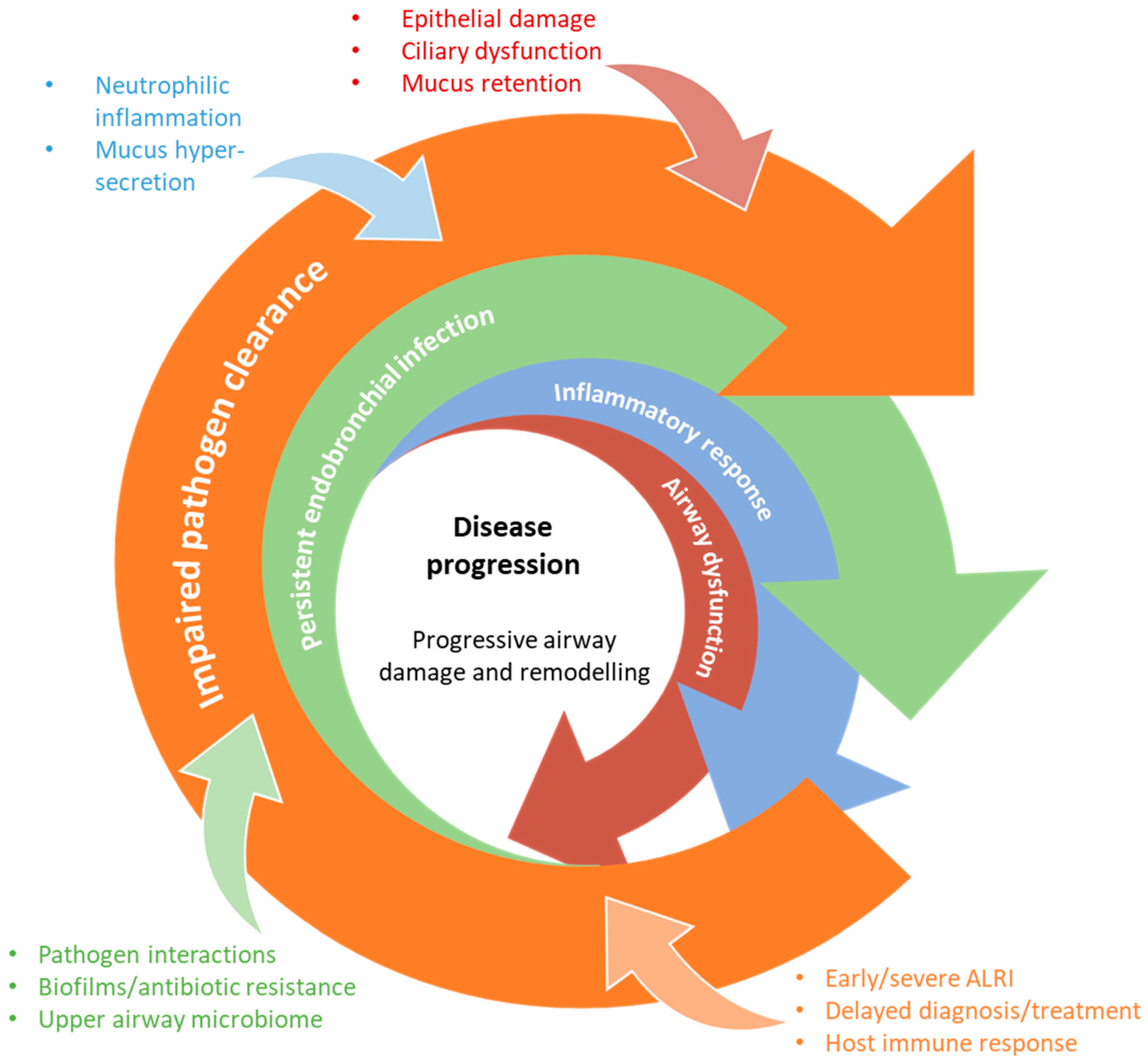
:1. Introduction
2. The Lower Airway Microbiome among Children with PBB or Bronchiectasis
2.1. Bacteriome
2.2. Virome
2.3. Mycobiome
3. Biofilm
4. Microbiome Dynamics
4.1. Interactions between Bacterial Pathobionts
4.2. Viral–Bacterial Interactions
4.3. The Gut–Lung Axis
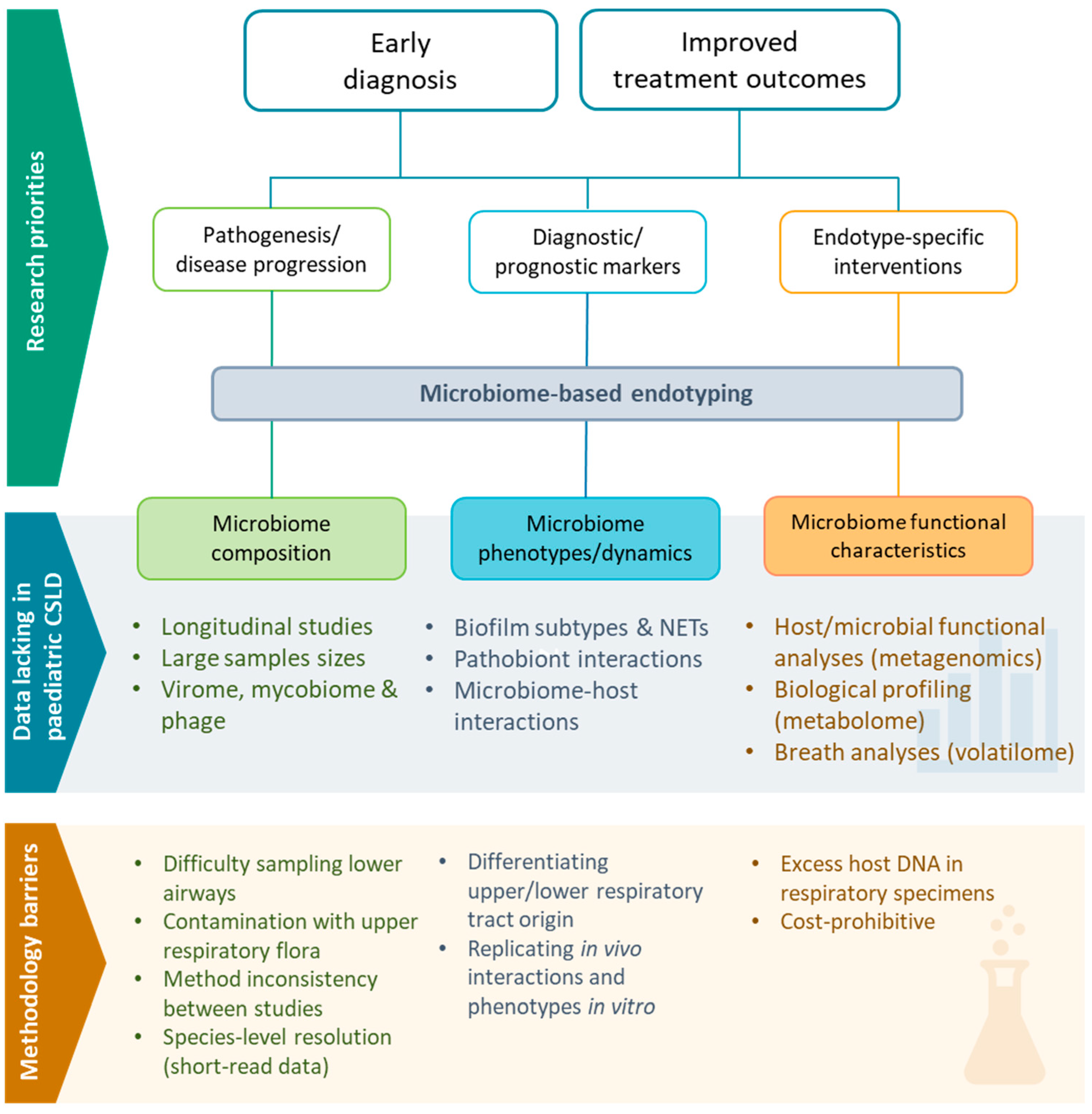
5. Moving beyond Amplicon Sequencing Data
5.1. Multiomics
5.2. Diagnostic Translation of ‘Omics Data
6. Emerging Therapeutic Approaches
6.1. Anti-Biofilm Therapies
6.2. Phage Therapy
6.3. Probiotics
7. Summary
8. Conclusions
Funding
Conflicts of Interest
References
- Bergmann, M.; Haasenritter, J.; Beidatsch, D.; Schwarm, S.; Hörner, K.; Bösner, S.; Grevenrath, P.; Schmidt, L.; Viniol, A.; Donner-Banzhoff, N. Coughing children in family practice and primary care: A systematic review of prevalence, aetiology and prognosis. BMC Pediatr. 2021, 21, 260. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.B.; Bell, S.C.; Torzillo, P.J.; King, P.T.; Maguire, G.P.; Byrnes, C.A.; Holland, A.E.; O’Mara, P.; Grimwood, K. Chronic suppurative lung disease and bronchiectasis in children and adults in Australia and New Zealand Thoracic Society of Australia and New Zealand guidelines. Med. J. Aust. 2015, 202, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Kantar, A.; Chang, A.B.; Shields, M.D.; Marchant, J.M.; Grimwood, K.; Grigg, J.; Priftis, K.N.; Cutrera, R.; Midulla, F.; Brand, P.L. ERS statement on protracted bacterial bronchitis in children. Eur. Respir. J. 2017, 50, 1602139. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, K.F.; Drescher, B.J.; Goyal, V.; Phillips, N.; Acworth, J.; Marchant, J.M.; Chang, A.B. Chronic cough postacute respiratory illness in children: A cohort study. Arch. Dis. Child. 2017, 102, 1044–1048. [Google Scholar] [CrossRef]
- O’Grady, K.-A.F.; Mahon, J.; Arnold, D.; Grimwood, K.; Hall, K.K.; Goyal, V.; Marchant, J.M.; Phillips, N.; Acworth, J.; King, A. Predictors of the development of protracted bacterial bronchitis following presentation to healthcare for an acute respiratory illness with cough: Analysis of three cohort studies. J. Clin. Med. 2021, 10, 5735. [Google Scholar] [CrossRef]
- Wurzel, D.F.; Marchant, J.M.; Yerkovich, S.T.; Upham, J.W.; Petsky, H.L.; Smith-Vaughan, H.; Masters, B.; Buntain, H.; Chang, A.B. Protracted Bacterial Bronchitis in Children: Natural History and Risk Factors for Bronchiectasis. Chest 2016, 150, 1101–1108. [Google Scholar] [CrossRef]
- Ruffles, T.J.C.; Marchant, J.M.; Masters, I.B.; Yerkovich, S.T.; Wurzel, D.F.; Gibson, P.G.; Busch, G.; Baines, K.J.; Simpson, J.L.; Smith-Vaughan, H.C.; et al. Outcomes of protracted bacterial bronchitis in children: A 5-year prospective cohort study. Respirology 2021, 26, 241–248. [Google Scholar] [CrossRef]
- Prime, S.J.; Carter, H.E.; McPhail, S.M.; Petsky, H.L.; Chang, A.B.; Graves, N.; Marchant, J.M. Chronic wet cough in Australian children: Societal costs and quality of life. Pediatr. Pulmonol. 2021, 56, 2707–2716. [Google Scholar] [CrossRef]
- Roberts, J.M.; Goyal, V.; Kularatna, S.; Chang, A.B.; Kapur, N.; Chalmers, J.D.; Goeminne, P.C.; Hernandez, F.; Marchant, J.M.; McPhail, S.M. The Economic Burden of Bronchiectasis: A Systematic Review. Chest 2023, 164, 1396–1421. [Google Scholar] [CrossRef]
- King, P.T.; Holdsworth, S.R.; Farmer, M.; Freezer, N.; Villanueva, E.; Holmes, P.W. Phenotypes of adult bronchiectasis: Onset of productive cough in childhood and adulthood. COPD J. Chronic Obstr. Pulm. Dis. 2009, 6, 130–136. [Google Scholar] [CrossRef]
- Di Filippo, P.; Scaparrotta, A.; Petrosino, M.I.; Attanasi, M.; Di Pillo, S.; Chiarelli, F.; Mohn, A. An underestimated cause of chronic cough: The protracted bacterial bronchitis. Ann. Thorac. Med. 2018, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.B.; Bell, S.C.; Byrnes, C.A.; Dawkins, P.; Holland, A.E.; Kennedy, E.; King, P.T.; Laird, P.; Mooney, S.; Morgan, L.; et al. Thoracic Society of Australia and New Zealand (TSANZ) position statement on chronic suppurative lung disease and bronchiectasis in children, adolescents and adults in Australia and New Zealand. Respirology 2023, 28, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Gallucci, M.; Pedretti, M.; Giannetti, A.; Di Palmo, E.; Bertelli, L.; Pession, A.; Ricci, G. When the cough does not improve: A review on protracted bacterial bronchitis in children. Front. Pediatr. 2020, 8, 433. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.B.; Yerkovich, S.T.; Gibson, P.G.; Anderson-James, S.; Petsky, H.L.; Carroll, M.L.; Masters, I.B.; Marchant, J.M.; Wurzel, D.; Upham, J.W. Pulmonary innate immunity in children with protracted bacterial bronchitis. J. Pediatr. 2012, 161, 621–625.e621. [Google Scholar] [CrossRef] [PubMed]
- Marchant, J.M.; Gibson, P.G.; Grissell, T.V.; Timmins, N.L.; Masters, I.B.; Chang, A.B. Prospective assessment of protracted bacterial bronchitis: Airway inflammation and innate immune activation. Pediatr. Pulmonol. 2008, 43, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Vandivier, R.W.; Henson, P.M.; Douglas, I.S. Burying the dead: The impact of failed apoptotic cell removal (efferocytosis) on chronic inflammatory lung disease. Chest 2006, 129, 1673–1682. [Google Scholar] [CrossRef]
- Kartsiouni, E.; Chatzipanagiotou, S.; Galani, A.; Moriki, D.; Sardeli, O.; Prountzos, S.; Alexopoulou, E.; Loukou, I.; Priftis, K.N.; Douros, K. Clinical course of children with chronic suppurative lung disease or bronchiectasis infected with Pseudomonas aeruginosa. Children 2022, 9, 1822. [Google Scholar] [CrossRef]
- Cole, P. Inflammation: A two-edged sword—The model of bronchiectasis. Eur. J. Respir. Diseases. Suppl. 1986, 147, 6–15. [Google Scholar]
- Pizzutto, S.J.; Hare, K.M.; Upham, J.W. Bronchiectasis in children: Current concepts in immunology and microbiology. Front. Pediatr. 2017, 5, 123. [Google Scholar] [CrossRef]
- Keir, H.R.; Chalmers, J.D. Pathophysiology of Bronchiectasis. Semin. Respir. Crit. Care Med. 2021, 42, 499–512. [Google Scholar] [CrossRef]
- Flume, P.A.; Chalmers, J.D.; Olivier, K.N. Advances in bronchiectasis: Endotyping, genetics, microbiome, and disease heterogeneity. Lancet 2018, 392, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Keir, H.R.; Chalmers, J.D. Endotypes in bronchiectasis: Moving towards precision medicine. A narrative review. Pulmonology 2023, 29, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mac Aogain, M.; Xu, T.; Jaggi, T.K.; Chan, L.L.Y.; Qu, J.; Wei, L.; Liao, S.; Cheng, H.S.; Keir, H.R.; et al. Neisseria species as pathobionts in bronchiectasis. Cell Host Microbe 2022, 30, 1311–1327.e1318. [Google Scholar] [CrossRef] [PubMed]
- Kompare, M.; Weinberger, M. Protracted bacterial bronchitis in young children: Association with airway malacia. J. Pediatr. 2012, 160, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Hare, K.M.; Pizzutto, S.J.; Chang, A.B.; Smith-Vaughan, H.C.; McCallum, G.B.; Beissbarth, J.; Versteegh, L.; Grimwood, K. Defining lower airway bacterial infection in children with chronic endobronchial disorders. Pediatr. Pulmonol. 2018, 53, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Craven, V.; Everard, M.L. Protracted bacterial bronchitis: Reinventing an old disease. Arch. Dis. Child. 2013, 98, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Broderick, D.T.J.; Waite, D.W.; Marsh, R.L.; Camargo, C.A., Jr.; Cardenas, P.; Chang, A.B.; Cookson, W.O.C.; Cuthbertson, L.; Dai, W.; Everard, M.L.; et al. Bacterial Signatures of Paediatric Respiratory Disease: An Individual Participant Data Meta-Analysis. Front. Microbiol. 2021, 12, 711134. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, L.; Craven, V.; Bingle, L.; Cookson, W.O.; Everard, M.L.; Moffatt, M.F. The impact of persistent bacterial bronchitis on the pulmonary microbiome of children. PLoS ONE 2017, 12, e0190075. [Google Scholar] [CrossRef]
- Marsh, R.L.; Smith-Vaughan, H.C.; Chen, A.C.; Marchant, J.M.; Yerkovich, S.T.; Gibson, P.G.; Pizzutto, S.J.; Hodge, S.; Upham, J.W.; Chang, A.B. Multiple respiratory microbiota profiles are associated with lower airway inflammation in children with protracted bacterial bronchitis. Chest 2019, 155, 778–786. [Google Scholar] [CrossRef]
- Marsh, R.L.; Kaestli, M.; Chang, A.B.; Binks, M.J.; Pope, C.E.; Hoffman, L.R.; Smith-Vaughan, H.C. The microbiota in bronchoalveolar lavage from young children with chronic lung disease includes taxa present in both the oropharynx and nasopharynx. Microbiome 2016, 4, 37. [Google Scholar] [CrossRef]
- Pillarisetti, N.; Broderick, D.; Ainsworth, A.; Mulholland, A.; Wagner Mackenzie, B.; Middleton, D.; Byrnes, C.A.; Taylor, M.W. The airway microbiota in children newly diagnosed with bronchiectasis largely retains its diversity. Eur. Respir. J. 2019, 54, 1900704. [Google Scholar] [CrossRef] [PubMed]
- van der Gast, C.J.; Cuthbertson, L.; Rogers, G.B.; Pope, C.; Marsh, R.L.; Redding, G.J.; Bruce, K.D.; Chang, A.B.; Hoffman, L.R. Three clinically distinct chronic pediatric airway infections share a common core microbiota. Ann. Am. Thorac. Soc. 2014, 11, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Broderick, D.T.J.; Regtien, T.; Ainsworth, A.; Taylor, M.W.; Pillarisetti, N. Dynamic Upper and Lower Airway Microbiotas in Paediatric Bronchiectasis Exacerbations: A Pilot Study. Front. Cell. Infect. Microbiol. 2021, 11, 773496. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Ryu, S.; Keir, H.R.; Giam, Y.H.; Dicker, A.J.; Perea, L.; Richardson, H.; Huang, J.T.J.; Cant, E.; Blasi, F.; et al. Inflammatory Molecular Endotypes in Bronchiectasis: A European Multicenter Cohort Study. Am. J. Respir. Crit. Care Med. 2023, 208, 1147–1148. [Google Scholar] [CrossRef] [PubMed]
- Charlson, E.S.; Bittinger, K.; Haas, A.R.; Fitzgerald, A.S.; Frank, I.; Yadav, A.; Bushman, F.D.; Collman, R.G. Topographical continuity of bacterial populations in the healthy human respiratory tract. Am. J. Respir. Crit. Care Med. 2011, 184, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Marsh, R.L.; Binks, M.J.; Smith-Vaughan, H.C.; Janka, M.; Clark, S.; Richmond, P.; Chang, A.B.; Thornton, R.B. Prevalence and subtyping of biofilms present in bronchoalveolar lavage from children with protracted bacterial bronchitis or non-cystic fibrosis bronchiectasis: A cross-sectional study. Lancet Microbe 2022, 3, e215–e223. [Google Scholar] [CrossRef] [PubMed]
- Richardson, H.; Dicker, A.J.; Barclay, H.; Chalmers, J.D. The microbiome in bronchiectasis. Eur. Respir. Rev. 2019, 28, 190048. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.T.; Pope, C.E.; Marsh, R.L.; Wolter, D.J.; Weiss, E.J.; Hager, K.R.; Vo, A.T.; Brittnacher, M.J.; Radey, M.C.; Hayden, H.S.; et al. Human and Extracellular DNA Depletion for Metagenomic Analysis of Complex Clinical Infection Samples Yields Optimized Viable Microbiome Profiles. Cell Rep. 2019, 26, 2227–2240.e2225. [Google Scholar] [CrossRef]
- Ben-Shimol, S.; Ramilo, O.; Leber, A.L.; van der Beek, B.A.; Everhart, K.; Mertz, S.; Mejias, A.; Dagan, R. A Hypothesis-Generating Prospective Longitudinal Study to Assess the Relative Contribution of Common Respiratory Viruses to Severe Lower Respiratory Infections in Young Children. Pediatr. Infect. Dis. J. 2023, 42, 396–404. [Google Scholar] [CrossRef]
- Kapur, N.; Mackay, I.M.; Sloots, T.P.; Masters, I.B.; Chang, A.B. Respiratory viruses in exacerbations of non-cystic fibrosis bronchiectasis in children. Arch. Dis. Child. 2014, 99, 749–753. [Google Scholar] [CrossRef]
- McCallum, G.B.; Oguoma, V.M.; Versteegh, L.A.; Wilson, C.A.; Bauert, P.; Spain, B.; Chang, A.B. Comparison of profiles of first nations and non-first nations children with bronchiectasis over two 5-year periods in the Northern Territory, Australia. Chest 2021, 160, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Gonzalez, R.; Xie, Z.; Zhang, J.; Liu, C.; Li, J.; Li, Y.; Wang, Z.; Kong, X.; Yao, Y. WU and KI polyomavirus present in the respiratory tract of children, but not in immunocompetent adults. J. Clin. Virol. 2008, 43, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Sarna, M.; Lambert, S.B.; Sloots, T.P.; Whiley, D.M.; Alsaleh, A.; Mhango, L.; Bialasiewicz, S.; Wang, D.; Nissen, M.D.; Grimwood, K. Viruses causing lower respiratory symptoms in young children: Findings from the ORChID birth cohort. Thorax 2018, 73, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, Q.; Dai, W.; Feng, X.; Lu, Z.; Yang, Z.; Liu, Y.; Xie, G.; Yang, Y.; Shen, K. Lung microbiota and pulmonary inflammatory cytokines expression vary in children with tracheomalacia and adenoviral or mycoplasma pneumoniae pneumonia. Front. Pediatr. 2019, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hao, C.; Ji, W.; Lu, Y.; Wu, M.; Chen, S.; Wang, K.; Shao, X. Detecting respiratory viruses in children with protracted bacterial bronchitis. Respir. Med. 2019, 151, 55–58. [Google Scholar] [CrossRef]
- Wurzel, D.F.; Mackay, I.M.; Marchant, J.M.; Wang, C.Y.; Yerkovich, S.T.; Upham, J.W.; Smith-Vaughan, H.C.; Petsky, H.L.; Chang, A.B. Adenovirus species C is associated with chronic suppurative lung diseases in children. Clin. Infect. Dis. 2014, 59, 34–40. [Google Scholar] [CrossRef] [PubMed]
- McCallum, G.B.; Chatfield, M.D.; Morris, P.S.; Chang, A.B. Risk factors for adverse outcomes of Indigenous infants hospitalized with bronchiolitis. Pediatr. Pulmonol. 2016, 51, 613–623. [Google Scholar] [CrossRef]
- Garnett, C.; Erdman, D.; Xu, W.; Gooding, L.R. Prevalence and quantitation of species C adenovirus DNA in human mucosal lymphocytes. J. Virol. 2002, 76, 10608–10616. [Google Scholar] [CrossRef]
- Wurzel, D.F.; Marchant, J.M.; Yerkovich, S.T.; Upham, J.W.; Mackay, I.M.; Masters, I.B.; Chang, A.B. Prospective characterization of protracted bacterial bronchitis in children. Chest 2014, 145, 1271–1278. [Google Scholar] [CrossRef]
- Becroft, D. Bronchiolitis obliterans, bronchiectasis, and other sequelae of adenovirus type 21 infection in young children. J. Clin. Pathol. 1971, 24, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.A.; Taylor, L.J.; Dothard, M.I.; Leiby, J.S.; Fitzgerald, A.S.; Khatib, L.A.; Collman, R.G.; Bushman, F.D. Redondoviridae, a family of small, circular DNA viruses of the human oro-respiratory tract associated with periodontitis and critical illness. Cell Host Microbe 2019, 25, 719–729.e4. [Google Scholar] [CrossRef] [PubMed]
- Dodi, G.; Attanasi, M.; Di Filippo, P.; Di Pillo, S.; Chiarelli, F. Virome in the lungs: The role of anelloviruses in childhood respiratory diseases. Microorganisms 2021, 9, 1357. [Google Scholar] [CrossRef] [PubMed]
- Maggi, F.; Pifferi, M.; Tempestini, E.; Lanini, L.; De Marco, E.; Fornai, C.; Andreoli, E.; Presciuttini, S.; Vatteroni, M.L.; Pistello, M. Correlation between Torque tenovirus infection and serum levels of eosinophil cationic protein in children hospitalized for acute respiratory diseases. J. Infect. Dis. 2004, 190, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Bernardinello, N.; Cocconcelli, E.; Giraudo, C.; Daverio, M.; Castelli, G.; Petrarulo, S.; Bovo, M.; Fichera, G.; Cavinato, S.; Cattelan, A.M. Predictors of pulmonary sequelae after COVID-19 pneumonia: A 12-month follow-up study. Front. Med. 2023, 10, 1084002. [Google Scholar] [CrossRef]
- Ribeiro Carvalho, C.R.; Lamas, C.A.; Chate, R.C.; Salge, J.M.; Sawamura, M.V.Y.; de Albuquerque, A.L.; Toufen Junior, C.; Lima, D.M.; Garcia, M.L.; Scudeller, P.G. Long-term respiratory follow-up of ICU hospitalized COVID-19 patients: Prospective cohort study. PLoS ONE 2023, 18, e0280567. [Google Scholar] [CrossRef]
- Dadali, Y.; Özkaçmaz, S.; Ünlü, E.; Akyol, R.; Alparslan, M. Computed tomography findings of COVID-19 in pediatric patients. Turk. J. Pediatr. 2022, 64. [Google Scholar] [CrossRef] [PubMed]
- Einsiedel, L.; Fernandes, L.; Spelman, T.; Steinfort, D.; Gotuzzo, E. Bronchiectasis is associated with human T-lymphotropic virus 1 infection in an Indigenous Australian population. Clin. Infect. Dis. 2012, 54, 43–50. [Google Scholar] [CrossRef]
- Tzani-Tzanopoulou, P.; Skliros, D.; Megremis, S.; Xepapadaki, P.; Andreakos, E.; Chanishvili, N.; Flemetakis, E.; Kaltsas, G.; Taka, S.; Lebessi, E. Interactions of bacteriophages and bacteria at the airway mucosa: New insights into the pathophysiology of asthma. Front. Allergy 2021, 1, 617240. [Google Scholar] [CrossRef]
- Kapur, N.; Grimwood, K.; Masters, I.B.; Morris, P.S.; Chang, A.B. Lower airway microbiology and cellularity in children with newly diagnosed non-CF bronchiectasis. Pediatr. Pulmonol. 2012, 47, 300–307. [Google Scholar] [CrossRef]
- Thornton, R.B.; Wiertsema, S.P.; Kirkham, L.-A.S.; Rigby, P.J.; Vijayasekaran, S.; Coates, H.L.; Richmond, P.C. Neutrophil extracellular traps and bacterial biofilms in middle ear effusion of children with recurrent acute otitis media—A potential treatment target. PLoS ONE 2013, 8, e53837. [Google Scholar] [CrossRef]
- Silva, N.; Marques, L.; Röder, D. Diagnosis of biofilm infections: Current methods used, challenges and perspectives for the future. J. Appl. Microbiol. 2021, 131, 2148–2160. [Google Scholar] [CrossRef]
- Chalmers, J.D.; Hill, A.T. Mechanisms of immune dysfunction and bacterial persistence in non-cystic fibrosis bronchiectasis. Mol. Immunol. 2013, 55, 27–34. [Google Scholar] [CrossRef]
- Kahn, F.W.; Jones, J.M. Diagnosing bacterial respiratory infection by bronchoalveolar lavage. J. Infect. Dis. 1987, 155, 862–869. [Google Scholar] [CrossRef]
- Pizzichini, E.; Pizzichini, M.M.; Efthimiadis, A.; Hargreave, F.E.; Dolovich, J. Measurement of inflammatory indices in induced sputum: Effects of selection of sputum to minimize salivary contamination. Eur. Respir. J. 1996, 9, 1174–1180. [Google Scholar] [CrossRef]
- Marsh, R.L.; Thornton, R.B.; Smith-Vaughan, H.C.; Richmond, P.; Pizzutto, S.J.; Chang, A.B. Detection of biofilm in bronchoalveolar lavage from children with non-cystic fibrosis bronchiectasis. Pediatr. Pulmonol. 2015, 50, 284–292. [Google Scholar] [CrossRef]
- Welp, A.L.; Bomberger, J.M. Bacterial community interactions during chronic respiratory disease. Front. Cell. Infect. Microbiol. 2020, 10, 213. [Google Scholar] [CrossRef]
- Hare, K.M.; Grimwood, K.; Leach, A.J.; Smith-Vaughan, H.; Torzillo, P.J.; Morris, P.S.; Chang, A.B. Respiratory bacterial pathogens in the nasopharynx and lower airways of Australian indigenous children with bronchiectasis. J. Pediatr. 2010, 157, 1001–1005. [Google Scholar] [CrossRef]
- Weimer, K.E.; Juneau, R.A.; Murrah, K.A.; Pang, B.; Armbruster, C.E.; Richardson, S.H.; Swords, W.E. Divergent mechanisms for passive pneumococcal resistance to β-lactam antibiotics in the presence of Haemophilus influenzae. J. Infect. Dis. 2011, 203, 549–555. [Google Scholar] [CrossRef]
- Schaar, V.; Nordström, T.; Mörgelin, M.; Riesbeck, K. Moraxella catarrhalis outer membrane vesicles carry β-lactamase and promote survival of Streptococcus pneumoniae and Haemophilus influenzae by inactivating amoxicillin. Antimicrob. Agents Chemother. 2011, 55, 3845–3853. [Google Scholar] [CrossRef]
- Perez, A.C.; Pang, B.; King, L.B.; Tan, L.; Murrah, K.A.; Reimche, J.L.; Wren, J.T.; Richardson, S.H.; Ghandi, U.; Swords, W.E. Residence of Streptococcus pneumoniae and Moraxella catarrhalis within polymicrobial biofilm promotes antibiotic resistance and bacterial persistence in vivo. Pathog. Dis. 2014, 70, 280–288. [Google Scholar] [CrossRef]
- Tikhomirova, A.; Zilm, P.S.; Trappetti, C.; Paton, J.C.; Kidd, S.P. The central role of arginine in Haemophilus influenzae survival in a polymicrobial environment with Streptococcus pneumoniae and Moraxella catarrhalis. PLoS ONE 2022, 17, e0271912. [Google Scholar] [CrossRef]
- Ikryannikova, L.; Malakhova, M.; Lominadze, G.; Karpova, I.Y.; Kostryukova, E.; Mayansky, N.; Kruglov, A.; Klimova, E.; Lisitsina, E.; Ilina, E. Inhibitory effect of streptococci on the growth of M. catarrhalis strains and the diversity of putative bacteriocin-like gene loci in the genomes of S. pneumoniae and its relatives. AMB Express 2017, 7, 218. [Google Scholar] [CrossRef]
- Tikhomirova, A.; Kidd, S.P. Haemophilus influenzae and Streptococcus pneumoniae: Living together in a biofilm. Pathog. Dis. 2013, 69, 114–126. [Google Scholar] [CrossRef]
- Bair, K.L.; Campagnari, A.A. Moraxella catarrhalis promotes stable polymicrobial biofilms with the major otopathogens. Front. Microbiol. 2020, 10, 3006. [Google Scholar] [CrossRef]
- Atto, B.; Eapen, M.S.; Sharma, P.; Frey, U.; Ammit, A.J.; Markos, J.; Chia, C.; Larby, J.; Haug, G.; Weber, H.C. New therapeutic targets for the prevention of infectious acute exacerbations of COPD: Role of epithelial adhesion molecules and inflammatory pathways. Clin. Sci. 2019, 133, 1663–1703. [Google Scholar] [CrossRef]
- Geme III, J.W.S. Molecular and cellular determinants of non-typeable Haemophilus influenzae adherence and invasion. Cell. Microbiol. 2002, 4, 191–200. [Google Scholar] [CrossRef]
- Clark, S.E.; Weiser, J.N. Microbial modulation of host immunity with the small molecule phosphorylcholine. Infect. Immun. 2013, 81, 392–401. [Google Scholar] [CrossRef]
- Cundell, D.R.; Gerard, C.; Idanpaan-Heikkila, I.; Tuomanen, E.I.; Gerard, N.P. PAf receptor anchors Streptococcus pneumoniae to activated human endothelial cells. In Platelet-Activating Factor and Related Lipid Mediators 2: Roles in Health and Disease; Plenum Press: New York, NY, USA, 1996; pp. 89–94. [Google Scholar]
- Slevogt, H.; Zabel, S.; Opitz, B.; Hocke, A.; Eitel, J.; N’Guessan, P.D.; Lucka, L.; Riesbeck, K.; Zimmermann, W.; Zweigner, J. CEACAM1 inhibits Toll-like receptor 2–triggered antibacterial responses of human pulmonary epithelial cells. Nat. Immunol. 2008, 9, 1270–1278. [Google Scholar] [CrossRef]
- Avadhanula, V.; Rodriguez, C.A.; Ulett, G.C.; Bakaletz, L.O.; Adderson, E.E. Nontypeable Haemophilus influenzae adheres to intercellular adhesion molecule 1 (ICAM-1) on respiratory epithelial cells and upregulates ICAM-1 expression. Infect. Immun. 2006, 74, 830–838. [Google Scholar] [CrossRef]
- Pickering, J.L.; Prosser, A.; Corscadden, K.J.; de Gier, C.; Richmond, P.C.; Zhang, G.; Thornton, R.B.; Kirkham, L.A. Haemophilus haemolyticus Interaction with Host Cells Is Different to Nontypeable Haemophilus influenzae and Prevents NTHi Association with Epithelial Cells. Front. Cell. Infect. Microbiol. 2016, 6, 50. [Google Scholar] [CrossRef]
- Atto, B.; Kunde, D.; Gell, D.A.; Tristram, S. Haemophilin-Producing Strains of Haemophilus haemolyticus Protect Respiratory Epithelia from NTHi Colonisation and Internalisation. Pathogens 2021, 10, 29. [Google Scholar] [CrossRef]
- Bomar, L.; Brugger, S.D.; Yost, B.H.; Davies, S.S.; Lemon, K.P. Corynebacterium accolens Releases Antipneumococcal Free Fatty Acids from Human Nostril and Skin Surface Triacylglycerols. mBio 2016, 7, e01725-15. [Google Scholar] [CrossRef]
- Sherrard, L.J.; McGrath, S.J.; McIlreavey, L.; Hatch, J.; Wolfgang, M.C.; Muhlebach, M.S.; Gilpin, D.F.; Elborn, J.S.; Tunney, M.M. Production of extended-spectrum beta-lactamases and the potential indirect pathogenic role of Prevotella isolates from the cystic fibrosis respiratory microbiota. Int. J. Antimicrob. Agents 2016, 47, 140–145. [Google Scholar] [CrossRef]
- Korppi, M. Mixed microbial aetiology of community-acquired pneumonia in children. Apmis 2002, 110, 515–522. [Google Scholar] [CrossRef]
- Gu, L.; Deng, H.; Ren, Z.; Zhao, Y.; Yu, S.; Guo, Y.; Dai, J.; Chen, X.; Li, K.; Li, R. Dynamic changes in the microbiome and mucosal immune microenvironment of the lower respiratory tract by influenza virus infection. Front. Microbiol. 2019, 10, 2491. [Google Scholar] [CrossRef]
- Molyneaux, P.L.; Mallia, P.; Cox, M.J.; Footitt, J.; Willis-Owen, S.A.; Homola, D.; Trujillo-Torralbo, M.-B.; Elkin, S.; Kon, O.M.; Cookson, W.O. Outgrowth of the bacterial airway microbiome after rhinovirus exacerbation of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2013, 188, 1224–1231. [Google Scholar] [CrossRef]
- Perez, G.F.; Pérez-Losada, M.; Isaza, N.; Rose, M.C.; Colberg-Poley, A.M.; Nino, G. Nasopharyngeal microbiome in premature infants and stability during rhinovirus infection. J. Investig. Med. 2017, 65, 984–990. [Google Scholar] [CrossRef]
- DeMuri, G.P.; Gern, J.E.; Eickhoff, J.C.; Lynch, S.V.; Wald, E.R. Dynamics of bacterial colonization with Streptococcus pneumoniae, Haemophilus influenzae, and Moraxella catarrhalis during symptomatic and asymptomatic viral upper respiratory tract infection. Clin. Infect. Dis. 2018, 66, 1045–1053. [Google Scholar] [CrossRef]
- Binks, M.J.; Cheng, A.C.; Smith-Vaughan, H.; Sloots, T.; Nissen, M.; Whiley, D.; McDonnell, J.; Leach, A.J. Viral-bacterial co-infection in Australian Indigenous children with acute otitis media. BMC Infect. Dis. 2011, 11, 161. [Google Scholar] [CrossRef]
- Rowe, H.M.; Meliopoulos, V.A.; Iverson, A.; Bomme, P.; Schultz-Cherry, S.; Rosch, J.W. Direct interactions with influenza promote bacterial adherence during respiratory infections. Nat. Microbiol. 2019, 4, 1328–1336. [Google Scholar] [CrossRef]
- Plotkowski, M.-C.; Puchelle, E.; Beck, G.; Jacquot, J.; Hannoun, C. Adherence of type I Streptococcus pneumoniae to tracheal epithelium of mice infected with influenza A/PR8 virus. Am. Rev. Respir. Dis. 1986, 134, 1040–1044. [Google Scholar] [CrossRef]
- Yokota, S.-i.; Okabayashi, T.; Yoto, Y.; Hori, T.; Tsutsumi, H.; Fujii, N. Fosfomycin suppresses RS-virus-induced Streptococcus pneumoniae and Haemophilus influenzae adhesion to respiratory epithelial cells via the platelet-activating factor receptor. FEMS Microbiol. Lett. 2010, 310, 84–90. [Google Scholar] [CrossRef]
- Toone, S.L.; Ratkiewicz, M.; Novotny, L.A.; Phong, B.L.; Bakaletz, L.O. Nontypeable haemophilus influenzae type IV pilus mediates augmented adherence to rhinovirus-infected human airway epithelial cells. Infect. Immun. 2020, 88, e00248-20. [Google Scholar] [CrossRef]
- Park, S.-S.; Gonzalez-Juarbe, N.; Riegler, A.N.; Im, H.; Hale, Y.; Platt, M.P.; Croney, C.; Briles, D.E.; Orihuela, C.J. Streptococcus pneumoniae binds to host GAPDH on dying lung epithelial cells worsening secondary infection following influenza. Cell Rep. 2021, 35, 109267. [Google Scholar] [CrossRef]
- Mokrzan, E.M.; Dairo, K.A.; Novotny, L.A.; Bakaletz, L.O. Nontypeable Haemophilus influenzae responds to virus-infected cells with a significant increase in type IV pilus expression. mSphere 2020, 5, e00384-20. [Google Scholar] [CrossRef]
- Ghoneim, H.E.; Thomas, P.G.; McCullers, J.A. Depletion of alveolar macrophages during influenza infection facilitates bacterial superinfections. J. Immunol. 2013, 191, 1250–1259. [Google Scholar] [CrossRef]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut–lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef]
- Madan, J.; Koestler, D.; Stanton, B.; Davidson, L.; Moulton, L.; Housman, M.; Moore, J.; Guill, M.F.; Morrison, H.; Sogin, M. Serial analysis of the gut and respiratory microbiome in cystic fibrosis in infancy: Interaction between intestinal and respiratory tracts and impact of nutritional exposures. mBio 2012, 3, e00251-12. [Google Scholar] [CrossRef]
- Keely, S.; Talley, N.J.; Hansbro, P.M. Pulmonary-intestinal cross-talk in mucosal inflammatory disease. Mucosal Immunol. 2012, 5, 7–18. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.-S.; Peng, S.-H.; Deng, X.-Y.; Zhu, D.-M.; Javidiparsijani, S.; Wang, G.-R.; Li, D.-Q.; Li, L.-X.; Wang, Y.-C. Gut-lung crosstalk in pulmonary involvement with inflammatory bowel diseases. World J. Gastroenterol. WJG 2013, 19, 6794. [Google Scholar] [CrossRef] [PubMed]
- Yazar, A.; Atis, S.; Konca, K.; Pata, C.; Akbay, E.; Calikoglu, M.; Hafta, A. Respiratory symptoms and pulmonary functional changes in patients with irritable bowel syndrome. Am. J. Gastroenterol. 2001, 96, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.D.; Budden, K.F.; Neal, R.; Hansbro, P.M. Microbiome effects on immunity, health and disease in the lung. Clin. Transl. Immunol. 2017, 6, e133. [Google Scholar] [CrossRef] [PubMed]
- Ruane, D.; Brane, L.; Reis, B.S.; Cheong, C.; Poles, J.; Do, Y.; Zhu, H.; Velinzon, K.; Choi, J.-H.; Studt, N. Lung dendritic cells induce migration of protective T cells to the gastrointestinal tract. J. Exp. Med. 2013, 210, 1871–1888. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.; Bazin, T.; Truchetet, M.-E.; Schaeverbeke, T.; Delhaes, L.; Pradeu, T. Protective microbiota: From localized to long-reaching co-immunity. Front. Immunol. 2017, 8, 1678. [Google Scholar] [CrossRef]
- Grier, A.; McDavid, A.; Wang, B.; Qiu, X.; Java, J.; Bandyopadhyay, S.; Yang, H.; Holden-Wiltse, J.; Kessler, H.A.; Gill, A.L. Neonatal gut and respiratory microbiota: Coordinated development through time and space. Microbiome 2018, 6, 193. [Google Scholar] [CrossRef]
- Tamburini, S.; Clemente, J.C. Gut microbiota: Neonatal gut microbiota induces lung immunity against pneumonia. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 263–264. [Google Scholar] [CrossRef]
- Gray, J.; Oehrle, K.; Worthen, G.; Alenghat, T.; Whitsett, J.; Deshmukh, H. Intestinal commensal bacteria mediate lung mucosal immunity and promote resistance of newborn mice to infection. Sci. Transl. Med. 2017, 9, eaaf9412. [Google Scholar] [CrossRef]
- Ji, J.J.; Sun, Q.M.; Nie, D.Y.; Wang, Q.; Zhang, H.; Qin, F.F.; Wang, Q.S.; Lu, S.F.; Pang, G.M.; Lu, Z.G. Probiotics protect against RSV infection by modulating the microbiota-alveolar-macrophage axis. Acta Pharmacol. Sin. 2021, 42, 1630–1641. [Google Scholar] [CrossRef]
- Sikder, M.A.A.; Rashid, R.B.; Ahmed, T.; Sebina, I.; Howard, D.R.; Ullah, M.A.; Rahman, M.M.; Lynch, J.P.; Curren, B.; Werder, R.B.; et al. Maternal diet modulates the infant microbiome and intestinal Flt3L necessary for dendritic cell development and immunity to respiratory infection. Immunity 2023, 56, 1098–1114.e1010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, S.; Wang, N.; Tan, H.-Y.; Zhang, Z.; Feng, Y. The cross-talk between gut microbiota and lungs in common lung diseases. Front. Microbiol. 2020, 11, 301. [Google Scholar] [CrossRef] [PubMed]
- Soret, P.; Vandenborght, L.-E.; Francis, F.; Coron, N.; Enaud, R.; Avalos, M.; Schaeverbeke, T.; Berger, P.; Fayon, M.; Thiebaut, R. Respiratory mycobiome and suggestion of inter-kingdom network during acute pulmonary exacerbation in cystic fibrosis. Sci. Rep. 2020, 10, 3589. [Google Scholar] [CrossRef] [PubMed]
- Frati, F.; Salvatori, C.; Incorvaia, C.; Bellucci, A.; Di Cara, G.; Marcucci, F.; Esposito, S. The role of the microbiome in asthma: The gut–lung axis. Int. J. Mol. Sci. 2018, 20, 123. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Nuñez, M.; Millares, L.; Pomares, X.; Ferrari, R.; Pérez-Brocal, V.; Gallego, M.; Espasa, M.; Moya, A.; Monsó, E. Severity-related changes of bronchial microbiome in chronic obstructive pulmonary disease. J. Clin. Microbiol. 2014, 52, 4217–4223. [Google Scholar] [CrossRef] [PubMed]
- Narayana, J.K.; Aliberti, S.; Mac Aogain, M.; Jaggi, T.K.; Ali, N.; Ivan, F.X.; Cheng, H.S.; Yip, Y.S.; Vos, M.I.G.; Low, Z.S.; et al. Microbial Dysregulation of the Gut-Lung Axis in Bronchiectasis. Am. J. Respir. Crit. Care Med. 2023, 207, 908–920. [Google Scholar] [CrossRef] [PubMed]
- Björkstén, B.; Sepp, E.; Julge, K.; Voor, T.; Mikelsaar, M. Allergy development and the intestinal microflora during the first year of life. J. Allergy Clin. Immunol. 2001, 108, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Stiemsma, L.T.; Arrieta, M.-C.; Dimitriu, P.A.; Cheng, J.; Thorson, L.; Lefebvre, D.L.; Azad, M.B.; Subbarao, P.; Mandhane, P.; Becker, A. Shifts in Lachnospira and Clostridium sp. in the 3-month stool microbiome are associated with preschool age asthma. Clin. Sci. 2016, 130, 2199–2207. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef]
- Kalliomäki, M.; Kirjavainen, P.; Eerola, E.; Kero, P.; Salminen, S.; Isolauri, E. Distinct patterns of neonatal gut microflora in infants in whom atopy was and was not developing. J. Allergy Clin. Immunol. 2001, 107, 129–134. [Google Scholar] [CrossRef]
- Narayana, J.K.; Tsaneva-Atanasova, K.; Chotirmall, S.H. Microbiomics-focused Data Integration: A Fresh Solve for the Rubik’s Cube of Endophenotyping? Am. J. Respir. Crit. Care Med. 2022, 206, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Charalampous, T.; Kay, G.L.; Richardson, H.; Aydin, A.; Baldan, R.; Jeanes, C.; Rae, D.; Grundy, S.; Turner, D.J.; Wain, J.; et al. Nanopore metagenomics enables rapid clinical diagnosis of bacterial lower respiratory infection. Nat. Biotechnol. 2019, 37, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.R.; Rawat, A.; Tang, P.; Jithesh, P.V.; Thomas, E.; Tan, R.; Tilley, P. Depletion of Human DNA in Spiked Clinical Specimens for Improvement of Sensitivity of Pathogen Detection by Next-Generation Sequencing. J. Clin. Microbiol. 2016, 54, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Gan, M.; Wu, B.; Yan, G.; Li, G.; Sun, L.; Lu, G.; Zhou, W. Combined nanopore adaptive sequencing and enzyme-based host depletion efficiently enriched microbial sequences and identified missing respiratory pathogens. BMC Genom. 2021, 22, 732. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, L.; Heyderman, R. The challenges of defining the human nasopharyngeal resistome. Trends Microbiol. 2023, 31, 816–831. [Google Scholar] [CrossRef] [PubMed]
- Mac Aogáin, M.; Narayana, J.K.; Tiew, P.Y.; Ali, N.A.t.B.M.; Yong, V.F.L.; Jaggi, T.K.; Lim, A.Y.H.; Keir, H.R.; Dicker, A.J.; Thng, K.X. Integrative microbiomics in bronchiectasis exacerbations. Nat. Med. 2021, 27, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Grimwood, K.; Bell, S.C.; Chang, A.B. Antimicrobial treatment of non-cystic fibrosis bronchiectasis. Expert Rev. Anti-Infect. Ther. 2014, 12, 1277–1296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, D.; Zhou, J.; Yang, Q.; Ying, Y.; Li, N.; Cao, L.; Wang, W.; Ma, X. The Utility of Metagenomic Next-Generation Sequencing (mNGS) in the Management of Patients With Bronchiectasis: A Single-Center Retrospective Study of 93 Cases. Open Forum Infect. Dis. 2023, 10, ofad425. [Google Scholar] [CrossRef]
- Liu, X.; Locasale, J.W. Metabolomics: A Primer. Trends Biochem. Sci. 2017, 42, 274–284. [Google Scholar] [CrossRef]
- Drabinska, N.; Flynn, C.; Ratcliffe, N.; Belluomo, I.; Myridakis, A.; Gould, O.; Fois, M.; Smart, A.; Devine, T.; Costello, B.L. A literature survey of all volatiles from healthy human breath and bodily fluids: The human volatilome. J. Breath Res. 2021, 15, 034001. [Google Scholar] [CrossRef]
- Filipiak, W.; Sponring, A.; Baur, M.M.; Ager, C.; Filipiak, A.; Wiesenhofer, H.; Nagl, M.; Troppmair, J.; Amann, A. Characterization of volatile metabolites taken up by or released from Streptococcus pneumoniae and Haemophilus influenzae by using GC-MS. Microbiology 2012, 158, 3044–3053. [Google Scholar] [CrossRef] [PubMed]
- Mellors, T.R.; Rees, C.A.; Franchina, F.A.; Burklund, A.; Patel, C.; Hathaway, L.J.; Hill, J.E. The volatile molecular profiles of seven Streptococcus pneumoniae serotypes. J. Chromatogr. B 2018, 1096, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Abd El Qader, A.; Lieberman, D.; Shemer Avni, Y.; Svobodin, N.; Lazarovitch, T.; Sagi, O.; Zeiri, Y. Volatile organic compounds generated by cultures of bacteria and viruses associated with respiratory infections. Biomed. Chromatogr. 2015, 29, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- Neerincx, A.H.; Geurts, B.P.; van Loon, J.; Tiemes, V.; Jansen, J.J.; Harren, F.J.; Kluijtmans, L.A.; Merkus, P.J.; Cristescu, S.M.; Buydens, L.M.; et al. Detection of Staphylococcus aureus in cystic fibrosis patients using breath VOC profiles. J. Breath Res. 2016, 10, 046014. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, F.J.; Bright-Thomas, R.J.; Jones, A.M.; Smith, D.; Spanel, P.; Webb, A.K.; Lenney, W. Hydrogen cyanide concentrations in the breath of adult cystic fibrosis patients with and without Pseudomonas aeruginosa infection. J. Breath Res. 2013, 7, 026010. [Google Scholar] [CrossRef] [PubMed]
- Španěl, P.; Sovová, K.; Dryahina, K.; Doušová, T.; Dřevínek, P.; Smith, D. Do linear logistic model analyses of volatile biomarkers in exhaled breath of cystic fibrosis patients reliably indicate Pseudomonas aeruginosa infection? J. Breath Res. 2016, 10, 036013. [Google Scholar] [CrossRef] [PubMed]
- Savelev, S.U.; Perry, J.D.; Bourke, S.J.; Jary, H.; Taylor, R.; Fisher, A.J.; Corris, P.A.; Petrie, M.; De Soyza, A. Volatile biomarkers of Pseudomonas aeruginosa in cystic fibrosis and noncystic fibrosis bronchiectasis. Lett. Appl. Microbiol. 2011, 52, 610–613. [Google Scholar] [CrossRef]
- Nasir, M.; Bean, H.D.; Smolinska, A.; Rees, C.A.; Zemanick, E.T.; Hill, J.E. Volatile molecules from bronchoalveolar lavage fluid can ‘rule-in’ Pseudomonas aeruginosa and ‘rule-out’ Staphylococcus aureus infections in cystic fibrosis patients. Sci. Rep. 2018, 8, 826. [Google Scholar] [CrossRef]
- Pollock, J.; Chalmers, J.D. The immunomodulatory effects of macrolide antibiotics in respiratory disease. Pulm. Pharmacol. Ther. 2021, 71, 102095. [Google Scholar] [CrossRef]
- Nickel, J.C.; Ruseska, I.; Wright, J.B.; Costerton, J.W. Tobramycin resistance of Pseudomonas aeruginosa cells growing as a biofilm on urinary catheter material. Antimicrob. Agents Chemother. 1985, 27, 619–624. [Google Scholar] [CrossRef]
- Reimche, J.L.; Kirse, D.J.; Whigham, A.S.; Swords, W.E. Resistance of non-typeable Haemophilus influenzae biofilms is independent of biofilm size. Pathog. Dis. 2017, 75, ftw112. [Google Scholar] [CrossRef] [PubMed]
- Altenburg, J.; De Graaff, C.; Van Der Werf, T.; Boersma, W. Immunomodulatory effects of macrolide antibiotics–part 1: Biological mechanisms. Respiration 2010, 81, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Starner, T.D.; Shrout, J.D.; Parsek, M.R.; Appelbaum, P.C.; Kim, G. Subinhibitory concentrations of azithromycin decrease nontypeable Haemophilus influenzae biofilm formation and diminish established biofilms. Antimicrob. Agents Chemother. 2008, 52, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Dhand, R. The rationale and evidence for use of inhaled antibiotics to control Pseudomonas aeruginosa infection in non-cystic fibrosis bronchiectasis. J. Aerosol Med. Pulm. Drug Deliv. 2018, 31, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Cipolla, D.; Blanchard, J.; Gonda, I. Development of liposomal ciprofloxacin to treat lung infections. Pharmaceutics 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.-T.; Vidaillac, C.; Yu, H.; Yong, V.F.; Roizman, D.; Chandrasekaran, R.; Lim, A.Y.; Low, T.B.; Tan, G.L.; Abisheganaden, J.A. A new therapeutic avenue for bronchiectasis: Dry powder inhaler of ciprofloxacin nanoplex exhibits superior ex vivo mucus permeability and antibacterial efficacy to its native ciprofloxacin counterpart. Int. J. Pharm. 2018, 547, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.-T.; Hadinoto, K. Ternary nanoparticle complex of antibiotic, polyelectrolyte, and mucolytic enzyme as a potential antibiotic delivery system in bronchiectasis therapy. Colloids Surf. B Biointerfaces 2020, 193, 111095. [Google Scholar] [CrossRef]
- Blasi, F.; Page, C.; Rossolini, G.M.; Pallecchi, L.; Matera, M.G.; Rogliani, P.; Cazzola, M. The effect of N-acetylcysteine on biofilms: Implications for the treatment of respiratory tract infections. Respir. Med. 2016, 117, 190–197. [Google Scholar] [CrossRef]
- Oliva, A.; Pallecchi, L.; Rossolini, G.; Travaglino, F.; Zanatta, P. Rationale and evidence for the adjunctive use of N-acetylcysteine in multidrug-resistant infections. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 4316–4325. [Google Scholar]
- Tran, T.-T.; Hadinoto, K. A Potential Quorum-sensing inhibitor for bronchiectasis therapy: Quercetin–chitosan nanoparticle complex exhibiting superior inhibition of biofilm formation and swimming motility of Pseudomonas aeruginosa to the native quercetin. Int. J. Mol. Sci. 2021, 22, 1541. [Google Scholar] [CrossRef]
- Zemke, A.C.; Shiva, S.; Burns, J.L.; Moskowitz, S.M.; Pilewski, J.M.; Gladwin, M.T.; Bomberger, J.M. Nitrite modulates bacterial antibiotic susceptibility and biofilm formation in association with airway epithelial cells. Free. Radic. Biol. Med. 2014, 77, 307–316. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, A.E.; Barker, A.F.; Ilowite, J.S.; Fick, R.B.; rhDNase Study Group. Treatment of idiopathic bronchiectasis with aerosolized recombinant human DNase I. Chest 1998, 113, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Goodman, S.D.; Bakaletz, L.O. Bacterial biofilms utilize an underlying extracellular DNA matrix structure that can be targeted for biofilm resolution. Microorganisms 2022, 10, 466. [Google Scholar] [CrossRef] [PubMed]
- Brockson, M.E.; Novotny, L.A.; Mokrzan, E.M.; Malhotra, S.; Jurcisek, J.A.; Akbar, R.; Devaraj, A.; Goodman, S.D.; Bakaletz, L.O. Evaluation of the kinetics and mechanism of action of anti-integration host factor-mediated disruption of bacterial biofilms. Mol. Microbiol. 2014, 93, 1246–1258. [Google Scholar] [CrossRef]
- Mokrzan, E.M.; Ahearn, C.P.; Buzzo, J.R.; Novotny, L.A.; Zhang, Y.; Goodman, S.D.; Bakaletz, L.O. Nontypeable Haemophilus influenzae newly released (NRel) from biofilms by antibody-mediated dispersal versus antibody-mediated disruption are phenotypically distinct. Biofilm 2020, 2, 100039. [Google Scholar] [CrossRef] [PubMed]
- Mokrzan, E.M.; Novotny, L.A.; Brockman, K.L.; Bakaletz, L.O. Antibodies against the majority subunit (PilA) of the type IV pilus of nontypeable Haemophilus influenzae disperse Moraxella catarrhalis from a dual-species biofilm. mBio 2018, 9, e02423-18. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.S. Phage therapy—Constraints and possibilities. Upsala J. Med. Sci. 2014, 119, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Ecology of anti-biofilm agents I: Antibiotics versus bacteriophages. Pharmaceuticals 2015, 8, 525–558. [Google Scholar] [CrossRef]
- Tkhilaishvili, T.; Lombardi, L.; Klatt, A.-B.; Trampuz, A.; Di Luca, M. Bacteriophage Sb-1 enhances antibiotic activity against biofilm, degrades exopolysaccharide matrix and targets persisters of Staphylococcus aureus. Int. J. Antimicrob. Agents 2018, 52, 842–853. [Google Scholar] [CrossRef]
- Gordillo Altamirano, F.; Forsyth, J.H.; Patwa, R.; Kostoulias, X.; Trim, M.; Subedi, D.; Archer, S.K.; Morris, F.C.; Oliveira, C.; Kielty, L. Bacteriophage-resistant Acinetobacter baumannii are resensitized to antimicrobials. Nat. Microbiol. 2021, 6, 157–161. [Google Scholar] [CrossRef]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Hawkins, C.; Änggård, E.; Harper, D. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. Clin. Otolaryngol. 2009, 34, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Samaee, H.R.; Eslami, G.; Rahimzadeh, G.; Saeedi, M.; Badabi, A.D.; Asare-Addo, K.; Nokhodchi, A.; Roozbeh, F.; Moosazadeh, M.; Ghasemian, R. Inhalation phage therapy as a new approach to preventing secondary bacterial pneumonia in patients with moderate to severe COVID-19: A double-blind clinical trial study. J. Drug Deliv. Sci. Technol. 2023, 84, 104486. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Souli, M.; Billard, M.; Campbell, J.; Conrad, D.; Ellison, D.W.; Evans, B.; Evans, S.R.; Greenwood-Quaintance, K.E.; Filippov, A.A. Safety and microbiological activity of phage therapy in persons with cystic fibrosis colonized with Pseudomonas aeruginosa: Study protocol for a phase 1b/2, multicenter, randomized, double-blind, placebo-controlled trial. Trials 2022, 23, 1057. [Google Scholar] [CrossRef]
- Angrill, J.; Agusti, C.; De Celis, R.; Rañó, A.; Gonzalez, J.; Solé, T.; Xaubet, A.; Rodriguez-Roisin, R.; Torres, A. Bacterial colonisation in patients with bronchiectasis: Microbiological pattern and risk factors. Thorax 2002, 57, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Köhler, T.; Luscher, A.; Falconnet, L.; Resch, G.; McBride, R.; Mai, Q.-A.; Simonin, J.L.; Chanson, M.; Maco, B.; Galiotto, R. Personalized aerosolised bacteriophage treatment of a chronic lung infection due to multidrug-resistant Pseudomonas aeruginosa. Nat. Commun. 2023, 14, 3629. [Google Scholar] [CrossRef] [PubMed]
- Levêque, M.; Cassir, N.; Mathias, F.; Fevre, C.; Daviet, F.; Bermudez, J.; Brioude, G.; Peyron, F.; Reynaud-Gaubert, M.; Coiffard, B. Refractory Pseudomonas aeruginosa Bronchopulmonary Infection After Lung Transplantation for Common Variable Immunodeficiency Despite Maximal Treatment Including IgM/IgA-Enriched Immunoglobulins and Bacteriophage Therapy. Infect. Drug Resist. 2023, 16, 4265–4271. [Google Scholar] [CrossRef]
- Li, L.; Zhong, Q.; Zhao, Y.; Bao, J.; Liu, B.; Zhong, Z.; Wang, J.; Yang, L.; Zhang, T.; Cheng, M. First-in-human application of double-stranded RNA bacteriophage in the treatment of pulmonary Pseudomonas aeruginosa infection. Microb. Biotechnol. 2023, 16, 862–867. [Google Scholar] [CrossRef]
- Racenis, K.; Lacis, J.; Rezevska, D.; Mukane, L.; Vilde, A.; Putnins, I.; Djebara, S.; Merabishvili, M.; Pirnay, J.-P.; Kalnina, M. Successful Bacteriophage-Antibiotic Combination Therapy against Multidrug-Resistant Pseudomonas aeruginosa Left Ventricular Assist Device Driveline Infection. Viruses 2023, 15, 1210. [Google Scholar] [CrossRef]
- Debarbieux, L.; Leduc, D.; Maura, D.; Morello, E.; Criscuolo, A.; Grossi, O.; Balloy, V.; Touqui, L. Bacteriophages can treat and prevent Pseudomonas aeruginosa lung infections. J. Infect. Dis. 2010, 201, 1096–1104. [Google Scholar] [CrossRef]
- Le Guellec, S.; Pardessus, J.; Bodier-Montagutelli, E.; L’hostis, G.; Dalloneau, E.; Piel, D.; Samaï, H.C.; Guillon, A.; Mujic, E.; Guillot-Combe, E. Administration of Bacteriophages via Nebulization during Mechanical Ventilation: In Vitro Study and Lung Deposition in Macaques. Viruses 2023, 15, 602. [Google Scholar] [CrossRef] [PubMed]
- Prokopczuk, F.I.; Im, H.; Campos-Gomez, J.; Orihuela, C.J.; Martínez, E. Engineered superinfective Pf phage prevents dissemination of Pseudomonas aeruginosa in a mouse burn model. mBio 2023, 14, e00472-23. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.M.; Neill, D.R.; Kaman, B.; Sahota, J.S.; Clokie, M.R.; Winstanley, C.; Kadioglu, A. Phage therapy is highly effective against chronic lung infections with Pseudomonas aeruginosa. Thorax 2017, 72, 666–667. [Google Scholar] [CrossRef]
- Nicholls, P.; Clark, J.R.; Gu Liu, C.; Terwilliger, A.; Maresso, A.W. Class-Driven Synergy and Antagonism between a Pseudomonas Phage and Antibiotics. Infect. Immun. 2023, 14, e00065-23. [Google Scholar] [CrossRef]
- Martin, I.; Morales, S.; Alton, E.W.; Davies, J.C. Lytic Bacteriophage Is a Promising Adjunct to Common Antibiotics across Cystic Fibrosis Clinical Strains and Culture Models of Pseudomonas aeruginosa Infection. Antibiotics 2023, 12, 593. [Google Scholar] [CrossRef] [PubMed]
- Kaabi, S.A.G.; Musafer, H.K. An experimental mouse model for phage therapy of bacterial pathogens causing bacteremia. Microb. Pathog. 2019, 137, 103770. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, K.; Gerstmans, H.; Saafan, A.; Dishisha, T.; Briers, Y. The preclinical and clinical progress of bacteriophages and their lytic enzymes: The parts are easier than the whole. Viruses 2019, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Iszatt, J.J.; Larcombe, A.N.; Chan, H.-K.; Stick, S.M.; Garratt, L.W.; Kicic, A. Phage therapy for multi-drug resistant respiratory tract infections. Viruses 2021, 13, 1809. [Google Scholar] [CrossRef]
- Saha, S.; Ojobor, C.D.; Li, A.S.C.; Mackinnon, E.; North, O.I.; Bondy-Denomy, J.; Lam, J.S.; Ensminger, A.W.; Maxwell, K.L.; Davidson, A.R. F-Type Pyocins Are Diverse Noncontractile Phage Tail-Like Weapons for Killing Pseudomonas aeruginosa. J. Bacteriol. 2023, 205, e00029-23. [Google Scholar] [CrossRef]
- Wang, C.; Li, P.; Niu, W.; Yuan, X.; Liu, H.; Huang, Y.; An, X.; Fan, H.; Zhangxiang, L.; Mi, L. Protective and therapeutic application of the depolymerase derived from a novel KN1 genotype of Klebsiella pneumoniae bacteriophage in mice. Res. Microbiol. 2019, 170, 156–164. [Google Scholar] [CrossRef]
- Liu, H.; Hu, Z.; Li, M.; Yang, Y.; Lu, S.; Rao, X. Therapeutic potential of bacteriophage endolysins for infections caused by Gram-positive bacteria. J. Biomed. Sci. 2023, 30, 29. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, J.M.; Djurkovic, S.; Fischetti, V.A. Phage lytic enzyme Cpl-1 as a novel antimicrobial for pneumococcal bacteremia. Infect. Immun. 2003, 71, 6199–6204. [Google Scholar] [CrossRef] [PubMed]
- Letrado, P.; Corsini, B.; Díez-Martínez, R.; Bustamante, N.; Yuste, J.E.; García, P. Bactericidal synergism between antibiotics and phage endolysin Cpl-711 to kill multidrug-resistant pneumococcus. Future Microbiol. 2018, 13, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cerrato, V.; García, P.; Del Prado, G.; García, E.; Gracia, M.; Huelves, L.; Ponte, C.; Lopez, R.; Soriano, F. In vitro interactions of LytA, the major pneumococcal autolysin, with two bacteriophage lytic enzymes (Cpl-1 and Pal), cefotaxime and moxifloxacin against antibiotic-susceptible and-resistant Streptococcus pneumoniae strains. J. Antimicrob. Chemother. 2007, 60, 1159–1162. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Feng, C.; Ren, J.; Zhuang, X.; Zhang, Y.; Zhu, Y.; Dong, K.; He, P.; Guo, X.; Qin, J. A novel antimicrobial endolysin, LysPA26, against Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 293. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Walmagh, M.; Grymonprez, B.; Biebl, M.; Pirnay, J.-P.; Defraine, V.; Michiels, J.; Cenens, W.; Aertsen, A.; Miller, S. Art-175 is a highly efficient antibacterial against multidrug-resistant strains and persisters of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 3774–3784. [Google Scholar] [CrossRef] [PubMed]
- Fauconnier, A. Phage therapy regulation: From night to dawn. Viruses 2019, 11, 352. [Google Scholar] [CrossRef]
- Chang, A.B.; Masel, J.P.; Boyce, N.C.; Wheaton, G.; Torzillo, P.J. Non-CF bronchiectasis: Clinical and HRCT evaluation. Pediatr. Pulmonol. 2003, 35, 477–483. [Google Scholar] [CrossRef]
- McCallum, G.B.; Binks, M.J. The epidemiology of chronic suppurative lung disease and bronchiectasis in children and adolescents. Front. Pediatr. 2017, 5, 27. [Google Scholar] [CrossRef]
- Singleton, R.J.; Valery, P.C.; Morris, P.; Byrnes, C.A.; Grimwood, K.; Redding, G.; Torzillo, P.J.; McCallum, G.; Chikoyak, L.; Mobberly, C.; et al. Indigenous children from three countries with non-cystic fibrosis chronic suppurative lung disease/bronchiectasis. Pediatr. Pulmonol. 2014, 49, 189–200. [Google Scholar] [CrossRef]
- Singleton, R.; Morris, A.; Redding, G.; Poll, J.; Holck, P.; Martinez, P.; Kruse, D.; Bulkow, L.R.; Petersen, K.M.; Lewis, C. Bronchiectasis in Alaska Native children: Causes and clinical courses. Pediatr. Pulmonol. 2000, 29, 182–187. [Google Scholar] [CrossRef]
- Das, L.; Kovesi, T.A. Bronchiectasis in children from Qikiqtani (Baffin) Region, Nunavut, Canada. Ann. Am. Thorac. Soc. 2015, 12, 96–100. [Google Scholar] [CrossRef]
- Amenyogbe, N.; Kollmann, T.R.; Ben-Othman, R. Early-Life Host-Microbiome Interphase: The Key Frontier for Immune Development. Front. Pediatr. 2017, 5, 111. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed]
- Uzan-Yulzari, A.; Turta, O.; Belogolovski, A.; Ziv, O.; Kunz, C.; Perschbacher, S.; Neuman, H.; Pasolli, E.; Oz, A.; Ben-Amram, H.; et al. Neonatal antibiotic exposure impairs child growth during the first six years of life by perturbing intestinal microbial colonization. Nat. Commun. 2021, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.J.; Lynch, D.B.; Murphy, K.; Ulaszewska, M.; Jeffery, I.B.; O’Shea, C.A.; Watkins, C.; Dempsey, E.; Mattivi, F.; Tuohy, K.; et al. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET Cohort. Microbiome 2017, 5, 4. [Google Scholar] [CrossRef]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The microbiome in early life: Implications for health outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Maldonado Galdeano, C.; Cazorla, S.I.; Lemme Dumit, J.M.; Velez, E.; Perdigon, G. Beneficial Effects of Probiotic Consumption on the Immune System. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Stiemsma, L.T.; Dimitriu, P.A.; Thorson, L.; Russell, S.; Yurist-Doutsch, S.; Kuzeljevic, B.; Gold, M.J.; Britton, H.M.; Lefebvre, D.L.; et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 2015, 7, 307ra152. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.K.Y.; Tao, J.; Chan, O.S.; Li, H.B.; Pang, H. Preventing Respiratory Tract Infections by Synbiotic Interventions: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv. Nutr. 2020, 11, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Dong, B.R.; Wu, T. Probiotics for preventing acute upper respiratory tract infections. Cochrane Database Syst. Rev. 2015, 8, CD006895. [Google Scholar] [CrossRef] [PubMed]
- Kukkonen, K.; Savilahti, E.; Haahtela, T.; Juntunen-Backman, K.; Korpela, R.; Poussa, T.; Tuure, T.; Kuitunen, M. Long-term safety and impact on infection rates of postnatal probiotic and prebiotic (synbiotic) treatment: Randomized, double-blind, placebo-controlled trial. Pediatrics 2008, 122, 8–12. [Google Scholar] [CrossRef]
- Luoto, R.; Ruuskanen, O.; Waris, M.; Kalliomaki, M.; Salminen, S.; Isolauri, E. Prebiotic and probiotic supplementation prevents rhinovirus infections in preterm infants: A randomized, placebo-controlled trial. J. Allergy Clin. Immunol. 2014, 133, 405–413. [Google Scholar] [CrossRef]
- Panigrahi, P.; Parida, S.; Nanda, N.C.; Satpathy, R.; Pradhan, L.; Chandel, D.S.; Baccaglini, L.; Mohapatra, A.; Mohapatra, S.S.; Misra, P.R.; et al. A randomized synbiotic trial to prevent sepsis among infants in rural India. Nature 2017, 548, 407–412. [Google Scholar] [CrossRef]
- Puccio, G.; Cajozzo, C.; Meli, F.; Rochat, F.; Grathwohl, D.; Steenhout, P. Clinical evaluation of a new starter formula for infants containing live Bifidobacterium longum BL999 and prebiotics. Nutrition 2007, 23, 1–8. [Google Scholar] [CrossRef]
- Rautava, S.; Salminen, S.; Isolauri, E. Specific probiotics in reducing the risk of acute infections in infancy—A randomised, double-blind, placebo-controlled study. Br. J. Nutr. 2009, 101, 1722–1726. [Google Scholar] [CrossRef]
- Taipale, T.; Pienihakkinen, K.; Isolauri, E.; Larsen, C.; Brockmann, E.; Alanen, P.; Jokela, J.; Soderling, E. Bifidobacterium animalis subsp. lactis BB-12 in reducing the risk of infections in infancy. Br. J. Nutr. 2011, 105, 409–416. [Google Scholar] [CrossRef]
- Taipale, T.J.; Pienihakkinen, K.; Isolauri, E.; Jokela, J.T.; Soderling, E.M. Bifidobacterium animalis subsp. lactis BB-12 in reducing the risk of infections in early childhood. Pediatr. Res. 2016, 79, 65–69. [Google Scholar] [CrossRef]
- Segal, L.N.; Clemente, J.C.; Tsay, J.C.; Koralov, S.B.; Keller, B.C.; Wu, B.G.; Li, Y.; Shen, N.; Ghedin, E.; Morris, A.; et al. Enrichment of the lung microbiome with oral taxa is associated with lung inflammation of a Th17 phenotype. Nat. Microbiol. 2016, 1, 16031. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.G.; Sulaiman, I.; Tsay, J.J.; Perez, L.; Franca, B.; Li, Y.; Wang, J.; Gonzalez, A.N.; El-Ashmawy, M.; Carpenito, J.; et al. Episodic Aspiration with Oral Commensals Induces a MyD88-dependent, Pulmonary T-Helper Cell Type 17 Response that Mitigates Susceptibility to Streptococcus pneumoniae. Am. J. Respir. Crit. Care Med. 2021, 203, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; de Steenhuijsen Piters, W.A.A.; van Houten, M.A.; Chu, M.; Biesbroek, G.; Kool, J.; Pernet, P.; de Groot, P.C.M.; Eijkemans, M.J.C.; Keijser, B.J.F.; et al. Maturation of the Infant Respiratory Microbiota, Environmental Drivers, and Health Consequences. A Prospective Cohort Study. Am. J. Respir. Crit. Care Med. 2017, 196, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.P.; Capozzi, V.; Russo, P.; Drider, D.; Spano, G.; Fiocco, D. Immunobiosis and probiosis: Antimicrobial activity of lactic acid bacteria with a focus on their antiviral and antifungal properties. Appl. Microbiol. Biotechnol. 2018, 102, 9949–9958. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chai, W.; Burwinkel, M.; Twardziok, S.; Wrede, P.; Palissa, C.; Esch, B.; Schmidt, M.F.G. Inhibitory Influence of Enterococcus faecium on the Propagation of Swine Influenza A Virus In Vitro. PLoS ONE 2013, 8, e53043. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Huang, L.; Zhu, L.; Mou, C.; Hou, Q.; Yu, Q. Inhibition of H9N2 Virus Invasion into Dendritic Cells by the S-Layer Protein from L. acidophilus ATCC 4356. Front. Cell. Infect. Microbiol. 2016, 6, 137. [Google Scholar] [CrossRef] [PubMed]
- Al Kassaa, I.; Hober, D.; Hamze, M.; Chihib, N.E.; Drider, D. Antiviral Potential of Lactic Acid Bacteria and Their Bacteriocins. Probiotics Antimicrob. Proteins 2014, 6, 177–185. [Google Scholar] [CrossRef]
- Oslund, K.L.; Baumgarth, N. Influenza-induced innate immunity: Regulators of viral replication, respiratory tract pathology & adaptive immunity. Future Virol. 2011, 6, 951–962. [Google Scholar] [CrossRef]
- Garcia-Crespo, K.E.; Chan, C.C.; Gabryszewski, S.J.; Percopo, C.M.; Rigaux, P.; Dyer, K.D.; Domachowske, J.B.; Rosenberg, H.F. Lactobacillus priming of the respiratory tract: Heterologous immunity and protection against lethal pneumovirus infection. Antivir. Res. 2013, 97, 270–279. [Google Scholar] [CrossRef]
- Clua, P.; Kanmani, P.; Zelaya, H.; Tada, A.; Kober, A.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Peptidoglycan from Immunobiotic Lactobacillus rhamnosus Improves Resistance of Infant Mice to Respiratory Syncytial Viral Infection and Secondary Pneumococcal Pneumonia. Front. Immunol. 2017, 8, 948. [Google Scholar] [CrossRef]
- Groeger, D.; Schiavi, E.; Grant, R.; Kurnik-Łucka, M.; Michalovich, D.; Williamson, R.; Beinke, S.; Kiely, B.; Akdis, C.A.; Hessel, E.M.; et al. Intranasal Bifidobacterium longum protects against viral-induced lung inflammation and injury in a murine model of lethal influenza infection. EBioMedicine 2020, 60, 102981. [Google Scholar] [CrossRef] [PubMed]
- Atto, B.; Kunde, D.; Gell, D.A.; Tristram, S. Oropharyngeal Carriage of hpl-Containing Haemophilus haemolyticus Predicts Lower Prevalence and Density of NTHi Colonisation in Healthy Adults. Pathogens 2021, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, L.; Jiang, N.; Liao, Y.; Kong, L.; Song, Y.; Xu, J.; Cao, J.; Li, Y.; Que, C.; et al. Effect of OM-85 BV on reducing bronchiectasis exacerbation in Chinese patients: The iPROBE study. J. Thorac. Dis. 2021, 13, 1641–1651. [Google Scholar] [CrossRef] [PubMed]
- Mac Aogáin, M.; Chotirmall, S.H. Microbiology and the Microbiome in Bronchiectasis. Clin. Chest Med. 2022, 43, 23–34. [Google Scholar] [CrossRef]
- Earl, J.P.; Adappa, N.D.; Krol, J.; Bhat, A.S.; Balashov, S.; Ehrlich, R.L.; Palmer, J.N.; Workman, A.D.; Blasetti, M.; Sen, B.; et al. Species-level bacterial community profiling of the healthy sinonasal microbiome using Pacific Biosciences sequencing of full-length 16S rRNA genes. Microbiome 2018, 6, 190. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
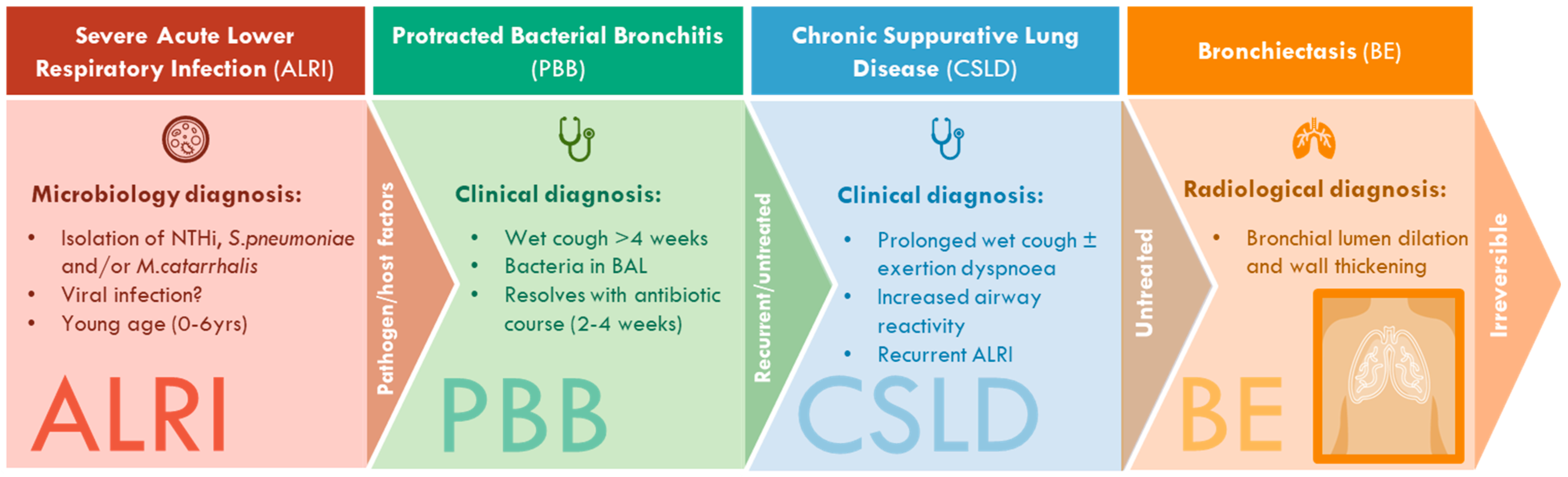
| Condition | Clinical Definition |
|---|---|
| PBB | Isolated chronic wet or productive cough without signs of another cause which typically resolves following a 2–4 week course of appropriate oral antibiotics [2]. |
| CSLD | Recurrent wet or productive cough episodes (≥3 per year), each lasting for >4 weeks, with or without other features (for example, exertional dyspnoea, symptoms of airway hyperresponiveness, recurrent chest infections, growth failure, clubbing, hyperinflation or chest wall deformity [3]. |
| BE | Symptoms and/or signs outlined for CLSD as well as characteristic radiographic features on chest high-resolution computed tomography (such as bronchial lumen dilation and wall thickening) [3]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atto, B.; Anteneh, Y.; Bialasiewicz, S.; Binks, M.J.; Hashemi, M.; Hill, J.; Thornton, R.B.; Westaway, J.; Marsh, R.L. The Respiratory Microbiome in Paediatric Chronic Wet Cough: What Is Known and Future Directions. J. Clin. Med. 2024, 13, 171. https://doi.org/10.3390/jcm13010171
Atto B, Anteneh Y, Bialasiewicz S, Binks MJ, Hashemi M, Hill J, Thornton RB, Westaway J, Marsh RL. The Respiratory Microbiome in Paediatric Chronic Wet Cough: What Is Known and Future Directions. Journal of Clinical Medicine. 2024; 13(1):171. https://doi.org/10.3390/jcm13010171
Chicago/Turabian StyleAtto, Brianna, Yitayal Anteneh, Seweryn Bialasiewicz, Michael J. Binks, Mostafa Hashemi, Jane Hill, Ruth B. Thornton, Jacob Westaway, and Robyn L. Marsh. 2024. "The Respiratory Microbiome in Paediatric Chronic Wet Cough: What Is Known and Future Directions" Journal of Clinical Medicine 13, no. 1: 171. https://doi.org/10.3390/jcm13010171
APA StyleAtto, B., Anteneh, Y., Bialasiewicz, S., Binks, M. J., Hashemi, M., Hill, J., Thornton, R. B., Westaway, J., & Marsh, R. L. (2024). The Respiratory Microbiome in Paediatric Chronic Wet Cough: What Is Known and Future Directions. Journal of Clinical Medicine, 13(1), 171. https://doi.org/10.3390/jcm13010171