
Biomarkers of Postpartum Depression: A Narrative Review

 ,
, 
Abstract
1. Introduction
2. Methods
3. Immunological Markers: Relevance to PPD
3.1. Kynurenine Pathway
3.2. Hormone Markers: Relevance to PPD
4. Multiple Pregnancy and Subsequent Pregnancies
5. Review of Hormonal Markers
5.1. Estrogen and Progesterone, Allopreganolone
5.2. HPA Axis
5.3. Prolactin and Oxytocin
5.4. Thyroid Hormones
5.5. Conclusions
6. Review of Inflammatory Markers
6.1. Tumor Necrosis Factor α (TNF-α)
6.2. C Reactive Protein
6.3. Transforming Growth Factor Beta (TGF-Beta β)
6.4. IL-10
6.5. IL-18
6.6. Chemokines
6.7. IL-1β
6.8. IFN-γ
6.9. IL-6
6.10. IL-2 and Soluble Interleukin-2 Receptor (sIL-2R)
6.11. IL-4
6.12. IL-8
6.13. IL-17
6.14. BDNF
6.15. IL-3
6.16. Conclusions
7. Nutrients
7.1. Vitamin A
7.2. B Vitamins
7.3. Vitamin C
7.4. Vitamin D
7.5. Vitamin E
7.6. Zinc
7.7. Iron
7.8. Selenium
7.9. Magnesium
7.10. Conclusions
8. Review of Metabolic Markers and Other Metabolic Substances
8.1. Insulin
8.2. Uric Acid
8.3. Homocysteine
8.4. Tyrosine
8.5. Vanillylmandelic Acid
8.6. Alanine
8.7. Conclusions
9. Metabolic Lipid Markers
9.1. PUFAs
9.2. Cholesterol
9.3. Conclusions
10. Oxidative Stress in Pregnancy
10.1. MPO
10.2. NO and NOS
10.3. MnSOD
10.4. Lipid Peroxidase
10.5. Malondialdehyde (MDA)
10.6. Conclusions
11. Genetic and Epigenetic Factors
12. Limitations
13. Conclusions
Funding
Conflicts of Interest
References
- Zhu, J.; Jin, J.; Tang, J. Inflammatory pathophysiological mechanisms implicated in postpartum depression. Front. Pharmacol. 2022, 13, 955672. [Google Scholar] [CrossRef] [PubMed]
- Waszkiewicz, N. Mentally Sick or Not-(Bio)Markers of Psychiatric Disorders Needed. J. Clin. Med. 2020, 9, 2375. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Stevens, W.; Peeters, D.; DeClerck, L.; Scharpe, S.; Bridts, C.; Schotte, C.; Cosyns, P. A study on the blunted natural killer cell activity in severely depressed patients. Life Sci. 1992, 50, 505–513. [Google Scholar] [CrossRef]
- Grosse, L.; Carvalho, L.A.; Birkenhager, T.K.; Hoogendijk, W.J.; Kushner, S.A.; Drexhage, H.A.; Bergink, V. Circulating cytotoxic T cells and natural killer cells as potential predictors for antidepressant response in melancholic depression. Restoration of T regulatory cell populations after antidepressant therapy. Psychopharmacology 2016, 233, 1679–1688. [Google Scholar] [CrossRef]
- Groer, M.W.; Morgan, K. Immune, health and endocrine characteristics of depressed postpartum mothers. Psychoneuroendocrinology 2007, 32, 133–139. [Google Scholar] [CrossRef]
- Groer, M.W.; El-Badri, N.; Djeu, J.; Williams, S.N.; Kane, B.; Szekeres, K. Suppression of natural killer cell cytotoxicity in postpartum women: Time course and potential mechanisms. Biol. Res. Nurs. 2014, 16, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Maes, M. Postpartum depression: Psychoneuroimmunological underpinnings and treatment. Neuropsychiatr. Dis. Treat. 2013, 9, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Colucci, F.; Kieckbusch, J. Maternal uterine natural killer cells nurture fetal growth: In medio stat virtus. Trends Mol. Med. 2015, 21, 60–67. [Google Scholar] [CrossRef]
- Moffett, A.; Colucci, F. Uterine NK cells: Active regulators at the maternal-fetal interface. J. Clin. Investig. 2014, 124, 1872–1879. [Google Scholar] [CrossRef]
- Ren, S.; Correia, M.A. Heme: A regulator of rat hepatic tryptophan 2,3-dioxygenase? Arch. Biochem. Biophys. 2000, 377, 195–203. [Google Scholar] [CrossRef]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [PubMed]
- Maes, M. Major depression and activation of the inflammatory response system. Adv. Exp. Med. Biol. 1999, 461, 25–46. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Walter, M.; Gos, T.; Guillemin, G.J.; Bernstein, H.G.; Sarnyai, Z.; Mawrin, C.; Brisch, R.; Bielau, H.; Meyer zu Schwabedissen, L.; et al. Severe depression is associated with increased microglial quinolinic acid in subregions of the anterior cingulate gyrus: Evidence for an immune-modulated glutamatergic neurotransmission? J. Neuroinflammation 2011, 8, 94. [Google Scholar] [CrossRef]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef] [PubMed]
- Slavich, G.M.; Irwin, M.R. From stress to inflammation and major depressive disorder: A social signal transduction theory of depression. Psychol. Bull. 2014, 140, 774–815. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.S.; Repasky, E.A.; Fisher, D.T. Fever and the thermal regulation of immunity: The immune system feels the heat. Nat. Rev. Immunol. 2015, 15, 335–349. [Google Scholar] [CrossRef]
- Larocca, L.; Ramhorst, R.; Roca, V.; Calafat, M.; Aisemberg, J.; Franchi, A.; Pérez Leirós, C. Neuroimmune-endocrine interactions during early pregnancy in an autoimmune context: Focus on macrophage activation. Neuroimmunomodulation 2008, 15, 84–90. [Google Scholar] [CrossRef]
- Nobis, A.; Zalewski, D.; Waszkiewicz, N. Peripheral Markers of Depression. J. Clin. Med. 2020, 9, 3793. [Google Scholar] [CrossRef]
- Amin, S.; Peterson, E.J.; Reed, A.M.; Mueller, D.L. Pregnancy and rheumatoid arthritis: Insights into the immunology of fetal tolerance and control of autoimmunity. Curr. Rheumatol. Rep. 2011, 13, 449–455. [Google Scholar] [CrossRef]
- Oracz, A.; Modzelewski, S.; Iłendo, K.; Sokół, A. Brexanolone and current methods of treating postpartum and perinatal depression. Pharmacother. Psychiatry Neurol. Farmakoter. W Psychiatr. I Neurol. 2023, 39, 53–64. [Google Scholar] [CrossRef]
- Balan, I.; Patterson, R.; Boero, G.; Krohn, H.; O’Buckley, T.K.; Meltzer-Brody, S.; Morrow, A.L. Brexanolone therapeutics in post-partum depression involves inhibition of systemic inflammatory pathways. EBioMedicine 2023, 89, 104473. [Google Scholar] [CrossRef]
- Bloch, M.; Daly, R.C.; Rubinow, D.R. Endocrine factors in the etiology of postpartum depression. Compr. Psychiatry 2003, 44, 234–246. [Google Scholar] [CrossRef]
- Freire-Garabal, M.; Núñez, M.J.; Balboa, J.; López-Delgado, P.; Gallego, R.; García-Caballero, T.; Fernández-Roel, M.D.; Brenlla, J.; Rey-Méndez, M. Serotonin upregulates the activity of phagocytosis through 5-HT1A receptors. Br. J. Pharmacol. 2003, 139, 457–463. [Google Scholar] [CrossRef]
- Worthen, R.J.; Beurel, E. Inflammatory and neurodegenerative pathophysiology implicated in postpartum depression. Neurobiol. Dis. 2022, 165, 105646. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, J.; Liu, Y.; Li, Z.; Li, X. TLR4-NF- κB Signal Involved in Depressive-Like Behaviors and Cytokine Expression of Frontal Cortex and Hippocampus in Stressed C57BL/6 and ob/ob Mice. Neural Plast. 2018, 2018, 7254016. [Google Scholar] [CrossRef]
- Sha, Q.; Madaj, Z.; Keaton, S.; Escobar Galvis, M.L.; Smart, L.; Krzyzanowski, S.; Fazleabas, A.T.; Leach, R.; Postolache, T.T.; Achtyes, E.D.; et al. Cytokines and tryptophan metabolites can predict depressive symptoms in pregnancy. Transl. Psychiatry 2022, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Achtyes, E.; Keaton, S.A.; Smart, L.; Burmeister, A.R.; Heilman, P.L.; Krzyzanowski, S.; Nagalla, M.; Guillemin, G.J.; Escobar Galvis, M.L.; Lim, C.K.; et al. Inflammation and kynurenine pathway dysregulation in post-partum women with severe and suicidal depression. Brain Behav. Immun. 2020, 83, 239–247. [Google Scholar] [CrossRef]
- Wang, S.; Quan, C.; Tan, Y.; Wen, S.; Zhang, J.; Duan, K. [Correlation between kynurenine metabolites and postpartum depression]. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2018, 43, 725–731. [Google Scholar] [CrossRef]
- Tulchinsky, D.; Hobel, C.J.; Yeager, E.; Marshall, J.R. Plasma estrone, estradiol, estriol, progesterone, and 17-hydroxyprogesterone in human pregnancy. I. Normal pregnancy. Am. J. Obstet. Gynecol. 1972, 112, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.S. The macrophage theory of depression. Med. Hypotheses 1991, 35, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Malyala, A.; Kelly, M.J.; Rønnekleiv, O.K. Estrogen modulation of hypothalamic neurons: Activation of multiple signaling pathways and gene expression changes. Steroids 2005, 70, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.; Maguire, J. The impact of tonic GABAA receptor-mediated inhibition on neuronal excitability varies across brain region and cell type. Front. Neural Circuits 2014, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Walton, N.; Maguire, J. Allopregnanolone-based treatments for postpartum depression: Why/how do they work? Neurobiol. Stress 2019, 11, 100198. [Google Scholar] [CrossRef] [PubMed]
- Maguire, J.; Ferando, I.; Simonsen, C.; Mody, I. Excitability changes related to GABAA receptor plasticity during pregnancy. J. Neurosci. 2009, 29, 9592–9601. [Google Scholar] [CrossRef] [PubMed]
- Patchev, V.K.; Shoaib, M.; Holsboer, F.; Almeida, O.F. The neurosteroid tetrahydroprogesterone counteracts corticotropin-releasing hormone-induced anxiety and alters the release and gene expression of corticotropin-releasing hormone in the rat hypothalamus. Neuroscience 1994, 62, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Stirone, C.; Duckles, S.P.; Krause, D.N.; Procaccio, V. Estrogen increases mitochondrial efficiency and reduces oxidative stress in cerebral blood vessels. Mol. Pharmacol. 2005, 68, 959–965. [Google Scholar] [CrossRef]
- Scharfman, H.E.; MacLusky, N.J. Estrogen-growth factor interactions and their contributions to neurological disorders. Headache 2008, 48 (Suppl. 2), S77–S89. [Google Scholar] [CrossRef]
- Khodadad, M.; Bahadoran, P.; Kheirabadi, G.R.; Sabzghabaee, A.M. Can Vitamin B6 Help to Prevent Postpartum Depression? A Randomized Controlled Trial. Int. J. Prev. Med. 2021, 12, 136. [Google Scholar]
- Barth, C.; Villringer, A.; Sacher, J. Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods. Front. Neurosci. 2015, 9, 37. [Google Scholar] [CrossRef]
- Mastorakos, G.; Ilias, I. Maternal and fetal hypothalamic-pituitary-adrenal axes during pregnancy and postpartum. Ann. N. Y Acad. Sci. 2003, 997, 136–149. [Google Scholar] [CrossRef]
- Chrousos, G.P.; Torpy, D.J.; Gold, P.W. Interactions between the hypothalamic-pituitary-adrenal axis and the female reproductive system: Clinical implications. Ann. Intern. Med. 1998, 129, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Ouakinin, S.R.S.; Barreira, D.P.; Gois, C.J. Depression and Obesity: Integrating the Role of Stress, Neuroendocrine Dysfunction and Inflammatory Pathways. Front. Endocrinol. 2018, 9, 431. [Google Scholar] [CrossRef]
- Maes, M.; Bosmans, E.; Meltzer, H.Y.; Scharpé, S.; Suy, E. Interleukin-1 beta: A putative mediator of HPA axis hyperactivity in major depression? Am. J. Psychiatry 1993, 150, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Pham, K.; McEwen, B.S.; Ledoux, J.E.; Nader, K. Fear learning transiently impairs hippocampal cell proliferation. Neuroscience 2005, 130, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.R.Y.B.; Tariq, A.; Lau, G.; Tok, N.W.K.; Tam, W.W.S.; Ho, C.S.H. Vitamin E, Alpha-Tocopherol, and Its Effects on Depression and Anxiety: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 656. [Google Scholar] [CrossRef]
- Smith, M.A.; Makino, S.; Kvetnansky, R.; Post, R.M. Stress and glucocorticoids affect the expression of brain-derived neurotrophic factor and neurotrophin-3 mRNAs in the hippocampus. J. Neurosci. 1995, 15, 1768–1777. [Google Scholar] [CrossRef]
- Hanson, N.D.; Owens, M.J.; Nemeroff, C.B. Depression, antidepressants, and neurogenesis: A critical reappraisal. Neuropsychopharmacology 2011, 36, 2589–2602. [Google Scholar] [CrossRef]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P.M. Maternal Immunological Adaptation During Normal Pregnancy. Front. Immunol. 2020, 11, 575197. [Google Scholar] [CrossRef]
- Kinder, J.M.; Turner, L.H.; Stelzer, I.A.; Miller-Handley, H.; Burg, A.; Shao, T.Y.; Pham, G.; Way, S.S. CD8+ T Cell Functional Exhaustion Overrides Pregnancy-Induced Fetal Antigen Alloimmunization. Cell Rep. 2020, 31, 107784. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.H.; Ertelt, J.M.; Xin, L.; Way, S.S. Pregnancy imprints regulatory memory that sustains anergy to fetal antigen. Nature 2012, 490, 102–106. [Google Scholar] [CrossRef]
- Bloch, M.; Schmidt, P.J.; Danaceau, M.; Murphy, J.; Nieman, L.; Rubinow, D.R. Effects of gonadal steroids in women with a history of postpartum depression. Am. J. Psychiatry 2000, 157, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.H.; Zhang, Z.H. Risk factors for postpartum depression: An evidence-based systematic review of systematic reviews and meta-analyses. Asian J. Psychiatr. 2020, 53, 102353. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Bishai, D.; Minkovitz, C.S. Multiple births are a risk factor for postpartum maternal depressive symptoms. Pediatrics 2009, 123, 1147–1154. [Google Scholar] [CrossRef]
- Liu, S.; Yan, Y.; Gao, X.; Xiang, S.; Sha, T.; Zeng, G.; He, Q. Risk factors for postpartum depression among Chinese women: Path model analysis. BMC Pregnancy Childbirth 2017, 17, 133. [Google Scholar] [CrossRef] [PubMed]
- Vigod, S.N.; Villegas, L.; Dennis, C.L.; Ross, L.E. Prevalence and risk factors for postpartum depression among women with preterm and low-birth-weight infants: A systematic review. BJOG 2010, 117, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Sibolboro Mezzacappa, E.; Endicott, J. Parity mediates the association between infant feeding method and maternal depressive symptoms in the postpartum. Arch. Womens Ment. Health 2007, 10, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, B.; Canário, C.; Field, T. Breastfeeding is negatively affected by prenatal depression and reduces postpartum depression. Psychol. Med. 2014, 44, 927–936. [Google Scholar] [CrossRef]
- Workman, J.L.; Barha, C.K.; Galea, L.A. Endocrine substrates of cognitive and affective changes during pregnancy and postpartum. Behav. Neurosci. 2012, 126, 54–72. [Google Scholar] [CrossRef]
- Meltzer-Brody, S.; Colquhoun, H.; Riesenberg, R.; Epperson, C.N.; Deligiannidis, K.M.; Rubinow, D.R.; Li, H.; Sankoh, A.J.; Clemson, C.; Schacterle, A.; et al. Brexanolone injection in post-partum depression: Two multicentre, double-blind, randomised, placebo-controlled, phase 3 trials. Lancet 2018, 392, 1058–1070. [Google Scholar] [CrossRef]
- Stefaniak, M.; Dmoch-Gajzlerska, E.; Jankowska, K.; Rogowski, A.; Kajdy, A.; Maksym, R.B. Progesterone and Its Metabolites Play a Beneficial Role in Affect Regulation in the Female Brain. Pharmaceuticals 2023, 16, 520. [Google Scholar] [CrossRef]
- Tiranini L, Nappi RE. Management of Premenstrual Syndrome: Green-top Guideline No. 48. BJOG 2017, 124, e73–e105. [Google Scholar] [CrossRef]
- O’Hara, M.W.; Schlechte, J.A.; Lewis, D.A.; Varner, M.W. Controlled prospective study of postpartum mood disorders: Psychological, environmental, and hormonal variables. J. Abnorm. Psychol. 1991, 100, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Harris, B. Biological and hormonal aspects of postpartum depressed mood. Br. J. Psychiatry 1994, 164, 288–292. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Abou-Saleh, M.T.; Ghubash, R.; Karim, L.; Krymski, M.; Bhai, I. Hormonal aspects of postpartum depression. Psychoneuroendocrinology 1998, 23, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Nott, P.N.; Franklin, M.; Armitage, C.; Gelder, M.G. Hormonal changes and mood in the puerperium. Br. J. Psychiatry 1976, 128, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Harris, B.; Johns, S.; Fung, H.; Thomas, R.; Walker, R.; Read, G.; Riad-Fahmy, D. The hormonal environment of post-natal depression. Br. J. Psychiatry 1989, 154, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Zonana, J.; Gorman, J.M. The neurobiology of postpartum depression. CNS Spectr. 2005, 10, 792–799, 805. [Google Scholar] [CrossRef]
- Pearson Murphy, B.E.; Steinberg, S.I.; Hu, F.Y.; Allison, C.M. Neuroactive ring A-reduced metabolites of progesterone in human plasma during pregnancy: Elevated levels of 5 alpha-dihydroprogesterone in depressed patients during the latter half of pregnancy. J. Clin. Endocrinol. Metab. 2001, 86, 5981–5987. [Google Scholar] [CrossRef] [PubMed]
- Goland, R.S.; Wardlaw, S.L.; Stark, R.I.; Brown, L.S.; Frantz, A.G. High levels of corticotropin-releasing hormone immunoactivity in maternal and fetal plasma during pregnancy. J. Clin. Endocrinol. Metab. 1986, 63, 1199–1203. [Google Scholar] [CrossRef]
- Thomson, M. The physiological roles of placental corticotropin releasing hormone in pregnancy and childbirth. J. Physiol. Biochem. 2013, 69, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.; Parker, G. The dexamethasone suppression test in the puerperium. Aust. N. Z. J. Psychiatry 1984, 18, 282–284. [Google Scholar] [CrossRef]
- Yim, I.S.; Glynn, L.M.; Dunkel-Schetter, C.; Hobel, C.J.; Chicz-DeMet, A.; Sandman, C.A. Risk of postpartum depressive symptoms with elevated corticotropin-releasing hormone in human pregnancy. Arch. Gen. Psychiatry 2009, 66, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Meltzer-Brody, S.; Stuebe, A.; Dole, N.; Savitz, D.; Rubinow, D.; Thorp, J. Elevated corticotropin releasing hormone (CRH) during pregnancy and risk of postpartum depression (PPD). J. Clin. Endocrinol. Metab. 2011, 96, E40–E47. [Google Scholar] [CrossRef]
- Melón, L.C.; Hooper, A.; Yang, X.; Moss, S.J.; Maguire, J. Inability to suppress the stress-induced activation of the HPA axis during the peripartum period engenders deficits in postpartum behaviors in mice. Psychoneuroendocrinology 2018, 90, 182–193. [Google Scholar] [CrossRef]
- Kammerer, M.; Taylor, A.; Glover, V. The HPA axis and perinatal depression: A hypothesis. Arch. Womens Ment. Health 2006, 9, 187–196. [Google Scholar] [CrossRef]
- Lamers, F.; de Jonge, P.; Nolen, W.A.; Smit, J.H.; Zitman, F.G.; Beekman, A.T.; Penninx, B.W. Identifying depressive subtypes in a large cohort study: Results from the Netherlands Study of Depression and Anxiety (NESDA). J. Clin. Psychiatry 2010, 71, 1582–1589. [Google Scholar] [CrossRef]
- Szpunar, M.J.; Parry, B.L. A systematic review of cortisol, thyroid-stimulating hormone, and prolactin in peripartum women with major depression. Arch. Womens Ment. Health 2018, 21, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Ystrom, E. Breastfeeding cessation and symptoms of anxiety and depression: A longitudinal cohort study. BMC Pregnancy Childbirth 2012, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Deems, N.P.; Leuner, B. Pregnancy, postpartum and parity: Resilience and vulnerability in brain health and disease. Front. Neuroendocr. 2020, 57, 100820. [Google Scholar] [CrossRef] [PubMed]
- Parry, B.L.; Sorenson, D.L.; Meliska, C.J.; Basavaraj, N.; Zirpoli, G.G.; Gamst, A.; Hauger, R. Hormonal basis of mood and postpartum disorders. Curr. Womens Health Rep. 2003, 3, 230–235. [Google Scholar]
- Okun, M.L.; Luther, J.; Prather, A.A.; Perel, J.M.; Wisniewski, S.; Wisner, K.L. Changes in sleep quality, but not hormones predict time to postpartum depression recurrence. J. Affect. Disord. 2011, 130, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.P.; Hibbeln, J.R.; Wisner, K.L.; Brumbach, B.H.; Watchman, M.; Gelenberg, A.J. Randomized dose-ranging pilot trial of omega-3 fatty acids for postpartum depression. Acta Psychiatr. Scand. 2006, 113, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Massey, S.H.; Backes, K.A.; Schuette, S.A. Plasma Oxytocin Concentration and Depressive Symptoms: A Review of Current Evidence and Directions for Future Research. Depress. Anxiety 2016, 33, 316–322. [Google Scholar] [CrossRef]
- Wysokiński, A.; Kłoszewska, I. Level of thyroid-stimulating hormone (TSH) in patients with acute schizophrenia, unipolar depression or bipolar disorder. Neurochem. Res. 2014, 39, 1245–1253. [Google Scholar] [CrossRef]
- O’Leary, P.C.; Boyne, P.; Atkinson, G.; Mileham, K.J.; James, I. Longitudinal study of serum thyroid hormone levels during normal pregnancy. Int. J. Gynaecol. Obstet. 1992, 38, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.S.; Sakowicz, A.; Roy, A.; Yang, A.; Sullivan, J.T.; Grobman, W.A.; Wisner, K.L. Plasma and cerebrospinal fluid inflammatory cytokines in perinatal depression. Am. J. Obstet. Gynecol. 2019, 220, 271.e1–271.e10. [Google Scholar] [CrossRef]
- Romanowska-Próchnicka, K.; Felis-Giemza, A.; Olesińska, M.; Wojdasiewicz, P.; Paradowska-Gorycka, A.; Szukiewicz, D. The Role of TNF-α and Anti-TNF-α Agents during Preconception, Pregnancy, and Breastfeeding. Int. J. Mol. Sci. 2021, 22, 2922. [Google Scholar] [CrossRef] [PubMed]
- Corwin, E.J.; Pajer, K.; Paul, S.; Lowe, N.; Weber, M.; McCarthy, D.O. Bidirectional psychoneuroimmune interactions in the early postpartum period influence risk of postpartum depression. Brain Behav. Immun. 2015, 49, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Buglione-Corbett, R.; Deligiannidis, K.M.; Leung, K.; Zhang, N.; Lee, M.; Rosal, M.C.; Moore Simas, T.A. Expression of inflammatory markers in women with perinatal depressive symptoms. Arch. Womens Ment. Health 2018, 21, 671–679. [Google Scholar] [CrossRef]
- Simpson, W.; Steiner, M.; Coote, M.; Frey, B.N. Relationship between inflammatory biomarkers and depressive symptoms during late pregnancy and the early postpartum period: A longitudinal study. Braz. J. Psychiatry 2016, 38, 190–196. [Google Scholar] [CrossRef]
- Karlsson, L.; Nousiainen, N.; Scheinin, N.M.; Maksimow, M.; Salmi, M.; Lehto, S.M.; Tolvanen, M.; Lukkarinen, H.; Karlsson, H. Cytokine profile and maternal depression and anxiety symptoms in mid-pregnancy-the FinnBrain Birth Cohort Study. Arch. Womens Ment. Health 2017, 20, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Szpakowski, M.; Nowak, M.; Oszukowski, P.; Wieczorek, A.; Skotnicka, A. [C-reactive protein in normal pregnancy]. Ginekol. Pol. 1996, 67, 17–20. [Google Scholar]
- Liu, H.; Zhang, Y.; Gao, Y.; Zhang, Z. Elevated levels of Hs-CRP and IL-6 after delivery are associated with depression during the 6 months post partum. Psychiatry Res. 2016, 243, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.S.; Hoxha, D.; Pinheiro, E.; Grobman, W.A.; Wisner, K.L. The association of serum C-reactive protein with the occurrence and course of postpartum depression. Arch. Womens Ment. Health 2019, 22, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Corwin, E.J.; Murray-Kolb, L.E.; Beard, J.L. Low hemoglobin level is a risk factor for postpartum depression. J. Nutr. 2003, 133, 4139–4142. [Google Scholar] [CrossRef]
- Sanjabi, S.; Zenewicz, L.A.; Kamanaka, M.; Flavell, R.A. Anti-inflammatory and pro-inflammatory roles of TGF-beta, IL-10, and IL-22 in immunity and autoimmunity. Curr. Opin. Pharmacol. 2009, 9, 447–453. [Google Scholar] [CrossRef]
- Moore, A.G.; Brown, D.A.; Fairlie, W.D.; Bauskin, A.R.; Brown, P.K.; Munier, M.L.C.; Russell, P.K.; Salamonsen, L.A.; Wallace, E.M.; Breit, S.N. The Transforming Growth Factor-β Superfamily Cytokine Macrophage Inhibitory Cytokine-1 Is Present in High Concentrations in the Serum of Pregnant Women1. J. Clin. Endocrinol. Metab. 2000, 85, 4781–4788. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhou, L.; Chen, J.; Li, M.; Xie, R. [Association between postpartum depression and concentrations of transforming growth factor-β in human colostrum: A nested cohort study]. Nan Fang. Yi Ke Da Xue Xue Bao 2022, 42, 1426–1430. [Google Scholar] [CrossRef]
- Gałecki, P.; Talarowska, M. Inflammatory theory of depression. Psychiatr. Pol. 2018, 52, 437–447. [Google Scholar] [CrossRef]
- Yu, Y.; Liang, H.-F.; Chen, J.; Li, Z.-B.; Han, Y.-S.; Chen, J.-X.; Li, J.-C. Postpartum Depression: Current Status and Possible Identification Using Biomarkers. Front. Psychiatry 2021, 12, 620371. [Google Scholar] [CrossRef]
- Köhler, C.A.; Freitas, T.H.; Maes, M.; de Andrade, N.Q.; Liu, C.S.; Fernandes, B.S.; Stubbs, B.; Solmi, M.; Veronese, N.; Herrmann, N.; et al. Peripheral cytokine and chemokine alterations in depression: A meta-analysis of 82 studies. Acta Psychiatr. Scand. 2017, 135, 373–387. [Google Scholar] [CrossRef]
- Dinarello, C.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 Binding Protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 regulates both Th1 and Th2 responses. Annu. Rev. Immunol. 2001, 19, 423–474. [Google Scholar] [CrossRef] [PubMed]
- Löb, S.; Ochmann, B.; Ma, Z.; Vilsmaier, T.; Kuhn, C.; Schmoeckel, E.; Herbert, S.L.; Kolben, T.; Wöckel, A.; Mahner, S.; et al. The role of Interleukin-18 in recurrent early pregnancy loss. J. Reprod. Immunol. 2021, 148, 103432. [Google Scholar] [CrossRef] [PubMed]
- Trifu, S.; Vladuti, A.; Popescu, A. The Neuroendocrinological Aspects of Pregnancy and Postpartum Depression. Acta Endocrinol. 2019, 15, 410–415. [Google Scholar] [CrossRef]
- Fransson, E.; Dubicke, A.; Byström, B.; Ekman-Ordeberg, G.; Hjelmstedt, A.; Lekander, M. Negative Emotions and Cytokines in Maternal and Cord Serum at Preterm Birth. Am. J. Reprod. Immunol. 2012, 67, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Osimo, E.F.; Pillinger, T.; Rodriguez, I.M.; Khandaker, G.M.; Pariante, C.M.; Howes, O.D. Inflammatory markers in depression: A meta-analysis of mean differences and variability in 5166 patients and 5083 controls. Brain Behav. Immun. 2020, 87, 901–909. [Google Scholar] [CrossRef]
- Camacho-Arroyo, I.; Flores-Ramos, M.; Mancilla-Herrera, I.; Cruz, F.M.C.; Hernández-Ruiz, J.; Diaz, G.P.; Labonne, B.F.; Del Pilar Meza-Rodríguez, M.; Gelman, P.L. Chemokine profile in women with moderate to severe anxiety and depression during pregnancy. BMC Pregnancy Childbirth 2021, 21, 807. [Google Scholar] [CrossRef]
- Edvinsson, Å.; Bränn, E.; Hellgren, C.; Freyhult, E.; White, R.; Kamali-Moghaddam, M.; Olivier, J.; Bergquist, J.; Boström, A.E.; Schiöth, H.B.; et al. Lower inflammatory markers in women with antenatal depression brings the M1/M2 balance into focus from a new direction. Psychoneuroendocrinology 2017, 80, 15–25. [Google Scholar] [CrossRef]
- Maes, M.; Abe, Y.; Sirichokchatchawan, W.; Suwimonteerabutr, J.; Sangkomkamhangd, U.; Almulla, A.F.; Satthapisit, S. The Cytokine, Chemokine, and Growth Factor Network of Prenatal Depression. Brain Sci. 2023, 13, 727. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Satyam, A.; Sharma, J.B. Leptin, IL-10 and inflammatory markers (TNF-alpha, IL-6 and IL-8) in pre-eclamptic, normotensive pregnant and healthy non-pregnant women. Am. J. Reprod. Immunol. 2007, 58, 21–30. [Google Scholar] [CrossRef]
- Petralia, M.C.; Mazzon, E.; Fagone, P.; Falzone, L.; Bramanti, P.; Nicoletti, F.; Basile, M.S. Retrospective follow-up analysis of the transcriptomic patterns of cytokines, cytokine receptors and chemokines at preconception and during pregnancy, in women with post-partum depression. Exp. Ther. Med. 2019, 18, 2055–2062. [Google Scholar] [CrossRef]
- Eyre, H.A.; Air, T.; Pradhan, A.; Johnston, J.; Lavretsky, H.; Stuart, M.J.; Baune, B.T. A meta-analysis of chemokines in major depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 68, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, N.; Kudo, K.; Mabuchi, T.; Takemoto, K.; Fujimaki, K.; Wati, H.; Iguchi, H.; Tezuka, H.; Kanba, S. Suppression of cell proliferation by interferon-alpha through interleukin-1 production in adult rat dentate gyrus. Neuropsychopharmacology 2006, 31, 2619–2626. [Google Scholar] [CrossRef] [PubMed]
- Corwin, E.J.; Johnston, N.; Pugh, L. Symptoms of postpartum depression associated with elevated levels of interleukin-1 beta during the first month postpartum. Biol. Res. Nurs. 2008, 10, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Fišar, Z.; Medina, M.; Scapagnini, G.; Nowak, G.; Berk, M. New drug targets in depression: Inflammatory, cell-mediated immune, oxidative and nitrosative stress, mitochondrial, antioxidant, and neuroprogressive pathways. And new drug candidates--Nrf2 activators and GSK-3 inhibitors. Inflammopharmacology 2012, 20, 127–150. [Google Scholar] [CrossRef] [PubMed]
- Sunico, C.R.; Portillo, F.; González-Forero, D.; Moreno-López, B. Nitric-oxide-directed synaptic remodeling in the adult mammal CNS. J. Neurosci. 2005, 25, 1448–1458. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K.; Glaser, R. Depression and immune function: Central pathways to morbidity and mortality. J. Psychosom. Res. 2002, 53, 873–876. [Google Scholar] [CrossRef]
- Chrousos, G.P. The hypothalamic-pituitary-adrenal axis and immune-mediated inflammation. N. Engl. J. Med. 1995, 332, 1351–1362. [Google Scholar] [CrossRef]
- Lenz, K.M.; Post, C.; Castaneda, A.J.; Banta, P.; Nelson, L.H.; Saulsbery, A.I.; Leuner, B. Abstract # 3185 Central immune alterations in a gestational stress animal model of postpartum depression. Brain Behav. Immun. 2019, 76, e38. [Google Scholar]
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK cells regulate key developmental processes at the human fetal-maternal interface. Nat. Med. 2006, 12, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Groer, M.E.; Jevitt, C.; Ji, M. Immune changes and dysphoric moods across the postpartum. Am. J. Reprod. Immunol. 2015, 73, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Han, Q.Q.; Gong, W.Q.; Pan, D.H.; Wang, L.Z.; Hu, W.; Yang, M.; Li, B.; Yu, J.; Liu, Q. Microglial activation mediates chronic mild stress-induced depressive- and anxiety-like behavior in adult rats. J. Neuroinflammation 2018, 15, 21. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Solmi, M.; Sanches, M.; Machado, M.O.; Stubbs, B.; Ajnakina, O.; Sherman, C.; Sun, Y.R.; Liu, C.S.; Brunoni, A.R.; et al. Evidence-based umbrella review of 162 peripheral biomarkers for major mental disorders. Transl. Psychiatry 2020, 10, 152. [Google Scholar] [CrossRef]
- Woodward, E.A.; Prêle, C.M.; Nicholson, S.E.; Kolesnik, T.B.; Hart, P.H. The anti-inflammatory effects of interleukin-4 are not mediated by suppressor of cytokine signalling-1 (SOCS1). Immunology 2010, 131, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Hu, X.; Jin, X. IL-4 as a potential biomarker for differentiating major depressive disorder from bipolar depression. Medicine 2023, 102, e33439. [Google Scholar] [CrossRef]
- Min, Z.; Li, Y.; Ying, H. Blood T-helper 17 cells and interleukin-17A correlate with the elevated risk of postpartum depression and anxiety. J. Clin. Lab. Anal. 2022, 36, e24559. [Google Scholar] [CrossRef]
- Szpunar, M.J.; Malaktaris, A.; Baca, S.A.; Hauger, R.L.; Lang, A.J. Are alterations in estradiol, cortisol, and inflammatory cytokines associated with depression during pregnancy and postpartum? An exploratory study. Brain Behav. Immun. Health 2021, 16, 100309. [Google Scholar] [CrossRef]
- Hong, J.; Hutton, G.J. Regulatory effects of interferon-β on osteopontin and interleukin-17 expression in multiple sclerosis. J. Interferon Cytokine Res. 2010, 30, 751–757. [Google Scholar] [CrossRef]
- Saraykar, S.; Cao, B.; Barroso, L.S.; Pereira, K.S.; Bertola, L.; Nicolau, M.; Ferreira, J.D.; Dias, N.S.; Vieira, E.L.; Teixeira, A.L.; et al. Plasma IL-17A levels in patients with late-life depression. Braz. J. Psychiatry 2018, 40, 212–215. [Google Scholar] [CrossRef]
- Kang, N.I.; Park, J.I.; Kim, Y.K.; Yang, J.C. Decreased Plasma BDNF Levels of Patients with Somatization Disorder. Psychiatry Investig. 2016, 13, 526–530. [Google Scholar] [CrossRef][Green Version]
- Gao, X.; Wang, J.; Yao, H.; Cai, Y.; Cheng, R. Serum BDNF concentration after delivery is associated with development of postpartum depression: A 3-month follow up study. J. Affect. Disord. 2016, 200, 25–30. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, K.H.; Lee, B.H.; Kim, Y.K. Plasma level of brain-derived neurotrophic factor (BDNF) in patients with postpartum depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 109, 110245. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.S.; Berk, M.; Turck, C.W.; Steiner, J.; Gonçalves, C.A. Decreased peripheral brain-derived neurotrophic factor levels are a biomarker of disease activity in major psychiatric disorders: A comparative meta-analysis. Mol. Psychiatry 2014, 19, 750–751. [Google Scholar] [CrossRef] [PubMed]
- Xiu, M.H.; Lin, C.G.; Tian, L.; Tan, Y.L.; Chen, J.; Chen, S.; Tan, S.P.; Wang, Z.R.; Yang, F.D.; Chen, D.C.; et al. Increased IL-3 serum levels in chronic patients with schizophrenia: Associated with psychopathology. Psychiatry Res. 2015, 229, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Rupanagunta, G.P.; Nandave, M.; Rawat, D.; Upadhyay, J.; Rashid, S.; Ansari, M.N. Postpartum depression: Aetiology, pathogenesis and the role of nutrients and dietary supplements in prevention and management. Saudi Pharm. J. 2023, 31, 1274–1293. [Google Scholar] [CrossRef]
- Sparling, T.M.; Waid, J.L.; Wendt, A.S.; Gabrysch, S. Depression among women of reproductive age in rural Bangladesh is linked to food security, diets and nutrition. Public. Health Nutr. 2020, 23, 660–673. [Google Scholar] [CrossRef]
- Mousa, A.; Naqash, A.; Lim, S. Macronutrient and Micronutrient Intake during Pregnancy: An Overview of Recent Evidence. Nutrients 2019, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Wisner, K.L. Nutrition and depression: Implications for improving mental health among childbearing-aged women. Biol. Psychiatry 2005, 58, 679–685. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, K.; Krishnan, A.; Cervenka, E.; Hu, G.; Guadagno, E.; Trakadis, Y. Biomarkers for major depressive and bipolar disorders using metabolomics: A systematic review. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2019, 180, 122–137. [Google Scholar] [CrossRef]
- Rouillon, F.; Thalassinos, M.; Miller, H.D.; Lemperiere, T. Folates and post partum depression. J. Affect. Disord. 1992, 25, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Abou-Saleh, M.T.; Ghubash, R.; Karim, L.; Krymski, M.; Anderson, D.N. The role of pterins and related factors in the biology of early postpartum depression. Eur. Neuropsychopharmacol. 1999, 9, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Ford, T.C.; Downey, L.A.; Simpson, T.; McPhee, G.; Oliver, C.; Stough, C. The Effect of a High-Dose Vitamin B Multivitamin Supplement on the Relationship between Brain Metabolism and Blood Biomarkers of Oxidative Stress: A Randomized Control Trial. Nutrients 2018, 10, 1860. [Google Scholar] [CrossRef]
- Rogne, T.; Tielemans, M.J.; Chong, M.F.; Yajnik, C.S.; Krishnaveni, G.V.; Poston, L.; Jaddoe, V.W.; Steegers, E.A.; Joshi, S.; Chong, Y.S.; et al. Associations of Maternal Vitamin B12 Concentration in Pregnancy with the Risks of Preterm Birth and Low Birth Weight: A Systematic Review and Meta-Analysis of Individual Participant Data. Am. J. Epidemiol. 2017, 185, 212–223. [Google Scholar] [CrossRef]
- Pierce, M.A.; Johnson, M.D.; Maciunas, R.J.; Murray, M.J.; Allen, G.S.; Harbison, M.A.; Creasy, J.L.; Kessler, R.M. Evaluating contrast-enhancing brain lesions in patients with AIDS by using positron emission tomography. Ann. Intern. Med. 1995, 123, 594–598. [Google Scholar] [CrossRef]
- Lindblad, M.; Tveden-Nyborg, P.; Lykkesfeldt, J. Regulation of vitamin C homeostasis during deficiency. Nutrients 2013, 5, 2860–2879. [Google Scholar] [CrossRef]
- Tveden-Nyborg, P.; Lykkesfeldt, J. Does vitamin C deficiency increase lifestyle-associated vascular disease progression? Evidence based on experimental and clinical studies. Antioxid. Redox Signal 2013, 19, 2084–2104. [Google Scholar] [CrossRef] [PubMed]
- Nauser, T.; Koppenol, W.H.; Schöneich, C. Protein thiyl radical reactions and product formation: A kinetic simulation. Free Radic. Biol. Med. 2015, 80, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Moritz, B.; Schmitz, A.E.; Rodrigues, A.L.S.; Dafre, A.L.; Cunha, M.P. The role of vitamin C in stress-related disorders. J. Nutr. Biochem. 2020, 85, 108459. [Google Scholar] [CrossRef]
- Ward, M.S.; Lamb, J.; May, J.M.; Harrison, F.E. Behavioral and monoamine changes following severe vitamin C deficiency. J. Neurochem. 2013, 124, 363–375. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Norman, A.W. From vitamin D to hormone D: Fundamentals of the vitamin D endocrine system essential for good health. Am. J. Clin. Nutr. 2008, 88, 491S–499S. [Google Scholar] [CrossRef]
- Heaney, R.P.; Weaver, C.M. Calcium and vitamin D. Endocrinol. Metab. Clin. N. Am. 2003, 32, 181–194. [Google Scholar] [CrossRef]
- Deluca, H.F.; Cantorna, M.T. Vitamin D: Its role and uses in immunology. FASEB J. 2001, 15, 2579–2585. [Google Scholar] [CrossRef] [PubMed]
- Amini, S.; Jafarirad, S.; Amani, R. Postpartum depression and vitamin D: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1514–1520. [Google Scholar] [CrossRef] [PubMed]
- Lansdowne, A.T.; Provost, S.C. Vitamin D3 enhances mood in healthy subjects during winter. Psychopharmacology 1998, 135, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Bertone-Johnson, E.R. Vitamin D and the occurrence of depression: Causal association or circumstantial evidence? Nutr. Rev. 2009, 67, 481–492. [Google Scholar] [CrossRef]
- Prohan, M.; Amani, R.; Nematpour, S.; Jomehzadeh, N.; Haghighizadeh, M.H. Total antioxidant capacity of diet and serum, dietary antioxidant vitamins intake, and serum hs-CRP levels in relation to depression scales in university male students. Redox Rep. 2014, 19, 133–139. [Google Scholar] [CrossRef]
- Maes, M.; De Vos, N.; Pioli, R.; Demedts, P.; Wauters, A.; Neels, H.; Christophe, A. Lower serum vitamin E concentrations in major depression. Another marker of lowered antioxidant defenses in that illness. J. Affect. Disord. 2000, 58, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Gautam, M.; Agrawal, M.; Sharma, P.; Gautam, A.S.; Gautam, S. Role of antioxidants in generalised anxiety disorder and depression. Indian J. Psychiatry 2012, 54, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, C.; Klabnik, J.J.; O’Donnell, J.M. Novel therapeutic targets in depression and anxiety: Antioxidants as a candidate treatment. Curr. Neuropharmacol. 2014, 12, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Grzeszczak, K.; Kwiatkowski, S.; Kosik-Bogacka, D. The Role of Fe, Zn, and Cu in Pregnancy. Biomolecules 2020, 10, 1176. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Evers, S.E.; Avison, W.R.; Campbell, M.K. Higher zinc intake buffers the impact of stress on depressive symptoms in pregnancy. Nutr. Res. 2010, 30, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.J.; Gong, B.; Xu, F.Y.; Luo, Y. Four trace elements in pregnant women and their relationships with adverse pregnancy outcomes. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4690–4697. [Google Scholar]
- Roomruangwong, C.; Kanchanatawan, B.; Sirivichayakul, S.; Mahieu, B.; Nowak, G.; Maes, M. Lower Serum Zinc and Higher CRP Strongly Predict Prenatal Depression and Physio-somatic Symptoms, Which All Together Predict Postnatal Depressive Symptoms. Mol. Neurobiol. 2017, 54, 1500–1512. [Google Scholar] [CrossRef]
- Aoki, C.; Imai, K.; Owaki, T.; Kobayashi-Nakano, T.; Ushida, T.; Iitani, Y.; Nakamura, N.; Kajiyama, H.; Kotani, T. The Possible Effects of Zinc Supplementation on Postpartum Depression and Anemia. Medicina 2022, 58, 731. [Google Scholar] [CrossRef] [PubMed]
- Ezzeddin, N.; Zavoshy, R.; Noroozi, M.; Sarichloo, M.E.; Jahanihashemi, H. The Association Between Postpartum Depression and Pica During Pregnancy. Glob. J. Health Sci. 2015, 8, 253–259. [Google Scholar] [CrossRef]
- Wang, J.; Um, P.; Dickerman, B.A.; Liu, J. Zinc, Magnesium, Selenium and Depression: A Review of the Evidence, Potential Mechanisms and Implications. Nutrients 2018, 10, 584. [Google Scholar] [CrossRef]
- Mokhber, N.; Namjoo, M.; Tara, F.; Boskabadi, H.; Rayman, M.P.; Ghayour-Mobarhan, M.; Sahebkar, A.; Majdi, M.R.; Tavallaie, S.; Azimi-Nezhad, M.; et al. Effect of supplementation with selenium on postpartum depression: A randomized double-blind placebo-controlled trial. J. Matern. Fetal Neonatal Med. 2011, 24, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Abumaria, N.; Yin, B.; Zhang, L.; Li, X.Y.; Chen, T.; Descalzi, G.; Zhao, L.; Ahn, M.; Luo, L.; Ran, C.; et al. Effects of elevation of brain magnesium on fear conditioning, fear extinction, and synaptic plasticity in the infralimbic prefrontal cortex and lateral amygdala. J. Neurosci. 2011, 31, 14871–14881. [Google Scholar] [CrossRef]
- Iosifescu, D.V.; Bolo, N.R.; Nierenberg, A.A.; Jensen, J.E.; Fava, M.; Renshaw, P.F. Brain bioenergetics and response to triiodothyronine augmentation in major depressive disorder. Biol. Psychiatry 2008, 63, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.S.; Sakowicz, A.; Roy, A.; Wang, A.; Yang, A.; Ciolino, J.; Grobman, W.A.; Wisner, K.L.; Yee, L.M. Is peripartum magnesium sulfate associated with a reduction in postpartum depressive symptoms? Am. J. Obstet. Gynecol. MFM 2021, 3, 100407. [Google Scholar] [CrossRef] [PubMed]
- Tarleton, E.K.; Littenberg, B.; MacLean, C.D.; Kennedy, A.G.; Daley, C. Role of magnesium supplementation in the treatment of depression: A randomized clinical trial. PLoS ONE 2017, 12, e0180067. [Google Scholar] [CrossRef]
- Jacka, F.N.; Overland, S.; Stewart, R.; Tell, G.S.; Bjelland, I.; Mykletun, A. Association between magnesium intake and depression and anxiety in community-dwelling adults: The Hordaland Health Study. Aust. N. Z. J. Psychiatry 2009, 43, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Yary, T.; Lehto, S.M.; Tolmunen, T.; Tuomainen, T.P.; Kauhanen, J.; Voutilainen, S.; Ruusunen, A. Dietary magnesium intake and the incidence of depression: A 20-year follow-up study. J. Affect. Disord. 2016, 193, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Guintivano, J.; Aberg, K.A.; Clark, S.L.; Rubinow, D.R.; Sullivan, P.F.; Meltzer-Brody, S.; van den Oord, E.J.C.G. Transcriptome-wide association study for postpartum depression implicates altered B-cell activation and insulin resistance. Mol. Psychiatry 2022, 27, 2858–2867. [Google Scholar] [CrossRef]
- Rasgon, N.L.; Kenna, H.A.; Williams, K.E.; Powers, B.; Wroolie, T.; Schatzberg, A.F. Rosiglitazone add-on in treatment of depressed patients with insulin resistance: A pilot study. Sci. World J. 2010, 10, 321–328. [Google Scholar] [CrossRef]
- Rasgon, N.L.; Kenna, H.A.; Reynolds-May, M.F.; Stemmle, P.G.; Vemuri, M.; Marsh, W.; Wang, P.; Ketter, T.A. Metabolic dysfunction in women with bipolar disorder: The potential influence of family history of type 2 diabetes mellitus. Bipolar Disord. 2010, 12, 504–513. [Google Scholar] [CrossRef]
- Sepanjnia, K.; Modabbernia, A.; Ashrafi, M.; Modabbernia, M.J.; Akhondzadeh, S. Pioglitazone adjunctive therapy for moderate-to-severe major depressive disorder: Randomized double-blind placebo-controlled trial. Neuropsychopharmacology 2012, 37, 2093–2100. [Google Scholar] [CrossRef]
- Kim, C. Maternal outcomes and follow-up after gestational diabetes mellitus. Diabet. Med. 2014, 31, 292–301. [Google Scholar] [CrossRef]
- Ruohomäki, A.; Toffol, E.; Upadhyaya, S.; Keski-Nisula, L.; Pekkanen, J.; Lampi, J.; Voutilainen, S.; Tuomainen, T.P.; Heinonen, S.; Kumpulainen, K.; et al. The association between gestational diabetes mellitus and postpartum depressive symptomatology: A prospective cohort study. J. Affect. Disord. 2018, 241, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Tokuda, N.; Kobayashi, Y.; Tanaka, H.; Sawai, H.; Shibahara, H.; Takeshima, Y.; Shima, M.; Group, J.E.a.C.s.S. Association between the serum insulin-like growth factor-1 concentration in the first trimester of pregnancy and postpartum depression. Psychiatry Clin. Neurosci. 2021, 75, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Fei, Y.; Li, J.; Shi, Y.; Yang, X. A Novel Review of Homocysteine and Pregnancy Complications. Biomed. Res. Int. 2021, 2021, 6652231. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.M.; Fernandez-Ballart, J.D. Homocysteine in pregnancy. Adv. Clin. Chem. 2011, 53, 105–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zou, W.; Huang, Y.; Wen, X.; Huang, J.; Wang, Y.; Sheng, X. A Preliminary Study of Uric Metabolomic Alteration for Postpartum Depression Based on Liquid Chromatography Coupled to Quadrupole Time-of-Flight Mass Spectrometry. Dis. Markers 2019, 2019, 4264803. [Google Scholar] [CrossRef]
- Liu, Y. Demonstrations of AIDS-associated malignancies and infections at FDG PET-CT. Ann. Nucl. Med. 2011, 25, 536–546. [Google Scholar] [CrossRef]
- McLean, A.; Rubinsztein, J.S.; Robbins, T.W.; Sahakian, B.J. The effects of tyrosine depletion in normal healthy volunteers: Implications for unipolar depression. Psychopharmacology 2004, 171, 286–297. [Google Scholar] [CrossRef]
- Doornbos, B.; Fekkes, D.; Tanke, M.A.; de Jonge, P.; Korf, J. Sequential serotonin and noradrenalin associated processes involved in postpartum blues. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1320–1325. [Google Scholar] [CrossRef]
- Mukta, F.Y.; Akhter, Q.S.; Islam, S.; Layla, K.N.; Azad, A.B.; Rahman, K.L.; Sarker, S.; Shahid, S.T.B. The evaluation of urinary vanillylmandelic acid level in patients with generalized anxiety disorder. World J. Adv. Res. Rev. 2021, 12, 001–005. [Google Scholar] [CrossRef]
- Wang, W.; Guo, H.; Zhang, S.X.; Li, J.; Cheng, K.; Bai, S.J.; Yang, D.Y.; Wang, H.Y.; Liang, Z.H.; Liao, L.; et al. Targeted Metabolomic Pathway Analysis and Validation Revealed Glutamatergic Disorder in the Prefrontal Cortex among the Chronic Social Defeat Stress Mice Model of Depression. J. Proteome Res. 2016, 15, 3784–3792. [Google Scholar] [CrossRef]
- Jiang, R.; Wu, S.; Fang, C.; Wang, C.; Yang, Y.; Liu, C.; Hu, J.; Huang, Y. Amino acids levels in early pregnancy predict subsequent gestational diabetes. J. Diabetes 2020, 12, 503–511. [Google Scholar] [CrossRef]
- Horrobin, D.F. Phospholipid metabolism and depression: The possible roles of phospholipase A2 and coenzyme A-independent transacylase. Hum. Psychopharmacol. 2001, 16, 45–52. [Google Scholar] [CrossRef]
- Innis, S.M. Essential fatty acid transfer and fetal development. Placenta 2005, 26 (Suppl. A), S70–S75. [Google Scholar] [CrossRef] [PubMed]
- Tahaei, H.; Gignac, F.; Pinar, A.; Fernandez-Barrés, S.; Romaguera, D.; Vioque, J.; Santa-Marina, L.; Subiza-Pérez, M.; Llop, S.; Soler-Blasco, R.; et al. Omega-3 Fatty Acid Intake during Pregnancy and Child Neuropsychological Development: A Multi-Centre Population-Based Birth Cohort Study in Spain. Nutrients 2022, 14, 518. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.P.; Reichel, M.; Mühle, C.; Rhein, C.; Gulbins, E.; Kornhuber, J. Brain membrane lipids in major depression and anxiety disorders. Biochim. Biophys. Acta 2015, 1851, 1052–1065. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Christophe, A.; Delanghe, J.; Altamura, C.; Neels, H.; Meltzer, H.Y. Lowered omega3 polyunsaturated fatty acids in serum phospholipids and cholesteryl esters of depressed patients. Psychiatry Res. 1999, 85, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.Y.; Huang, S.Y.; Su, K.P. A meta-analytic review of polyunsaturated fatty acid compositions in patients with depression. Biol. Psychiatry 2010, 68, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Wahab, R.J.; Jaddoe, V.W.V.; Mezzoiuso, A.G.; Gaillard, R. Maternal polyunsaturated fatty acid concentrations during pregnancy and childhood liver fat accumulation. Clin. Nutr. 2022, 41, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Hamazaki, K.; Matsumura, K.; Tsuchida, A.; Kasamatsu, H.; Tanaka, T.; Ito, M.; Inadera, H.; Group, J.E.a.C.s.S. Dietary intake of fish and n-3 polyunsaturated fatty acids and risk of postpartum depression: A nationwide longitudinal study—The Japan Environment and Children’s Study (JECS). Psychol. Med. 2020, 50, 2416–2424. [Google Scholar] [CrossRef]
- Llorente, A.M.; Jensen, C.L.; Voigt, R.G.; Fraley, J.K.; Berretta, M.C.; Heird, W.C. Effect of maternal docosahexaenoic acid supplementation on postpartum depression and information processing. Am. J. Obstet. Gynecol. 2003, 188, 1348–1353. [Google Scholar] [CrossRef]
- Freeman, M.P.; Davis, M.; Sinha, P.; Wisner, K.L.; Hibbeln, J.R.; Gelenberg, A.J. Omega-3 fatty acids and supportive psychotherapy for perinatal depression: A randomized placebo-controlled study. J. Affect. Disord. 2008, 110, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Marangell, L.B.; Martinez, J.M.; Zboyan, H.A.; Chong, H.; Puryear, L.J. Omega-3 fatty acids for the prevention of postpartum depression: Negative data from a preliminary, open-label pilot study. Depress. Anxiety 2004, 19, 20–23. [Google Scholar] [CrossRef]
- Parekh, A.; Smeeth, D.; Milner, Y.; Thure, S. The Role of Lipid Biomarkers in Major Depression. Healthcare 2017, 5, 5. [Google Scholar] [CrossRef]
- Ramachandran Pillai, R.; Wilson, A.B.; Premkumar, N.R.; Kattimani, S.; Sagili, H.; Rajendiran, S. Low serum levels of High-Density Lipoprotein cholesterol (HDL-c) as an indicator for the development of severe postpartum depressive symptoms. PLoS ONE 2018, 13, e0192811. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best. Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J. Oxygen, the Janus gas; its effects on human placental development and function. J. Anat. 2009, 215, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jin, H.; Qiu, Y.; Liu, Y.; Wen, L.; Fu, Y.; Qi, H.; Baker, P.N.; Tong, C. Reactive Oxygen Species are Essential for Placental Angiogenesis During Early Gestation. Oxid. Med. Cell Longev. 2022, 2022, 4290922. [Google Scholar] [CrossRef] [PubMed]
- Hubel, C.A.; Roberts, J.M.; Taylor, R.N.; Musci, T.J.; Rogers, G.M.; McLaughlin, M.K. Lipid peroxidation in pregnancy: New perspectives on preeclampsia. Am. J. Obstet. Gynecol. 1989, 161, 1025–1034. [Google Scholar] [CrossRef]
- Cranfield, L.M.; Gollan, J.L.; White, A.G.; Dormandy, T.L. Serum antioxidant activity in normal and abnormal subjects. Ann. Clin. Biochem. 1979, 16, 299–306. [Google Scholar] [CrossRef]
- Hansson, M.; Olsson, I.; Nauseef, W.M. Biosynthesis, processing, and sorting of human myeloperoxidase. Arch. Biochem. Biophys. 2006, 445, 214–224. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Biological reactivity and biomarkers of the neutrophil oxidant, hypochlorous acid. Toxicology 2002, 181–182, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Hammer, A.; Desoye, G.; Dohr, G.; Sattler, W.; Malle, E. Myeloperoxidase-dependent generation of hypochlorite-modified proteins in human placental tissues during normal pregnancy. Lab. Investig. 2001, 81, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Kindzelskii, A.L.; Clark, A.J.; Espinoza, J.; Maeda, N.; Aratani, Y.; Romero, R.; Petty, H.R. Myeloperoxidase accumulates at the neutrophil surface and enhances cell metabolism and oxidant release during pregnancy. Eur. J. Immunol. 2006, 36, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Vaccarino, V.; Brennan, M.L.; Miller, A.H.; Bremner, J.D.; Ritchie, J.C.; Lindau, F.; Veledar, E.; Su, S.; Murrah, N.V.; Jones, L.; et al. Association of major depressive disorder with serum myeloperoxidase and other markers of inflammation: A twin study. Biol. Psychiatry 2008, 64, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Boeldt, D.S.; Yi, F.X.; Bird, I.M. eNOS activation and NO function: Pregnancy adaptive programming of capacitative entry responses alters nitric oxide (NO) output in vascular endothelium—New insights into eNOS regulation through adaptive cell signaling. J. Endocrinol. 2011, 210, 243–258. [Google Scholar] [CrossRef]
- Hong, Y.C.; Lee, K.H.; Im, M.W.; Kim, Y.J.; Ha, E.H. Placental superoxide dismutase, genetic polymorphism, and neonatal birth weight. J. Prev. Med. Public. Health 2004, 37, 306–311. [Google Scholar] [PubMed]
- Scapagnini, G.; Davinelli, S.; Drago, F.; De Lorenzo, A.; Oriani, G. Antioxidants as antidepressants: Fact or fiction? CNS Drugs 2012, 26, 477–490. [Google Scholar] [CrossRef]
- Marcus, C.; Feizi, P.; Hogg, J.; Summerfield, H.; Castellani, R.; Sriwastava, S.; Marano, G.D. Imaging in Differentiating Cerebral Toxoplasmosis and Primary CNS Lymphoma with Special Focus on FDG PET/CT. AJR Am. J. Roentgenol. 2021, 216, 157–164. [Google Scholar] [CrossRef]
- Gupta, S.; Aziz, N.; Sekhon, L.; Agarwal, R.; Mansour, G.; Li, J.; Agarwal, A. Lipid peroxidation and antioxidant status in preeclampsia: A systematic review. Obstet. Gynecol. Surv. 2009, 64, 750–759. [Google Scholar] [CrossRef]
- Bilici, M.; Efe, H.; Köroğlu, M.A.; Uydu, H.A.; Bekaroğlu, M.; Değer, O. Antioxidative enzyme activities and lipid peroxidation in major depression: Alterations by antidepressant treatments. J. Affect. Disord. 2001, 64, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Gałecki, P.; Szemraj, J.; Bieńkiewicz, M.; Florkowski, A.; Gałecka, E. Lipid peroxidation and antioxidant protection in patients during acute depressive episodes and in remission after fluoxetine treatment. Pharmacol. Rep. 2009, 61, 436–447. [Google Scholar] [CrossRef]
- Kodydková, J.; Vávrová, L.; Zeman, M.; Jirák, R.; Macásek, J.; Stanková, B.; Tvrzická, E.; Zák, A. Antioxidative enzymes and increased oxidative stress in depressive women. Clin. Biochem. 2009, 42, 1368–1374. [Google Scholar] [CrossRef]
- Herken, H.; Gurel, A.; Selek, S.; Armutcu, F.; Ozen, M.E.; Bulut, M.; Kap, O.; Yumru, M.; Savas, H.A.; Akyol, O. Adenosine deaminase, nitric oxide, superoxide dismutase, and xanthine oxidase in patients with major depression: Impact of antidepressant treatment. Arch. Med. Res. 2007, 38, 247–252. [Google Scholar] [CrossRef]
- Stefanescu, C.; Ciobica, A. The relevance of oxidative stress status in first episode and recurrent depression. J. Affect. Disord. 2012, 143, 34–38. [Google Scholar] [CrossRef]
- Jiménez-Fernández, S.; Gurpegui, M.; Díaz-Atienza, F.; Pérez-Costillas, L.; Gerstenberg, M.; Correll, C.U. Oxidative stress and antioxidant parameters in patients with major depressive disorder compared to healthy controls before and after antidepressant treatment: Results from a meta-analysis. J. Clin. Psychiatry 2015, 76, 1658–1667. [Google Scholar] [CrossRef]
- Mazereeuw, G.; Herrmann, N.; Andreazza, A.C.; Khan, M.M.; Lanctôt, K.L. A meta-analysis of lipid peroxidation markers in major depression. Neuropsychiatr. Dis. Treat. 2015, 11, 2479–2491. [Google Scholar] [CrossRef]
- da Cruz, K.L.O.; Salla, D.H.; de Oliveira, M.P.; da Silva, L.E.; Dela Vedova, L.M.; Mendes, T.F.; Bressan, C.B.C.; Costa, A.B.; da Silva, M.R.; Réus, G.Z.; et al. The impact of obesity-related neuroinflammation on postpartum depression: A narrative review. Int. J. Dev. Neurosci. 2022, 82, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Masuya, M.; Matsumoto, T.; Ito, N.; Ohishi, K.; Maeda, M.; Katayama, N. High-intensity signals in the basal ganglia from gadolinium-enhanced T1-weighted MRI as an early change in toxoplasma encephalitis in an AIDS patient. J. Infect. Chemother. 2010, 16, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Roomruangwong, C.; Kanchanatawan, B.; Sirivichayakul, S.; Anderson, G.; Carvalho, A.F.; Duleu, S.; Geffard, M.; Maes, M. IgM-mediated autoimmune responses to oxidative specific epitopes, but not nitrosylated adducts, are significantly decreased in pregnancy: Association with bacterial translocation, perinatal and lifetime major depression and the tryptophan catabolite (TRYCAT) pathway. Metab. Brain Dis. 2017, 32, 1571–1583. [Google Scholar] [CrossRef] [PubMed]
- Roomruangwong, C.; Anderson, G.; Berk, M.; Stoyanov, D.; Carvalho, A.F.; Maes, M. A neuro-immune, neuro-oxidative and neuro-nitrosative model of prenatal and postpartum depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 81, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Ait Tayeb, A.E.K.; Poinsignon, V.; Chappell, K.; Bouligand, J.; Becquemont, L.; Verstuyft, C. Major Depressive Disorder and Oxidative Stress: A Review of Peripheral and Genetic Biomarkers According to Clinical Characteristics and Disease Stages. Antioxidants 2023, 12, 942. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D. The heterogeneity of “major depression”. World Psychiatry 2011, 10, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Pasco, J.A.; Nicholson, G.C.; Williams, L.J.; Jacka, F.N.; Henry, M.J.; Kotowicz, M.A.; Schneider, H.G.; Leonard, B.E.; Berk, M. Association of high-sensitivity C-reactive protein with de novo major depression. Br. J. Psychiatry 2010, 197, 372–377. [Google Scholar] [CrossRef]
- Mondin, T.C.; de Azevedo Cardoso, T.; Moreira, F.P.; Wiener, C.; Oses, J.P.; de Mattos Souza, L.D.; Jansen, K.; da Silva Magalhães, P.V.; Kapczinski, F.; da Silva, R.A. Circadian preferences, oxidative stress and inflammatory cytokines in bipolar disorder: A community study. J. Neuroimmunol. 2016, 301, 23–29. [Google Scholar] [CrossRef]
- Gadad, B.S.; Jha, M.K.; Czysz, A.; Furman, J.L.; Mayes, T.L.; Emslie, M.P.; Trivedi, M.H. Peripheral biomarkers of major depression and antidepressant treatment response: Current knowledge and future outlooks. J. Affect. Disord. 2018, 233, 3–14. [Google Scholar] [CrossRef]
- Ripke, S.; Wray, N.R.; Lewis, C.M.; Hamilton, S.P.; Weissman, M.M.; Breen, G.; Byrne, E.M.; Blackwood, D.H.; Boomsma, D.I.; Cichon, S.; et al. A mega-analysis of genome-wide association studies for major depressive disorder. Mol. Psychiatry 2013, 18, 497–511. [Google Scholar] [CrossRef]
- Couto, T.C.; Brancaglion, M.Y.; Alvim-Soares, A.; Moreira, L.; Garcia, F.D.; Nicolato, R.; Aguiar, R.A.; Leite, H.V.; Corrêa, H. Postpartum depression: A systematic review of the genetics involved. World J. Psychiatry 2015, 5, 103–111. [Google Scholar] [CrossRef]
- Zheng, Y.; Fan, W.; Zhang, X.; Dong, E. Gestational stress induces depressive-like and anxiety-like phenotypes through epigenetic regulation of BDNF expression in offspring hippocampus. Epigenetics 2016, 11, 150–162. [Google Scholar] [CrossRef]
- D’Addario, C.; Dell’Osso, B.; Galimberti, D.; Palazzo, M.C.; Benatti, B.; Di Francesco, A.; Scarpini, E.; Altamura, A.C.; Maccarrone, M. Epigenetic modulation of BDNF gene in patients with major depressive disorder. Biol. Psychiatry 2013, 73, e6–e7. [Google Scholar] [CrossRef] [PubMed]
- Comasco, E.; Sylvén, S.M.; Papadopoulos, F.C.; Sundström-Poromaa, I.; Oreland, L.; Skalkidou, A. Postpartum depression symptoms: A case-control study on monoaminergic functional polymorphisms and environmental stressors. Psychiatr. Genet. 2011, 21, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Alvim-Soares, A.; Miranda, D.; Campos, S.B.; Figueira, P.; Romano-Silva, M.A.; Correa, H. Postpartum depression symptoms associated with Val158Met COMT polymorphism. Arch. Womens Ment. Health 2013, 16, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Doornbos, B.; Dijck-Brouwer, D.A.; Kema, I.P.; Tanke, M.A.; van Goor, S.A.; Muskiet, F.A.; Korf, J. The development of peripartum depressive symptoms is associated with gene polymorphisms of MAOA, 5-HTT and COMT. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 1250–1254. [Google Scholar] [CrossRef] [PubMed]


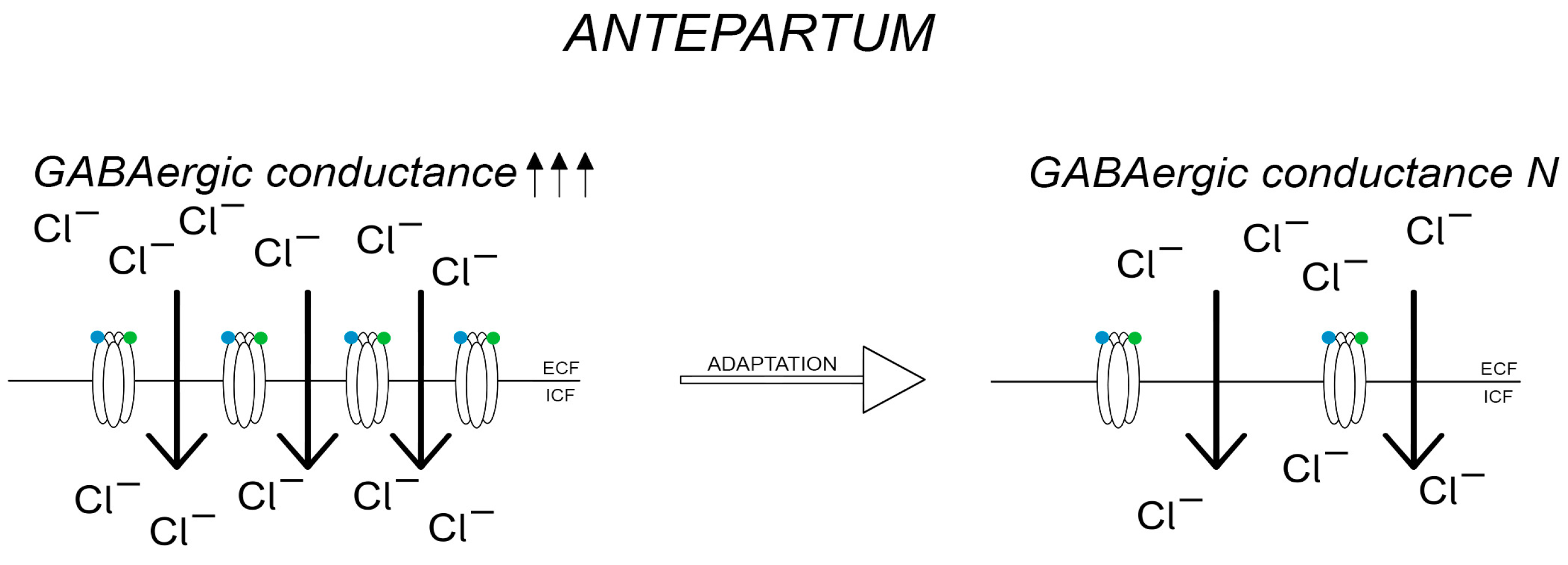
 —GABA,
—GABA,  —allopregnanolone.
—GABA, —allopregnanolone.
—allopregnanolone.
—GABA, —allopregnanolone.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormone | Concentration in PPD | |
|---|---|---|
| estrogen | ↓ | |
| progesterone | ||
| allopregnanolone | ||
| prolactin | ↓ | |
| oxytocin | ↑/↓ | |
| TSH | ↓/N/↑ | |
| HPA | CRH | ↓ |
| ACTH | ||
| cortisol | ↓/↑ | |
| Inflammation Biomarker | Concentration in PPD | |
|---|---|---|
| Chemokines | CX3CL1 | ↓ |
| CXCL8 | ↑ | |
| CCL2 | ↑ | |
| CCR2 | ↓ | |
| CCL4 | ↑ | |
| CCL5 | ↑ | |
| CCL11 | ↑ | |
| TNF-α | ↑ | |
| TGF-β1 | ↓ | |
| TGF-β2 | ↑ | |
| TGF-β3 | ↓ | |
| INF-γ | ↓ | |
| Interleukins | IL-1β | ↑ |
| IL-2 i IL-2R | ↑ | |
| IL-3 | ↑ | |
| IL-4 | ↓ | |
| IL-6 | ↑ | |
| IL-8 | ↑ | |
| IL-10 | ↓ | |
| IL-17 | ↑ | |
| IL-18 | ↑ | |
| BDNF | ↓ | |
| CRP | ↑ | |
| Nutrient | Concentration in PPD | |
|---|---|---|
| Vitamin A | ↓ | |
| Vitamin B6 | ||
| Vitamin B9 | ||
| Vitamin B12 | ||
| Vitamin C | ||
| Vitamin D | ||
| Vitamin E | ||
| Elements | Zinc | ↓ |
| Iron | ||
| Selenium | ||
| Magnesium | ||
| Metabolic Markers and Other Metabolic Substances | Concentration in PPD |
|---|---|
| Insulin | ↓ |
| Uric acid | ↓ |
| Homocysteine | ↓ |
| Tyrosine | ↓ |
| Vanillin-malic acid | ↓ |
| Alanine | ↓ |
| Lipid Marker | Concentration in PPD |
|---|---|
| PUFA | ↓ |
| HDL-c | ↓ |
| cholesterol—total | ↓ |
| Enzyme | Concentration in PPD |
|---|---|
| MPO | ↑ |
| NO and NOS | ↑ |
| MnSOD | ↑ |
| lipid peroxidase | ↑ |
| MDA | ↑ |
| Substance | Pregnancy—Postpartum Concentration | Possible Concentrations in PPD | |
|---|---|---|---|
| hormones | estrogen | ↑↑↑-N/↓ | N/↓↓ |
| progesterone | ↑↑-N/↓ | N/↓↓ | |
| allopregnanolone | ↑↑-↓ | ↓↓ | |
| cortisol | ↑-N/↑ | ↑/N/↓ | |
| inflammatory markers | IL-1β | ↑/↓-N | ↑↑↑ |
| IL-6 | ↑/↓-N | ↑↑↑ | |
| IL-10 | ↑-↓ | ↓↓ 1 | |
| TNF-α | ↑/↓-N | ↑↑ | |
| BDNF | ↓/N-↑ | ↓↓ | |
| CCL-2 | ↑-N | ↑↑ | |
| lipid markers | PUFA | ↑-↓ | ↓↓ |
| cholesterol | ↑-↑ | ↓↓ | |
| oxidative stress enzymes | MPO | ↑-↓ | ↑↑ |
| lipid peroxidase | ↑-↓ | ↑↑ | |
| nutrients | B vitamins | ↓-↓/N | ↓↓ |
| vitamin D | ↓-↓/N | ↓↓ | |
| metabolic markers | alanine | N | ↑↑ |
| vanillylmandelic acid | N | ↑↑ | |
| homocysteine | N | ↑↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modzelewski, S.; Oracz, A.; Iłendo, K.; Sokół, A.; Waszkiewicz, N. Biomarkers of Postpartum Depression: A Narrative Review. J. Clin. Med. 2023, 12, 6519. https://doi.org/10.3390/jcm12206519
Modzelewski S, Oracz A, Iłendo K, Sokół A, Waszkiewicz N. Biomarkers of Postpartum Depression: A Narrative Review. Journal of Clinical Medicine. 2023; 12(20):6519. https://doi.org/10.3390/jcm12206519
Chicago/Turabian StyleModzelewski, Stefan, Aleksandra Oracz, Kamila Iłendo, Aleksandra Sokół, and Napoleon Waszkiewicz. 2023. "Biomarkers of Postpartum Depression: A Narrative Review" Journal of Clinical Medicine 12, no. 20: 6519. https://doi.org/10.3390/jcm12206519
APA StyleModzelewski, S., Oracz, A., Iłendo, K., Sokół, A., & Waszkiewicz, N. (2023). Biomarkers of Postpartum Depression: A Narrative Review. Journal of Clinical Medicine, 12(20), 6519. https://doi.org/10.3390/jcm12206519