

Chronic Fatigue, Depression and Anxiety Symptoms in Long COVID Are Strongly Predicted by Neuroimmune and Neuro-Oxidative Pathways Which Are Caused by the Inflammation during Acute Infection

 ,
, 
 ,
,
Abstract
1. Introduction
2. Participants and Methods
2.1. Participants
2.2. Clinical Measurements
2.3. Assays
2.4. Statistical Analysis
3. Results
3.1. Socio-Demographic Data
3.2. Differences in Psychiatric Rating Scales between Study Groups
3.3. Differences in Biomarkers between Study Groups
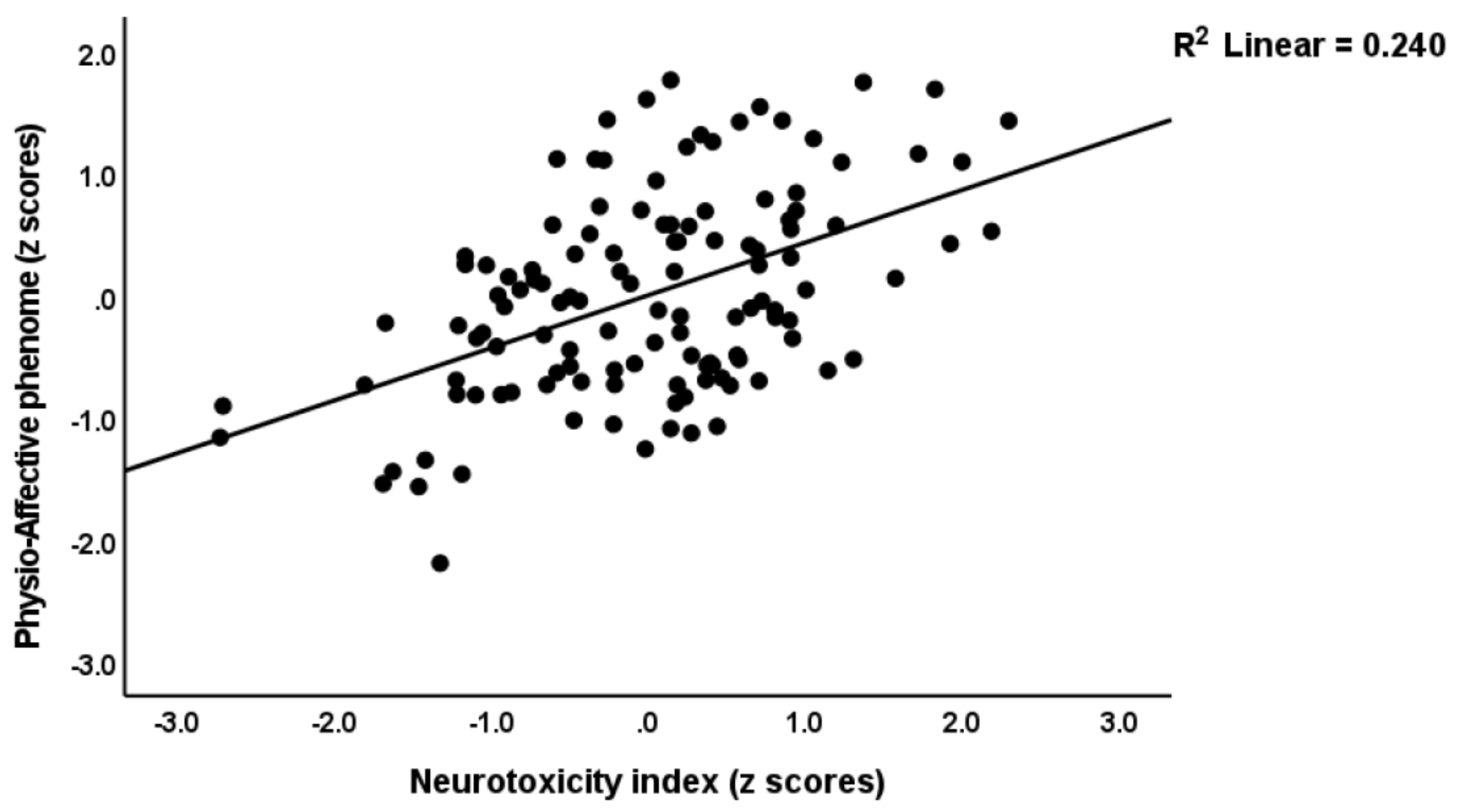
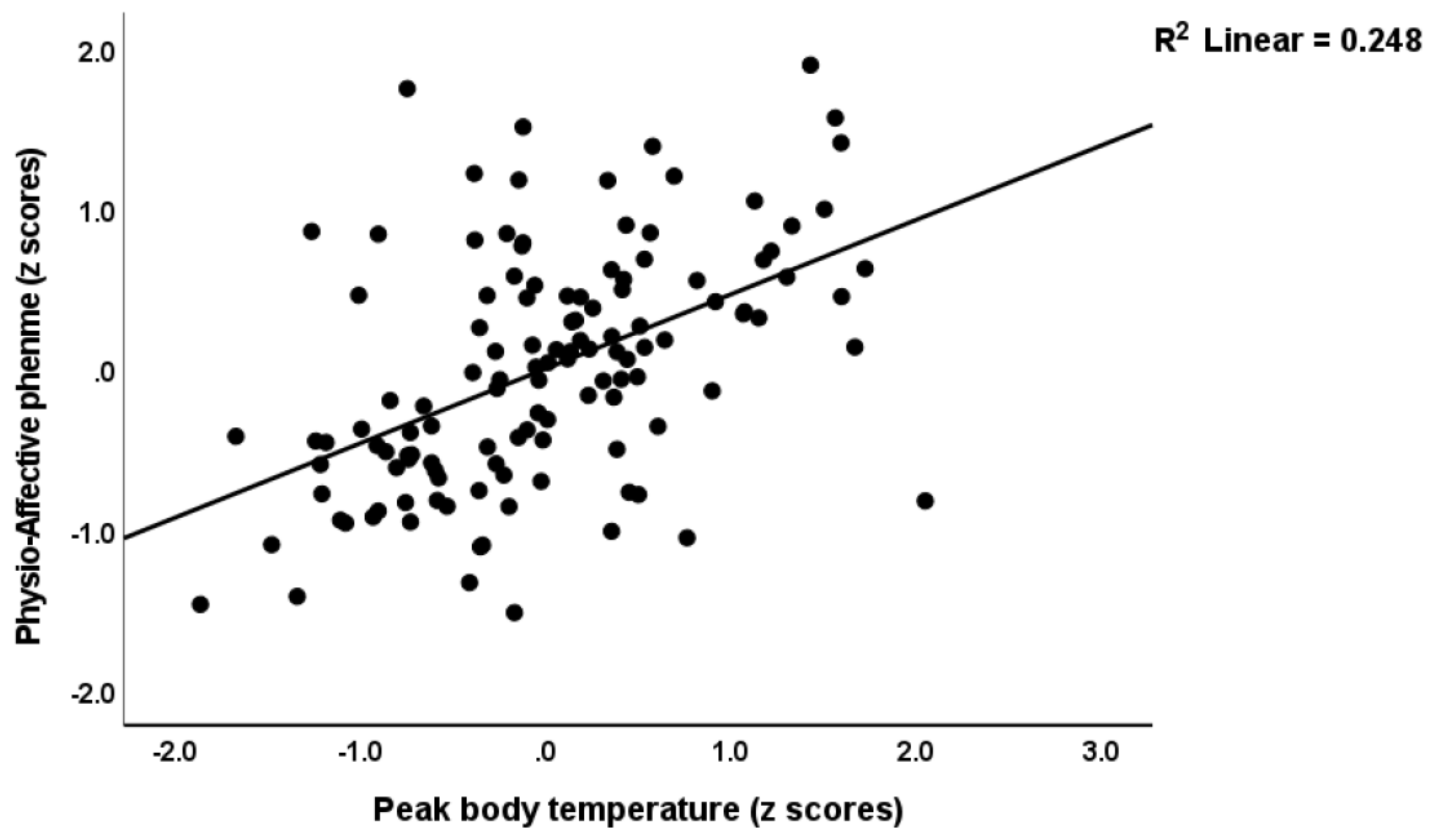
3.4. Prediction of the Physio-Affective Phenome Using TO2, NT, and Total Ca
3.5. Prediction of the Physio-Somatic and Affective Domains Using Biomarkers
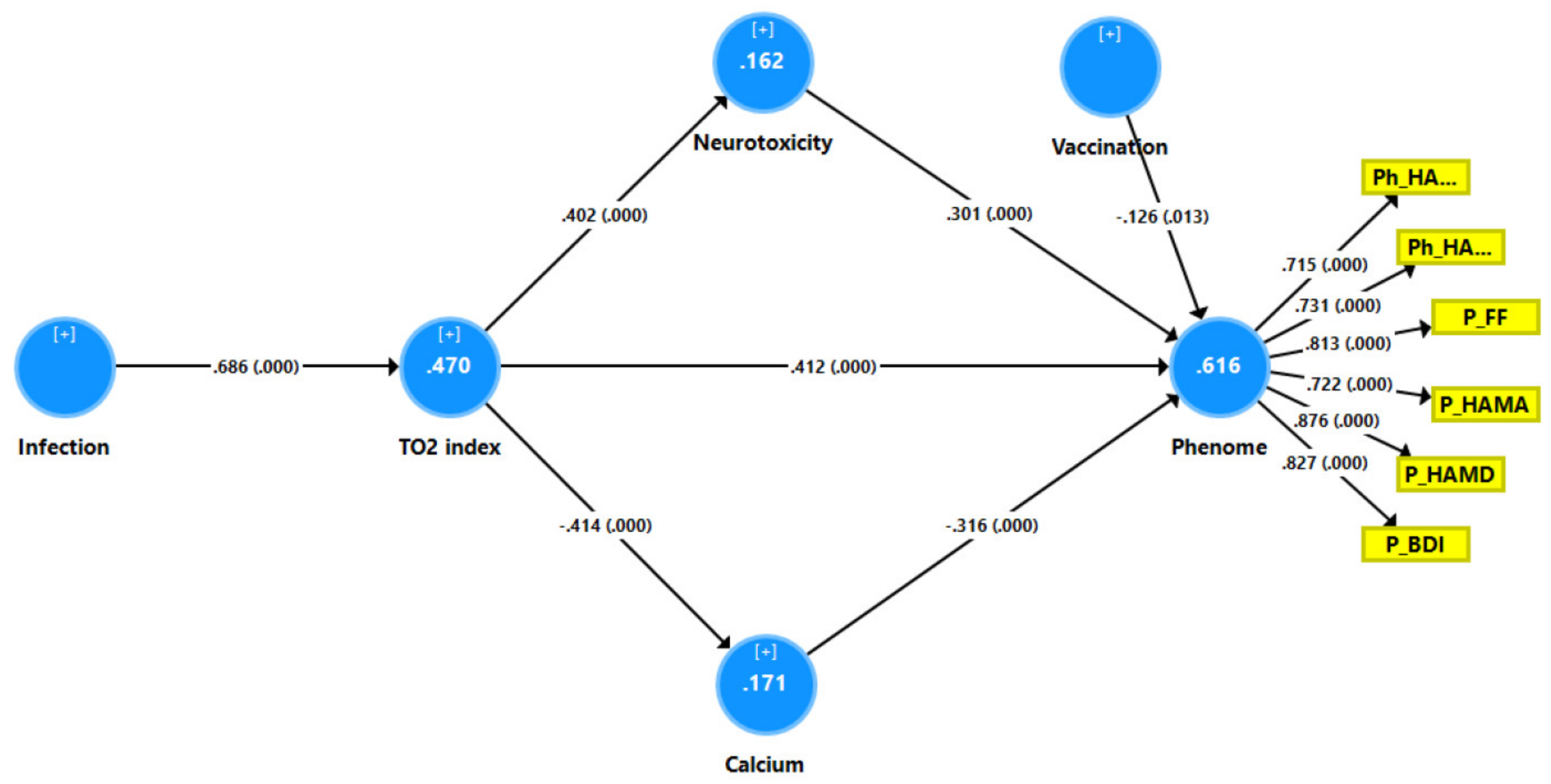
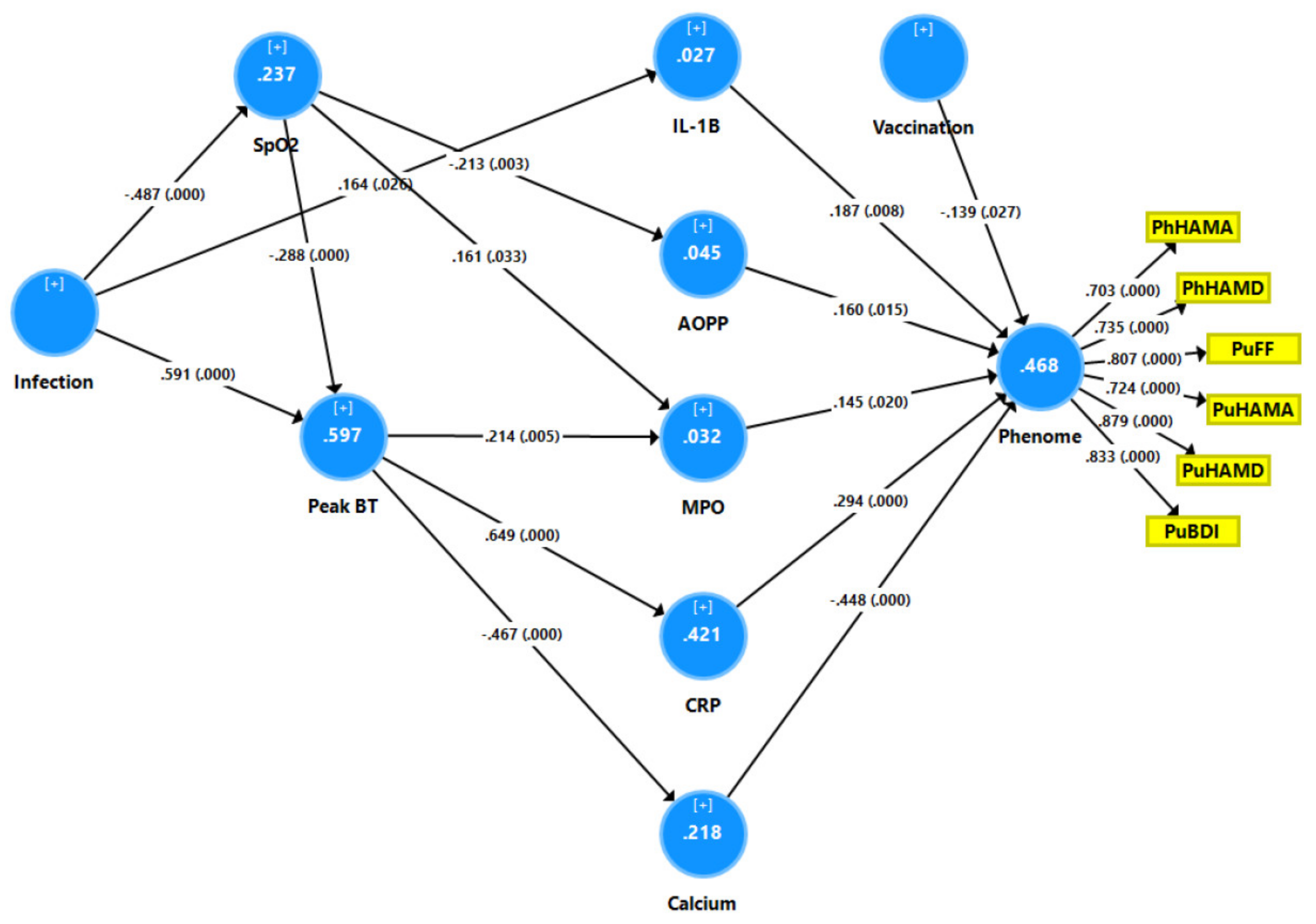
3.6. Results of PLS Analysis
4. Discussion
4.1. The Physio-Affective Phenome of Long COVID
4.2. Increased NT due to NLRP3 Activation Predicts the Physio-Affective Phenome
4.3. Increased NT due to Increased CRP Predicts the Physio-Affective Core
4.4. Increased NT due to Oxidative Stress Predicts the Physio-Affective Phenome
4.5. Lowered Total Ca Levels Predict the Physio-Affective Phenome
4.6. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Sachdeva, R.; Gower, C.; Ramsay, M.; Lopez Bernal, J. Effectiveness of COVID-19 booster vaccines against COVID-19-related symptoms, hospitalization and death in England. Nat. Med. 2022, 28, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Moghadas, S.M.; Vilches, T.N.; Zhang, K.; Wells, C.R.; Shoukat, A.; Singer, B.H.; Meyers, L.A.; Neuzil, K.M.; Langley, J.M.; Fitzpatrick, M.C.; et al. The Impact of Vaccination on Coronavirus Disease 2019 (COVID-19) Outbreaks in the United States. Clin. Infect. Dis. 2021, 73, 2257–2264. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.M.; Robinson, D.A.; Yang, L.; Williams, C.F.; Newman, L.M.; Breen, J.J.; Eisinger, R.W.; Schneider, J.S.; Adimora, A.A.; Erbelding, E.J. Toward Understanding COVID-19 Recovery: National Institutes of Health Workshop on Postacute COVID-19. Ann. Intern. Med. 2021, 174, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. More than 50 long-term effects of COVID-19: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 16144. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Kang, L.; Guo, L.; Liu, M.; Zhou, X.; et al. 6-month consequences of COVID-19 in patients discharged from hospital: A cohort study. Lancet 2021, 397, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhou, F.; Hou, W.; Silver, Z.; Wong, C.Y.; Chang, O.; Huang, E.; Zuo, Q.K. The prevalence of depression, anxiety, and sleep disturbances in COVID-19 patients: A meta-analysis. Ann. N. Y. Acad. Sci. 2021, 1486, 90–111. [Google Scholar] [CrossRef] [PubMed]
- Fernández-de-Las-Peñas, C.; Palacios-Ceña, D.; Gómez-Mayordomo, V.; Cuadrado, M.L.; Florencio, L.L. Defining Post-COVID Symptoms (Post-Acute COVID, Long COVID, Persistent Post-COVID): An Integrative Classification. Int. J. Environ. Res. Public Health 2021, 18, 2621. [Google Scholar] [CrossRef]
- Arnold, D.T.; Hamilton, F.W.; Milne, A.; Morley, A.J.; Viner, J.; Attwood, M.; Noel, A.; Gunning, S.; Hatrick, J.; Hamilton, S.; et al. Patient outcomes after hospitalisation with COVID-19 and implications for follow-up: Results from a prospective UK cohort. Thorax 2021, 76, 399–401. [Google Scholar] [CrossRef]
- Carfì, A.; Bernabei, R.; Landi, F. Persistent Symptoms in Patients After Acute COVID-19. JAMA 2020, 324, 603–605. [Google Scholar] [CrossRef]
- Renaud-Charest, O.; Lui, L.M.W.; Eskander, S.; Ceban, F.; Ho, R.; Di Vincenzo, J.D.; Rosenblat, J.D.; Lee, Y.; Subramaniapillai, M.; McIntyre, R.S. Onset and frequency of depression in post-COVID-19 syndrome: A systematic review. J. Psychiatr. Res. 2021, 144, 129–137. [Google Scholar] [CrossRef]
- Sandler, C.X.; Wyller, V.B.B.; Moss-Morris, R.; Buchwald, D.; Crawley, E.; Hautvast, J.; Katz, B.Z.; Knoop, H.; Little, P.; Taylor, R.; et al. Long COVID and Post-infective Fatigue Syndrome: A Review. Open Forum Infect. Dis. 2021, 8, ofab440. [Google Scholar] [CrossRef]
- Shah, W.; Hillman, T.; Playford, E.D.; Hishmeh, L. Managing the long term effects of COVID-19: Summary of NICE, SIGN, and RCGP rapid guideline. BMJ 2021, 372, n136. [Google Scholar] [CrossRef]
- Cares-Marambio, K.; Montenegro-Jiménez, Y.; Torres-Castro, R.; Vera-Uribe, R.; Torralba, Y.; Alsina-Restoy, X.; Vasconcello-Castillo, L.; Vilaró, J. Prevalence of potential respiratory symptoms in survivors of hospital admission after coronavirus disease 2019 (COVID-19): A systematic review and meta-analysis. Chron. Respir. Dis. 2021, 18, 14799731211002240. [Google Scholar] [CrossRef]
- Davis, H.E.; Assaf, G.S.; McCorkell, L.; Wei, H.; Low, R.J.; Re’em, Y.; Redfield, S.; Austin, J.P.; Akrami, A. Characterizing long COVID in an international cohort: 7 months of symptoms and their impact. eClinicalMedicine 2021, 38, 101019. [Google Scholar] [CrossRef]
- Simani, L.; Ramezani, M.; Darazam, I.A.; Sagharichi, M.; Aalipour, M.A.; Ghorbani, F.; Pakdaman, H. Prevalence and correlates of chronic fatigue syndrome and post-traumatic stress disorder after the outbreak of the COVID-19. J. Neurovirol. 2021, 27, 154–159. [Google Scholar] [CrossRef]
- Bellan, M.; Soddu, D.; Balbo, P.E.; Baricich, A.; Zeppegno, P.; Avanzi, G.C.; Baldon, G.; Bartolomei, G.; Battaglia, M.; Battistini, S.; et al. Respiratory and Psychophysical Sequelae Among Patients With COVID-19 Four Months After Hospital Discharge. JAMA Netw. Open 2021, 4, e2036142. [Google Scholar] [CrossRef]
- Cirulli, E.T.; Schiabor Barrett, K.M.; Riffle, S.; Bolze, A.; Neveux, I.; Dabe, S.; Grzymski, J.J.; Lu, J.T.; Washington, N.L. Long-term COVID-19 symptoms in a large unselected population. medRxiv 2010. [Google Scholar] [CrossRef]
- Garrigues, E.; Janvier, P.; Kherabi, Y.; Le Bot, A.; Hamon, A.; Gouze, H.; Doucet, L.; Berkani, S.; Oliosi, E.; Mallart, E.; et al. Post-discharge persistent symptoms and health-related quality of life after hospitalization for COVID-19. J. Infect. 2020, 81, e4–e6. [Google Scholar] [CrossRef]
- Van den Borst, B.; Peters, J.B.; Brink, M.; Schoon, Y.; Bleeker-Rovers, C.P.; Schers, H.; van Hees, H.W.H.; van Helvoort, H.; van den Boogaard, M.; van der Hoeven, H.; et al. Comprehensive Health Assessment 3 Months after Recovery from Acute Coronavirus Disease 2019 (COVID-19). Clin. Infect. Dis. 2021, 73, e1089–e1098. [Google Scholar] [CrossRef]
- Maes, M.; Tedesco Junior, W.L.D.; Lozovoy, M.A.B.; Mori, M.T.E.; Danelli, T.; Almeida, E.R.D.d.; Tejo, A.M.; Tano, Z.N.; Reiche, E.M.V.; Simão, A.N.C. In COVID-19, NLRP3 inflammasome genetic variants are associated with critical disease and these effects are partly mediated by the sickness symptom complex: A nomothetic network approach. Mol. Psychiatry 2022, 27, 1945–1955. [Google Scholar] [CrossRef]
- Yalcinkaya, M.; Liu, W.; Islam, M.N.; Kotini, A.G.; Gusarova, G.A.; Fidler, T.P.; Papapetrou, E.P.; Bhattacharya, J.; Wang, N.; Tall, A.R. Modulation of the NLRP3 inflammasome by SARS-CoV-2 Envelope protein. Sci. Rep. 2021, 11, 24432. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Di, B.; Xu, L.-L. The NLRP3 inflammasome and COVID-19: Activation, pathogenesis and therapeutic strategies. Cytokine Growth Factor Rev. 2021, 61, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Sagulkoo, P.; Plaimas, K.; Suratanee, A.; Colado Simão, A.N.; Vissoci Reiche, E.M.; Maes, M. Immunopathogenesis and Immunogenetic Variants in COVID-19. Curr. Pharm. Des. 2022, 28, 1780–1797. [Google Scholar] [CrossRef] [PubMed]
- Vora, S.M.; Lieberman, J.; Wu, H. Inflammasome activation at the crux of severe COVID-19. Nat. Rev. Immunol. 2021, 21, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, C.; Crespo, Ã.n.; Ranjbar, S.; Lewandrowski, M.; Ingber, J.; de Lacerda, L.B.; Parry, B.; Ravid, S.; Clark, S.; Ho, F.; et al. SARS-CoV-2 infects blood monocytes to activate NLRP3 and AIM2 inflammasomes, pyroptosis and cytokine release. Res. Sq. 2021, 1–42. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; de Sá, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Gonçalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218, e20201707. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.V.; Deng, M.; Ting, J.P. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Tharakan, S.; Nomoto, K.; Miyashita, S.; Ishikawa, K. Body temperature correlates with mortality in COVID-19 patients. Crit. Care 2020, 24, 298. [Google Scholar] [CrossRef]
- Al-Hadrawi, D.S.; Al-Rubaye, H.T.; Almulla, A.F.; Al-Hakeim, H.K.; Maes, M. Lowered oxygen saturation and increased body temperature in acute COVID-19 largely predict chronic fatigue syndrome and affective symptoms due to Long COVID: A precision nomothetic approach. Acta Neuropsychiatr. 2022, 1–12. [Google Scholar] [CrossRef]
- Maes, M. Precision Nomothetic Medicine in Depression Research: A New Depression Model, and New Endophenotype Classes and Pathway Phenotypes, and A Digital Self. J. Pers. Med. 2022, 12, 403. [Google Scholar] [CrossRef]
- Al-Jassas, H.K.; Al-Hakeim, H.K.; Maes, M. Intersections between pneumonia, lowered oxygen saturation percentage and immune activation mediate depression, anxiety, and chronic fatigue syndrome-like symptoms due to COVID-19: A nomothetic network approach. J. Affect. Disord. 2022, 297, 233–245. [Google Scholar] [CrossRef]
- Al-Hakeim, H.K.; Al-Rubaye, H.T.; Al-Hadrawi, D.S.; Almulla, A.F.; Maes, M. Long-COVID post-viral chronic fatigue and affective symptoms are associated with oxidative damage, lowered antioxidant defenses and inflammation: A proof of concept and mechanism study. Mol. Psychiatry 2022. [CrossRef]
- Maes, M.; Carvalho, A.F. The Compensatory Immune-Regulatory Reflex System (CIRS) in Depression and Bipolar Disorder. Mol. Neurobiol. 2018, 55, 8885–8903. [Google Scholar] [CrossRef]
- Maes, M.; Bonifacio, K.L.; Morelli, N.R.; Vargas, H.O.; Moreira, E.G.; St Stoyanov, D.; Barbosa, D.S.; Carvalho, A.F.; Nunes, S.O.V. Generalized Anxiety Disorder (GAD) and Comorbid Major Depression with GAD Are Characterized by Enhanced Nitro-oxidative Stress, Increased Lipid Peroxidation, and Lowered Lipid-Associated Antioxidant Defenses. Neurotox. Res. 2018, 34, 489–510. [Google Scholar] [CrossRef]
- Maes, M.; Kubera, M.; Kotańska, M. Aberrations in the Cross-Talks Among Redox, Nuclear Factor-κB, and Wnt/β-Catenin Pathway Signaling Underpin Myalgic Encephalomyelitis and Chronic Fatigue Syndrome. Front. Psychiatry 2022, 13, 822382. [Google Scholar] [CrossRef]
- Almulla, F.A.; Maes, M. The Tryptophan Catabolite or Kynurenine Pathway’s Role in Major Depression. Curr. Top. Med. Chem. 2022, 22, 1731–1735. [Google Scholar] [CrossRef]
- Zhou, X.; Fernando, S.M.; Pan, A.Y.; Laposa, R.; Cullen, K.R.; Klimes-Dougan, B.; Andreazza, A.C. Characterizing the NLRP3 Inflammasome in Mood Disorders: Overview, Technical Development, and Measures of Peripheral Activation in Adolescent Patients. Int. J. Mol. Sci. 2021, 22, 12513. [Google Scholar] [CrossRef]
- Kaufmann, F.N.; Costa, A.P.; Ghisleni, G.; Diaz, A.P.; Rodrigues, A.L.S.; Peluffo, H.; Kaster, M.P. NLRP3 inflammasome-driven pathways in depression: Clinical and preclinical findings. Brain Behav. Immun. 2017, 64, 367–383. [Google Scholar] [CrossRef]
- Zhang, Z.T.; Du, X.M.; Ma, X.J.; Zong, Y.; Chen, J.K.; Yu, C.L.; Liu, Y.G.; Chen, Y.C.; Zhao, L.J.; Lu, G.C. Activation of the NLRP3 inflammasome in lipopolysaccharide-induced mouse fatigue and its relevance to chronic fatigue syndrome. J. Neuroinflamm. 2016, 13, 71. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, X.; Xia, Z.; Chen, J.; Liu, Y.; Chen, Y.; Zhu, J.; Li, J.; Yu, H.; Zong, Y.; et al. NLRP3 inflammasome activation mediates fatigue-like behaviors in mice via neuroinflammation. Neuroscience 2017, 358, 115–123. [Google Scholar] [CrossRef]
- Kuwar, R.; Rolfe, A.; Di, L.; Blevins, H.; Xu, Y.; Sun, X.; Bloom, G.S.; Zhang, S.; Sun, D. A Novel Inhibitor Targeting NLRP3 Inflammasome Reduces Neuropathology and Improves Cognitive Function in Alzheimer’s Disease Transgenic Mice. J. Alzheimers Dis. 2021, 82, 1769–1783. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Trageser, K.J.; Wu, H.; Herman, F.J.; Iqbal, U.H.; Sebastian-Valverde, M.; Frolinger, T.; Zeng, E.; Pasinetti, G.M. Anxiolytic effects of NLRP3 inflammasome inhibition in a model of chronic sleep deprivation. Transl. Psychiatry 2021, 11, 52. [Google Scholar] [CrossRef] [PubMed]
- Aghaie, F.; Moradifar, F.; Hosseini, A. Rapamycin attenuates depression and anxiety-like behaviors through modulation of the NLRP3 pathway in pentylenetetrazole-kindled male Wistar rats. Fundam. Clin. Pharmacol. 2021, 35, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.H.; Stoyanov, D. False dogmas in mood disorders research: Towards a nomothetic network approach. World J. Psychiatry 2022, 12, 651–667. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. A Clinical Case Definition of Post COVID-19 Condition by a Delphi Consensus, 6 October 2021; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Morris, G.; Maes, M. Myalgic encephalomyelitis/chronic fatigue syndrome and encephalomyelitis disseminata/multiple sclerosis show remarkable levels of similarity in phenomenology and neuroimmune characteristics. BMC Med. 2013, 11, 205. [Google Scholar] [CrossRef]
- Zachrisson, O.; Regland, B.; Jahreskog, M.; Kron, M.; Gottfries, C.G. A rating scale for fibromyalgia and chronic fatigue syndrome (the FibroFatigue scale). J. Psychosom. Res. 2002, 52, 501–509. [Google Scholar] [CrossRef]
- Hamilton, M. A rating scale for depression. J. Neurol. Neurosurg Psychiatry 1960, 23, 56–62. [Google Scholar] [CrossRef]
- Beck, A.T.; Steer, R.A.; Brown, G. Beck Depression Inventory–II (BDI-II) [Database Record]. In APA PsycTests; American Psychological Association: Washington, DC, USA; p. 1996. [CrossRef]
- Hamilton, M. The assessment of anxiety states by rating. Br. J. Med. Psychol. 1959, 32, 50–55. [Google Scholar] [CrossRef]
- Lin, L.; Xu, L.; Lv, W.; Han, L.; Xiang, Y.; Fu, L.; Jin, M.; Zhou, R.; Chen, H.; Zhang, A. An NLRP3 inflammasome-triggered cytokine storm contributes to Streptococcal toxic shock-like syndrome (STSLS). PLoS Pathog. 2019, 15, e1007795. [Google Scholar] [CrossRef]
- Tisoncik Jennifer, R.; Korth Marcus, J.; Simmons Cameron, P.; Farrar, J.; Martin Thomas, R.; Katze Michael, G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef]
- Li, M.X.; Zheng, H.L.; Luo, Y.; He, J.G.; Wang, W.; Han, J.; Zhang, L.; Wang, X.; Ni, L.; Zhou, H.Y.; et al. Gene deficiency and pharmacological inhibition of caspase-1 confers resilience to chronic social defeat stress via regulating the stability of surface AMPARs. Mol. Psychiatry 2018, 23, 556–568. [Google Scholar] [CrossRef]
- Towers, A.E.; Oelschlager, M.L.; Patel, J.; Gainey, S.J.; McCusker, R.H.; Freund, G.G. Acute fasting inhibits central caspase-1 activity reducing anxiety-like behavior and increasing novel object and object location recognition. Metabolism 2017, 71, 70–82. [Google Scholar] [CrossRef]
- Swartz, J.R.; Prather, A.A.; Di Iorio, C.R.; Bogdan, R.; Hariri, A.R. A Functional Interleukin-18 Haplotype Predicts Depression and Anxiety through Increased Threat-Related Amygdala Reactivity in Women but Not Men. Neuropsychopharmacology 2017, 42, 419–426. [Google Scholar] [CrossRef]
- Hardcastle, S.L.; Brenu, E.W.; Johnston, S.; Nguyen, T.; Huth, T.; Ramos, S.; Staines, D.; Marshall-Gradisnik, S. Serum Immune Proteins in Moderate and Severe Chronic Fatigue Syndrome/Myalgic Encephalomyelitis Patients. Int. J. Med. Sci. 2015, 12, 764–772. [Google Scholar] [CrossRef]
- VanElzakker, M.B.; Brumfield, S.A.; Lara Mejia, P.S. Neuroinflammation and Cytokines in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): A Critical Review of Research Methods. Front. Neurol. 2019, 9, 1033. [Google Scholar] [CrossRef]
- Song, L.; Pei, L.; Yao, S.; Wu, Y.; Shang, Y. NLRP3 Inflammasome in Neurological Diseases, from Functions to Therapies. Front. Cell. Neurosci. 2017, 11, 63. [Google Scholar] [CrossRef]
- Cannon, J.G. Inflammatory Cytokines in Nonpathological States. News Physiol Sci. 2000, 15, 298–303. [Google Scholar] [CrossRef]
- Alvarez, J.I.; Dodelet-Devillers, A.; Kebir, H.; Ifergan, I.; Fabre, P.J.; Terouz, S.; Sabbagh, M.; Wosik, K.; Bourbonnière, L.; Bernard, M.; et al. The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence. Science 2011, 334, 1727–1731. [Google Scholar] [CrossRef]
- Ferrari, C.C.; Depino, A.M.; Prada, F.; Muraro, N.; Campbell, S.; Podhajcer, O.; Perry, V.H.; Anthony, D.C.; Pitossi, F.J. Reversible demyelination, blood-brain barrier breakdown, and pronounced neutrophil recruitment induced by chronic IL-1 expression in the brain. Am. J. Pathol. 2004, 165, 1827–1837. [Google Scholar] [CrossRef]
- Nakahira, M.; Ahn, H.J.; Park, W.R.; Gao, P.; Tomura, M.; Park, C.S.; Hamaoka, T.; Ohta, T.; Kurimoto, M.; Fujiwara, H. Synergy of IL-12 and IL-18 for IFN-gamma gene expression: IL-12-induced STAT4 contributes to IFN-gamma promoter activation by up-regulating the binding activity of IL-18-induced activator protein 1. J. Immunol. 2002, 168, 1146–1153. [Google Scholar] [CrossRef]
- Bossù, P.; Ciaramella, A.; Salani, F.; Vanni, D.; Palladino, I.; Caltagirone, C.; Scapigliati, G. Interleukin-18, from neuroinflammation to Alzheimer’s disease. Curr. Pharm. Des. 2010, 16, 4213–4224. [Google Scholar] [CrossRef] [PubMed]
- Felderhoff-Mueser, U.; Schmidt, O.I.; Oberholzer, A.; Bührer, C.; Stahel, P.F. IL-18: A key player in neuroinflammation and neurodegeneration? Trends Neurosci. 2005, 28, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Licastro, F.; Pedrini, S.; Caputo, L.; Annoni, G.; Davis, L.J.; Ferri, C.; Casadei, V.; Grimaldi, L.M. Increased plasma levels of interleukin-1, interleukin-6 and alpha-1-antichymotrypsin in patients with Alzheimer’s disease: Peripheral inflammation or signals from the brain? J. Neuroimmunol. 2000, 103, 97–102. [Google Scholar] [CrossRef] [PubMed]
- De Jong, B.A.; Huizinga, T.W.; Bollen, E.L.; Uitdehaag, B.M.; Bosma, G.P.; van Buchem, M.A.; Remarque, E.J.; Burgmans, A.C.; Kalkers, N.F.; Polman, C.H.; et al. Production of IL-1beta and IL-1Ra as risk factors for susceptibility and progression of relapse-onset multiple sclerosis. J. Neuroimmunol. 2002, 126, 172–179. [Google Scholar] [CrossRef]
- Huang, W.X.; Huang, P.; Hillert, J. Increased expression of caspase-1 and interleukin-18 in peripheral blood mononuclear cells in patients with multiple sclerosis. Mult. Scler. J. 2004, 10, 482–487. [Google Scholar] [CrossRef]
- Mandal, S.; Barnett, J.; Brill, S.E.; Brown, J.S.; Denneny, E.K.; Hare, S.S.; Heightman, M.; Hillman, T.E.; Jacob, J.; Jarvis, H.C.; et al. ‘Long-COVID’: A cross-sectional study of persisting symptoms, biomarker and imaging abnormalities following hospitalisation for COVID-19. Thorax 2021, 76, 396. [Google Scholar] [CrossRef]
- Gameil, M.A.; Marzouk, R.E.; Elsebaie, A.H.; Rozaik, S.E. Long-term clinical and biochemical residue after COVID-19 recovery. Egypt. Liver J. 2021, 11, 74. [Google Scholar] [CrossRef]
- Kuta, A.E.; Baum, L.L. C-reactive protein is produced by a small number of normal human peripheral blood lymphocytes. J. Exp. Med. 1986, 164, 321–326. [Google Scholar] [CrossRef]
- Song, I.U.; Kim, J.S.; Chung, S.W.; Lee, K.S. Is there an association between the level of high-sensitivity C-reactive protein and idiopathic Parkinson’s disease? A comparison of Parkinson’s disease patients, disease controls and healthy individuals. Eur. Neurol. 2009, 62, 99–104. [Google Scholar] [CrossRef]
- Windgassen, E.B.; Funtowicz, L.; Lunsford, T.N.; Harris, L.A.; Mulvagh, S.L. C-reactive protein and high-sensitivity C-reactive protein: An update for clinicians. Postgrad. Med. 2011, 123, 114–119. [Google Scholar] [CrossRef]
- Hsuchou, H.; Kastin, A.J.; Mishra, P.K.; Pan, W. C-reactive protein increases BBB permeability: Implications for obesity and neuroinflammation. Cell. Physiol. Biochem. 2012, 30, 1109–1119. [Google Scholar] [CrossRef]
- Belin, C.; Devic, P.; Ayrignac, X.; Dos Santos, A.; Paix, A.; Sirven-Villaros, L.; Simard, C.; Lamure, S.; Gastinne, T.; Ursu, R.; et al. Description of neurotoxicity in a series of patients treated with CAR T-cell therapy. Sci. Rep. 2020, 10, 18997. [Google Scholar] [CrossRef]
- Ge, X.; Xu, X.-y.; Feng, C.-h.; Wang, Y.; Li, Y.-l.; Feng, B. Relationships among serum C-reactive protein, receptor for advanced glycation products, metabolic dysfunction, and cognitive impairments. BMC Neurol. 2013, 13, 110. [Google Scholar] [CrossRef]
- Elkind, M.S.; Tai, W.; Coates, K.; Paik, M.C.; Sacco, R.L. High-sensitivity C-reactive protein, lipoprotein-associated phospholipase A2, and outcome after ischemic stroke. Arch Intern. Med. 2006, 166, 2073–2080. [Google Scholar] [CrossRef]
- Di Napoli, M.; Papa, F.; Bocola, V. C-reactive protein in ischemic stroke: An independent prognostic factor. Stroke 2001, 32, 917–924. [Google Scholar] [CrossRef]
- Masotti, L.; Ceccarelli, E.; Forconi, S.; Cappelli, R. Prognostic role of C-reactive protein in very old patients with acute ischaemic stroke. J. Intern. Med. 2005, 258, 145–152. [Google Scholar] [CrossRef]
- Montaner, J.; Fernandez-Cadenas, I.; Molina, C.A.; Ribó, M.; Huertas, R.; Rosell, A.; Penalba, A.; Ortega, L.; Chacón, P.; Alvarez-Sabín, J. Poststroke C-reactive protein is a powerful prognostic tool among candidates for thrombolysis. Stroke 2006, 37, 1205–1210. [Google Scholar] [CrossRef]
- Howren, M.B.; Lamkin, D.M.; Suls, J. Associations of depression with C-reactive protein, IL-1, and IL-6: A meta-analysis. Psychosom. Med. 2009, 71, 171–186. [Google Scholar] [CrossRef]
- Roomruangwong, C.; Kanchanatawan, B.; Sirivichayakul, S.; Mahieu, B.; Nowak, G.; Maes, M. Lower Serum Zinc and Higher CRP Strongly Predict Prenatal Depression and Physio-somatic Symptoms, Which All Together Predict Postnatal Depressive Symptoms. Mol. Neurobiol. 2017, 54, 1500–1512. [Google Scholar] [CrossRef]
- Vasupanrajit, A.; Jirakran, K.; Tunvirachaisakul, C.; Solmi, M.; Maes, M. Inflammation and nitro-oxidative stress in current suicidal attempts and current suicidal ideation: A systematic review and meta-analysis. Mol. Psychiatry 2022, 27, 1350–1361. [Google Scholar] [CrossRef]
- Costello, H.; Gould, R.L.; Abrol, E.; Howard, R. Systematic review and meta-analysis of the association between peripheral inflammatory cytokines and generalised anxiety disorder. BMJ Open 2019, 9, e027925. [Google Scholar] [CrossRef] [PubMed]
- Strawbridge, R.; Sartor, M.L.; Scott, F.; Cleare, A.J. Inflammatory proteins are altered in chronic fatigue syndrome-A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2019, 107, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Arce Rentería, M.; Gillett, S.R.; McClure, L.A.; Wadley, V.G.; Glasser, S.P.; Howard, V.J.; Kissela, B.M.; Unverzagt, F.W.; Jenny, N.S.; Manly, J.J.; et al. C-reactive protein and risk of cognitive decline: The REGARDS study. PLoS ONE 2020, 15, e0244612. [Google Scholar] [CrossRef] [PubMed]
- Mathy-Hartert, M.; Bourgeois, E.; Grülke, S.; Deby-Dupont, G.; Caudron, I.; Deby, C.; Lamy, M.; Serteyn, D. Purification of myeloperoxidase from equine polymorphonuclear leucocytes. Can. J. Vet. Res. 1998, 62, 127–132. [Google Scholar] [PubMed]
- Da Cruz Nizer, W.S.; Inkovskiy, V.; Overhage, J. Surviving Reactive Chlorine Stress: Responses of Gram-Negative Bacteria to Hypochlorous Acid. Microorganisms 2020, 8, 1220. [Google Scholar] [CrossRef]
- Maes, M.; Landucci Bonifacio, K.; Morelli, N.R.; Vargas, H.O.; Barbosa, D.S.; Carvalho, A.F.; Nunes, S.O.V. Major Differences in Neurooxidative and Neuronitrosative Stress Pathways Between Major Depressive Disorder and Types I and II Bipolar Disorder. Mol. Neurobiol. 2019, 56, 141–156. [Google Scholar] [CrossRef]
- Gray, E.; Thomas, T.L.; Betmouni, S.; Scolding, N.; Love, S. Elevated Activity and Microglial Expression of Myeloperoxidase in Demyelinated Cerebral Cortex in Multiple Sclerosis. Brain Pathol. 2008, 18, 86–95. [Google Scholar] [CrossRef]
- Green, P.S.; Mendez, A.J.; Jacob, J.S.; Crowley, J.R.; Growdon, W.; Hyman, B.T.; Heinecke, J.W. Neuronal expression of myeloperoxidase is increased in Alzheimer’s disease. J. Neurochem. 2004, 90, 724–733. [Google Scholar] [CrossRef]
- Vaccarino, V.; Brennan, M.-L.; Miller, A.H.; Bremner, J.D.; Ritchie, J.C.; Lindau, F.; Veledar, E.; Su, S.; Murrah, N.V.; Jones, L.; et al. Association of major depressive disorder with serum myeloperoxidase and other markers of inflammation: A twin study. Biol. Psychiatry 2008, 64, 476–483. [Google Scholar] [CrossRef]
- Liang, S.; Li, X.; Huang, W.; Gong, H. Change of serum myeloperoxidase and lipoxin A4 level in coronary heart disease patients with anxiety and/or depression. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2013, 38, 370–375. [Google Scholar] [CrossRef]
- Talarowska, M.; Szemraj, J.; Gałecki, P. Myeloperoxidase gene expression and cognitive functions in depression. Adv. Med. Sci. 2015, 60, 1–5. [Google Scholar] [CrossRef]
- Shrivastava, S.; Chelluboina, S.; Jedge, P.; Doke, P.; Palkar, S.; Mishra, A.C.; Arankalle, V.A. Elevated Levels of Neutrophil Activated Proteins, Alpha-Defensins (DEFA1), Calprotectin (S100A8/A9) and Myeloperoxidase (MPO) Are Associated with Disease Severity in COVID-19 Patients. Front. Cell. Infect. Microbiol. 2021, 11, 1056. [Google Scholar] [CrossRef]
- Islam, M.R.; Islam, M.R.; Shalahuddin Qusar, M.M.A.; Islam, M.S.; Kabir, M.H.; Mustafizur Rahman, G.K.M.; Islam, M.S.; Hasnat, A. Alterations of serum macro-minerals and trace elements are associated with major depressive disorder: A case-control study. BMC Psychiatry 2018, 18, 94. [Google Scholar] [CrossRef]
- Bowden, C.L.; Huang, L.G.; Javors, M.A.; Johnson, J.M.; Seleshi, E.; McIntyre, K.; Contreras, S.; Maas, J.W. Calcium function in affective disorders and healthy controls. Biol. Psychiatry 1988, 23, 367–376. [Google Scholar] [CrossRef]
- Al-Dujaili, A.H.; Al-Hakeim, H.K.; Twayej, A.J.; Maes, M. Total and ionized calcium and magnesium are significantly lowered in drug-naïve depressed patients: Effects of antidepressants and associations with immune activation. Metab. Brain Dis. 2019, 34, 1493–1503. [Google Scholar] [CrossRef]
- Jung, K.I.; Ock, S.M.; Chung, J.H.; Song, C.H. Associations of serum Ca and Mg levels with mental health in adult women without psychiatric disorders. Biol. Trace Elem. Res. 2010, 133, 153–161. [Google Scholar] [CrossRef]
- Paul, I.A. Antidepressant activity and calcium signaling cascades. Hum. Psychopharmacol. 2001, 16, 71–80. [Google Scholar] [CrossRef]
- Toescu, E.C.; Verkhratsky, A. The importance of being subtle: Small changes in calcium homeostasis control cognitive decline in normal aging. Aging Cell 2007, 6, 267–273. [Google Scholar] [CrossRef]
- Hurst, K. Primary hyperparathyroidism as a secondary cause of depression. J. Am. Board Fam. Med. 2010, 23, 677–680. [Google Scholar] [CrossRef]
- Grützner, T.M.; Listunova, L.; Fabian, G.A.; Kramer, B.A.; Flach, D.; Weisbrod, M.; Roesch-Ely, D.; Sharma, A. Serum calcium levels and neuropsychological performance in depression and matched healthy controls: Reversal of correlation a marker of the aging cognitive clock? Psychoneuroendocrinology 2018, 91, 198–205. [Google Scholar] [CrossRef]
- Almulla, A.F.; Thipakorn, Y.; Vasupanrajit, A.; Abo Algon, A.A.; Tunvirachaisakul, C.; Hashim Aljanabi, A.A.; Ox-enkrug, G.; Al-Hakeim, H.K.; Maes, M. The tryptophan catabolite or kynurenine pathway in major depressive and bi-polar disorder: A systematic review and meta-analysis. Brain Behav. Immun. Health 2022, 26, 100537. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Leonard, B.E.; Myint, A.M.; Kubera, M.; Verkerk, R. The new ‘5-HT’ hypothesis of depression: Cell-mediated immune activation induces indoleamine 2,3-dioxygenase, which leads to lower plasma tryptophan and an increased synthesis of detrimental tryptophan catabolites (TRYCATs), both of which contribute to the onset of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 702–721. [Google Scholar] [CrossRef]
- Maes, M.; Song, C.; Lin, A.H.; Bonaccorso, S.; Kenis, G.; De Jongh, R.; Bosmans, E.; Scharpé, S. Negative immunoregulatory effects of antidepressants: Inhibition of interferon-gamma and stimulation of interleukin-10 secretion. Neuropsychopharmacology 1999, 20, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Seneff, S.; Nigh, G.; Kyriakopoulos, A.M.; McCullough, P.A. Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food Chem. Toxicol. 2022, 164, 113008. [Google Scholar] [CrossRef]
- Couzin-Frankel, J.; Vogel, G. Vaccines may cause rare, Long COVID-like symptoms. Science 2022, 375, 364–366. [Google Scholar] [CrossRef]
- Spinicci, M. Different SARS-CoV-2 variants may give rise to different long COVID symptoms, study suggests. In Proceedings of the European Congress of Clinical Microbiology & Infectious Diseases (ECCMID 2022), Lisbon, Portugal, 23–26 April 2022. [Google Scholar]
- Al-Rashedi, N.A.M.; Alburkat, H.; Hadi, A.O.; Munahi, M.G.; Jasim, A.; Hameed, A.; Oda, B.S.; Lilo, K.M.; AlObaidi, L.A.H.; Vapalahti, O.; et al. High prevalence of an alpha variant lineage with a premature stop codon in ORF7a in Iraq, winter 2020–2021. PLoS ONE 2022, 17, e0267295. [Google Scholar] [CrossRef]
- Huaxia. Iraq Reports First Cases of COVID-19 Omicron Variant. Available online: https://english.news.cn/20220106/e6eedae158c44eea8433caf983a10738/c.html (accessed on 13 October 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | HC (n = 39) A | Low TO-NT (n = 56) B | High TO-NT (n = 30) C | F/X2 | df | p |
|---|---|---|---|---|---|---|
| Age (years) | 28.3 (7.6) | 27.6 (5.4) | 29.8 (7.3) | 1.15 | 2/122 | 0.320 |
| Sex (M/F) | 42/15 | 40/16 | 22/8 | 1.42 | 2 | 0.512 |
| Marital state (Ma/S) | 22/17 | 31/25 | 18/12 | 0.18 | 2 | 0.946 |
| Smoking (Y/N) | 13/26 | 16/40 | 11/19 | 0.64 | 2 | 0.732 |
| Residency (U/R) | 31/8 | 46/10 | 25/5 | 0.19 | 2 | 0.914 |
| Vaccination (A/Pf/S) | 9/21/9 | 14/31/11 | 6/17/7 | 0.42 | 4 | 0.979 |
| BMI (Kg/m2) | 25.60 (3.97) | 25.71 (3.74) | 26.96 (5.75) | 0.99 | 2/122 | 0.372 |
| Education (years) | 15.0 (1.3) | 15.7 (1.8) | 15.5 (1.7) | 2.73 | 2/122 | 0.069 |
| Peak BT (℃) | 36.86 (0.25) B,C | 38.20 (0.64) A,C | 39.31 (0.83) A,B | 138.38 | 2/122 | <0.001 |
| Lowest SpO2 (%) | 95.08 (1.52) B,C | 92.41 (2.74) A,C | 88.27 (4.62) A,B | 42.81 | 2/122 | <0.001 |
| TO2 index (z scores) | −1.01 (0.240) B,C | 0.054 (0.52) A,C | 1.21 (0.86) A,B | 131.43 | 2/122 | <0.001 |
| Composite NT (z score) | −0.595 (0.85) B,C | −0.143 (0.80) A,C | 1.04 (0.66) A,B | 37.91 | 2/122 | <0.001 |
| Variables | HC (n = 39) A | Low TO-NT (n = 56) B | High TO-NT (n = 30) C | F/X2 df = 2/119 | p |
|---|---|---|---|---|---|
| Total HAMD | 5.5 (0.67) B,C | 15.7 (0.6) A,C | 19.1 (0.8) A,B | 105.21 | <0.001 |
| Total BDI | 8.6 (1.0) B,C | 23.5 (0.9) A,C | 27.0 (1.2) A,B | 85.35 | <0.001 |
| Total HAMA | 7.8 (1.1) B,C | 15.4 (0.9) A,C | 19.5 (1.3) A,B | 26.23 | <0.001 |
| Total FF | 10.9 (1.8) B,C | 24.7 (1.5) A,C | 35.4 (2.0) A,B | 43.05 | <0.001 |
| Pure FF (z score) | −0.877 (0.122) B,C | 0.146 (0.101) A,C | 0.867 (0.138) A,B | 46.06 | <0.001 |
| Pure HAMD (z score) | −0.964 (0.118) B,C | 0.296 (0.098) A,C | 0.702 (0.133) A,B | 51.04 | <0.001 |
| Pure BDI (z score) | −1.030 (0.113) B,C | 0.348 (0.094) A,C | 0.700 (0.128) A,B | 62.79 | <0.001 |
| Pure HAMA (z score) | −0.606 (0.139) B,C | 0.190 (0.115) A | 0.432 (0.157) A | 14.37 | <0.001 |
| Physiosom HAMA (z score) | −0.495 (0.145) B,C | 0.029 (0.120) A,C | 0.588 (0.164) A,B | 12.21 | <0.001 |
| Physiosom HAMD (z score) | −1.031 (0.118) B,C | 0.347 (0.098) A,C | 0.702 (0.133) A,B | 57.93 | <0.001 |
| Biomarkers | HC (n = 39) A | Low TO-NT (n = 56) B | High TO-NT (n = 30) C | F/X2 df = 2/117 | p |
|---|---|---|---|---|---|
| Caspase 1 (pg/mL) | 73.90 (3.27) C | 71.83(2.75) C | 85.57(3.75) A,B | 4.54 | 0.013 |
| IL-1β (pg/mL) | 4.58 (0.33) C | 5.21(0.282) | 5.81(0.385) A | 2.95 | 0.056 |
| IL-18 (pg/mL) | 233.9 (11.91) | 231.62(10.02) | 261.32(13.67) | 1.67 | 0.192 |
| IL-10 (pg/mL) | 9.09 (1.08) B,C | 14.10(0.911) A | 13.06(1.24) A | 6.50 | 0.002 |
| CRP (mg/L) | 5.02 (0.53) B,C | 6.32 (0.443) A,C | 10.11 (0.604) A,B | 27.08 | <0.001 |
| MPO (ng/mL) | 43.1 (3.3) | 49.9 (2.8) | 51.3 (3.8) | 1.72 | 0.184 |
| TAC (U/mL) | 6.74 (0.51) | 6.78 (0.43) | 7.02 (0.58) | 0.08 | 0.925 |
| AOPP (µmol/g) | 0.92 (0.14) B,C | 1.29 (0.12) A,C | 1.76 (0.16) B,C | 7.80 | 0.001 |
| Total calcium (mM) | 2.56 (0.03) B,C | 2.26 (0.02) A | 2.33 (0.03) A | 40.45 | <0.001 |
| Dependent Variables | Explanatory Variables | B | t | p | Model R2 | F | df | p |
|---|---|---|---|---|---|---|---|---|
| #1 Physio-somatic phenome | Model Total calcium NT BMI | −0.414 0.420 0.164 | −6.00 6.16 2.42 | <0.001 <0.001 0.017 | 0.460 | 34.06 | 3/120 | <0.001 |
| #2 Physio-somatic phenome | Model Peak BT Calcium NT | 0.472 −0.253 0.229 | 6.29 −3.76 3.38 | <0.001 <0.001 <0.001 | 0.574 | 53.88 | 3/120 | <0.001 |
| #3 Pure FF | Model Total calcium CRP Education AOPP AstraZeneca vaccination BMI | −0.346 0.242 0.192 0.214 0.166 0.149 | −4.56 3.09 2.59 2.74 2.23 1.99 | <0.001 0.002 0.011 0.007 0.027 0.048 | 0.371 | 11.61 | 6/118 | <0.001 |
| #4 Pure HAMD | Model Total calcium CRP Education AstraZeneca vaccination AOPP MPO | −0.381 0.224 0.244 0.186 0.201 0.155 | −5.25 2.97 3.37 2.58 2.67 2.11 | <0.001 0.004 0.001 0.011 0.009 0.037 | 0.413 | 13.86 | 6/118 | <0.001 |
| #5 Pure BDI | Model Total calcium AOPP Education CRP Interleukin-1β | −0.368 0.247 0.197 0.218 0.174 | −4.87 3.17 2.61 2.80 2.22 | <0.001 0.002 0.010 0.006 0.028 | 0.372 | 14.12 | 5/119 | <0.001 |
| #6 Pure HAMA | Model Total calcium CRP MPO | −0.296 0.204 0.170 | −3.52 2.42 2.06 | 0.001 0.017 0.041 | 0.183 | 9.06 | 3/121 | <0.001 |
| #7 Physiosom HAMD | Model Total calcium CRP Interleukin 18 Vaccination Sinopharm | −0.432 0.301 0.181 −0.159 | −5.84 4.07 2.49 −2.19 | <0.001 <0.001 0.014 0.030 | 0.374 | 17.88 | 4/120 | <0.001 |
| #8 Physiosom HAMA | Model Total calcium MPO BMI | −0.301 0.231 0.207 | −3.70 2.85 2.53 | <0.001 0.005 0.013 | 0.217 | 11.17 | 3/121 | <0.001 |
| Sets | Variables | Canonical Loadings |
|---|---|---|
| Set 1 Dependent | Pure FF | 0.845 |
| Pure HAMD | 0.815 | |
| Physiosom HAMD | 0.784 | |
| Pure HAMA | 0.585 | |
| Physiosom HAMA | 0.629 | |
| Pure BDI | 0.825 | |
| Set 2 Explanatory | NT composite | 0.670 |
| TO2 index | 0.875 | |
| Total calcium | 0.674 | |
| Statistics | F (df) | 8.675 (18/325) |
| P | <0.001 | |
| Correlation | 0.799 | |
| Set 1/set 2 | 0.355 | |
| Set 2 by itself | 0.556 | |
| Set 1 by itself | 0.569 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Hakeim, H.K.; Al-Rubaye, H.T.; Almulla, A.F.; Al-Hadrawi, D.S.; Maes, M. Chronic Fatigue, Depression and Anxiety Symptoms in Long COVID Are Strongly Predicted by Neuroimmune and Neuro-Oxidative Pathways Which Are Caused by the Inflammation during Acute Infection. J. Clin. Med. 2023, 12, 511. https://doi.org/10.3390/jcm12020511
Al-Hakeim HK, Al-Rubaye HT, Almulla AF, Al-Hadrawi DS, Maes M. Chronic Fatigue, Depression and Anxiety Symptoms in Long COVID Are Strongly Predicted by Neuroimmune and Neuro-Oxidative Pathways Which Are Caused by the Inflammation during Acute Infection. Journal of Clinical Medicine. 2023; 12(2):511. https://doi.org/10.3390/jcm12020511
Chicago/Turabian StyleAl-Hakeim, Hussein Kadhem, Haneen Tahseen Al-Rubaye, Abbas F. Almulla, Dhurgham Shihab Al-Hadrawi, and Michael Maes. 2023. "Chronic Fatigue, Depression and Anxiety Symptoms in Long COVID Are Strongly Predicted by Neuroimmune and Neuro-Oxidative Pathways Which Are Caused by the Inflammation during Acute Infection" Journal of Clinical Medicine 12, no. 2: 511. https://doi.org/10.3390/jcm12020511
APA StyleAl-Hakeim, H. K., Al-Rubaye, H. T., Almulla, A. F., Al-Hadrawi, D. S., & Maes, M. (2023). Chronic Fatigue, Depression and Anxiety Symptoms in Long COVID Are Strongly Predicted by Neuroimmune and Neuro-Oxidative Pathways Which Are Caused by the Inflammation during Acute Infection. Journal of Clinical Medicine, 12(2), 511. https://doi.org/10.3390/jcm12020511